
Mechanical Sensing Element PDLIM5 Promotes Osteogenesis of Human Fibroblasts by Affecting the Activity of Microfilaments



 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Osteogenic Differentiation, and Treatments
2.2. Western Blotting
2.3. RNA Extraction and Quantitative Real-Time PCR
2.4. Immunofluorescence
2.5. Alkaline Phosphatase Staining
2.6. Lentivirus Transduction
2.7. Cell Proliferation Assay (CCK8 Assay)
2.8. Wound Healing
2.9. Transwell Migration Experiments
2.10. Statistical Analysis
3. Results
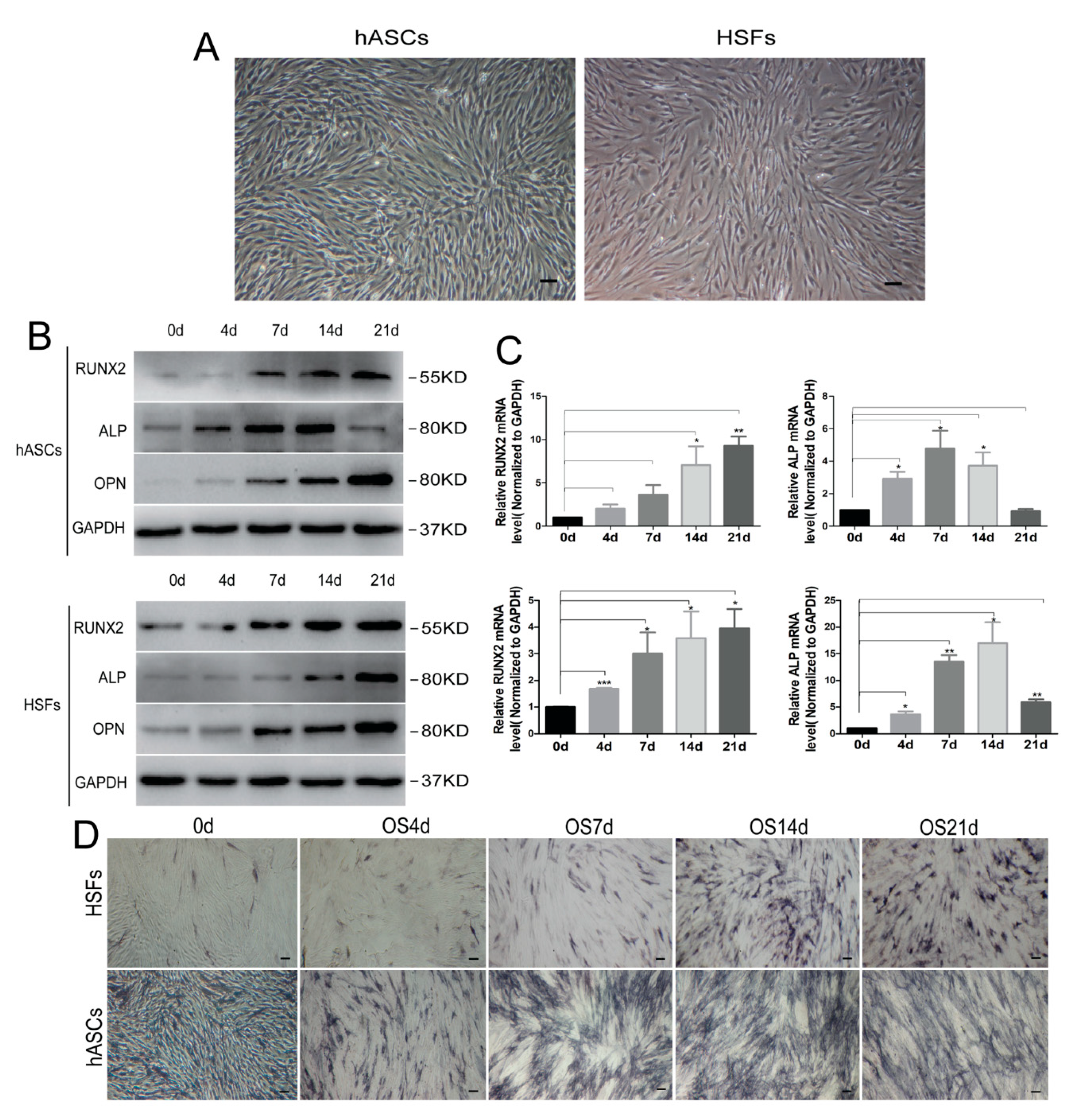
3.1. Cell Culture and Osteogenic Differentiation In Vitro
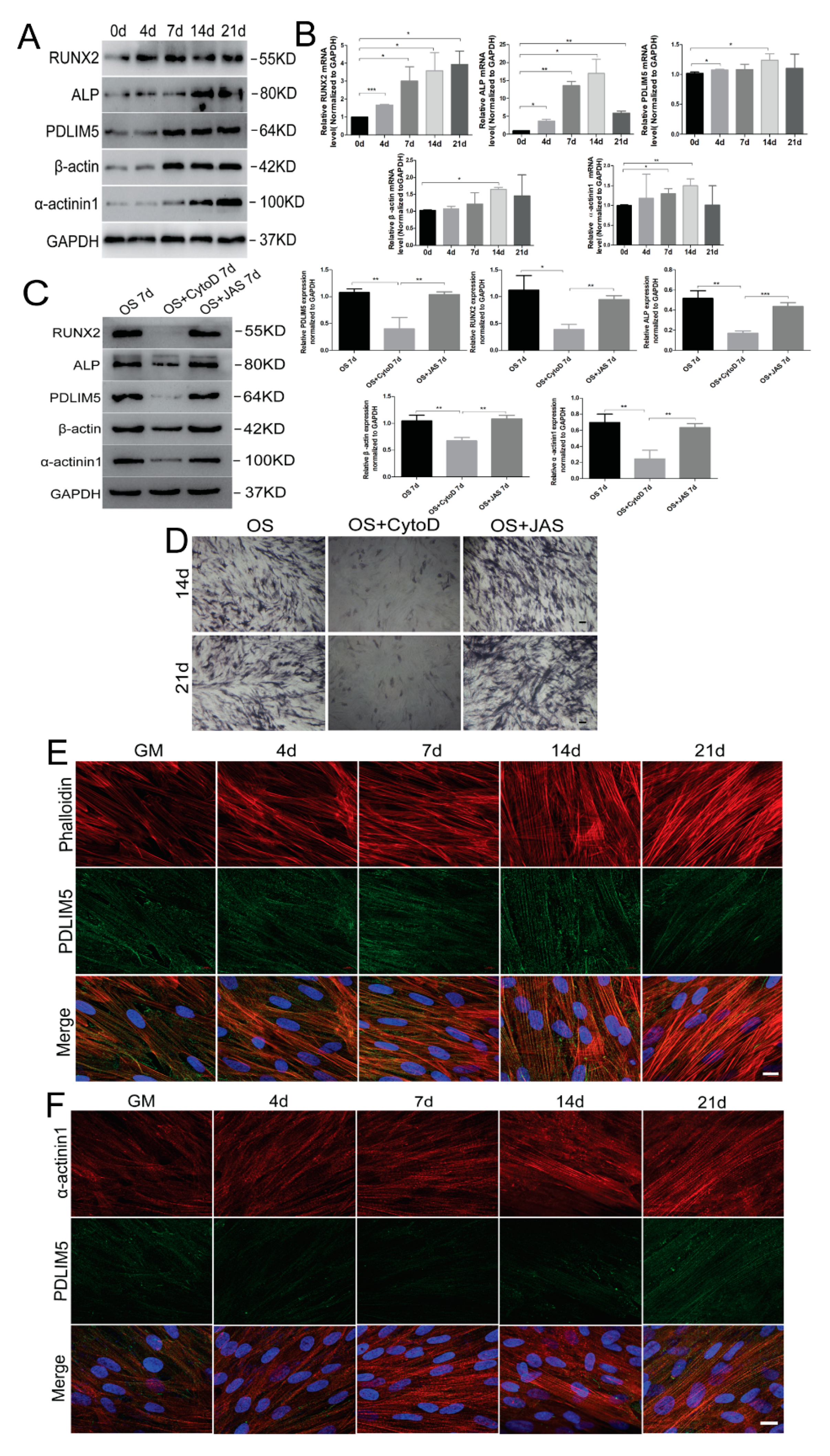
3.2. Microfilaments and Related Proteins Were Involved in Osteogenic Differentiation of HSFs
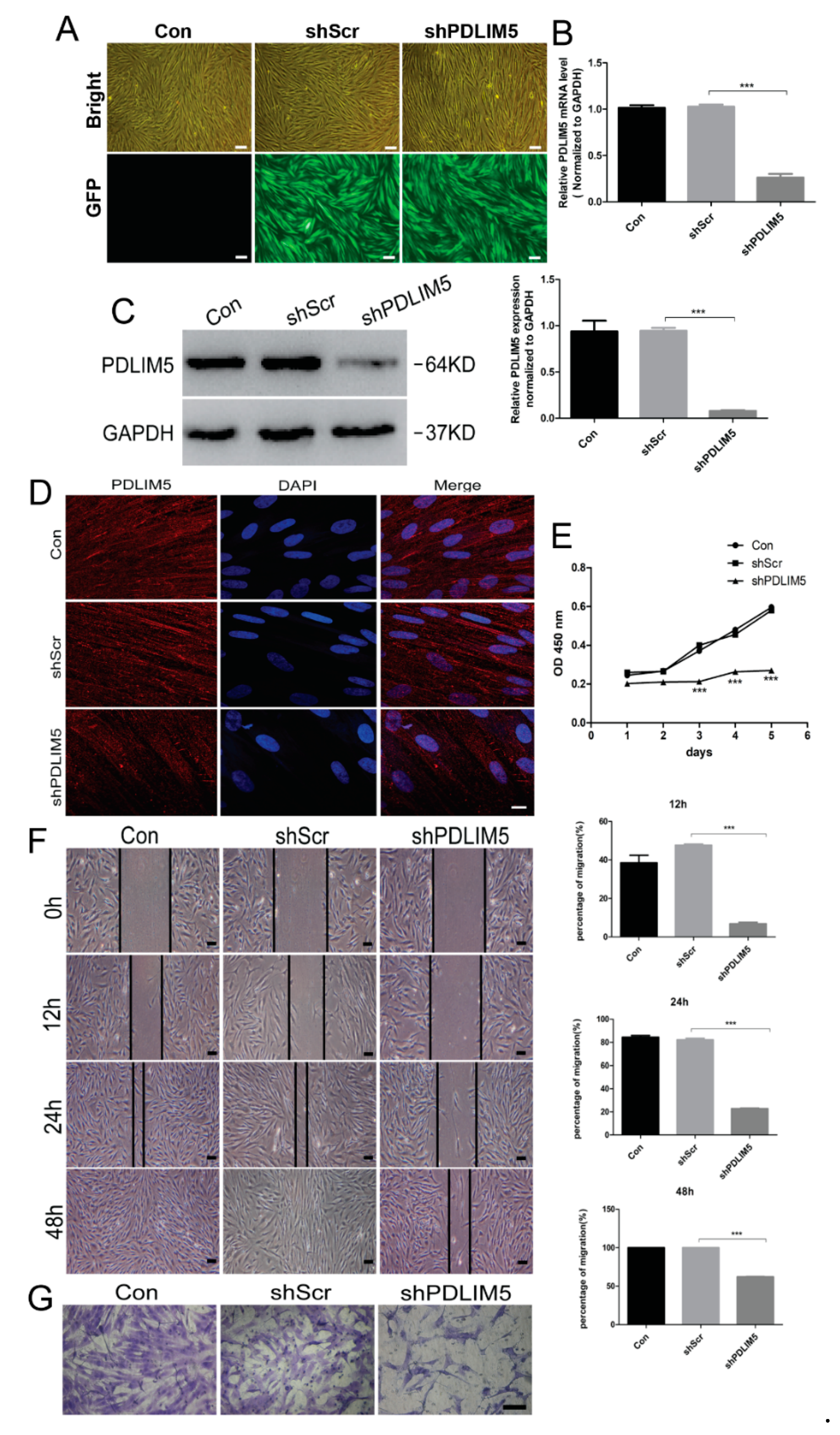
3.3. PDLIM5 Knockdown Inhibited the Proliferation, Movement, and Migration Ability of HSFs
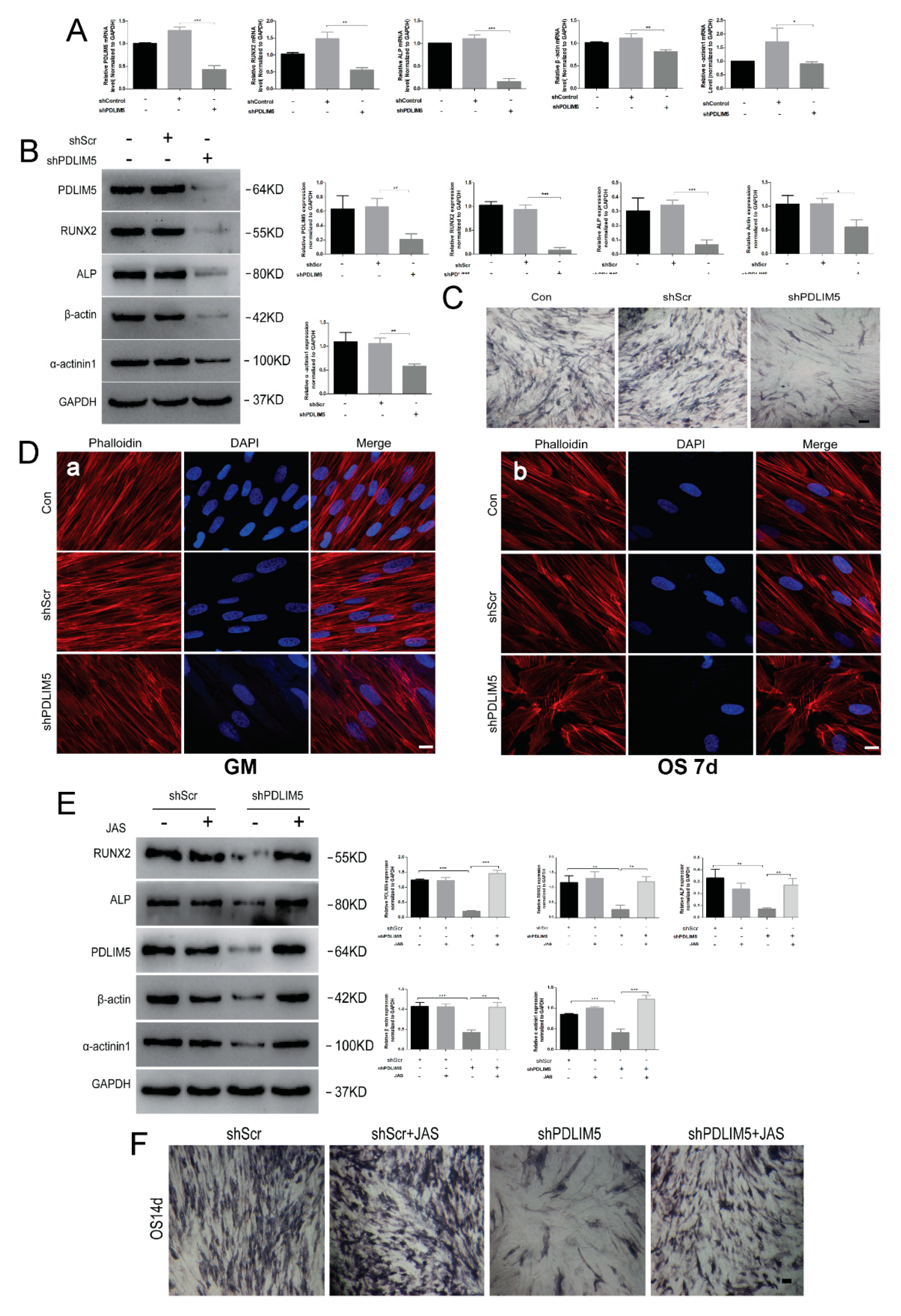
3.4. PDLIM5 Knockdown Attenuated the Osteogenic Differentiation of HSFs Mediated by Microfilament
3.5. Stable Microfilaments Reversed the Inhibitory Effect of Knockdown PDLIM5 on Osteogenic Differentiation of HSFs
3.6. PDLIM5-Knockdown Inhibited the Nuclear Localization of YAP
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Umezawa, A. Regenerative Medicine. Arerugi 2016, 65, 987–989. [Google Scholar]
- Trohatou, O.; Roubelakis, M.G. Mesenchymal Stem/Stromal Cells in Regenerative Medicine: Past, Present, and Future. Cell. Reprogram. 2017, 19, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Walmsley, G.G.; Ransom, R.C.; Zielins, E.R.; Leavitt, T.; Flacco, J.S.; Hu, M.S.; Lee, A.S.; Longaker, M.T.; Wan, D.C. Stem Cells in Bone Regeneration. Stem Cell Rev. Rep. 2016, 12, 524–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Gou, W.; Lu, Q.; Peng, J.; Lu, S. Role of mesenchymal stem cells in bone regeneration and fracture repair: A review. Int. Orthop. 2013, 37, 2491–2498. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, A.G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Yamanaka, S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell 2006, 126, 663–676. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Li, H.; Guo, Z. Mesenchymal stem cell-like properties in fibroblasts. Cell. Physiol. Biochem. 2014, 34, 703–714. [Google Scholar] [CrossRef] [PubMed]
- Christy, B.A.; Herzig, M.C.; Delavan, C.; Cantu, C.; Salgado, C.; Bynum, J.A.; Cap, A.P. Human primary fibroblasts perform similarly to MSCs in assays used to evaluate MSC safety and potency. Transfusion 2019, 59, 1593–1600. [Google Scholar] [CrossRef] [Green Version]
- Hee, C.K.; Nicoll, S. Induction of osteoblast differentiation markers in human dermal fibroblasts: Potential application to bonetissue engineering. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2006, 2006, 521–524. [Google Scholar] [PubMed] [Green Version]
- Tour, G.; Wendel, M.; Tcacencu, I. Human fibroblast-derived extracellular matrix constructs for bone tissue engineering applica tions. J. Biomed. Mater. Res. A 2013, 101, 2826–2837. [Google Scholar] [CrossRef]
- Kuroda, S.; Tokunaga, C.; Kiyohara, Y.; Higuchi, O.; Konishi, H.; Mizuno, K.; Gill, G.N.; Kikkawa, U. Protein-protein interaction of zinc finger LIM domains with protein kinase C. J. Biol. Chem. 1996, 271, 31029–31032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eeckhoute, J. A cell-type-specific transcriptional network required for estrogen regulation of cyclin D1 and cell cycle progression in breast cancer. Genes Dev. 2006, 20, 2513–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edlund, K.; Lindskog, C.; Saito, A.; Berglund, A.; Pontén, F.; Göransson-Kultima, H.; Isaksson, A.; Jirström, K.; Planck, M.; Johansson, L.; et al. CD99 is a novel prognostic stromal marker in non-small cell lung cancer. Int. J. Cancer 2012, 131, 2264–2273. [Google Scholar] [CrossRef] [PubMed]
- Heiliger, K.; Hess, J.; Vitagliano, D.; Salerno, P.; Braselmann, H.; Salvatore, G.; Ugolini, C.; Summerer, I.; Bogdanova, T.; Unger, K.; et al. Novel candidate genes of thyroid tumourigenesis identified in Trk-T1 transgenic mice. Endocr.-Relat. Cancer 2012, 19, 409–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Gao, Y.; Xu, Y.; Sun, X.; Song, X.; Ma, H.; Yang, M. Retracted: Si-RNA-Mediated Knockdown of PDLIM5 Suppresses Gastric Cancer Cell Proliferation in Vitro. Chem. Biol. Drug Des. 2015, 85, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chen, L.; Huang, H.; Lv, J.M.; Chen, M.; Qu, F.J.; Pan, X.W.; Li, L.; Yin, L.; Cui, X.G.; et al. High expression of PDLIM5 facilitates cell tumorigenesis and migration by maintaining AMPK activation in prostate cancer. Oncotarget 2017, 8, 98117–98134. [Google Scholar] [CrossRef]
- Krcmery, J.; Camarata, T.; Kulisz, A.; Simon, H.G. Nucleocytoplasmic functions of the PDZ-LIM protein family: New insights into organ development. Bioessays 2010, 32, 100–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loughran, G.; Healy, N.C.; Kiely, P.A.; Huigsloot, M.; Kedersha, N.L.; O’Connor, R. Mystique is a new insulin-like growth factor-I-regulated PDZ-LIM domain protein that promotes cell attachment and migration and suppresses Anchorage-independent growth. Mol. Biol. Cell 2005, 16, 1811–1822. [Google Scholar] [CrossRef] [Green Version]
- Jani, K.; Schock, F. Zasp is required for the assembly of functional integrin adhesion sites. J. Cell Biol. 2007, 179, 1583–1597. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, N.; Hoshijima, M.; Oyasu, M.; Saito, N.; Tanizawa, K.; Kuroda, S. ENH, Containing PDZ and LIM Domains, Heart/Skeletal Muscle-Specific Protein, Associates with Cytoskeletal Proteins through the PDZ Domain. Biochem. Biophys. Res. Commun. 2000, 272, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Qiu, H.; Liu, N.; Luo, L.; Zhong, J.; Tang, Z.; Kang, K.; Qu, J.; Peng, W.; Liu, L.; Li, L.; et al. MicroRNA-17-92 regulates myoblast proliferation and differentiation by targeting the ENH1/Id1 signaling axis. Cell Death Differ. 2016, 23, 1658–1669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ito, J.; Takita, M.; Takimoto, K.; Maturana, A. Enigma homolog 1 promotes myogenic gene expression and differentiation of C2C12 cells. Biochem. Biophys. Res. Commun. 2013, 435, 483–487. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Zhang, Y.; Yu, S.; Li, S.; Jiang, W.; Zhu, Y.; Xu, Y.; Yang, C.; Tian, G.; Mi, J.; et al. PDLIM5 identified by label-free quantitative proteomics as a potential novel biomarker of papillary thyroid carcinoma. Biochem. Biophys. Res. Commun. 2018, 499, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Ajeian, J.N.; Horton, E.R.; Astudillo, P.; Byron, A.; Askari, J.A.; Millon-Frémillon, A.; Knight, D.; Kimber, S.J.; Humphries, M.J.; Humphries, J.D. Proteomic analysis of integrin-associated complexes from mesenchymal stem cells. PROTEOMICS—Clin. Appl. 2016, 10, 51–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elbediwy, A.; Vanyai, H.; Diaz-De-La-Loza, M.; Frith, D.; Snijders, A.; Thompson, B.J. Enigma proteins regulate YAP mechanotransduction. J. Cell Sci. 2018, 131, s221788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Tsukamoto, O.; Nakano, A.; Kato, H.; Kioka, H.; Ito, N.; Higo, S.; Yamazaki, S.; Shintani, Y.; Matsuoka, K.; et al. Augmented AMPK activity inhibits cell migration by phosphorylating the novel substrate Pdlim5. Nat. Commun. 2015, 6, 6137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sluysmans, S.; Vasileva, E.; Spadaro, D.; Shah, J.; Rouaud, F.; Citi, S. The role of apical cell-cell junctions and associated cytoskeleton in mechanotransduction. Biol. Cell 2017, 109, 139–161. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.D.; Gerlach, B.D. The roles and regulation of the actin cytoskeleton, intermediate filaments and microtubules in smooth muscle cell migration. Respir. Res. 2017, 18, 54. [Google Scholar] [CrossRef] [Green Version]
- Mohan, R.; John, A. Microtubule-associated proteins as direct crosslinkers of actin filaments and microtubules. IUBMB Life 2015, 67, 395–403. [Google Scholar] [CrossRef]
- Svitkina, T.M. Ultrastructure of the actin cytoskeleton. Curr. Opin. Cell Biol. 2018, 54, 1–8. [Google Scholar] [CrossRef]
- Gourlay, C.W.; Ayscough, K.R. The actin cytoskeleton: A key regulator of apoptosis and ageing? Nat. Rev. Mol. Cell Biol. 2005, 6, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhuang, Y.; Ouyang, N.; Yu, H. Cytochalasin D Promotes Osteogenic Differentiation of MC3T3-E1 Cells via p38-MAPK Sig naling Pathway. Curr. Mol. Med. 2019, 20, 79–88. [Google Scholar] [CrossRef]
- Khan, A.U.; Qu, R.; Fan, T.; Ouyang, J.; Dai, J. A glance on the role of actin in osteogenic and adipogenic differentiation of mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 283. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Wu, F.; Chen, J.; Xu, H.; Wang, H.; Guo, F.; Tan, Y.; Ding, B.; Wang, J.; Wan, Y.; et al. Actin microfilament mediates osteoblast Cbfa1 responsiveness to BMP2 under simulated micrograv ity. PLoS ONE 2013, 8, e63661. [Google Scholar]
- Xu, H.; Wu, F.; Zhang, H.; Yang, C.; Li, K.; Wang, H.; Yang, H.; Liu, Y.; Ding, B.; Tan, Y.; et al. Actin cytoskeleton mediates BMP2-Smad signaling via calponin 1 in preosteoblast under simulated microgravity. Biochimie 2017, 138, 184–193. [Google Scholar] [CrossRef]
- Nakatani, M.; Ito, J.; Koyama, R.; Iijima, M.; Yoshimoto, N.; Niimi, T.; Kuroda, S.; Maturana, A.D. Scaffold protein enigma homolog 1 overcomes the repression of myogenesis activation by inhibitor of DNA binding 2. Biochem. Biophys. Res. Commun. 2016, 474, 413–420. [Google Scholar] [CrossRef]
- Lasorella, A.; Iavarone, A. The Protein ENH Is a Cytoplasmic Sequestration Factor for Id2 in Normal and Tumor Cells from the Nervous System. Proc. Nat. Acad. Sci. USA 2006, 103, 4976–4981. [Google Scholar] [CrossRef] [Green Version]
- Yuda, A.; Lee, W.S.; Petrovic, P.; McCulloch, C.A. Novel proteins that regulate cell extension formation in fibroblasts. Exp. Cell Res. 2018, 365, 85–96. [Google Scholar] [CrossRef]
- Mcintosh, K.; Zvonic, S.; Garrett, S.; Mitchell, J.B.; Floyd, Z.E.; Hammill, L.; Kloster, A.; Di Halvorsen, Y.; Ting, J.P.; Storms, R.W.; et al. The immunogenicity of human adipose-derived cells: Temporal changes in vitro. Stem Cells 2006, 24, 1246–1253. [Google Scholar] [CrossRef]
- Denu, R.A.; Nemcek, S.; Bloom, D.D.; Goodrich, A.D.; Kim, J.; Mosher, D.F.; Hematti, P. Fibroblasts and Mesenchymal Stromal/Stem Cells Are Phenotypically Indistinguishable. Acta Haematol. 2016, 136, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Soundararajan, M.; Kannan, S. Fibroblasts and mesenchymal stem cells: Two sides of the same coin? J. Cell Physiol. 2018, 233, 9099–9109. [Google Scholar] [CrossRef]
- Drabek, K.; van de Peppel, J.; Eijken, M.; van Leeuwen, J.P. GPM6B regulates osteoblast function and induction of mineralization by controlling cytoskeleton and matrix vesicle release. J. Bone Miner. Res. 2011, 26, 2045–2051. [Google Scholar] [CrossRef] [PubMed]
- Sen, B.; Xie, Z.; Uzer, G.; Thompson, W.R.; Styner, M.; Wu, X.; Rubin, J. Intranuclear Actin Regulates Osteogenesis. Stem Cells 2015, 33, 3065–3076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.K.; Yu, X.; Cohen, D.M.; Wozniak, M.A.; Yang, M.T.; Gao, L.; Eyckmans, J.; Chen, C.S. Bone morphogenetic protein-2-induced signaling and osteogenesis is regulated by cell shape, RhoA/ROCK, and cytoskeletal tension. Stem Cells Dev. 2012, 21, 1176–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, T.; Qu, R.; Jiang, X.; Yang, Y.; Sun, B.; Huang, X.; Zhou, Z.; Ouyang, J.; Zhong, S.; Dai, J. Spatial organization and crosstalk of vimentin and actin stress fibers regulate the osteogenic differentiation of human adipose-derived stem cells. FASEB J. 2021, 35. [Google Scholar] [CrossRef] [PubMed]
- Martins, C.S.; Ferraz, E.P.; De Castro-Raucci, L.M.; Teixeira, L.N.; Maximiano, W.M.; Rosa, A.L.; De Oliveira, P.T. Changes in actin and tubulin expression in osteogenic cells cultured on bioactive glass-based surfaces. Microsc. Res. Tech. 2015, 78, 1046–1053. [Google Scholar] [CrossRef] [PubMed]
- Guryanova, O.A.; Drazba, J.A.; Frolova, E.I.; Chumakov, P.M. Actin cytoskeleton remodeling by the alternatively spliced isoform of PDLIM4/RIL protein. J. Biol. Chem. 2011, 286, 26849–26859. [Google Scholar] [CrossRef] [Green Version]
- Vallenius, T.; Luukko, K.; Makela, T.P. CLP-36 PDZ-LIM protein associates with nonmuscle alpha-actinin-1 and alpha-actinin-4. J. Biol. Chem. 2000, 275, 11100–11105. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Wang, B.K.; Chang, M.L.; Wan, Z.Q.; Han, G.L. Cyclic Stretch Enhances Osteogenic Differentiation of Human Periodontal Ligament Cells via YAP Activation. Biomed. Res. Int. 2018, 2018, 2174824. [Google Scholar] [CrossRef]
- Du, Y.; Montoya, C.; Orrego, S.; Wei, X.; Ling, J.; Lelkes, P.I.; Yang, M. Topographic cues of a novel bilayered scaffold modulate dental pulp stem cells differentiation by regulating YAP signalling through cytoskeleton adjustments. Cell Prolif. 2019, 52, e12676. [Google Scholar] [CrossRef] [PubMed]
- Tong, Z.; Liu, Y.; Xia, R.; Chang, Y.; Hu, Y.; Liu, P.; Zhai, Z.; Zhang, J.; Li, H. F-actin Regulates Osteoblastic Differentiation of Mesenchymal Stem Cells on TiO2 Nanotubes Through MKL1 and YAP/TAZ. Nanoscale Res. Lett. 2020, 15, 183. [Google Scholar] [CrossRef] [PubMed]
- Pocaterra, A.; Romani, P.; Dupont, S. YAP/TAZ functions and their regulation at a glance. J. Cell Sci. 2020, 133, s230425. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Rowe, R.G.; Botvinick, E.L.; Kurup, A.; Putnam, A.J.; Seiki, M.; Weaver, V.M.; Keller, E.T.; Goldstein, S.; Dai, J.; et al. MT1-MMP-dependent control of skeletal stem cell commitment via a beta1-integ rin/YAP/TAZ signaling axis. Dev. Cell 2013, 25, 402–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (3′→5′) |
|---|---|---|
| RUNX2 | GCCTTCAAGGTGGTAGCCC | CGTTACCCGCCATGACAGTA |
| ALP | CTATCCTGGCTCCGTGCTC | CGCCAGTACTTGGGGTCTTT |
| PDLIM5 | TTAGTGGCACTGGGGAAATC | GATCTTCCTTTGGCATCGAC |
| β-actin | CTTCGCGGGCGACGAT | CCACATAGGAATCCTTCTGACC |
| α-actinin1 | ATTGGCAACGACCCCCAGAA | ATGTTGTAACCCATGGAGATCAGG |
| GAPDH | TCGGAGTCAACGGATTTGGT | TTCCCGTTCTCAGCCTTGAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Qu, R.; Peng, Y.; Yang, Y.; Fan, T.; Sun, B.; Khan, A.U.; Wu, S.; Wei, K.; Xu, C.; et al. Mechanical Sensing Element PDLIM5 Promotes Osteogenesis of Human Fibroblasts by Affecting the Activity of Microfilaments. Biomolecules 2021, 11, 759. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050759
Huang X, Qu R, Peng Y, Yang Y, Fan T, Sun B, Khan AU, Wu S, Wei K, Xu C, et al. Mechanical Sensing Element PDLIM5 Promotes Osteogenesis of Human Fibroblasts by Affecting the Activity of Microfilaments. Biomolecules. 2021; 11(5):759. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050759
Chicago/Turabian StyleHuang, Xiaolan, Rongmei Qu, Yan Peng, Yuchao Yang, Tingyu Fan, Bing Sun, Asmat Ullah Khan, Shutong Wu, Kuanhai Wei, Chujiang Xu, and et al. 2021. "Mechanical Sensing Element PDLIM5 Promotes Osteogenesis of Human Fibroblasts by Affecting the Activity of Microfilaments" Biomolecules 11, no. 5: 759. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11050759