
Docosahexaenoic Acid and Melatonin Prevent Impaired Oligodendrogenesis Induced by Intrauterine Growth Restriction (IUGR)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
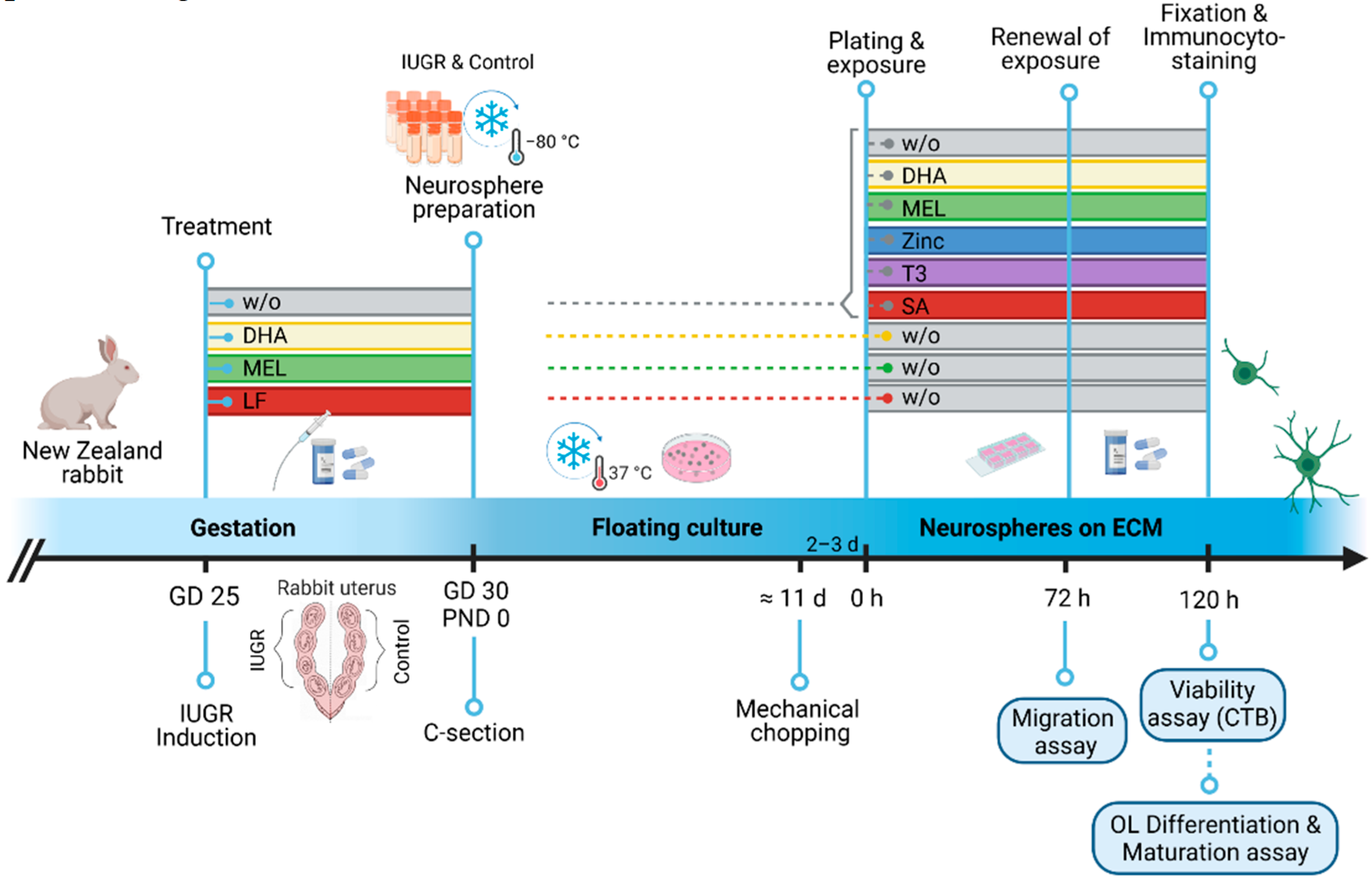
2.1. In Vivo Procedures: IUGR Induction and Administration of Therapies
2.2. Neurosphere Preparation
2.3. The Neurosphere Assay
2.3.1. In Vitro Testing of Potential Therapies
2.3.2. Migration Assay
2.3.3. OL Differentiation and Maturation Assay
2.3.4. Cell Viability
2.3.5. qRT-PCR
2.4. Statistics
3. Results
3.1. IUGR Decreases OL Differentiation
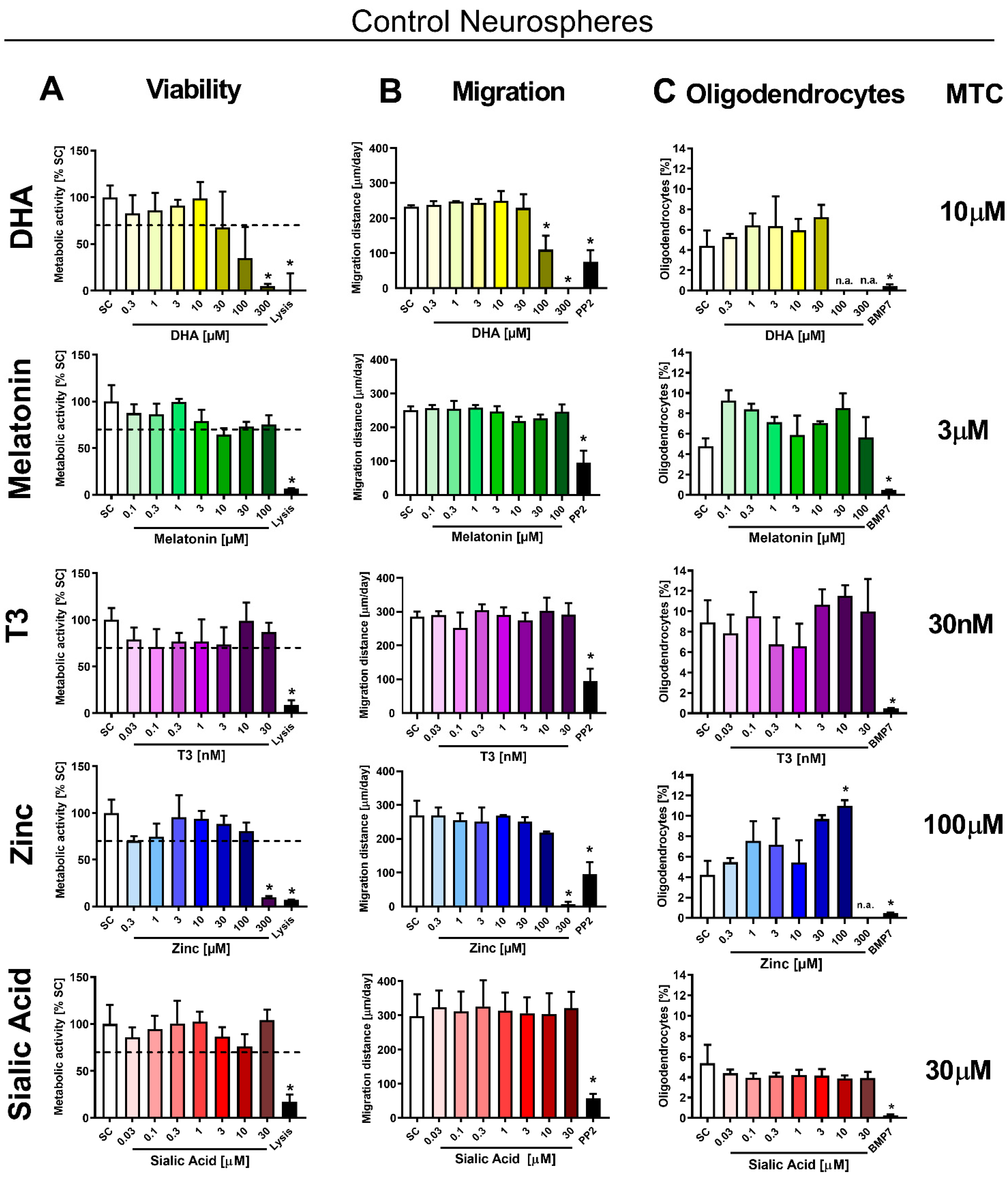
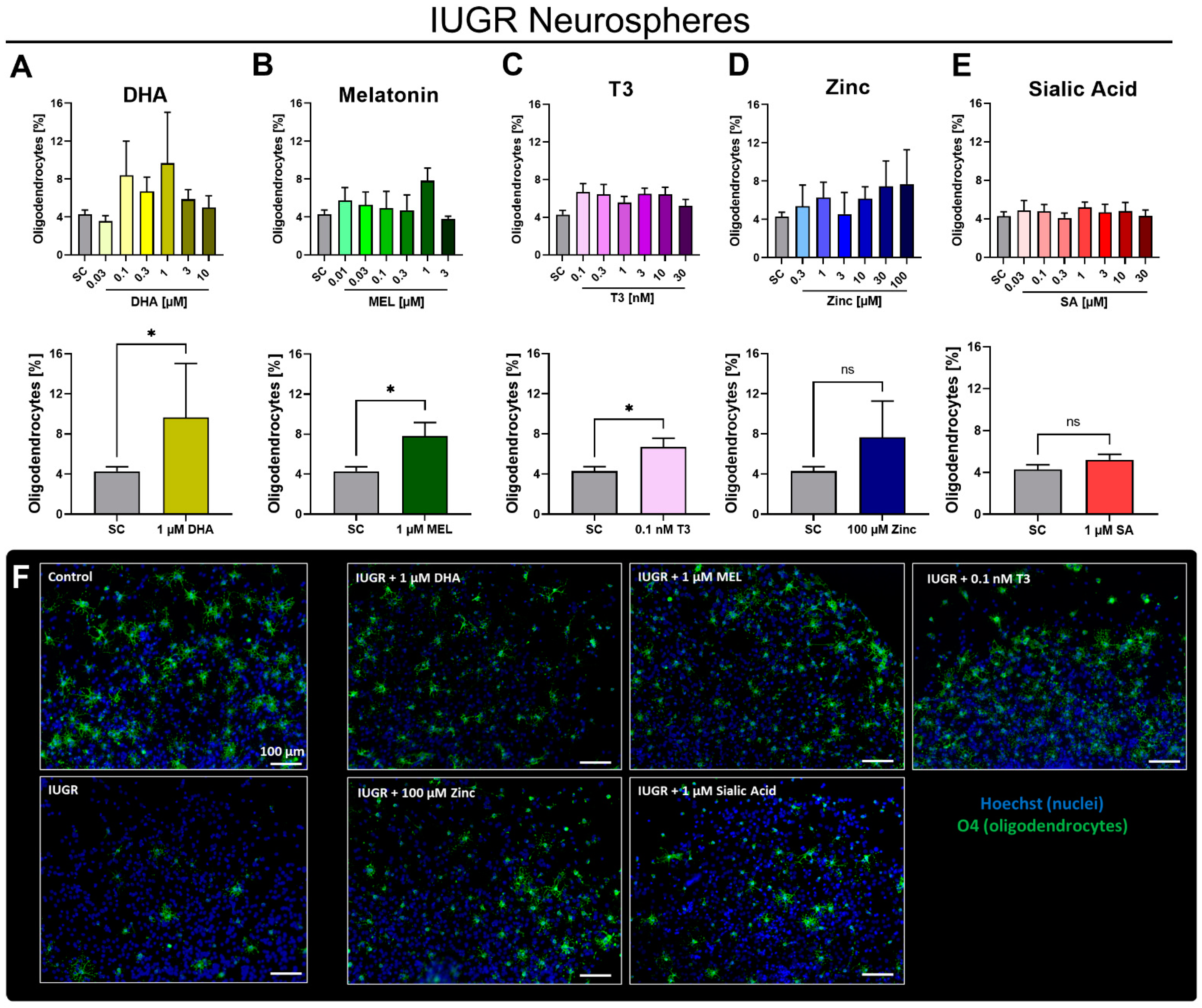
3.2. In Vitro Testing of Potential Therapies
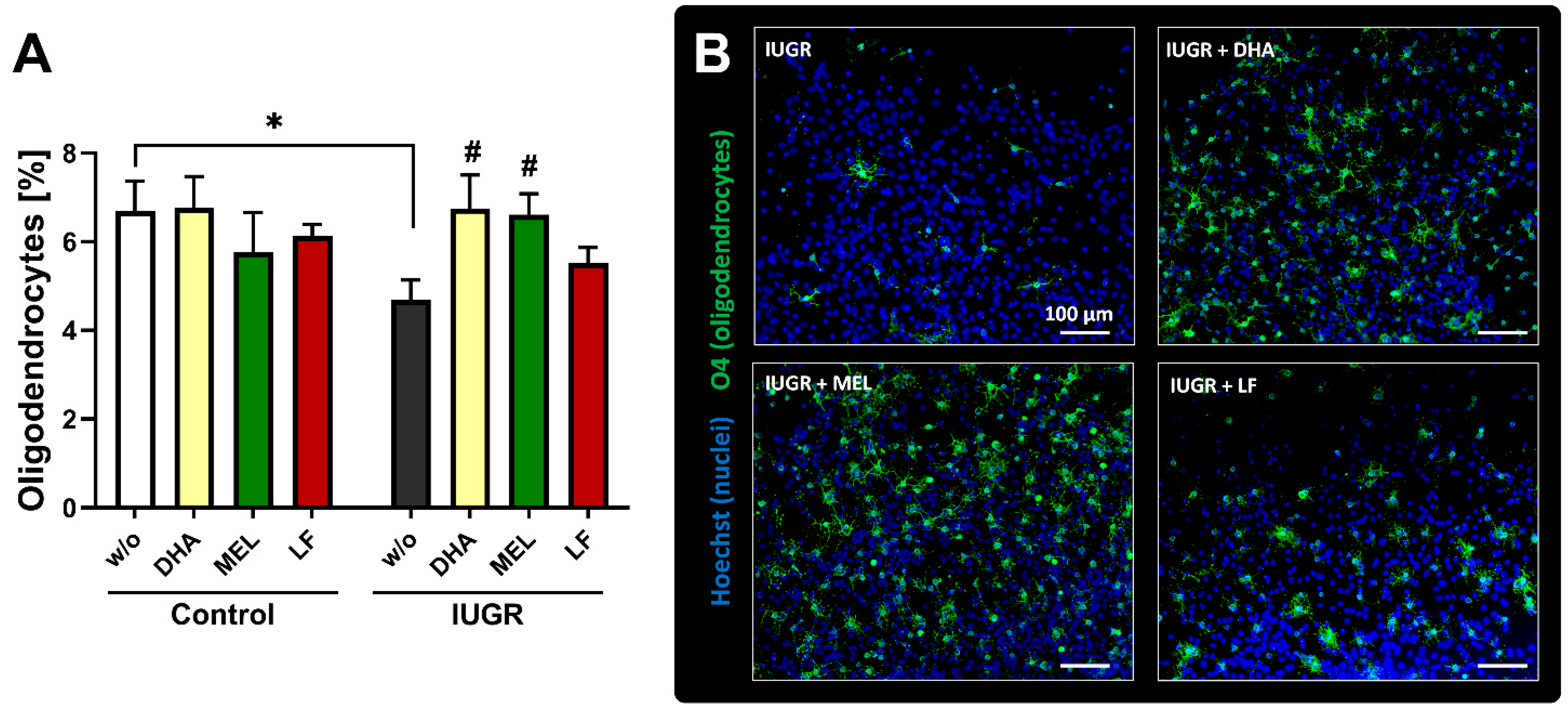
3.3. In Vivo Administration of Selected Therapies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mwaniki, M.K.; Atieno, M.; Lawn, J.E.; Newton, C.R. Long-term neurodevelopmental outcomes after intrauterine and neonatal insults: A systematic review. Lancet 2012, 379, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Kady, S.M.; Gardosi, J. Perinatal mortality and fetal growth restriction. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 397–410. [Google Scholar] [CrossRef] [PubMed]
- Esteban, F.J.; Padilla, N.; Sanz-Cortes, M.; de Miras, J.R.; Bargallo, N.; Villoslada, P.; Gratacos, E. Fractal-dimension analysis detects cerebral changes in preterm infants with and without intrauterine growth restriction. NeuroImage 2010, 53, 1225–1232. [Google Scholar] [CrossRef]
- Pla, L.; Illa, M.; Loreiro, C.; Lopez, M.C.; Vázquez-Aristizabal, P.; Kühne, B.A.; Barenys, M.; Eixarch, E.; Gratacós, E. Structural Brain Changes during the Neonatal Period in a Rabbit Model of Intrauterine Growth Restriction. Dev. Neurosci. 2020, 42, 217–229. [Google Scholar] [CrossRef]
- Reid, M.V.; Murray, K.A.; Marsh, E.D.; Golden, J.A.; Simmons, R.A.; Grinspan, J.B. Delayed Myelination in an Intrauterine Growth Retardation Model Is Mediated by Oxidative Stress Upregulating Bone Morphogenetic Protein 4. J. Neuropathol. Exp. Neurol. 2012, 71, 640–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novais, A.R.B.; Pham, H.; van de Looij, Y.; Bernal, M.; Mairesse, J.; Zana-Taieb, E.; Colella, M.; Jarreau, P.-H.; Pansiot, J.; Dumont, F.; et al. Transcriptomic regulations in oligodendroglial and microglial cells related to brain damage following fetal growth restriction. Glia 2016, 64, 2306–2320. [Google Scholar] [CrossRef] [Green Version]
- Eixarch, E.; Batalle, D.; Illa, M.; Muñoz-Moreno, E.; Arbat-Plana, A.; Amat-Roldan, I.; Figueras, F.; Gratacos, E. Neonatal Neurobehavior and Diffusion MRI Changes in Brain Reorganization Due to Intrauterine Growth Restriction in a Rabbit Model. PLoS ONE 2012, 7, e31497. [Google Scholar] [CrossRef] [Green Version]
- Tolcos, M.; Bateman, E.; O’Dowd, R.; Markwick, R.; Vrijsen, K.; Rehn, A.; Rees, S. Intrauterine growth restriction affects the maturation of myelin. Exp. Neurol. 2011, 232, 53–65. [Google Scholar] [CrossRef]
- Abel, K.M.; Dalman, C.; Svensson, A.C.; Susser, E.; Dal, H.; Idring, S.; Webb, R.T.; Rai, D.; Magnusson, C. Deviance in Fetal Growth and Risk of Autism Spectrum Disorder. Am. J. Psychiatry 2013, 170, 391–398. [Google Scholar] [CrossRef]
- Leitner, Y.; Fattal-Valevski, A.; Geva, R.; Eshel, R.; Toledano-Alhadef, H.; Rotstein, M.; Bassan, H.; Radianu, B.; Bitchonsky, O.; Jaffa, A.J.; et al. Neurodevelopmental Outcome of Children with Intrauterine Growth Retardation: A Longitudinal, 10-Year Prospective Study. J. Child Neurol. 2007, 22, 580–587. [Google Scholar] [CrossRef]
- Illa, M.; Eixarch, E.; Batalle, D.; Arbat-Plana, A.; Muñoz-Moreno, E.; Figueras, F.; Gratacos, E. Long-Term Functional Outcomes and Correlation with Regional Brain Connectivity by MRI Diffusion Tractography Metrics in a Near-Term Rabbit Model of Intrauterine Growth Restriction. PLoS ONE 2013, 8, e76453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleiss, B.; Wong, F.; Brownfoot, F.; Shearer, I.K.; Baud, O.; Walker, D.W.; Gressens, P.; Tolcos, M. Knowledge Gaps and Emerging Research Areas in Intrauterine Growth Restriction-Associated Brain Injury. Front. Endocrinol. 2019, 10, 188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolcos, M.; Petratos, S.; Hirst, J.J.; Wong, F.; Spencer, S.J.; Azhan, A.; Emery, B.; Walker, D.W. Blocked, delayed, or obstructed: What causes poor white matter development in intrauterine growth restricted infants? Prog. Neurobiol. 2017, 154, 62–77. [Google Scholar] [CrossRef] [PubMed]
- Barenys, M.; Illa, M.; Hofrichter, M.; Loreiro, C.; Pla, L.; Klose, J.; Kühne, B.A.; Gómez-Catalán, J.; Braun, J.M.; Crispi, F.; et al. Rabbit neurospheres as a novel in vitro tool for studying neurodevelopmental effects induced by intrauterine growth restriction. Stem Cells Transl. Med. 2021, 10, 209–221. [Google Scholar] [CrossRef]
- Eixarch, E.; Figueras, F.; Hernández-Andrade, E.; Crispi, F.; Nadal, A.; Torre, I.; Oliveira, S.; Gratacos, E. An Experimental Model of Fetal Growth Restriction Based on Selective Ligature of Uteroplacental Vessels in the Pregnant Rabbit. Fetal Diagn. Ther. 2009, 26, 203–211. [Google Scholar] [CrossRef]
- Eixarch, E.; Hernandez-Andrade, E.; Crispi, F.; Illa, M.; Torre, I.; Figueras, F.; Gratacos, E. Impact on fetal mortality and cardiovascular Doppler of selective ligature of uteroplacental vessels compared with undernutrition in a rabbit model of intrauterine growth restriction. Placenta 2011, 32, 304–309. [Google Scholar] [CrossRef]
- Baumann, J.; Barenys, M.; Gassmann, K.; Fritsche, E. Comparative Human and Rat “Neurosphere Assay” for Developmental Neurotoxicity Testing. Curr. Protoc. Toxicol. 2014, 59, 12–21. [Google Scholar] [CrossRef]
- Moors, M.; Rockel, T.D.; Abel, J.; Cline, J.E.; Gassmann, K.; Schreiber, T.; Schuwald, J.; Weinmann, N.; Fritsche, E. Human Neurospheres as Three-Dimensional Cellular Systems for Developmental Neurotoxicity Testing. Environ. Health Perspect. 2009, 117, 1131–1138. [Google Scholar] [CrossRef]
- Siqueira, L.D.S.; Majolo, F.; da Silva, A.P.B.; da Costa, J.C.; Marinowic, D.R. Neurospheres: A potential in vitro model for the study of central nervous system disorders. Mol. Biol. Rep. 2021, 48, 3649–3663. [Google Scholar] [CrossRef]
- Masjosthusmann, S.; Siebert, C.; Hübenthal, U.; Bendt, F.; Baumann, J.; Fritsche, E. Arsenite interrupts neurodevelopmental processes of human and rat neural progenitor cells: The role of reactive oxygen species and species-specific antioxidative defense. Chemosphere 2019, 235, 447–456. [Google Scholar] [CrossRef]
- Dach, K.; Bendt, F.; Huebenthal, U.; Giersiefer, S.; Lein, P.; Heuer, H.; Fritsche, E. BDE-99 impairs differentiation of human and mouse NPCs into the oligodendroglial lineage by species-specific modes of action. Sci. Rep. 2017, 7, srep44861. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, J.A.; Bell, S.J.; Ausdal, W. Van Omega-3 Fatty Acid Supplementation during Pregnancy. Rev. Obstet. Gynecol. 2008, 1, 162–169. [Google Scholar] [PubMed]
- Gil-Sánchez, A.; Larqué, E.; Demmelmair, H.; Acien, M.I.; Faber, F.L.; Parrilla, J.J.; Koletzko, B. Maternal-fetal in vivo transfer of [13C]docosahexaenoic and other fatty acids across the human placenta 12 h after maternal oral intake. Am. J. Clin. Nutr. 2010, 92, 115–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.S.; Ciappolino, V.; Agostoni, C. DHA Effects in Brain Development and Function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, S.L.; Yawno, T.; Alers, N.O.; Castillo-Melendez, M.; Supramaniam, V.G.; Vanzyl, N.; Sabaretnam, T.; Loose, J.M.; Drummond, G.R.; Walker, D.W.; et al. Antenatal antioxidant treatment with melatonin to decrease newborn neurodevelopmental deficits and brain injury caused by fetal growth restriction. J. Pineal Res. 2014, 56, 283–294. [Google Scholar] [CrossRef]
- Nuttall, J.R.; Oteiza, P.I. Zinc and the ERK Kinases in the Developing Brain. Neurotox. Res. 2012, 21, 128–141. [Google Scholar] [CrossRef] [Green Version]
- Adamo, A.M.; Oteiza, P.I. Zinc deficiency and neurodevelopment: The case of neurons. BioFactors 2010, 36, 117–124. [Google Scholar] [CrossRef] [Green Version]
- Ladd, F.V.L.; Ladd, A.A.B.L.; Ribeiro, A.A.C.M.; Costa, S.B.C.; Coutinho, B.P.; Feitosa, G.A.S.; de Andrade, G.M.; de Castro-Costa, C.M.; Magalhães, C.E.C.; Castro, I.C.; et al. Zinc and glutamine improve brain development in suckling mice subjected to early postnatal malnutrition. Nutrition 2010, 26, 662–670. [Google Scholar] [CrossRef] [Green Version]
- Mathur, N.B.; Agarwal, D.K. Zinc supplementation in preterm neonates and neurological development: A randomized controlled trial. Indian Pediatr. 2015, 52, 951–955. [Google Scholar] [CrossRef]
- Fernández, M.; Paradisi, M.; Del Vecchio, G.; Giardino, L.; Calzà, L. Thyroid hormone induces glial lineage of primary neurospheres derived from non-pathological and pathological rat brain: Implications for remyelination-enhancing therapies. Int. J. Dev. Neurosci. 2009, 27, 769–778. [Google Scholar] [CrossRef]
- Baud, O.; Berkane, N. Hormonal Changes Associated with Intra-Uterine Growth Restriction: Impact on the Developing Brain and Future Neurodevelopment. Front. Endocrinol. 2019, 10, 179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernal, J. Thyroid Hormones and Brain Development. Vitam. Horm. 2005, 71, 95–122. [Google Scholar] [CrossRef] [PubMed]
- van de Looij, Y.; Larpin, C.; Cabungcal, J.-H.; Sanches, E.F.; Toulotte, A.; Do, K.Q.; Sizonenko, S.V. Nutritional Intervention for Developmental Brain Damage: Effects of Lactoferrin Supplementation in Hypocaloric Induced Intrauterine Growth Restriction Rat Pups. Front. Endocrinol. 2019, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B. Molecular Determinants of Milk Lactoferrin as a Bioactive Compound in Early Neurodevelopment and Cognition. J. Pediatr. 2016, 173, S29–S36. [Google Scholar] [CrossRef] [Green Version]
- Baumann, N.; Pham-Dinh, D. Biology of Oligodendrocyte and Myelin in the Mammalian Central Nervous System. Physiol. Rev. 2001, 81, 871–927. [Google Scholar] [CrossRef]
- Fumarola, A.; Di Fiore, A.; Dainelli, M.; Grani, G.; Carbotta, G.; Calvanese, A. Therapy of Hyperthyroidism in Pregnancy and Breastfeeding. Obstet. Gynecol. Surv. 2011, 66, 378–385. [Google Scholar] [CrossRef]
- Drobyshevsky, A.; Jiang, R.; Derrick, M.; Luo, K.; Tan, S. Functional correlates of central white matter maturation in perinatal period in rabbits. Exp. Neurol. 2014, 261, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Workman, A.D.; Charvet, C.J.; Clancy, B.; Darlington, R.B.; Finlay, B.L. Modeling Transformations of Neurodevelopmental Sequences across Mammalian Species. J. Neurosci. 2013, 33, 7368–7383. [Google Scholar] [CrossRef]
- Chini, M.; Hanganu-Opatz, I.L. Prefrontal Cortex Development in Health and Disease: Lessons from Rodents and Humans. Trends Neurosci. 2021, 44, 227–240. [Google Scholar] [CrossRef]
- Back, S.A. White matter injury in the preterm infant: Pathology and mechanisms. Acta Neuropathol. 2017, 134, 331–349. [Google Scholar] [CrossRef]
- Back, S.A.; Luo, N.L.; Borenstein, N.S.; Volpe, J.J.; Kinney, H.C. Arrested Oligodendrocyte Lineage Progression during Human Cerebral White Matter Development: Dissociation between the Timing of Progenitor Differentiation and Myelinogenesis. J. Neuropathol. Exp. Neurol. 2002, 61, 197–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segovia, K.N.; McClure, M.; Moravec, M.; Luo, N.L.; Wan, Y.; Gong, X.; Riddle, A.; Craig, A.; Struve, J.; Sherman, L.S.; et al. Arrested oligodendrocyte lineage maturation in chronic perinatal white matter injury. Ann. Neurol. 2008, 63, 520–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- French, H.M.; Reid, M.; Mamontov, P.; Simmons, R.A.; Grinspan, J.B. Oxidative stress disrupts oligodendrocyte maturation. J. Neurosci. Res. 2009, 87, 3076–3087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueras, F.; Caradeux, J.; Crispi, F.; Eixarch, E.; Peguero, A.; Gratacos, E. Diagnosis and surveillance of late-onset fetal growth restriction. Am. J. Obstet. Gynecol. 2018, 218, S790–S802.e1. [Google Scholar] [CrossRef] [Green Version]
- Figueras, F.; Gratacos, E. An integrated approach to fetal growth restriction. Best Pract. Res. Clin. Obstet. Gynaecol. 2017, 38, 48–58. [Google Scholar] [CrossRef]
- Maršál, K. Obstetric management of intrauterine growth restriction. Best Pract. Res. Clin. Obstet. Gynaecol. 2009, 23, 857–870. [Google Scholar] [CrossRef]
- Suganuma, H.; Arai, Y.; Kitamura, Y.; Hayashi, M.; Okumura, A.; Shimizu, T. Maternal docosahexaenoic acid-enriched diet prevents neonatal brain injury. Neuropathology 2010, 30, 597–605. [Google Scholar] [CrossRef]
- Bernardo, A.; Giammarco, M.L.; De Nuccio, C.; Ajmone-Cat, M.A.; Visentin, S.; De Simone, R.; Minghetti, L. Docosahexaenoic acid promotes oligodendrocyte differentiation via PPAR-γ signalling and prevents tumor necrosis factor-α-dependent maturational arrest. Biochim. et Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2017, 1862, 1013–1023. [Google Scholar] [CrossRef]
- Gustafson, K.M.; Carlson, S.E.; Colombo, J.; Yeh, H.-W.; Shaddy, D.J.; Li, S.; Kerling, E.H. Effects of docosahexaenoic acid supplementation during pregnancy on fetal heart rate and variability: A randomized clinical trial. Prostaglandins Leukot. Essent. Fat. Acids 2013, 88, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Mozurkewich, E.L.; Clinton, C.M.; Chilimigras, J.L.; Hamilton, S.E.; Allbaugh, L.J.; Berman, D.R.; Marcus, S.M.; Romero, V.C.; Treadwell, M.C.; Keeton, K.L.; et al. The Mothers, Omega-3, and Mental Health Study: A double-blind, randomized controlled trial. Am. J. Obstet. Gynecol. 2013, 208, 313.e1–313.e9. [Google Scholar] [CrossRef] [Green Version]
- Reiter, R.J.; Tan, D.X.; Korkmaz, A.; Rosales-Corral, S.A. Melatonin and stable circadian rhythms optimize maternal, placental and fetal physiology. Hum. Reprod. Update 2014, 20, 293–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbets, A. Melatonin, placental growth factor and placental hormones at placental insufficiency. Cell Organ Transpl. 2019, 7, 103–107. [Google Scholar] [CrossRef]
- Berbets, A.M.; Davydenko, I.S.; Barbe, A.M.; Konkov, D.H.; Albota, O.M.; Yuzko, O.M. Melatonin 1A and 1B Receptors’ Expression Decreases in the Placenta of Women with Fetal Growth Restriction. Reprod. Sci. 2021, 28, 197–206. [Google Scholar] [CrossRef]
- Castillo-Melendez, M.; Yawno, T.; Sutherland, A.; Jenkin, G.; Wallace, E.M.; Miller, S.L. Effects of Antenatal Melatonin Treatment on the Cerebral Vasculature in an Ovine Model of Fetal Growth Restriction. Dev. Neurosci. 2017, 39, 323–337. [Google Scholar] [CrossRef] [PubMed]
- Olivier, P.; Fontaine, R.H.; Loron, G.; Van Steenwinckel, J.; Biran, V.; Massonneau, V.; Kaindl, A.; Dalous, J.; Charriaut-Marlangue, C.; Aigrot, M.-S.; et al. Melatonin Promotes Oligodendroglial Maturation of Injured White Matter in Neonatal Rats. PLoS ONE 2009, 4, e7128. [Google Scholar] [CrossRef]
- Palmer, K.R.; Mockler, J.C.; Davies-Tuck, M.L.; Miller, S.L.; Goergen, S.K.; Fahey, M.C.; Anderson, P.J.; Groom, K.M.; Wallace, E.M. Protect-me: A parallel-group, triple blinded, placebo-controlled randomised clinical trial protocol assessing antenatal maternal melatonin supplementation for fetal neuroprotection in early-onset fetal growth restriction. BMJ Open 2019, 9, e028243. [Google Scholar] [CrossRef] [PubMed]
- Klose, J.; Pahl, M.; Bartmann, K.; Bendt, F.; Blum, J.; Dolde, X.; Förster, N.; Holzer, A.-K.; Hübenthal, U.; Keßel, H.E.; et al. Neurodevelopmental toxicity assessment of flame retardants using a human DNT in vitro testing battery. Cell Biol. Toxicol. 2021, 5, 1–45. [Google Scholar] [CrossRef]
- Kilby, M.D.; Gittoes, N.; McCabe, C.; Verhaeg, J.; Franklyn, J.A. Expression of thyroid receptor isoforms in the human fetal central nervous system and the effects of intrauterine growth restriction. Clin. Endocrinol. 2000, 53, 469–477. [Google Scholar] [CrossRef]
- LaFranchi, S.H. Thyroid Function in Preterm/Low Birth Weight Infants: Impact on Diagnosis and Management of Thyroid Dysfunction. Front. Endocrinol. 2021, 12, 371. [Google Scholar] [CrossRef]
- van de Looij, Y.; Ginet, V.; Chatagner, A.; Toulotte, A.; Somm, E.; Hüppi, P.S.; Sizonenko, S.V. Lactoferrin during lactation protects the immature hypoxic-ischemic rat brain. Ann. Clin. Transl. Neurol. 2014, 1, 955–967. [Google Scholar] [CrossRef]
- Bashmakov, Y.K.; Petyaev, I.M.; Chalyk, N.E.; Klochkov, V.A.; Pristensky, D.V.; Chernyshova, M.P.; Kyle, N.H. Pharmacokinetics and oxidation parameters in volunteers supplemented with microencapsulated docosahexaenoic acid. Int. J. Appl. Basic Med. Res. 2018, 8, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Harpsøe, N.G.; Andersen, L.P.H.; Gögenur, I.; Rosenberg, J. Clinical pharmacokinetics of melatonin: A systematic review. Eur. J. Clin. Pharmacol. 2015, 71, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Candia, A.; Veliz, M.; Araya, C.; Quezada, S.; Ebensperger, G.; Serón-Ferré, M.; Reyes, R.V.; Llanos, A.J.; Herrera, E.A. Potential adverse effects of antenatal melatonin as a treatment for intrauterine growth restriction: Findings in pregnant sheep. Am. J. Obstet. Gynecol. 2016, 215, 245.e1–245.e7. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.B.; Jacob, S. A simple practice guide for dose conversion between animals and human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehnder, A.M.; Hawkins, M.G.; Trestrail, E.A.; Holt, R.W.; Kent, M.S. Calculation of body surface area via computed tomography–guided modeling in domestic rabbits (Oryctolagus cuniculus). Am. J. Vet. Res. 2012, 73, 1859–1863. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dose (mg/kg bw/day) | Number of Control Pups | Birth Weight [g] ± SEM | Number of IUGR Pups | Birth Weight [g] ± SEM |
|---|---|---|---|---|---|
| Rabbit Doe | Control Pups | IUGR Pups | |||
| w/o | - | 12 | 48.52 ± 1.93 | 10 | 31.72 ± 2.17 * |
| DHA | 37 | 2 | 57.05 ± 3.90 | 2 | 34.76 ± 5.20 * |
| MEL | 10 | 2 | 52.01 ± 9.38 | 2 | 27.94 ± 2.52 * |
| LF | 166 | 2 | 59.72 ± 1.57 | 2 | 37.86 ± 3.71 * |
| Compound (Synonym) | CAS Number | Solubility | Concentration In Vitro | MTC |
|---|---|---|---|---|
| DHA | 6217-54-5 | 300 µM (DMSO) | 300–100–30–10–3–1–0.3 µM | 10 µM |
| MEL | 73-31-4 | 100 µM (DMSO) | 100–30–10–3–1–0.3–0.1 µM | 3 µM |
| T3 | 55-06-1 | 30 nM (HCl/EtOH) | 30–10–3–1–0.3–0.1–0.03 nM | 30 nM |
| Zinc | 7440-66-6 | 300 µM (H2O) | 300–100–30–10–3–1–0.3 µM | 100 µM |
| LF | 339615-76-8 | 10 mg/mL (H2O) | 30–10–3–1–0.3–0.1–0.03 µM | 30 µM |
| SA | 131-48-6 | 30 µM (DMSO) | 30–10–3–1–0.3–0.1–0.03 µM | 30 µM |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kühne, B.A.; Vázquez-Aristizabal, P.; Fuentes-Amell, M.; Pla, L.; Loreiro, C.; Gómez-Catalán, J.; Gratacós, E.; Illa, M.; Barenys, M. Docosahexaenoic Acid and Melatonin Prevent Impaired Oligodendrogenesis Induced by Intrauterine Growth Restriction (IUGR). Biomedicines 2022, 10, 1205. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051205
Kühne BA, Vázquez-Aristizabal P, Fuentes-Amell M, Pla L, Loreiro C, Gómez-Catalán J, Gratacós E, Illa M, Barenys M. Docosahexaenoic Acid and Melatonin Prevent Impaired Oligodendrogenesis Induced by Intrauterine Growth Restriction (IUGR). Biomedicines. 2022; 10(5):1205. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051205
Chicago/Turabian StyleKühne, Britta Anna, Paula Vázquez-Aristizabal, Mercè Fuentes-Amell, Laura Pla, Carla Loreiro, Jesús Gómez-Catalán, Eduard Gratacós, Miriam Illa, and Marta Barenys. 2022. "Docosahexaenoic Acid and Melatonin Prevent Impaired Oligodendrogenesis Induced by Intrauterine Growth Restriction (IUGR)" Biomedicines 10, no. 5: 1205. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines10051205