
Osteopontin in Cardiovascular Diseases

1
Department of Cardiovascular Medicine, Graduate School of Medicine, Juntendo University, Bunkyo-ku, Tokyo 1138421, Japan
2
Department of Cardiology, Keio University School of Medicine, Shinjuku-ku, Tokyo 1608582, Japan
*
Author to whom correspondence should be addressed.
Biomolecules 2021, 11(7), 1047; https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071047
Submission received: 21 May 2021
/
Revised: 14 July 2021
/
Accepted: 14 July 2021
/
Published: 16 July 2021
(This article belongs to the Special Issue Molecular Biomarkers In Cardiology 2021)
{kind=link}
{kind=link}
Abstract
:Unprecedented advances in secondary prevention have greatly improved the prognosis of cardiovascular diseases (CVDs); however, CVDs remain a leading cause of death globally. These findings suggest the need to reconsider cardiovascular risk and optimal medical therapy. Numerous studies have shown that inflammation, pro-thrombotic factors, and gene mutations are focused not only on cardiovascular residual risk but also as the next therapeutic target for CVDs. Furthermore, recent clinical trials, such as the Canakinumab Anti-inflammatory Thrombosis Outcomes Study trial, showed the possibility of anti-inflammatory therapy for patients with CVDs. Osteopontin (OPN) is a matricellular protein that mediates diverse biological functions and is involved in a number of pathological states in CVDs. OPN has a two-faced phenotype that is dependent on the pathological state. Acute increases in OPN have protective roles, including wound healing, neovascularization, and amelioration of vascular calcification. By contrast, chronic increases in OPN predict poor prognosis of a major adverse cardiovascular event independent of conventional cardiovascular risk factors. Thus, OPN can be a therapeutic target for CVDs but is not clinically available. In this review, we discuss the role of OPN in the development of CVDs and its potential as a therapeutic target.
1. Introduction
Recent advances in preventive medicine have greatly improved the prognosis of cardiovascular diseases (CVDs) [1,2]. Current standard therapies for secondary prevention include the use of antiplatelet agents, lifestyle modifications, optimal medical therapy, and coronary revascularization. Despite unprecedented advances in secondary prevention, CVDs remain the leading cause of death worldwide. This suggests that the risks of CVDs are underestimated and current optimal medical therapy for secondary prevention is required for patients with CVDs.
Recently, beyond classical cardiovascular risk factors, other drivers, including inflammation, pro-thrombotic factors, and gene mutations, have been focused on not only the residual risk but also the next therapeutic target for CVDs [1,3,4]. Several biological processes contribute to the pathogenesis of CVD. In particular, the targeting of inflammatory processes in experimental animal models has been demonstrated to be beneficial in promoting healing and attenuating myocardial injury [5,6,7]. Furthermore, recent clinical trials have revealed that inflammatory activity contributes to an increased risk of CVDs in humans [8,9]. For instance, increased levels of high-sensitivity C-reactive protein (hsCRP) have been recognized as an independent predictor of both recurrent ischemia and death among patients with coronary artery disease (CAD) [10,11,12] and the reduction in hsCRP by rosuvastatin significantly reduced the incidence of major cardiovascular events (MACE) in healthy controls without hyperlipidemia [13]. In the Canakinumab Anti-inflammatory Thrombosis Outcomes Study (CANTOS) trial, canakinumab, which is an IL-1β inhibitor, resulted in a lower incidence of recurrent non-fatal myocardial infarction (MI), non-fatal stroke, or cardiovascular death among patients with stable CAD and hsCRP levels of >2 mg/L [9]. Lowering of hsCRP levels results in a reduction in MACE, cardiovascular death, and all-cause mortality without any effect on LDL-C [9]. Furthermore, the Colchicine Cardiovascular Outcomes Trial demonstrated a reduction in the primary composite outcome of cardiovascular death, cardiac arrest, non-fatal MI, stroke, or angina resulting in revascularization in patients with CAD [8]. Low-Dose Colchicine 2 (LoDoCo2), which is a proteomic substudy, revealed that colchicine treatment attenuated the NLRP3 inflammasome pathway, including interleukin (IL)-18, IL1 receptor antagonist, and IL-6 in patients with chronic CAD [14]. These data suggest that inflammation contributes to the pathogenesis of CVDs and that anti-inflammatory therapy is clinically applicable to humans.
Osteopontin (OPN) is a matricellular protein that mediates diverse biological functions [15,16,17]. OPN functions as a proinflammatory cytokine and promotes cell-mediated immune responses [18,19,20,21]. In addition to its inflammatory functions, OPN also has protective functions such as biomineralization [20,22,23] and wound healing [16,24,25,26]. OPN is involved in a number of diseases, including MI, atherosclerosis, kidney injury, diabetes, and other chronic inflammatory diseases in experimental animal models [15,19,24,25,27,28,29] and is also a strong predictor of adverse outcomes in patients with CVDs [30,31,32]. Thus, OPN is not only a risk factor but also a potential therapeutic target for CVDs. This review will explore the evidence linking OPN to CVDs and potential future therapeutic options to attenuate inflammation as a residual cardiovascular risk factor.
2. Diversity of the Origin and Regulation of OPN in CVDs
OPN has a two-faced phenotype dependent on the pathological state of a number of CVDs. OPN is hardly expressed under physiological conditions [24,33], but its expression markedly increases under a number of pathological conditions [24,33,34,35,36,37]. The major source of OPN varies depending on the organ and pathological conditions. Acute increases in OPN have protective roles, including wound healing and neovascularization [15,24]. In contrast, chronic increases in OPN predict poor prognosis of a major adverse cardiovascular event independent of conventional risk factors [16,30,31,36,38,39].
2.1. The Source of OPN
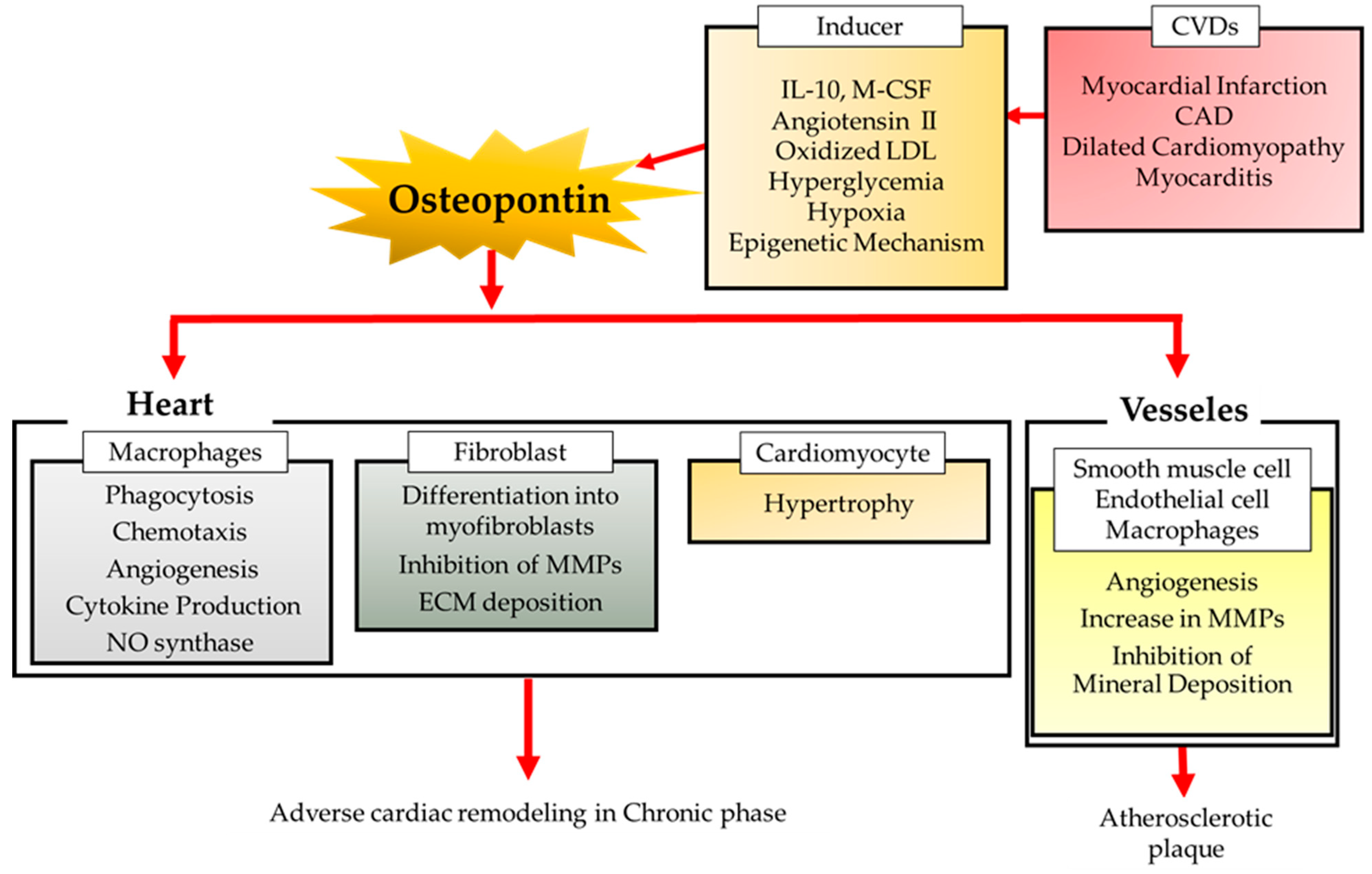
In the human heart after MI, macrophages infiltrating the infarcted myocardium make up the major source of OPN [24,40]. We previously showed that OPN is hardly expressed in normal hearts, but its expression increases markedly after MI. The hearts in Spp1 (encoding OPN) knockout (KO) mice have fewer single collagen filaments compared to the hearts in WT-sham mice, although there is no significant difference in cardiac function [33]. OPN is almost exclusively produced by galectin-3 hi CD206 + macrophages, which appear specifically in the infarcted myocardium after MI, determined by flow cytometric analysis of EGFP-Spp1-KI reporter mice heart cells subjected to MI [24]. In experimental models of chronic viral myocarditis or myocardial necrosis by transdiaphragmatic freeze-thaw, infiltrating macrophages have also been reported as the major source of OPN [34,40]. OPN has been implicated in multiple functions of macrophages, including phagocytosis, chemotaxis, angiogenesis, cytokine production, and the expression of inducible nitric oxide synthase [41]. OPN enhances phagocytosis through integrin [42] and induces macrophage migration via interaction of the C-terminal fragment with CD44 surface receptors [43] and via the SLAYGLR domain with integrin receptors [44]. OPN secreted by alternatively activated macrophages directly contributes to the phagocytic clearance of dead cells and to the reparative fibrotic response in wound healing [27,45,46,47,48,49]. OPN also induces the differentiation of cardiac fibroblasts into α-smooth muscle actin+ myofibroblasts [50]. Spp1 KO mice showed exaggerated post-MI left ventricular (LV) chamber dilatation associated with reduced collagen accumulation and no increase in collagen 1(α1) mRNA in infarcted myocardium after MI [24,33].
Cardiomyocytes have also been reported as a major source of OPN in diseased hearts. OPN expression is elevated in hypertrophied myocardium [51], where it modulates the activation of p38 kinase and JNKs and the development of myocardial hypertrophy in response to chronic pressure overload in mice [52]. The increase in OPN expression during streptozotocin-induced diabetic cardiomyopathy has been reported to induce cardiomyocyte apoptosis, hypertrophy, and fibrosis by modulating protein kinase C activity. Genetic depletion of OPN improves cardiac hypertrophy, function, and fibrosis in these mice [53].
Increased OPN expression protects the post-MI heart and plays a beneficial role in regulating LV remodeling after MI by promoting collagen synthesis and accumulation [24,33]. The extracellular matrix (ECM) plays an important role in the pathological remodeling of the myocardium [54,55,56]. ECM remodeling is complicated since it contributes to the development of LV dysfunction and the progression of heart failure [54,55,56]. Cardiac fibroblasts play a fundamental role in tissue remodeling after MI by modulating ECM deposition [57,58,59]. OPN in cardiac fibroblasts plays an essential role in tissue repair by enhancing their proliferative and adhesive functions. Cardiac fibroblasts derived from OPN KO mice exhibit a reduction in adhesion to ECM proteins such as collagen I compared with those derived from wild-type (WT) mice [60,61]. OPN induces cardiac fibroblasts to differentiate into α-smooth muscle actin+ myofibroblasts [50]. In addition, the arginine-glycine-aspartate (RGD) sequence of OPN binds to beta3 integrins on the fibroblast to promote fibroblast binding to collagen [61]. Furthermore, OPN expression is increased not only in macrophages but also in smooth muscle and endothelial cells in human coronary atherosclerotic plaques [62].
Thus, OPN is involved in a number of CVDs and the major source and role of OPN depends on pathological states. In order to use OPN as a therapeutic target, it is necessary to understand the induction and mechanism of action of OPN in each pathological state.
2.2. Inducer of OPN
A number of stimuli have been shown to promote OPN expression, including cytokines, reactive oxygen species (ROS), angiotensin II (Ang II), high glucose, and hypoxia [24,28,63,64,65].
We previously reported that the IL-10-STAT3-galectin-3 axis is essential for Spp1 transcriptional activation in cardiac macrophages after MI [24]. IL-10 and M-CSF act synergistically to activate STAT3 and ERK in cardiac macrophages, which in turn upregulate the expression of galectin-3 and MerTK, resulting in the functional maturation of OPN-producing macrophages [65].
Ang II upregulated Spp1 expression in adult rat cardiac fibroblasts by ROS-mediated activation of ERK1/2 and JNK pathways and IL-1beta and TNF-alpha further act synergistically with Ang II to increase Spp1 expression [64]. Ang II activates the mitogen-activated protein kinase (p42/44 MAPK) pathway followed by the induction of Spp1 expression in cardiac microvascular endothelial cells [64]. Interestingly, blocking Ang-II inhibits Spp1 expression in non-infarcted myocardium of post-MI hearts [63] and olmesartan, which is an Ang II receptor blocker, significantly decreases plasma OPN levels in patients with hypertension [66].
Furthermore, hyperglycemia and hypoxia synergistically increase OPN expression in smooth muscle cells or proximal tubule epithelial cells in the kidney [28,67]. Oxidized low-density lipoprotein (LDL) promotes proliferation and migration of smooth muscle cells derived from the human coronary artery via upregulation of OPN and MMP-9 [68]. Glucocorticoid dexamethasone has also been shown to increase Spp1 expression in cardiomyocytes and microvascular endothelial cells in vitro and OPN suppressed NOS2 activity by regulating the location and extent of NOS2 induction in these cells [69].
Thus, mechanisms that regulate the expression of Spp1 varies among cell types and pathologies, which results in the complexity of OPN as a therapeutic target for CVD.
2.3. OPN Isoforms
Multiple OPN isoforms exist in human as the result of alternative splicing and these isoforms have distinct biologic functions [70]. Full-length OPN can be modified by thrombin cleavage. Thrombin-cleaved OPN exposes an epitope for integrin receptors of α4β1, α9β1, and α9β4. OPN isoform includes OPNa (the full-length isoform), OPNb (which lacks exon 5 and 3) and OPNc (which lacks exon 4) [15]. In CVDs, the question of which OPN isoforms are involved or what roles each isoform have with respect to their involvement in pathological states remain elusive and thus further studies are required.
3. OPN in CVDs
Accumulating evidence suggests that OPN is a potent biomarker and mediator in CVDs. In this section, we discuss the roles of OPN in CVDs, specifically the ischemic heart diseases, hypertension, heart failure, dilated cardiomyopathy, atherosclerosis, and several cardiomyopathies (Table S1).
3.1. Ischemic Heart Diseases
Plasma OPN levels drastically changed in a time-dependent manner in patients who underwent successful reperfusion after anterior-wall acute MI and they began to increase on day two, peaking on day three, and persisting until day 14 (420 ng/mL for control subjects versus 1139 ng/mL for day three) [71]. These data are consistent with temporal Spp1 expression in an experimental animal model [24]. We analyzed EGFP-Spp1-KI reporter mice hearts subjected to MI and GFP (the activity of Spp1 transcription in situ) was expressed almost exclusively in cardiac macrophages of the infarcted myocardium. Temporal Spp1 expression in cardiac macrophages peaked on day three and OPN secreted by cardiac macrophages directly contributed to the phagocytic clearance of dead cells and the reparative fibrotic response in wound healing after MI [24].
Plasma OPN levels also increased in patients with stable CAD [72,73] and LV ejection fraction (EF) was inversely correlated with plasma OPN levels in these patients [71,72,74]. Furthermore, in patients with stable CAD with preserved EF on optimal medical therapy, plasma OPN levels were independently associated with adverse cardiovascular outcomes [30]. In patients with a previous anterior wall MI, OPN is assumed to be released from the heart into the coronary circulation [72]. However, in a mouse model of acute MI caused by coronary artery ligation, we confirmed that there is little transcriptional activation of Spp1 in the heart 28 days after MI (Shirakawa et al., unpublished data). In patients with post-MI, OPN may continue to be released continuously from the residual coronary plaque.
Spp1 KO mice showed exaggerated post-MI LV chamber dilatation associated with reduced collagen accumulation in the infarcted myocardium after MI [24,27,33]. Cardiac fibroblasts play a fundamental role in the tissue remodeling process after MI by modulating the extent and composition of the ECM. Transforming growth factor-β1 (TGF-β1) induces the differentiation of fibroblasts to myofibroblasts in WT mice, but this does not occur in Spp1 KO mice. Cardiac fibroblasts derived from Spp1 KO mice showed a reduction in stress fibers, focal adhesions, spreading ability, and resistance to detachment by shear stress compared to those derived from WT mice [50]. These results suggest that OPN is required for myofibroblast differentiation induced by TGF-β1.
Matrix metalloproteinases (MMPs) are members of the ECM proteases and play a fundamental role in tissue remodeling in a number of CVDs, including pressure overload-induced hypertrophy, MI, or dilated cardiomyopathy (DCM) [50,75,76]. MMPs are responsible for collagen degradation in the ECM and the activation of MMP induces a reduction in cardiac tissue tensile strength and causes systolic and diastolic dysfunction [77]. OPN causes the upregulation of tissue inhibitors of MMP and collagen and the downregulation of MMP-1 expression in cardiac fibroblasts [78]. OPN inhibits IL-1β-induced activation of MMP-2 and MMP-9 via the involvement of β3 integrins and activation of protein kinase C-ζ in adult rat cardiac fibroblasts and this resulted in enhanced collagen deposition after MI [79]. MMP inhibition decreases post-MI LV dilation in Spp1 KO mice, but not in WT mice [45]. Furthermore, cardiac transgenic MMP-2 expression induces impaired contractility [80] and the deletion of MMP-2 attenuates cardiac rupture and LV dysfunction after MI [81].
OPN is also reported to be involved in angiogenesis, which is a critical and beneficial process in post-ischemic repair, after MI. Angiogenesis enhances the oxygen and nutrient supply necessary for the wound healing process and decreases the severity of ischemic events [82,83]. OPN plays an important role in post-ischemic neovascularization [84,85] and is upregulated in patients with peripheral artery disease or diabetes mellitus [86,87]. Genetic depletion of Spp1 reduces angiogenesis in cardiovascular endothelial cells after MI. OPN KO mice show impaired myocardial angiogenic response and results in adverse remodeling post-MI [88].
Thus, OPN has a complex effect on cardiac remodeling after MI (Figure 1). Inhibition of OPN may not only ameliorate excessive fibrosis, but also promote MMP activity and inhibit angiogenesis. Furthermore, the major source of OPN in the chronic phase of MI remains unclear. There are still many issues that need to be resolved to achieve secondary prevention of ischemic heart disease by targeting OPN.
3.2. Hypertension
An increase in OPN expression in aortic tissues has been observed in a number of animal models of hypertension and its expression is correlated with systolic blood pressure [89]. OPN expression is increased via the Akt1/AP-1 dependent pathway and promotes the production of MMP-2 in vascular smooth muscle cells followed by vascular remodeling [89]. The effects of OPN on MMP are opposite in cardiac fibroblasts and vascular smooth muscle cells and the action of OPN may be target cell-dependent. Furthermore, OPN plays an essential role in modulating compensatory cardiac hypertrophy in response to chronic pressure overload and MMP regulation by OPN is important for this process [52].
3.3. Heart Failure
Plasma OPN levels were elevated in patients with chronic heart failure (HF) due to LV dysfunction and correlated with the severity as assessed by the New York Heart Association class (NYHA). Furthermore, plasma OPN levels predicted death within four years of follow-up in these patients (382 ng/mL for control subjects versus 532 ng/mL for patients with HF and 479 ng/mL for the New York Heart Association (NYHA) class I/II versus 672 ng/mL for NYHA class III/IV) [90]. OPN expression is increased in the myocardium of patients with hypertensive heart disease and HF compared to the controls and the increase in OPN expression is correlated with lysyl oxidase, insoluble collagen, pulmonary capillary wedge pressure, and LV chamber stiffness while being inversely correlated with LVEF [91]. Furthermore, serum levels of OPN are predictors of ventricular tachycardia and fibrillation in patients with HF [92].
Col4a3−/− mice show a phenotype similar to that of heart failure with preserved EF, including diastolic dysfunction, cardiac hypertrophy and fibrosis, pulmonary edema, and impaired mitochondrial function. Genetic depletion of OPN ameliorates these phenotypes by regulating mitochondrial oxoglutarate dehydrogenase L [32].
However, the major source of OPN remains unclear. HF is a complex multiple organ disease and thus OPN production may increase not only in the heart but also in other organs, including the kidney, lung, and liver. Elucidation of the major source and inducer of OPN results in novel therapies for HF.
3.4. Dilated Cardiomyopathy
Plasma OPN levels are elevated and associated with the severity of HF, as assessed by NYHA class in patients with DCM [32]. Analysis of the myocardium obtained from patients with DCM revealed that the main source of OPN was cardiomyocytes and the extent of Spp1 expression showed a positive correlation with LV end-systolic diameter and a negative correlation with LVEF in these patients [93]. Furthermore, Spp1 expression levels in the myocardium are elevated, especially in patients with a LVEF of <50% and correlated with the accumulation of collagen type I [94], serving as a biomarker for prognosis in patients with DCM [95].
Cardiac-specific integrin-linked kinase (ILK) is a serine/threonine kinase that has been linked to HF. ILK KO mice spontaneously develop phenotypes such as DCM, including apoptosis, fibrosis, and inflammation. Blocking OPN attenuates the decline in cardiac function in these mice [96]. Furthermore, the inhibition of OPN suppressed collagen I secretion in fibroblasts through a FAK/Akt-dependent pathway and reduced LV remodeling and dysfunction in mice with DCM [97]. Thus, although effective treatments for DCM are scarce, OPN may be a future therapeutic target for DCM.
3.5. Atherosclerosis
OPN is primarily expressed in endothelial cells, macrophages, and smooth muscle-derived foam cells, with an increased expression in human atherosclerotic plaques in the aorta, carotid, and coronary arteries [62,98]. The increase in OPN expression in plaques is reported to be correlated with the formation of ulceration, inflammation, and unstable plaques in patients who underwent carotid endarterectomy [99]. Serum OPN levels were higher in patients with acute coronary syndrome than in those with stable CAD and were associated with rapid coronary plaque progression and in-stent restenosis (684 ng/mL for no progression versus 977 ng/mL for progression) [100]. On the contrary, it has been reported that OPN expression levels of coronary plaque in patients who underwent endarterectomy during coronary bypass surgery are lower in unstable plaques and plaques with large calcification deposits [101].
In mice, the overexpression of OPN induced the development of medial thickening and neointimal formation and OPN KO mice showed attenuation of atherosclerosis [102]. The overexpression of OPN develops early fatty-streak and mononuclear cell-rich lesions in a high-fat diet (HFD)-induced atherosclerosis model of OPN transgenic mice [103]. Thus, OPN plays a fundamental role in the development and progression of atherosclerotic plaques.
3.6. Cardiovascular Calcification
OPN is recognized as an inhibitor of mineral deposition in the vascular wall and cardiac valves [15]. OPN was significantly and positively correlated to the coronary calcium score evaluated by cardiac computed tomography [104]. Smooth muscle cells isolated from the aortas of Spp1 KO mice showed higher calcification compared to those of WT mice in the presence of inorganic phosphate, which is an inducer of vascular calcification [105]. Furthermore, OPN phosphorylation is required for the inhibition of human vascular smooth muscle cell calcification [106,107]. Epigenetic modifications of the genome, such as DNA methylation and histone modifications, play an important role in the development of CVDs [108,109]. For instance, H3K9a, H3K27ac, or H3K4 have been reported to increase in atherosclerotic smooth muscle cells or macrophages [110]. There are several reports on epigenetic regulation of OPN. OPN is markedly upregulated in the alveolar macrophages of the bleomycin model of lung fibrosis. The HIF1α-driven glycolysis in alveolar macrophages altered the epigenetic modification of OPN, with the metabolic intermediate 3-phosphoglyceric acid, and reduced its expression [111]. Epigenetic regulations, including Methyl-CpG binding protein 2 and the activation of nuclear calcium signaling, have been demonstrated to be involved in OPNc production [112]. Epigenetic regulation of Spp1 in CVDs remain unclear and thus further studies are required. Hydroxyapatite deposition in atherosclerotic coronary plaques is associated with OPN expression [113]. OPN has been reported to inhibit calcification by blocking hydroxyapatite crystal formation and regulating acidification, while carbonic anhydrase II in macrophages and implanted glutaraldehyde-fixed aortic valve leaflets showed accelerated calcification in OPN KO mice compared to WT mice [114,115]. Exogenous recombinant histidine-fused OPN, which is phosphorylated and contains a functional RGD cell adhesive domain, attenuated ectopic calcification of glutaraldehyde-fixed bovine pericardium in rats [115]. Furthermore, OPN promotes the differentiation of monocytes derived from patients with vascular calcification into alternatively activated macrophages and inhibits their differentiation into osteoclasts [116].
In addition, foamy macrophages persist in atherosclerotic plaques and promote disease progression [117]. In the context of atherosclerosis, OPN is generally regarded as a proinflammatory and proatherogenic molecule. A significantly higher expression of OPN was observed in foam cells of atherosclerotic plaques [118,119]. However, recent single-cell RNA sequencing analysis revealed that non-foamy macrophages formed a number of inflammatory transcripts, including IL-1β, and foamy macrophages expressed fewer inflammatory genes than non-foamy macrophages in the atherosclerotic aorta [120,121]. These data suggest that OPN-producing macrophages may have a protective effect on vascular calcification in atherosclerotic plaques. OPN promotes atherosclerosis but inhibits the calcification. This is expected to be dependent on the type of OPN and the localization of OPN-producing cells in atherosclerotic plaques.
3.7. Other Heart Diseases
OPN is involved in the pathogenesis of CVDs. Cardiac-specific overexpression of OPN showed cardiomyocyte loss, severe fibrosis, and Th1 cell infiltration followed by LV diastolic and systolic function and the development of myocarditis in mice [122]. Furthermore, OPN expression is increased in macrophages, resulting in cardiac fibrosis and enhanced cardiac remodeling in a model of coxsackievirus B3-induced myocarditis [123]. MMP-9 is increased in the hearts of dystrophin-deficient mdx mice, which possess point mutation in the Duchenne muscular dystrophy-related gene, and genetic ablation of MMP9 diminishes the activation of ERK1/2 and Akt kinase followed by an improvement in cardiac function [124]. OPN has been shown to enhance MMP-9 expression in cardiac and skeletal muscle, resulting in the development of myopathy in these mice. Furthermore, OPN induces myocardial MMP-2 activation followed by the development of cardiac remodeling, inflammation, and interstitial fibrosis in a mouse model of chronic Chagas heart disease [125]. In patients with amyloid light-chain amyloidosis, serum OPN levels are an independent predictor of all-cause mortality [126]. OPN/p38 MAPK signaling pathways are suggested to protect against ascending aortic aneurysm progression [127]. Thus, OPN plays an essential role in the pathogenesis of several heart diseases.
4. OPN in Multi-Organ Crosstalk
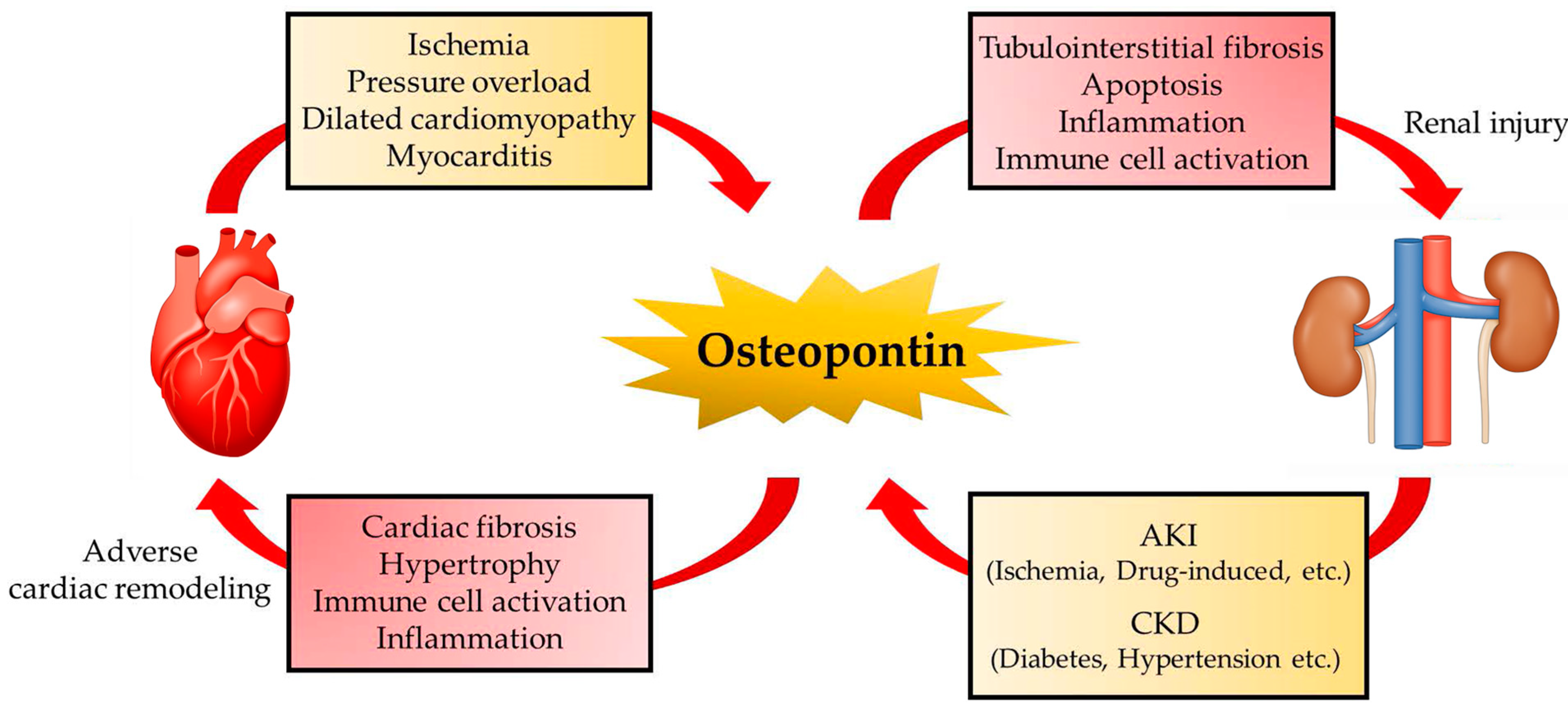
Multi-organ crosstalk is defined as a complex biological interaction between distant organs. CVDs are not only single organ diseases, but rather a complex multi-organ syndrome with impairments including the kidney, endocrine, lung, skeletal muscle, liver, and hematopoietic system. Many molecules, including proinflammatory cytokines and growth factors, are reported to be involved in multi-organ dysfunction in CVDs. In particular, the interplay between the heart, kidney, and diabetes plays an essential role in the pathogenesis of CVD. In this section, we review the role of OPN in the kidney, diabetes, and cardiorenal syndrome.
4.1. OPN in the Kidney
In both humans and mice, OPN is reported to be mainly expressed in the thick ascending limb of the loops of Henle and distal nephrons in the steady state [19]. The physiological role of OPN in normal kidneys remains unclear and the transcriptional activation of OPN rarely occurs in normal kidneys [19]. Furthermore, Spp1 KO mice showed no abnormalities in the kidney. In contrast, in a number of pathological conditions, OPN is highly upregulated and plays a critical role in interstitial fibrosis, especially in the proximal tubules [128]. Multiple factors, including cytokines, hormones, hyperglycemia, and hypoxia, induce OPN upregulation in the kidneys [128]. Tubular injury can result in abnormalities in many metabolic and signaling pathways, which contribute to tubulointerstitial fibrosis and kidney dysfunction. The different cell types and pathological states differ in their regulatory mechanisms of Spp1 expression in the kidney.
4.1.1. Acute Kidney Injury
The plasma OPN levels were elevated and independently predicted the 28 day survival rate in patients with acute kidney injury (AKI) [129]. Urine levels of OPN at admission and on day three were associated with an increased risk of AKI and mortality in survivors after out-of-hospital cardiac arrest [130]. OPN is also recognized as a predictor of early kidney transplant rejection [131]. OPN can activate NK cells to mediate the apoptosis of the tubular epithelial cells [132]. Blockade of OPN inhibited the infiltration of NK cells into the kidney, resulting in a reduction in the expression of injury markers, including neutrophil gelatinase-associated lipocalin and kidney injury molecule-1, and apoptosis of renal tissue in mice after renal ischemia reperfusion injury [133]. Genetic depletion of Spp1 results in a reduction in apoptosis in both the tubular epithelium and interstitium in the acute phase of ischemia-reperfusion injury in mice [134]. Furthermore, following ischemia-reperfusion injury, OPN KO mice showed less macrophage infiltration and interstitial fibrosis compared to WT mice [134]. These results suggest that inflammation and oxidative stress induced by ischemia-reperfusion injury may result in the upregulation of OPN, which causes renal damage in AKI.
4.1.2. Chronic Kidney Injury
OPN is involved in AKI as well as chronic kidney disease (CKD). The plasma OPN levels, but not urine levels, were significantly higher in diabetic patients than in healthy controls [95,135]. The increase in plasma levels of OPN is associated with the development of microalbuminuria and negatively correlated with eGFR in patients with diabetes [136,137]. In experimental diabetic animal models, OPN expression in the proximal tubules was also reported to be markedly upregulated in db/db and streptozocin-induced diabetic mice [138,139]. Hyperglycemic conditions induce histone acetylation and methylation, which in turn results in upregulation of OPN gene expression [138]. Furthermore, Ang II accelerates macrophage infiltration and fibrosis in the kidney and subsequently results in the elevation of blood pressure and the increase in urinary albumin/creatinine ratios in mice. Genetic depletion of OPN protects Ang II-induced inflammation, oxidative stress, and fibrosis of the kidney [140,141]. Thus, OPN has been recognized as a facilitator of tubulointerstitial injury in the pathogenesis of CKD.
4.2. Diabetes
As previously mentioned, OPN plays an essential role in the pathogenesis of diabetic neuropathy in mice [138,139]. OPN has been reported to be increased in the circulating blood of obese diabetic and insulin-resistant patients [136,142] and to enhance visceral adipose inflammation, which results in insulin resistance [143,144]. We previously reported that a CD153+PD-1+CD44hiCD4+T cell population, which accumulates in the visceral adipose tissue of HFD-induced obese mice, causes adipose inflammation by producing large amounts of OPN [29]. Interestingly, CD153+PD-1+CD44hiCD4+T cells acquire a senescent phenotype and are not easily eliminated as a negative legacy of obesity after weight reduction [145]. Furthermore, it has been reported that OPN expression increases in the epicardial fat from patients with CAD compared to controls and is associated with the presence of calcified atherosclerotic plaques [146]. Flow cytometric analysis of epicardial fat in HFD-induced obese EGFP-Spp1-KI reporter mice demonstrated that CD153+PD-1+CD44hiCD4+T cells markedly increased and produced a large amount of OPN (Shirakawa K., unpublished observation). These findings suggest that senescent-adipose CD4 T cells in epicardial fat may play a pivotal role in the development of CVDs in patients with diabetes.
4.3. Cardiorenal Syndrome
Cardiorenal syndrome (CRS) is involved in a spectrum of disorders involving both the heart and kidneys and is currently the focus of much attention as a therapeutic target for HF. Acute or chronic dysfunction of the heart or kidneys can induce acute or chronic dysfunction in other organs and several factors are involved in the interactions between the heart and kidney [147]. OPN is recognized as one of the key molecules that mediates cardiorenal syndrome (Figure 2). In fact, plasma OPN and creatinine levels are elevated in patients with stable CAD. Furthermore, higher levels of OPN are associated with multivessel lesions and decline in the renal function of these patients [148]. Serum levels of OPN predict adverse cardiovascular events in patients with CKD [149]. However, the major source and role of OPN in CRS still remain elusive.
The cardioprotective effect of SGLT2 inhibition, which is responsible for the reabsorption of sodium and glucose in the proximal tubules of the kidney, has reaffirmed the significance of CRS in the pathogenesis of heart failure and CKD. Empagliflozin reduced the rates of death from cardiovascular causes, hospitalization for heart failure, and death from any cause in patients with type 2 diabetes who are at high risk for cardiovascular events and who received empagliflozin [150]. Canagliflozin also reduced the risk of cardiovascular death and hospitalization for heart failure in patients with type 2 diabetes with a high risk of CVD [151]. Furthermore, dapagliflozin resulted in a lower rate of cardiovascular death or hospitalization for HF in patients with type 2 diabetes and who had no atherosclerotic CVDs [152]. More recently, among patients with HF and a reduced EF, the risk of worsening HF or death was lower in the SGLT2 inhibitor group than in the placebo group, regardless of the presence or absence of diabetes [153]. These results suggest a beneficial effect of SGLT2 inhibition beyond glycemic control. However, the reason for the prognostic improvement of CVD with kidney-targeted treatment remains unclear.
Under healthy conditions, aerobic energy metabolism in the proximal tubule occurs with fatty acids as the energy substrate [154]. A recent systematic approach with transcriptomics, metabolomics, and metabolic flux analysis demonstrated that metabolic reprograming has occurred in proximal tubular epithelial cells in diabetes [155,156,157]; specifically, oxygen consumption is increased, glycolytic pathways are set in motion, and the production of ROS in mitochondria is enhanced [158]. Increased oxygen consumption in tubular epithelial cells results in a decrease in the partial pressure of tissue oxygen in the renal cortex (hypoxia) [159]. These changes were responsible for tubulointerstitial fibrosis. The SGLT2 inhibitors decrease oxygen consumption in proximal tubular epithelial cells and improve tissue oxygenation [159]. We found that in cultured proximal tubular epithelial cells, a high-glucose environment drives the glycolytic pathway and induces enhanced mitochondrial ROS and transcriptional activity of OPN [28], which could be suppressed by inhibiting the glycolytic pathway or by inhibiting SGLT2-mediated uptake of glucose into the cell [28]. In diabetes mellitus, myoinositol oxygenase (MIOX) is overexpressed in proximal tubular epithelial cells and activates the myoinositol pathway, which is a collateral pathway of glycolytic pathways [160]. This results in mitochondrial fragmentation and depolarization and simultaneously impairs mitophagy [160]. The resulting accumulation of impaired mitochondria induces excessive ROS production and cell death [161]. We found that SGLT2 inhibitors suppress the increased expression of MIOX in a high-glucose environment [28]. These results suggest that SGLT2 inhibitors may prevent the development of heart failure via CRS as a result of ameliorating abnormal glucose metabolism in the proximal tubules of the kidney and suppresses OPN production [162].
5. Conclusions
Accumulating evidence demonstrates the significance of OPN in the pathogenesis of CVD. The effects of OPN are disease-specific and consists of positive and negative effects. Therefore, OPN is a useful biomarker for predicting the risk of CVD; however, OPN itself cannot be directly targeted for treatment. Instead, the mechanism of OPN production specific to each condition should be the target of therapy.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/biom11071047/s1, Table S1: Osteopontin in cardiovascular diseases.
Author Contributions
Conceptualization, writing (original draft preparation, review, and editing), and funding acquisition; supervision and project administration; K.S. and M.S. Both authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the JSPS Grant-in-Aid for Scientific Research B Grant Numbers 18H02812.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
We are grateful to Nagahiro Minato and Yoko Hamazaki (Kyoto University, Japan) for their technical assistance. We would like to thank Editage for English language editing.
Conflicts of Interest
All authors have read the journal authorship agreement and policy on disclosure of potential conflicts of interest and have nothing to disclose.
References
- Dhindsa, D.S.; Sandesara, P.B.; Shapiro, M.D.; Wong, N.D. The Evolving Understanding and Approach to Residual Cardiovascular Risk Management. Front. Cardiovasc. Med. 2020, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W.; et al. 2019 ACC/AHA Guideline on the Primary Prevention of Cardiovascular Disease: Executive Summary: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e563–e595. [Google Scholar] [CrossRef]
- Libby, P. The changing landscape of atherosclerosis. Nature 2021, 592, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Stock, J.K. Residual inflammatory risk: Lessons from trials for the future. Atherosclerosis 2020, 311, 103–104. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Hervás, S.; González-Navarro, H. Anti-inflammatory Therapies for Cardiovascular Disease: Signaling Pathways and Mechanisms. Rev. Esp. Cardiol. 2019, 72, 767–773. [Google Scholar] [CrossRef]
- Lutgens, E.; Atzler, D.; Döring, Y.; Duchene, J.; Steffens, S.; Weber, C. Immunotherapy for cardiovascular disease. Eur. Heart J. 2019, 40, 3937–3946. [Google Scholar] [CrossRef]
- Golia, E.; Limongelli, G.; Natale, F.; Fimiani, F.; Maddaloni, V.; Pariggiano, I.; Bianchi, R.; Crisci, M.; D’Acierno, L.; Giordano, R.; et al. Inflammation and cardiovascular disease: From pathogenesis to therapeutic target. Curr. Atheroscler. Rep. 2014, 16, 435. [Google Scholar] [CrossRef]
- Nidorf, S.M.; Fiolet, A.T.L.; Mosterd, A.; Eikelboom, J.W.; Schut, A.; Opstal, T.S.J.; The, S.H.K.; Xu, X.F.; Ireland, M.A.; Lenderink, T.; et al. Colchicine in Patients with Chronic Coronary Disease. N. Engl. J. Med. 2020, 383, 1838–1847. [Google Scholar] [CrossRef]
- Ridker, P.M.; Everett, B.M.; Thuren, T.; MacFadyen, J.G.; Chang, W.H.; Ballantyne, C.; Fonseca, F.; Nicolau, J.; Koenig, W.; Anker, S.D.; et al. Antiinflammatory Therapy with Canakinumab for Atherosclerotic Disease. N. Engl. J. Med. 2017, 377, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Yousuf, O.; Mohanty, B.D.; Martin, S.S.; Joshi, P.H.; Blaha, M.J.; Nasir, K.; Blumenthal, R.S.; Budoff, M.J. High-sensitivity C-reactive protein and cardiovascular disease: A resolute belief or an elusive link? J. Am. Coll. Cardiol. 2013, 62, 397–408. [Google Scholar] [CrossRef] [Green Version]
- Sattar, N.; Murray, H.M.; McConnachie, A.; Blauw, G.J.; Bollen, E.L.; Buckley, B.M.; Cobbe, S.M.; Ford, I.; Gaw, A.; Hyland, M.; et al. C-reactive protein and prediction of coronary heart disease and global vascular events in the Prospective Study of Pravastatin in the Elderly at Risk (PROSPER). Circulation 2007, 115, 981–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ridker, P.M. Clinical application of C-reactive protein for cardiovascular disease detection and prevention. Circulation 2003, 107, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Ridker, P.M.; Danielson, E.; Fonseca, F.A.; Genest, J.; Gotto, A.M., Jr.; Kastelein, J.J.; Koenig, W.; Libby, P.; Lorenzatti, A.J.; MacFadyen, J.G.; et al. Rosuvastatin to prevent vascular events in men and women with elevated C-reactive protein. N. Engl. J. Med. 2008, 359, 2195–2207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Opstal, T.S.J.; Hoogeveen, R.M.; Fiolet, A.T.L.; Silvis, M.J.M.; The, S.H.K.; Bax, W.A.; de Kleijn, D.P.V.; Mosterd, A.; Stroes, E.S.G.; Cornel, J.H. Colchicine Attenuates Inflammation Beyond the Inflammasome in Chronic Coronary Artery Disease: A LoDoCo2 Proteomic Substudy. Circulation 2020, 142, 1996–1998. [Google Scholar] [CrossRef] [PubMed]
- Lok, Z.S.Y.; Lyle, A.N. Osteopontin in Vascular Disease. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 613–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelaziz Mohamed, I.; Gadeau, A.P.; Hasan, A.; Abdulrahman, N.; Mraiche, F. Osteopontin: A Promising Therapeutic Target in Cardiac Fibrosis. Cells 2019, 8, 1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waller, A.H.; Sanchez-Ross, M.; Kaluski, E.; Klapholz, M. Osteopontin in cardiovascular disease: A potential therapeutic target. Cardiol. Rev. 2010, 18, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Lamort, A.S.; Giopanou, I.; Psallidas, I.; Stathopoulos, G.T. Osteopontin as a Link between Inflammation and Cancer: The Thorax in the Spotlight. Cells 2019, 8, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaleta, B. The role of osteopontin in kidney diseases. Inflamm. Res. Off. J. Eur. Histamine Res. Soc. 2019, 68, 93–102. [Google Scholar] [CrossRef]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Cho, H.J.; Kim, H.S. Osteopontin: A multifunctional protein at the crossroads of inflammation, atherosclerosis, and vascular calcification. Curr. Atheroscler. Rep. 2009, 11, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Si, J.; Wang, C.; Zhang, D.; Wang, B.; Zhou, Y. Osteopontin in Bone Metabolism and Bone Diseases. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2020, 26, e919159. [Google Scholar] [CrossRef] [PubMed]
- Giachelli, C.M.; Steitz, S. Osteopontin: A versatile regulator of inflammation and biomineralization. Matrix Biol. J. Int. Soc. Matrix Biol. 2000, 19, 615–622. [Google Scholar] [CrossRef]
- Shirakawa, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Yoshida, N.; Yamamoto, T.; Isobe, S.; Moriyama, H.; Goto, S.; Kitakata, H.; et al. IL (Interleukin)-10-STAT3-Galectin-3 Axis Is Essential for Osteopontin-Producing Reparative Macrophage Polarization After Myocardial Infarction. Circulation 2018, 138, 2021–2035. [Google Scholar] [CrossRef] [PubMed]
- Weber, C.E.; Li, N.Y.; Wai, P.Y.; Kuo, P.C. Epithelial-mesenchymal transition, TGF-β, and osteopontin in wound healing and tissue remodeling after injury. J. Burn Care Res. Off. Publ. Am. Burn Assoc. 2012, 33, 311–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, M.D.; Pedraza, C.E.; Kaartinen, M.T. Osteopontin and wound healing in bone. Cells Tissues Organs 2011, 194, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Foster, C.R.; Dalal, S.; Singh, K. Osteopontin: Role in extracellular matrix deposition and myocardial remodeling post-MI. J. Mol. Cell. Cardiol. 2010, 48, 538–543. [Google Scholar] [CrossRef] [Green Version]
- Shirakawa, K.; Sano, M. Sodium-Glucose Co-Transporter 2 Inhibitors Correct Metabolic Maladaptation of Proximal Tubular Epithelial Cells in High-Glucose Conditions. Int. J. Mol. Sci. 2020, 21, 7676. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, K.; Yan, X.; Shinmura, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Yamamoto, T.; Anzai, A.; Isobe, S.; Yoshida, N.; et al. Obesity accelerates T cell senescence in murine visceral adipose tissue. J. Clin. Investig. 2016, 126, 4626–4639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalrhim, A.D.; Marroush, T.S.; Austin, E.E.; Gersh, B.J.; Solak, N.; Rizvi, S.A.; Bailey, K.R.; Kullo, I.J. Plasma Osteopontin Levels and Adverse Cardiovascular Outcomes in the PEACE Trial. PLoS ONE 2016, 11, e0156965. [Google Scholar] [CrossRef] [PubMed]
- Klingel, K.; Kandolf, R. Osteopontin: A biomarker to predict the outcome of inflammatory heart disease. Semin. Thromb. Hemost. 2010, 36, 195–202. [Google Scholar] [CrossRef]
- Yousefi, K.; Irion, C.I.; Takeuchi, L.M.; Ding, W.; Lambert, G.; Eisenberg, T.; Sukkar, S.; Granzier, H.L.; Methawasin, M.; Lee, D.I.; et al. Osteopontin Promotes Left Ventricular Diastolic Dysfunction Through a Mitochondrial Pathway. J. Am. Coll. Cardiol. 2019, 73, 2705–2718. [Google Scholar] [CrossRef] [PubMed]
- Trueblood, N.A.; Xie, Z.; Communal, C.; Sam, F.; Ngoy, S.; Liaw, L.; Jenkins, A.W.; Wang, J.; Sawyer, D.B.; Bing, O.H.; et al. Exaggerated left ventricular dilation and reduced collagen deposition after myocardial infarction in mice lacking osteopontin. Circ. Res. 2001, 88, 1080–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szalay, G.; Sauter, M.; Haberland, M.; Zuegel, U.; Steinmeyer, A.; Kandolf, R.; Klingel, K. Osteopontin: A fibrosis-related marker molecule in cardiac remodeling of enterovirus myocarditis in the susceptible host. Circ. Res. 2009, 104, 851–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sam, F.; Xie, Z.; Ooi, H.; Kerstetter, D.L.; Colucci, W.S.; Singh, M.; Singh, K. Mice lacking osteopontin exhibit increased left ventricular dilation and reduced fibrosis after aldosterone infusion. Am. J. Hypertens. 2004, 17, 188–193. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Sirokman, G.; Communal, C.; Robinson, K.G.; Conrad, C.H.; Brooks, W.W.; Bing, O.H.; Colucci, W.S. Myocardial osteopontin expression coincides with the development of heart failure. Hypertension 1999, 33, 663–670. [Google Scholar] [CrossRef] [Green Version]
- Williams, E.B.; Halpert, I.; Wickline, S.; Davison, G.; Parks, W.C.; Rottman, J.N. Osteopontin expression is increased in the heritable cardiomyopathy of Syrian hamsters. Circulation 1995, 92, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Carbone, F.; Rigamonti, F.; Burger, F.; Roth, A.; Bertolotto, M.; Spinella, G.; Pane, B.; Palombo, D.; Pende, A.; Bonaventura, A.; et al. Serum levels of osteopontin predict major adverse cardiovascular events in patients with severe carotid artery stenosis. Int. J. Cardiol. 2018, 255, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Bjerre, M.; Pedersen, S.H.; Møgelvang, R.; Lindberg, S.; Jensen, J.S.; Galatius, S.; Flyvbjerg, A. High osteopontin levels predict long-term outcome after STEMI and primary percutaneous coronary intervention. Eur. J. Prev. Cardiol. 2013, 20, 922–929. [Google Scholar] [CrossRef]
- Murry, C.E.; Giachelli, C.M.; Schwartz, S.M.; Vracko, R. Macrophages express osteopontin during repair of myocardial necrosis. Am. J. Pathol. 1994, 145, 1450–1462. [Google Scholar] [PubMed]
- Rittling, S.R. Osteopontin in macrophage function. Expert Rev. Mol. Med. 2011, 13, e15. [Google Scholar] [CrossRef]
- Schack, L.; Stapulionis, R.; Christensen, B.; Kofod-Olsen, E.; Skov Sørensen, U.B.; Vorup-Jensen, T.; Sørensen, E.S.; Höllsberg, P. Osteopontin enhances phagocytosis through a novel osteopontin receptor, the alphaXbeta2 integrin. J. Immunol. 2009, 182, 6943–6950. [Google Scholar] [CrossRef] [Green Version]
- Weber, G.F.; Zawaideh, S.; Hikita, S.; Kumar, V.A.; Cantor, H.; Ashkar, S. Phosphorylation-dependent interaction of osteopontin with its receptors regulates macrophage migration and activation. J. Leukoc. Biol. 2002, 72, 752–761. [Google Scholar] [PubMed]
- Lund, S.A.; Wilson, C.L.; Raines, E.W.; Tang, J.; Giachelli, C.M.; Scatena, M. Osteopontin mediates macrophage chemotaxis via α4 and α9 integrins and survival via the α4 integrin. J. Cell. Biochem. 2013, 114, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Krishnamurthy, P.; Peterson, J.T.; Subramanian, V.; Singh, M.; Singh, K. Inhibition of matrix metalloproteinases improves left ventricular function in mice lacking osteopontin after myocardial infarction. Mol. Cell. Biochem. 2009, 322, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Hulsmans, M.; Sager, H.B.; Roh, J.D.; Valero-Muñoz, M.; Houstis, N.E.; Iwamoto, Y.; Sun, Y.; Wilson, R.M.; Wojtkiewicz, G.; Tricot, B.; et al. Cardiac macrophages promote diastolic dysfunction. J. Exp. Med. 2018, 215, 423–440. [Google Scholar] [CrossRef]
- Zheng, Y.H.; Tian, C.; Meng, Y.; Qin, Y.W.; Du, Y.H.; Du, J.; Li, H.H. Osteopontin stimulates autophagy via integrin/CD44 and p38 MAPK signaling pathways in vascular smooth muscle cells. J. Cell. Physiol. 2012, 227, 127–135. [Google Scholar] [CrossRef]
- Toyonaga, T.; Nakase, H.; Ueno, S.; Matsuura, M.; Yoshino, T.; Honzawa, Y.; Itou, A.; Namba, K.; Minami, N.; Yamada, S.; et al. Osteopontin Deficiency Accelerates Spontaneous Colitis in Mice with Disrupted Gut Microbiota and Macrophage Phagocytic Activity. PLoS ONE 2015, 10, e0135552. [Google Scholar] [CrossRef]
- Shin, Y.J.; Kim, H.L.; Choi, J.S.; Choi, J.Y.; Cha, J.H.; Lee, M.Y. Osteopontin: Correlation with phagocytosis by brain macrophages in a rat model of stroke. Glia 2011, 59, 413–423. [Google Scholar] [CrossRef]
- Lenga, Y.; Koh, A.; Perera, A.S.; McCulloch, C.A.; Sodek, J.; Zohar, R. Osteopontin expression is required for myofibroblast differentiation. Circ. Res. 2008, 102, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Graf, K.; Do, Y.S.; Ashizawa, N.; Meehan, W.P.; Giachelli, C.M.; Marboe, C.C.; Fleck, E.; Hsueh, W.A. Myocardial osteopontin expression is associated with left ventricular hypertrophy. Circulation 1997, 96, 3063–3071. [Google Scholar] [CrossRef]
- Xie, Z.; Singh, M.; Singh, K. Osteopontin modulates myocardial hypertrophy in response to chronic pressure overload in mice. Hypertension 2004, 44, 826–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, V.; Krishnamurthy, P.; Singh, K.; Singh, M. Lack of osteopontin improves cardiac function in streptozotocin-induced diabetic mice. Am. J. Physiol. Heart Circ. Physiol. 2007, 292, H673–H683. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhao, Q.; Kong, W. Extracellular matrix remodeling and cardiac fibrosis. Matrix Biol. J. Int. Soc. Matrix Biol. 2018, 68–69, 490–506. [Google Scholar] [CrossRef]
- Frangogiannis, N.G. The extracellular matrix in myocardial injury, repair, and remodeling. J. Clin. Investig. 2017, 127, 1600–1612. [Google Scholar] [CrossRef] [Green Version]
- Burke, R.M.; Burgos Villar, K.N.; Small, E.M. Fibroblast contributions to ischemic cardiac remodeling. Cell. Signal. 2021, 77, 109824. [Google Scholar] [CrossRef] [PubMed]
- Humeres, C.; Frangogiannis, N.G. Fibroblasts in the Infarcted, Remodeling, and Failing Heart. JACC Basic Transl. Sci. 2019, 4, 449–467. [Google Scholar] [CrossRef]
- Fan, D.; Takawale, A.; Lee, J.; Kassiri, Z. Cardiac fibroblasts, fibrosis and extracellular matrix remodeling in heart disease. Fibrogenesis Tissue Repair 2012, 5, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zohar, R.; Zhu, B.; Liu, P.; Sodek, J.; McCulloch, C.A. Increased cell death in osteopontin-deficient cardiac fibroblasts occurs by a caspase-3-independent pathway. Am. J. Physiol. Heart Circ. Physiol. 2004, 287, H1730–H1739. [Google Scholar] [CrossRef]
- Ashizawa, N.; Graf, K.; Do, Y.S.; Nunohiro, T.; Giachelli, C.M.; Meehan, W.P.; Tuan, T.L.; Hsueh, W.A. Osteopontin is produced by rat cardiac fibroblasts and mediates A(II)-induced DNA synthesis and collagen gel contraction. J. Clin. Investig. 1996, 98, 2218–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, E.R.; Garvin, M.R.; Stewart, D.K.; Hinohara, T.; Simpson, J.B.; Schwartz, S.M.; Giachelli, C.M. Osteopontin is synthesized by macrophage, smooth muscle, and endothelial cells in primary and restenotic human coronary atherosclerotic plaques. Arterioscler. Thromb. J. Vasc. Biol. 1994, 14, 1648–1656. [Google Scholar] [CrossRef] [Green Version]
- Kusuyama, T.; Yoshiyama, M.; Omura, T.; Nishiya, D.; Enomoto, S.; Matsumoto, R.; Izumi, Y.; Akioka, K.; Takeuchi, K.; Iwao, H.; et al. Angiotensin blockade inhibits osteopontin expression in non-infarcted myocardium after myocardial infarction. J. Pharmacol. Sci. 2005, 98, 283–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Pimental, D.R.; Lohan, S.; Vasertriger, A.; Pligavko, C.; Colucci, W.S.; Singh, K. Regulation of angiotensin II-stimulated osteopontin expression in cardiac microvascular endothelial cells: Role of p42/44 mitogen-activated protein kinase and reactive oxygen species. J. Cell. Physiol. 2001, 188, 132–138. [Google Scholar] [CrossRef]
- Shirakawa, K.; Endo, J.; Kataoka, M.; Katsumata, Y.; Anzai, A.; Moriyama, H.; Kitakata, H.; Hiraide, T.; Ko, S.; Goto, S.; et al. MerTK Expression and ERK Activation Are Essential for the Functional Maturation of Osteopontin-Producing Reparative Macrophages After Myocardial Infarction. J. Am. Heart Assoc. 2020, 9, e017071. [Google Scholar] [CrossRef]
- Lorenzen, J.M.; Neunhöffer, H.; David, S.; Kielstein, J.T.; Haller, H.; Fliser, D. Angiotensin II receptor blocker and statins lower elevated levels of osteopontin in essential hypertension—Results from the EUTOPIA trial. Atherosclerosis 2010, 209, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Sodhi, C.P.; Phadke, S.A.; Batlle, D.; Sahai, A. Hypoxia stimulates osteopontin expression and proliferation of cultured vascular smooth muscle cells: Potentiation by high glucose. Diabetes 2001, 50, 1482–1490. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Ren, Y.; Kang, L.; Zhang, L. Oxidized low-density lipoprotein increases the proliferation and migration of human coronary artery smooth muscle cells through the upregulation of osteopontin. Int. J. Mol. Med. 2014, 33, 1341–1347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, K.; Balligand, J.L.; Fischer, T.A.; Smith, T.W.; Kelly, R.A. Glucocorticoids increase osteopontin expression in cardiac myocytes and microvascular endothelial cells. Role in regulation of inducible nitric oxide synthase. J. Biol. Chem. 1995, 270, 28471–28478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimba, E.R.; Tilli, T.M. Human osteopontin splicing isoforms: Known roles, potential clinical applications and activated signaling pathways. Cancer Lett. 2013, 331, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Suezawa, C.; Kusachi, S.; Murakami, T.; Toeda, K.; Hirohata, S.; Nakamura, K.; Yamamoto, K.; Koten, K.; Miyoshi, T.; Shiratori, Y. Time-dependent changes in plasma osteopontin levels in patients with anterior-wall acute myocardial infarction after successful reperfusion: Correlation with left-ventricular volume and function. J. Lab. Clin. Med. 2005, 145, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Tamura, A.; Shingai, M.; Aso, N.; Hazuku, T.; Nasu, M. Osteopontin is released from the heart into the coronary circulation in patients with a previous anterior wall myocardial infarction. Circ. J. Off. J. Jpn. Circ. Soc. 2003, 67, 742–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maniatis, K.; Siasos, G.; Oikonomou, E.; Vavuranakis, M.; Zaromytidou, M.; Mourouzis, K.; Paraskevopoulos, T.; Charalambous, G.; Papavassiliou, A.G.; Tousoulis, D. Osteoprotegerin and Osteopontin Serum Levels are Associated with Vascular Function and Inflammation in Coronary Artery Disease Patients. Curr. Vasc. Pharmacol. 2020, 18, 523–530. [Google Scholar] [CrossRef]
- Georgiadou, P.; Iliodromitis, E.K.; Kolokathis, F.; Varounis, C.; Gizas, V.; Mavroidis, M.; Capetanaki, Y.; Boudoulas, H.; Kremastinos, D.T. Osteopontin as a novel prognostic marker in stable ischaemic heart disease: A 3-year follow-up study. Eur. J. Clin. Investig. 2010, 40, 288–293. [Google Scholar] [CrossRef]
- Spinale, F.G. Myocardial matrix remodeling and the matrix metalloproteinases: Influence on cardiac form and function. Physiol. Rev. 2007, 87, 1285–1342. [Google Scholar] [CrossRef] [PubMed]
- Lindsey, M.L.; Mann, D.L.; Entman, M.L.; Spinale, F.G. Extracellular matrix remodeling following myocardial injury. Ann. Med. 2003, 35, 316–326. [Google Scholar] [CrossRef] [PubMed]
- Mujumdar, V.S.; Smiley, L.M.; Tyagi, S.C. Activation of matrix metalloproteinase dilates and decreases cardiac tensile strength. Int. J. Cardiol. 2001, 79, 277–286. [Google Scholar] [CrossRef]
- Pardo, A.; Gibson, K.; Cisneros, J.; Richards, T.J.; Yang, Y.; Becerril, C.; Yousem, S.; Herrera, I.; Ruiz, V.; Selman, M.; et al. Up-regulation and profibrotic role of osteopontin in human idiopathic pulmonary fibrosis. PLoS Med. 2005, 2, e251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Singh, M.; Siwik, D.A.; Joyner, W.L.; Singh, K. Osteopontin inhibits interleukin-1beta-stimulated increases in matrix metalloproteinase activity in adult rat cardiac fibroblasts: Role of protein kinase C-zeta. J. Biol. Chem. 2003, 278, 48546–48552. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.Y.; Bergman, M.R.; Nguyen, A.P.; Turcato, S.; Swigart, P.M.; Rodrigo, M.C.; Simpson, P.C.; Karliner, J.S.; Lovett, D.H.; Baker, A.J. Cardiac transgenic matrix metalloproteinase-2 expression directly induces impaired contractility. Cardiovasc. Res. 2006, 69, 688–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashidani, S.; Tsutsui, H.; Ikeuchi, M.; Shiomi, T.; Matsusaka, H.; Kubota, T.; Imanaka-Yoshida, K.; Itoh, T.; Takeshita, A. Targeted deletion of MMP-2 attenuates early LV rupture and late remodeling after experimental myocardial infarction. Am. J. Physiol. Heart Circ. Physiol. 2003, 285, H1229–H1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsman, P.; vant Hof, A.W.; de Boer, M.J.; Hoorntje, J.C.; Suryapranata, H.; Dambrink, J.H.; Zijlstra, F. Role of collateral circulation in the acute phase of ST-segment-elevation myocardial infarction treated with primary coronary intervention. Eur. Heart J. 2004, 25, 854–858. [Google Scholar] [CrossRef] [PubMed]
- Sabia, P.J.; Powers, E.R.; Ragosta, M.; Sarembock, I.J.; Burwell, L.R.; Kaul, S. An association between collateral blood flow and myocardial viability in patients with recent myocardial infarction. N. Engl. J. Med. 1992, 327, 1825–1831. [Google Scholar] [CrossRef]
- Duvall, C.L.; Weiss, D.; Robinson, S.T.; Alameddine, F.M.; Guldberg, R.E.; Taylor, W.R. The role of osteopontin in recovery from hind limb ischemia. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 290–295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lyle, A.N.; Joseph, G.; Fan, A.E.; Weiss, D.; Landázuri, N.; Taylor, W.R. Reactive oxygen species regulate osteopontin expression in a murine model of postischemic neovascularization. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1383–1391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahles, F.; Findeisen, H.M.; Bruemmer, D. Osteopontin: A novel regulator at the cross roads of inflammation, obesity and diabetes. Mol. Metab. 2014, 3, 384–393. [Google Scholar] [CrossRef]
- Koshikawa, M.; Aizawa, K.; Kasai, H.; Izawa, A.; Tomita, T.; Kumazaki, S.; Tsutsui, H.; Koyama, J.; Shimodaira, S.; Takahashi, M.; et al. Elevated osteopontin levels in patients with peripheral arterial disease. Angiology 2009, 60, 42–45. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Johnson, J.N.; Singh, K.; Singh, M. Impairment of myocardial angiogenic response in the absence of osteopontin. Microcirculation 2007, 14, 233–240. [Google Scholar] [CrossRef]
- Seo, K.W.; Lee, S.J.; Ye, B.H.; Kim, Y.W.; Bae, S.S.; Kim, C.D. Mechanical stretch enhances the expression and activity of osteopontin and MMP-2 via the Akt1/AP-1 pathways in VSMC. J. Mol. Cell. Cardiol. 2015, 85, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, M.; Zugck, C.; Nelles, M.; Juenger, C.; Frank, D.; Remppis, A.; Giannitsis, E.; Katus, H.A.; Frey, N. Osteopontin, a new prognostic biomarker in patients with chronic heart failure. Circ. Heart Fail. 2008, 1, 43–49. [Google Scholar] [CrossRef] [Green Version]
- López, B.; González, A.; Lindner, D.; Westermann, D.; Ravassa, S.; Beaumont, J.; Gallego, I.; Zudaire, A.; Brugnolaro, C.; Querejeta, R.; et al. Osteopontin-mediated myocardial fibrosis in heart failure: A role for lysyl oxidase? Cardiovasc. Res. 2013, 99, 111–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Francia, P.; Adduci, C.; Semprini, L.; Borro, M.; Ricotta, A.; Sensini, I.; Santini, D.; Caprinozzi, M.; Balla, C.; Simmaco, M.; et al. Osteopontin and galectin-3 predict the risk of ventricular tachycardia and fibrillation in heart failure patients with implantable defibrillators. J. Cardiovasc. Electrophysiol. 2014, 25, 609–616. [Google Scholar] [CrossRef]
- Satoh, M.; Nakamura, M.; Akatsu, T.; Shimoda, Y.; Segawa, I.; Hiramori, K. Myocardial osteopontin expression is associated with collagen fibrillogenesis in human dilated cardiomyopathy. Eur. J. Heart Fail. 2005, 7, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Cabiati, M.; Svezia, B.; Matteucci, M.; Botta, L.; Pucci, A.; Rinaldi, M.; Caselli, C.; Lionetti, V.; Del Ry, S. Myocardial Expression Analysis of Osteopontin and Its Splice Variants in Patients Affected by End-Stage Idiopathic or Ischemic Dilated Cardiomyopathy. PLoS ONE 2016, 11, e0160110. [Google Scholar] [CrossRef] [Green Version]
- Rubiś, P.; Wiśniowska-Śmiałek, S.; Dziewięcka, E.; Rudnicka-Sosin, L.; Kozanecki, A.; Podolec, P. Prognostic value of fibrosis-related markers in dilated cardiomyopathy: A link between osteopontin and cardiovascular events. Adv. Med. Sci. 2018, 63, 160–166. [Google Scholar] [CrossRef]
- Dai, J.; Matsui, T.; Abel, E.D.; Dedhar, S.; Gerszten, R.E.; Seidman, C.E.; Seidman, J.G.; Rosenzweig, A. Deep sequence analysis of gene expression identifies osteopontin as a downstream effector of integrin-linked kinase (ILK) in cardiac-specific ILK knockout mice. Circ. Heart Fail. 2014, 7, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, W.; Zhang, J.; Liang, T.; Fan, G.P.; Wang, Z.W.; Zhang, P.D.; Wang, X.; Zhang, J. Inhibition of osteopontin reduce the cardiac myofibrosis in dilated cardiomyopathy via focal adhesion kinase mediated signaling pathway. Am. J. Transl. Res. 2016, 8, 3645–3655. [Google Scholar]
- Ikeda, T.; Shirasawa, T.; Esaki, Y.; Yoshiki, S.; Hirokawa, K. Osteopontin mRNA is expressed by smooth muscle-derived foam cells in human atherosclerotic lesions of the aorta. J. Clin. Investig. 1993, 92, 2814–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobescu-Ciobanu, C.; Giuşcă, S.E.; Căruntu, I.D.; Amălinei, C.; Rusu, A.; Cojocaru, E.; Popa, R.F.; Lupaşcu, C.D. Osteopontin and osteoprotegerin in atherosclerotic plaque—Are they significant markers of plaque vulnerability? Rom. J. Morphol. Embryol. 2020, 61, 793–801. [Google Scholar] [CrossRef]
- Mazzone, A.; Parri, M.S.; Giannessi, D.; Ravani, M.; Vaghetti, M.; Altieri, P.; Casalino, L.; Maltinti, M.; Balbi, M.; Barsotti, A.; et al. Osteopontin plasma levels and accelerated atherosclerosis in patients with CAD undergoing PCI: A prospective clinical study. Coron. Artery Dis. 2011, 22, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polonskaya, Y.V.; Kashtanova, E.V.; Murashov, I.S.; Kurguzov, A.V.; Sadovski, E.V.; Maslatsov, N.A.; Stakhneva, E.M.; Chernyavskii, A.M.; Ragino, Y.I. The Influence of Calcification Factors and Endothelial-Dysfunction Factors on the Development of Unstable Atherosclerotic Plaques. Diagnostics 2020, 10, 1074. [Google Scholar] [CrossRef] [PubMed]
- Matsui, Y.; Rittling, S.R.; Okamoto, H.; Inobe, M.; Jia, N.; Shimizu, T.; Akino, M.; Sugawara, T.; Morimoto, J.; Kimura, C.; et al. Osteopontin deficiency attenuates atherosclerosis in female apolipoprotein E-deficient mice. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1029–1034. [Google Scholar] [CrossRef]
- Isoda, K.; Kamezawa, Y.; Ayaori, M.; Kusuhara, M.; Tada, N.; Ohsuzu, F. Osteopontin transgenic mice fed a high-cholesterol diet develop early fatty-streak lesions. Circulation 2003, 107, 679–681. [Google Scholar] [CrossRef] [Green Version]
- Kamińska, J.; Stopiński, M.; Mucha, K.; Pac, M.; Gołębiowski, M.; Niewczas, M.A.; Pączek, L.; Foroncewicz, B. Circulating Osteoprotegerin in Chronic Kidney Disease and All-Cause Mortality. Int. J. Gen. Med. 2021, 14, 2413–2420. [Google Scholar] [CrossRef]
- Speer, M.Y.; Chien, Y.C.; Quan, M.; Yang, H.Y.; Vali, H.; McKee, M.D.; Giachelli, C.M. Smooth muscle cells deficient in osteopontin have enhanced susceptibility to calcification in vitro. Cardiovasc. Res. 2005, 66, 324–333. [Google Scholar] [CrossRef] [Green Version]
- Jono, S.; Peinado, C.; Giachelli, C.M. Phosphorylation of osteopontin is required for inhibition of vascular smooth muscle cell calcification. J. Biol. Chem. 2000, 275, 20197–20203. [Google Scholar] [CrossRef] [Green Version]
- Grau, J.B.; Poggio, P.; Sainger, R.; Vernick, W.J.; Seefried, W.F.; Branchetti, E.; Field, B.C.; Bavaria, J.E.; Acker, M.A.; Ferrari, G. Analysis of osteopontin levels for the identification of asymptomatic patients with calcific aortic valve disease. Ann. Thorac. Surg. 2012, 93, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muka, T.; Koromani, F.; Portilla, E.; O’Connor, A.; Bramer, W.M.; Troup, J.; Chowdhury, R.; Dehghan, A.; Franco, O.H. The role of epigenetic modifications in cardiovascular disease: A systematic review. Int. J. Cardiol. 2016, 212, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Song, M.; Qu, J.; Liu, G.H. Epigenetic Modifications in Cardiovascular Aging and Diseases. Circ. Res. 2018, 123, 773–786. [Google Scholar] [CrossRef]
- Khyzha, N.; Alizada, A.; Wilson, M.D.; Fish, J.E. Epigenetics of Atherosclerosis: Emerging Mechanisms and Methods. Trends Mol. Med. 2017, 23, 332–347. [Google Scholar] [CrossRef]
- Zhang, W.; Li, Q.; Li, D.; Li, J.; Aki, D.; Liu, Y.C. The E3 ligase VHL controls alveolar macrophage function via metabolic-epigenetic regulation. J. Exp. Med. 2018, 215, 3180–3193. [Google Scholar] [CrossRef]
- Chang, S.; Huang, J.; Niu, H.; Wang, J.; Si, Y.; Bai, Z.; Cheng, S.; Ding, W. Epigenetic regulation of osteopontin splicing isoform c defines its role as a microenvironmental factor to promote the survival of colon cancer cells from 5-FU treatment. Cancer Cell Int. 2020, 20, 452. [Google Scholar] [CrossRef]
- Moss, A.J.; Sim, A.M.; Adamson, P.D.; Seidman, M.A.; Andrews, J.P.M.; Doris, M.K.; Shah, A.S.V.; BouHaidar, R.; Alcaide-Corral, C.J.; Williams, M.C.; et al. Ex vivo (18)F-fluoride uptake and hydroxyapatite deposition in human coronary atherosclerosis. Sci. Rep. 2020, 10, 20172. [Google Scholar] [CrossRef]
- Steitz, S.A.; Speer, M.Y.; McKee, M.D.; Liaw, L.; Almeida, M.; Yang, H.; Giachelli, C.M. Osteopontin inhibits mineral deposition and promotes regression of ectopic calcification. Am. J. Pathol. 2002, 161, 2035–2046. [Google Scholar] [CrossRef] [Green Version]
- Ohri, R.; Tung, E.; Rajachar, R.; Giachelli, C.M. Mitigation of ectopic calcification in osteopontin-deficient mice by exogenous osteopontin. Calcif. Tissue Int. 2005, 76, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Ruan, C.C.; Ma, Y.; Tang, X.F.; Wu, Q.H.; Wang, J.G.; Zhu, D.L.; Gao, P.J. Osteopontin regulates macrophage activation and osteoclast formation in hypertensive patients with vascular calcification. Sci. Rep. 2017, 7, 40253. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in atherosclerosis: A dynamic balance. Nat. Rev. Immunol. 2013, 13, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Wang, Y.; Zhang, R.; Gao, Q.; Liang, X.; Gao, L.; Jiang, Z.; Qiao, R.; Han, D.; Zhang, Y.; et al. MRI/optical dual-modality imaging of vulnerable atherosclerotic plaque with an osteopontin-targeted probe based on Fe(3)O(4) nanoparticles. Biomaterials 2017, 112, 336–345. [Google Scholar] [CrossRef]
- Li, S.; Gou, T.; Wang, Q.; Chen, M.; Chen, Z.; Xu, M.; Wang, Y.; Han, D.; Cao, R.; Liu, J.; et al. Ultrasound/Optical Dual-Modality Imaging for Evaluation of Vulnerable Atherosclerotic Plaques with Osteopontin Targeted Nanoparticles. Macromol. Biosci. 2020, 20, e1900279. [Google Scholar] [CrossRef]
- Kim, K.; Shim, D.; Lee, J.S.; Zaitsev, K.; Williams, J.W.; Kim, K.W.; Jang, M.Y.; Seok Jang, H.; Yun, T.J.; Lee, S.H.; et al. Transcriptome Analysis Reveals Nonfoamy Rather Than Foamy Plaque Macrophages Are Proinflammatory in Atherosclerotic Murine Models. Circ. Res. 2018, 123, 1127–1142. [Google Scholar] [CrossRef]
- Williams, J.W.; Zaitsev, K.; Kim, K.W.; Ivanov, S.; Saunders, B.T.; Schrank, P.R.; Kim, K.; Elvington, A.; Kim, S.H.; Tucker, C.G.; et al. Limited proliferation capacity of aortic intima resident macrophages requires monocyte recruitment for atherosclerotic plaque progression. Nat. Immunol. 2020, 21, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Renault, M.A.; Robbesyn, F.; Réant, P.; Douin, V.; Daret, D.; Allières, C.; Belloc, I.; Couffinhal, T.; Arnal, J.F.; Klingel, K.; et al. Osteopontin expression in cardiomyocytes induces dilated cardiomyopathy. Circ. Heart Fail. 2010, 3, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, I.A.; Mraiche, F. Targeting osteopontin, the silent partner of Na+/H+ exchanger isoform 1 in cardiac remodeling. J. Cell. Physiol. 2015, 230, 2006–2018. [Google Scholar] [CrossRef] [PubMed]
- Dahiya, S.; Givvimani, S.; Bhatnagar, S.; Qipshidze, N.; Tyagi, S.C.; Kumar, A. Osteopontin-stimulated expression of matrix metalloproteinase-9 causes cardiomyopathy in the mdx model of Duchenne muscular dystrophy. J. Immunol. 2011, 187, 2723–2731. [Google Scholar] [CrossRef] [Green Version]
- Caballero, E.P.; Santamaría, M.H.; Corral, R.S. Endogenous osteopontin induces myocardial CCL5 and MMP-2 activation that contributes to inflammation and cardiac remodeling in a mouse model of chronic Chagas heart disease. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 11–23. [Google Scholar] [CrossRef]
- Kristen, A.V.; Rosenberg, M.; Lindenmaier, D.; Merkle, C.; Steen, H.; Andre, F.; Schönland, S.O.; Schnabel, P.A.; Schuster, T.; Röcken, C.; et al. Osteopontin: A novel predictor of survival in patients with systemic light-chain amyloidosis. Amyloid Int. J. Exp. Clin. Investig. Off. J. Int. Soc. Amyloidosis 2014, 21, 202–210. [Google Scholar] [CrossRef]
- Pei, H.; Zhang, H.; Tian, C.; Sun, X.; Qian, X.; Meng, Y.; Guo, X.; Chang, Q. Proliferative Vascular Smooth Muscle Cells Stimulate Extracellular Matrix Production via Osteopontin/p38 MAPK Signaling Pathway. Cardiology 2021, 1–10. [Google Scholar] [CrossRef]
- Xie, Y.; Sakatsume, M.; Nishi, S.; Narita, I.; Arakawa, M.; Gejyo, F. Expression, roles, receptors, and regulation of osteopontin in the kidney. Kidney Int. 2001, 60, 1645–1657. [Google Scholar] [CrossRef] [Green Version]
- Lorenzen, J.M.; Hafer, C.; Faulhaber-Walter, R.; Kümpers, P.; Kielstein, J.T.; Haller, H.; Fliser, D. Osteopontin predicts survival in critically ill patients with acute kidney injury. Nephrol. Dial. Transplant. Off. Publ. Eur. Dial. Transpl. Assoc. Eur. Ren. Assoc. 2011, 26, 531–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beitland, S.; Nakstad, E.R.; Berg, J.P.; Trøseid, A.S.; Brusletto, B.S.; Brunborg, C.; Lundqvist, C.; Sunde, K. Urine β-2-Microglobulin, Osteopontin, and Trefoil Factor 3 May Early Predict Acute Kidney Injury and Outcome after Cardiac Arrest. Crit. Care Res. Pract. 2019, 2019, 4384796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Z.K.; Tian, P.X.; Wang, X.Z.; Xue, W.J.; Ding, X.M.; Zheng, J.; Ding, C.G.; Mao, T.C.; Duan, W.L.; Xi, M. Kidney injury molecule-1 and osteopontin: New markers for prediction of early kidney transplant rejection. Mol. Immunol. 2013, 54, 457–464. [Google Scholar] [CrossRef]
- Zhang, Z.X.; Shek, K.; Wang, S.; Huang, X.; Lau, A.; Yin, Z.; Sun, H.; Liu, W.; Garcia, B.; Rittling, S.; et al. Osteopontin expressed in tubular epithelial cells regulates NK cell-mediated kidney ischemia reperfusion injury. J. Immunol. 2010, 185, 967–973. [Google Scholar] [CrossRef]
- Cen, C.; Aziz, M.; Yang, W.L.; Nicastro, J.M.; Coppa, G.F.; Wang, P. Osteopontin Blockade Attenuates Renal Injury After Ischemia Reperfusion by Inhibiting NK Cell Infiltration. Shock 2017, 47, 52–60. [Google Scholar] [CrossRef]
- Persy, V.P.; Verhulst, A.; Ysebaert, D.K.; De Greef, K.E.; De Broe, M.E. Reduced postischemic macrophage infiltration and interstitial fibrosis in osteopontin knockout mice. Kidney Int. 2003, 63, 543–553. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, H.; Igarashi, M.; Hirata, A.; Tsuchiya, H.; Sugiyama, K.; Morita, Y.; Jimbu, Y.; Ohnuma, H.; Daimon, M.; Tominaga, M.; et al. Progression of diabetic nephropathy enhances the plasma osteopontin level in type 2 diabetic patients. Endocr. J. 2004, 51, 499–504. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Sano, M.; Lu, L.; Wang, W.; Zhang, Q.; Zhang, R.; Wang, L.; Chen, Q.; Fukuda, K.; Shen, W. Plasma concentrations of osteopontin, but not thrombin-cleaved osteopontin, are associated with the presence and severity of nephropathy and coronary artery disease in patients with type 2 diabetes mellitus. Cardiovasc. Diabetol. 2010, 9, 70. [Google Scholar] [CrossRef] [Green Version]
- Gordin, D.; Forsblom, C.; Panduru, N.M.; Thomas, M.C.; Bjerre, M.; Soro-Paavonen, A.; Tolonen, N.; Sandholm, N.; Flyvbjerg, A.; Harjutsalo, V.; et al. Osteopontin is a strong predictor of incipient diabetic nephropathy, cardiovascular disease, and all-cause mortality in patients with type 1 diabetes. Diabetes Care 2014, 37, 2593–2600. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Bompada, P.; Atac, D.; Laakso, M.; Groop, L.; De Marinis, Y. Epigenetic regulation of glucose-stimulated osteopontin (OPN) expression in diabetic kidney. Biochem. Biophys. Res. Commun. 2016, 469, 108–113. [Google Scholar] [CrossRef]
- Susztak, K.; Böttinger, E.; Novetsky, A.; Liang, D.; Zhu, Y.; Ciccone, E.; Wu, D.; Dunn, S.; McCue, P.; Sharma, K. Molecular profiling of diabetic mouse kidney reveals novel genes linked to glomerular disease. Diabetes 2004, 53, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Wolak, T.; Kim, H.; Ren, Y.; Kim, J.; Vaziri, N.D.; Nicholas, S.B. Osteopontin modulates angiotensin II-induced inflammation, oxidative stress, and fibrosis of the kidney. Kidney Int. 2009, 76, 32–43. [Google Scholar] [CrossRef] [Green Version]
- Irita, J.; Okura, T.; Jotoku, M.; Nagao, T.; Enomoto, D.; Kurata, M.; Desilva, V.R.; Miyoshi, K.; Matsui, Y.; Uede, T.; et al. Osteopontin deficiency protects against aldosterone-induced inflammation, oxidative stress, and interstitial fibrosis in the kidney. Am. J. Physiol. Ren. Physiol. 2011, 301, F833–F844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolak, T. Osteopontin—A multi-modal marker and mediator in atherosclerotic vascular disease. Atherosclerosis 2014, 236, 327–337. [Google Scholar] [CrossRef]
- Lancha, A.; Rodríguez, A.; Catalán, V.; Becerril, S.; Sáinz, N.; Ramírez, B.; Burrell, M.A.; Salvador, J.; Frühbeck, G.; Gómez-Ambrosi, J. Osteopontin deletion prevents the development of obesity and hepatic steatosis via impaired adipose tissue matrix remodeling and reduced inflammation and fibrosis in adipose tissue and liver in mice. PLoS ONE 2014, 9, e98398. [Google Scholar] [CrossRef] [PubMed]
- Nomiyama, T.; Perez-Tilve, D.; Ogawa, D.; Gizard, F.; Zhao, Y.; Heywood, E.B.; Jones, K.L.; Kawamori, R.; Cassis, L.A.; Tschöp, M.H.; et al. Osteopontin mediates obesity-induced adipose tissue macrophage infiltration and insulin resistance in mice. J. Clin. Investig. 2007, 117, 2877–2888. [Google Scholar] [CrossRef] [PubMed]
- Shirakawa, K.; Endo, J.; Katsumata, Y.; Yamamoto, T.; Kataoka, M.; Isobe, S.; Yoshida, N.; Fukuda, K.; Sano, M. Negative legacy of obesity. PLoS ONE 2017, 12, e0186303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luna-Luna, M.; Criales-Vera, S.; Medina-Leyte, D.; Díaz-Zamudio, M.; Flores-Zapata, A.; Cruz-Robles, D.; López-Meneses, M.; Olvera-Cruz, S.; Ramírez-Marroquín, S.; Flores-Castillo, C.; et al. Bone Morphogenetic Protein-2 and Osteopontin Gene Expression in Epicardial Adipose Tissue from Patients with Coronary Artery Disease Is Associated with the Presence of Calcified Atherosclerotic Plaques. Diabetes Metab. Syndr. Obes. Targets Ther. 2020, 13, 1943–1951. [Google Scholar] [CrossRef]
- Rangaswami, J.; Bhalla, V.; Blair, J.E.A.; Chang, T.I.; Costa, S.; Lentine, K.L.; Lerma, E.V.; Mezue, K.; Molitch, M.; Mullens, W.; et al. Cardiorenal Syndrome: Classification, Pathophysiology, Diagnosis, and Treatment Strategies: A Scientific Statement From the American Heart Association. Circulation 2019, 139, e840–e878. [Google Scholar] [CrossRef]
- Chen, J.; Lu, Y.; Huang, D.; Luo, X.; Zhang, Y. Relationship of osteopontin and renal function with severity of coronary artery lesions. Int. J. Clin. Exp. Med. 2014, 7, 1122–1127. [Google Scholar]
- Lorenzen, J.; Krämer, R.; Kliem, V.; Bode-Boeger, S.M.; Veldink, H.; Haller, H.; Fliser, D.; Kielstein, J.T. Circulating levels of osteopontin are closely related to glomerular filtration rate and cardiovascular risk markers in patients with chronic kidney disease. Eur. J. Clin. Investig. 2010, 40, 294–300. [Google Scholar] [CrossRef]
- Zinman, B.; Wanner, C.; Lachin, J.M.; Fitchett, D.; Bluhmki, E.; Hantel, S.; Mattheus, M.; Devins, T.; Johansen, O.E.; Woerle, H.J.; et al. Empagliflozin, Cardiovascular Outcomes, and Mortality in Type 2 Diabetes. N. Engl. J. Med. 2015, 373, 2117–2128. [Google Scholar] [CrossRef]
- Rådholm, K.; Figtree, G.; Perkovic, V.; Solomon, S.D.; Mahaffey, K.W.; de Zeeuw, D.; Fulcher, G.; Barrett, T.D.; Shaw, W.; Desai, M.; et al. Canagliflozin and Heart Failure in Type 2 Diabetes Mellitus: Results From the CANVAS Program. Circulation 2018, 138, 458–468. [Google Scholar] [CrossRef] [PubMed]
- Wiviott, S.D.; Raz, I.; Bonaca, M.P.; Mosenzon, O.; Kato, E.T.; Cahn, A.; Silverman, M.G.; Zelniker, T.A.; Kuder, J.F.; Murphy, S.A.; et al. Dapagliflozin and Cardiovascular Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2019, 380, 347–357. [Google Scholar] [CrossRef] [PubMed]
- McMurray, J.J.V.; Solomon, S.D.; Inzucchi, S.E.; Køber, L.; Kosiborod, M.N.; Martinez, F.A.; Ponikowski, P.; Sabatine, M.S.; Anand, I.S.; Bělohlávek, J.; et al. Dapagliflozin in Patients with Heart Failure and Reduced Ejection Fraction. N. Engl. J. Med. 2019, 381, 1995–2008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef]
- Kang, H.M.; Ahn, S.H.; Choi, P.; Ko, Y.A.; Han, S.H.; Chinga, F.; Park, A.S.; Tao, J.; Sharma, K.; Pullman, J.; et al. Defective fatty acid oxidation in renal tubular epithelial cells has a key role in kidney fibrosis development. Nat. Med. 2015, 21, 37–46. [Google Scholar] [CrossRef]
- Lan, R.; Geng, H.; Singha, P.K.; Saikumar, P.; Bottinger, E.P.; Weinberg, J.M.; Venkatachalam, M.A. Mitochondrial Pathology and Glycolytic Shift during Proximal Tubule Atrophy after Ischemic AKI. J. Am. Soc. Nephrol. 2016, 27, 3356–3367. [Google Scholar] [CrossRef] [Green Version]
- Sas, K.M.; Kayampilly, P.; Byun, J.; Nair, V.; Hinder, L.M.; Hur, J.; Zhang, H.; Lin, C.; Qi, N.R.; Michailidis, G.; et al. Tissue-specific metabolic reprogramming drives nutrient flux in diabetic complications. JCI Insight 2016, 1, e86976. [Google Scholar] [CrossRef]
- Zhan, M.; Usman, I.M.; Sun, L.; Kanwar, Y.S. Disruption of renal tubular mitochondrial quality control by Myo-inositol oxygenase in diabetic kidney disease. J. Am. Soc. Nephrol. 2015, 26, 1304–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, J.; Fasching, A.; Pihl, L.; Patinha, D.; Franzén, S.; Palm, F. Acute SGLT inhibition normalizes O2 tension in the renal cortex but causes hypoxia in the renal medulla in anaesthetized control and diabetic rats. Am. J. Physiol. Ren. Physiol. 2015, 309, F227–F234. [Google Scholar] [CrossRef] [Green Version]
- Sharma, I.; Deng, F.; Liao, Y.; Kanwar, Y.S. Myo-inositol Oxygenase (MIOX) Overexpression Drives the Progression of Renal Tubulointerstitial Injury in Diabetes. Diabetes 2020, 69, 1248–1263. [Google Scholar] [CrossRef]
- Fleury, C.; Mignotte, B.; Vayssière, J.L. Mitochondrial reactive oxygen species in cell death signaling. Biochimie 2002, 84, 131–141. [Google Scholar] [CrossRef]
- Sano, M.; Goto, S. Possible Mechanism of Hematocrit Elevation by Sodium Glucose Cotransporter 2 Inhibitors and Associated Beneficial Renal and Cardiovascular Effects. Circulation 2019, 139, 1985–1987. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
OPN in cardiac remodeling. OPN is involved in a number of CVDs, including ischemic heart diseases, hypertension, heart failure, dilated cardiomyopathy, atherosclerosis, and several cardiomyopathies and the major source and role of OPN is dependent on pathological states. A number of stimuli are shown to promote OPN expression, including cytokines, ROS, Ang II (angiotensin II), high glucose, hypoxia, and epigenetic mechanisms. Different cell types and pathological states differ in their regulatory mechanisms of OPN in the heart and vessels.
Figure 1.
OPN in cardiac remodeling. OPN is involved in a number of CVDs, including ischemic heart diseases, hypertension, heart failure, dilated cardiomyopathy, atherosclerosis, and several cardiomyopathies and the major source and role of OPN is dependent on pathological states. A number of stimuli are shown to promote OPN expression, including cytokines, ROS, Ang II (angiotensin II), high glucose, hypoxia, and epigenetic mechanisms. Different cell types and pathological states differ in their regulatory mechanisms of OPN in the heart and vessels.

Figure 2.
Vicious cycle of OPN in CRS. OPN is involved in the pathogenesis of CRS. Acute or chronic dysfunction of the heart or kidneys can induce OPN expression and may result in acute or chronic dysfunction in the other organ. Several stimuli induce OPN in the pathological conditions of the heart and kidney and OPN causes cardiac fibrosis, cardiac hypertrophy, LV dysfunction, and tubulointerstitial fibrosis.
Figure 2.
Vicious cycle of OPN in CRS. OPN is involved in the pathogenesis of CRS. Acute or chronic dysfunction of the heart or kidneys can induce OPN expression and may result in acute or chronic dysfunction in the other organ. Several stimuli induce OPN in the pathological conditions of the heart and kidney and OPN causes cardiac fibrosis, cardiac hypertrophy, LV dysfunction, and tubulointerstitial fibrosis.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shirakawa, K.; Sano, M. Osteopontin in Cardiovascular Diseases. Biomolecules 2021, 11, 1047. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071047
AMA Style
Shirakawa K, Sano M. Osteopontin in Cardiovascular Diseases. Biomolecules. 2021; 11(7):1047. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071047
Chicago/Turabian StyleShirakawa, Kohsuke, and Motoaki Sano. 2021. "Osteopontin in Cardiovascular Diseases" Biomolecules 11, no. 7: 1047. https://0-doi-org.brum.beds.ac.uk/10.3390/biom11071047
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.