Gut Microbiota and Colorectal Cancer Development: A Closer Look to the Adenoma-Carcinoma Sequence





Department of General Surgery and Medical-Surgical Specialties, University of Catania, Via S. Sofia 78, 95123 Catania, Italy
*
Author to whom correspondence should be addressed.
Biomedicines 2020, 8(11), 489; https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8110489
Submission received: 26 October 2020
/
Revised: 6 November 2020
/
Accepted: 8 November 2020
/
Published: 10 November 2020
(This article belongs to the Special Issue Colorectal Cancer: From Pathophysiology to Novel Therapeutic Approaches)
Abstract
:There is wide evidence that CRC could be prevented by regular physical activity, keeping a healthy body weight, and following a healthy and balanced diet. Many sporadic CRCs develop via the traditional adenoma-carcinoma pathway, starting as premalignant lesions represented by conventional, tubular or tubulovillous adenomas. The gut bacteria play a crucial role in regulating the host metabolism and also contribute to preserve intestinal barrier function and an effective immune response against pathogen colonization. The microbiota composition is different among people, and is conditioned by many environmental factors, such as diet, chemical exposure, and the use of antibiotic or other medication. The gut microbiota could be directly involved in the development of colorectal adenomas and the subsequent progression to CRC. Specific gut bacteria, such as Fusobacterium nucleatum, Escherichia coli, and enterotoxigenic Bacteroides fragilis, could be involved in colorectal carcinogenesis. Potential mechanisms of CRC progression may include DNA damage, promotion of chronic inflammation, and release of bioactive carcinogenic metabolites. The aim of this review was to summarize the current knowledge on the role of the gut microbiota in the development of CRC, and discuss major mechanisms of microbiota-related progression of the adenoma-carcinoma sequence.
1. Introduction
Colorectal cancer (CRC) is a leading cause of cancer mortality worldwide with approximately 900,000 deaths every year, and the increasing age-standardized incidence rate of CRC in most countries represents an important public health challenge [1]. Indeed, the global incidence of CRC was 1.8 million (95% UI 1.8–1.9) in 2017, with an age-standardized incidence rate of 23.2 per 100,000 person-years that raised by 9.5% (4.5–13.5) between 1990 and 2017 [2]. There is wide evidence that CRC risk is highly modifiable through diet and lifestyle [3]. Several studies suggested that a significant number of CRC cases could be prevented by regular physical activity, keeping a healthy body weight, and following a healthy and balanced diet [4,5,6].
Around 60–90% of sporadic CRCs arise via the traditional adenoma-carcinoma pathway, starting as premalignant lesions represented by conventional, tubular, or tubulovillous adenomas [7]. Cancers that derive from this pathway are frequently associated with male sex, and located in the distal colon. These tumors are characterized by chromosomal instability (CIN), inactivating mutations or losses in the adenomatous polyposis coli (APC) tumor suppressor gene, and in some cases mutations in the KRAS oncogene, SMAD4, PIK3CA, and TP53 genes [8,9].
The term “gut microbiota” indicates the collection of microorganisms (bacteria, archaea and eukarya) colonizing the human gastrointestinal tract. Overall, the number of these microorganisms has been calculated to exceed 1014, with a ratio of human:bacterial cells closer to 1:1 [10,11]. The gut bacteria play a crucial role in regulating host metabolism (i.e., absorption of indigestible carbohydrates and fat-soluble vitamins, and stimulation of innate and cell-mediated immunity) and also contribute to preserve intestinal barrier function and an effective immune response against pathogen colonization [12,13,14]. The microbiota composition is different among people, and is conditioned by many environmental factors, such as diet, chemical exposure, and the use of antibiotic or other medication [15].
Several studies suggested that the gut microbiota could be directly involved in the development of colorectal adenomas and the subsequent progression to CRC [16]. Patients with CRC could present changes in microbial composition and ecology, and functional studies in animal models underlined the importance of certain bacteria, such as Fusobacterium nucleatum, Escherichia coli, and Bacteroides fragilis, in colorectal carcinogenesis [17,18]. Possible mechanisms of CRC progression may include DNA damage, promotion of chronic inflammation, and release of bioactive carcinogenic metabolites [19,20,21].
The aim of this review was to summarize the current knowledge on the role of the gut microbiota in the development of CRC, including major mechanisms of microbiota-related progression of the adenoma-carcinoma sequence.
2. Risk Factors for the Development of Adenomas and CRC
Genetic alterations play a key role in the progression of adenomas to CRC; for instance, mutations may occur in oncogenes (i.e., KRAS), tumor suppressor genes such as APC, p53, and CTNNB1, as well as in pathways associated with CpG island methylation (CIMP), mismatch repair (MMR), and chromosomal and microsatellite instability (CIN and MSI) [22,23,24]. Ageing and family history have been also correlated with higher risk of adenomas and CRC [25,26,27].
It has been suggested that genetic predisposition and somatic mutations in combination with environmental factors could be responsible for CRC, in the way of a complex disease [28,29,30]. Lifestyle and dietary habits represent the most common environmental factors associated with colorectal adenomas and CRC [31,32,33]. Even if it is difficult to analyze the single dietary risk factors in epidemiological studies, preclinical animal models have shown the key role of nutrition in tumor development [34,35]. Nutrition may affect the incidence, natural progression and therapeutic response of cancer, modulating the release of endocrine factors, modifying inflammatory and immunological pathways, or by changing the gut microbiota composition [36,37,38].
An increased risk of adenomas and CRC has been observed in subjects consuming diets high in red meat or processed meat, food with a high glycemic index, salt and alcohol, and low daily water and fiber intake [39,40]. On the contrary, the consumption of white meat, vegetables and fish oils with a high omega-3 polyunsaturated fatty acids (PUFA) to omega-6 PUFA ratio could lower the risk of CRC [41,42,43]. A diet rich in fiber, vitamin B6, C, D, E, folic acid, magnesium and selenium, has also been suggested to decrease the risk of CRC [44]. Other risk factors that may contribute to the development of CRC are obesity, smoking, male sex, non-hispanic black ethnicity, and lack of physical activity [45,46,47].
There is growing evidence that diet may select for the microbiota composition, thus regulating many beneficial or harmful effects of gut bacteria [15,48]. For instance, dietary fiber are able to stimulate the colonic microbial production of anti-proliferative and counter carcinogenic substances, especially butyrate [49]. The adoption of a healthy lifestyle, and a diet rich in fiber, vegetables and fruit, could decrease the risk of CRC. Moreover, a recent study showed that higher fiber intake after the diagnosis of non-metastatic CRC (non-mCRC) was associated with decreased CRC-specific and overall mortality. Indeed, an increased fiber intake after CRC diagnosis could give supplementary advantages to patients with CRC due to the interaction with gut microbiota [50,51].
3. Dysbiosis, Inflammation and Toxic Bacterial Metabolites
The adenomas are the most frequent premalignant precursor lesions of almost all the sporadic CRCs [52]. Up to 40% of individuals aged 60 years or older may present adenomatous polyps, with a transformation rate into CRC of approximately 0.25% per year [53,54]. Inactivating mutations of the APC gene are considered as the initial step of the adenoma-carcinoma sequence. A loss of APC gene activity results in the accumulation of β-catenin, that leads to abnormal cell proliferation, and formation of adenomatous polyposis [55]. There is evidence that an interaction between gut microbiota and genetic could contribute to the genetic pattern of the adenoma-carcinoma sequence; indeed, bacterial drivers could be responsible for the initiation of precancerous lesions and the subsequent accumulation of gene mutations [56,57,58].
Chronic inflammation has also been suggested to play a crucial role in many aspects of CRC initiation, promotion, and progression [59,60]. A meta-analysis confirmed the association between circulating levels of C-reactive protein (CRP), a non-specific marker of systemic inflammation, and risk of colorectal adenoma [61]. Also, higher levels of pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), have been observed within adenoma tissues as an expression of an inflammatory state. TNF-α and IL-6 are also involved in cell growth, differentiation, and apoptosis [62,63].
At the phyla level, the colonic microbiota of healthy individuals usually shows a predominance of Gram-positive Firmicutes and Gram-negative Bacteroidetes, with a less presence of Verrucomicrobia and Actinobacteria. The Firmicutes phylum is represented by more than 200 different genera including Clostridium, Lactobacillus, Enterococcus, Bacillus, and Ruminicoccus. The Actinobacteria phylum mainly consists of the Bifidobacterium genus [64,65]. Variation in the composition of gut microbiota between phenotypically similar and healthy subjects may be influenced by age, gender, genetics, diet and diseases [66].
Some studied reported abnormalities in the normal bacterial community composition, known as dysbiosis, in CRC patients [67]. Dysbiosis of the gut microbiota is characterized by the reduction in commensal bacterial species (i.e., butyrate-producing bacteria) and the growth of detrimental bacterial strains (i.e., pro-inflammatory opportunistic pathogens) [68].
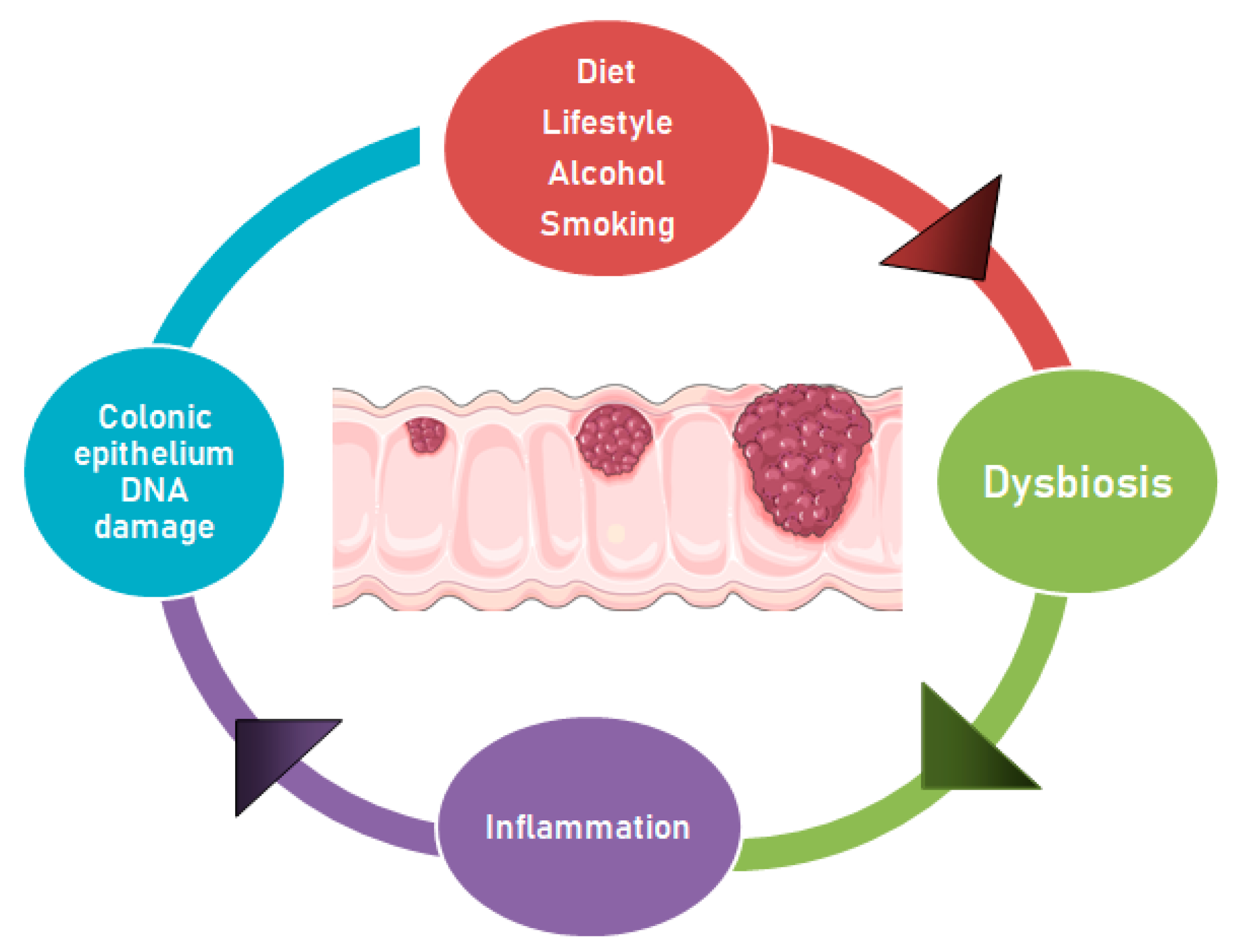
Changes in the balance of commensal bacteria may lead to a raise in mucosal permeability, bacterial translocation, and activation of factors of the innate and adaptive immune system to stimulate chronic inflammation [69]. Over-expression of proinflammatory cytokines, such as IL-12, IL-23, IFNγ and TNF-α by dendritic cells, macrophages, and natural killer (NK) cells, may further promote the activation of T and B cells and different inflammatory mediators. The activation of signaling pathways by transcription factors such as NF-κB and signal transducer and activator of transcription 3 (STAT3) in colonic epithelial cells, the production of reactive oxygen species (ROS) and the related oxidative stress, DNA damage, and abnormal cell proliferation, may favor the development of colorectal adenomas and cancer [70,71,72] (Figure 1).
During chronic inflammation, there is a general imbalance in the gut due to release of toxic compounds and procarcinogens. Actually, an abnormal generation of bacterial metabolites directly involved in tumor metabolism, such as polyamines and short-chain fatty acids (i.e., butyrate, propionate and acetate), has been observed in patients with adenomas and CRC [15,73]. Under homeostasis, the gut microbiota is metabolized to generate many beneficial compounds for the host, whereas under an unbalanced state, the bacterial growth and health of the host may be negatively influenced [74].
The microbiota initiates and supports the hypoxic environment of the gut that is fundamental for nutrient absorption, epithelial barrier function, and immune response. The response to hypoxia is regulated by hypoxia-inducible factors (HIFs), which modulate the expression of genes, including the ones involved in metabolism, that promote adaptation to hypoxia. Chronic HIF activation may aggravate disease conditions, leading to intestinal damage, inflammation, and CRC [75,76,77].
Overall, the fermentation of carbohydrates produces short-chain fatty acids, especially butyrate, which can be utilized by the host and shows antineoplastic properties, while proteolytic fermentation generates ammonia, sulphides, phenols, and cresols, which may exert a pro-inflammatory effect, increase tissue permeability and in turn contribute to the development of adenomas and CRC [78,79]. Great amounts of specific strains of bacteria may lead to the generation of other substances with anti- and/or pro-carcinogenic effects, such as enterotoxins, B vitamins, urolithins, cyclomodulins, lignans, and equol [16,80].
Changes of the microbiota profile in adenomas could enhance the production of primary and secondary bile acids, as well as sucrose, lipid, starch, and phenylpropanoid metabolism, thus supporting an intestinal environment that favors the growth of bile-resistant and sulfidogenic microorganisms including Desulfovibrio and Bilophilia [81,82].
It is well recognized that hydrogen sulfide (H2S) generated by bacteria in the gut is related to adenoma development and eventually CRC [83]. Many anaerobic bacterial strains such as Salmonella enterica, Clostridia, Escherichia coli, and Enterobacter aerogenes are able to convert cysteine to H2S, ammonia and pyruvate by cysteine desulfhydrase; moreover some gut bacteria (i.e., Escherichia coli, Salmonella, Enterobacter, Staphylococcus, Bacillus, Klebsiella, Corynebacterium, and Rhodococcus) may generate H2S by sulfite reduction [84]. H2S modulates inflammation, ischemia and/or perfusion injury and motility, and exerts a toxic activity on the colonic epithelium [85]. Phenolic substances such as amines, N-nitroso compounds (NOCs) found in processed meat, may also exert toxic activities favoring carcinogenesis [86,87].
Colibactin is a genotoxin produced by certain strains of bacteria, such as B2 phylogroup E. coli strains that colonize the human gut [88]. The synthesis of colibactin by the polyketide synthetase (pks) genomic island, especially in members of the family Enterobacteriaceae, may lead to chromosomal instability and DNA damage in eukaryotic cells, apoptosis of immune cells, and in turn the development of CRC [89].
4. Specific Bacteria Associated with Colorectal Adenoma and Cancer Development
Numerous studies have identified tumour-specific bacteria present in colorectal mucosal and/or faecal samples, and not detectable in healthy controls or tumour tissue versus the bordering healthy mucosa [90] (Table 1). A metagenome-wide association study (MGWAS) on stools from advanced adenoma and CRC patients and from healthy individuals, detected microbial genes, strains and functions enriched in each group. High consumption of red meat relative to fruits and vegetables seems to be associated with development of specific bacteria that could contribute to a more hostile intestinal milieu [91]. In general, microbial species associated with CRC development are represented by specific strains of Escherichia coli, Streptococcus gallolyticus, Bacteroides fragilis, Fusobacterium nucleatum, and Enterococcus faecalis among others [16].
Hale et al. observed significant abundances of multiple taxa in subjects with adenomas, such as Bilophila, Desulfovibrio, pro-inflammatory bacteria in the genus Mogibacterium, and Bacteroidetes spp. On the other hand, Veillonella, Firmicutes (class Clostridia), and Actinobacteria (family Bifidobacteriales) were more represented in patients without adenomas [81].
A study by Peters et al. analyzed for the first time the link between the gut microbiota and specific colorectal polyp types in 540 subjects, and showed that conventional adenomas (CA) cases had lower species diversity in faeces compared to controls (p = 0.03), especially with regard to advanced CA cases (p = 0.004). Only subjects with distal or advanced CA showed significant differences in general microbiota composition compared to controls (p = 0.02 and p = 0.002). Faeces of CA cases were characterized by the reduction in Clostridia from families Ruminococcaceae, Clostridiaceae, and Lachnospiraceae, and the increase in the classes Gammaproteobacteria and Bacilli, order Enterobacteriales, and genera Streptococcus and Actinomyces. There were not significant differences between sessile serrated adenoma (SSA) and hyperplastic polyps (HP) cases in diversity or composition compared to controls [92].
Feng et al. detected a great amount of Bacteroides and Parabacteroides, together with Bilophila wadsworthia, Lachnospiraceae bacterium, Alistipes putredinis, and Escherichia coli in CRC compared with both healthy and advanced adenoma. Also, gut commensals such as Bifidobactium animalis and Streptococcus thermophilus, were diminished in stools from adenoma or CRC patients, thus highlighting a divergence from healthy microbiota. Patients with advanced adenoma or CRC seem to be lacking in lactic acid-producing commensals such as Bifidobacterium that could facilitate epithelium regeneration and inhibition of opportunistic pathogens [91].
4.1. Fusobacterium nucleatum
F. nucleatum is an oral symbiont, and opportunistic pathogen that has been detected in intestinal cancers [93,94]. F. nucleatum may enhance CRC carcinogensis by stimulating the production of interleukin (IL)-17F/21/22/23/31/cluster of differentiation (CD)40L and protein expression of phospho-STAT3 (p-STAT3), p-STAT5, and phospho-extracellular regulated protein kinases (p-ERK)1/2 [95]. A great amount of Fusobacteria has been observed in SSA [108,109]; a study by Yu et al. reported that the prevalence of invasive Fusobacteria within proximal SSAs (78.8%) and HPs (65.7%) was significantly more elevated than that of proximal and distal traditional adenomas (28.9% and 24.4% respectively; p < 0.05) [96]. The presence of F. nucleatum has been associated with poor prognosis in CRC patients and development of chemoresistance [97,98]. F. nucleatum binds E-cadherin in the clonic epithelium and stimulates colorectal carcinogenesis through the fusobacterial adhesin FadA [110,111]. The interplay between Gal-GalNAc, a host polysaccharide, with fusobacterial lectin (Fap2) may promote the increase of F. nucleatum in colorectal adenoma and cancer [112]. A study by Mima et al. showed that multivariable hazard ratios (HRs) for CRC-specific mortality in F. nucleatum-low subjects and F. nucleatum-high subjects, compared with F. nucleatum-negative subjects, were 1.25 (95% C.I. 0.82 to 1.92) and 1.58 (95% C.I. 1.04 to 2.39), respectively (p for trend = 0.020). The quantity of F. nucleatum was correlated with microsatellite instability (MSI)-high (multivariable odd ratio (OR), 5.22; 95% CI 2.86 to 9.55) independent of the presence of CIMP and BRAF mutation. A significant association between CIMP and BRAF mutation with F. nucleatum was observed only in univariate analyses (p < 0.001) but not in multivariate analysis that adjusted for MSI status [97].
Yang et al. observed that an infection of CRC cells lines (HCT116, HT29, LoVo, and SW480) with F. nucleatum increased cell growth, invasiveness, and capability to form xenograft cancers in mice. F. nucleatum promoted Toll-like receptor 4 (TLR4) signaling to myeloid differentiation factor 88 (MYD88), activating NFκB signaling pathways and increasing the expression of microRNA-21 (miR21), which reduced the levels of the RAS GTPase p21 protein activator 1 (RASA1). Shorter survival times were observed for tumors with high amounts of F. nucleatum DNA and miR21 [113].
It has been also observed that F. nucleatum may promote LC3-II protein expression, autophagy pathway, and autophagosome production in CRC cells. F. nucleatum may favor the release of the autophagy-related proteins, pULK1, ULK1, and ATG7, contributing to the resistance to oxaliplatin and 5-fluorouracil regimens in CRC cells [98].
A study by Bullman et al. showed the persistance of F. nucleatum also in distal metastatic lesions of CRC patients. Administration of metronidazole in mice bearing a colon cancer xenograft decreased F. nucleatum load, tumor cell proliferation, and overall cancer development, thus suggesting that specific antibiotics could potentially be used to treat patients with Fusobacterium-associated CRC [114].
4.2. Streptococcus gallolyticus (Formerly S. bovis)
Streptococcus gallolyticus subsp. gallolyticus (SGG), formerly known as S. bovis biotype I, represents a common causative agent for bacteremia and endocarditis in older adults. Gut colonization by SGG is strongly correlated with the development of CRC [99,115]. Indeed, both American and European guidelines recommended colonoscopy in patients with SGG bacteremia [116,117].
A case-control study by Corredoira-Sánchez et al. carried out on 109 cases showed that the prevalence of CRC was higher in patients with SGG bacteremia compared to controls (70% vs. 32%; OR, 5.1; 95% CI 3.0–8.6). The study did not show significant differences when comparing nonadvanced adenomas (19% vs. 12%). However, significant differences were observed in advanced adenomas (40% vs. 16%; OR 3.5, 95% C.I. 2.0–6.1) and invasive CRC (12% vs. 5%, OR 2.9, 95% C.I. 1.2–6.9) [100].
A large epidemiological study by Butt et al. showed for the first time a statistically significant association between exposure to SGG antigens and CRC, and pointed out that the risk for CRC was stronger among subjects younger than 65 years [101].
Aymeric et al. observed that CRC-specific conditions may favor SGG colonization of the gut at the expense of commensal enterococci. Indeed, gut colonization by SGG is promoted by a bacteriocin called “gallocin”, which is enhanced by bile acids and may exert toxic activity to enterococci. Also, the stimulation of the Wnt pathway, and the reduced expression of the bile acid apical transporter gene Slc10A2, may act on the APC founding mutation, supporting the gut colonization by SGG [115].
4.3. Enterotoxigenic Bacteroides fragilis (ETBF)
Enterotoxigenic B. fragilis (ETBF) may support colorectal carcinogenesis by the production of pro-inflammatory cytokines and the stimulation of Wnt signaling. Expression of B. fragilis toxin (BFT), a 20 kDa metalloprotease produced by ETBF, is able to promote persistent colitis in mice, damage E-cadherin junctions, as well as stimulate B-catenin signaling and IL-8 production in colonic epithelial cells [118].
A study by Purcell et al. underlined the key role of ETBF in the development of colorectal low-grade dysplasia, tubular adenomas, and serrated polyps (p-values of 0.007, 0.027 and 0.007, respectively) [102]. Similar findings were reported in a study of patients with colonic adenomas that presented higher expression of the B. fragilis toxin gene (bft) associated with adenoma tissue compared to normal healthy mucosa [103].
Zamani et al. reported an increased positivity of ETBF in patients with precancerous and cancerous lesions compared to healthy controls. Higher ORs of ETBF were significantly associated with serrated lesions and adenoma with low-grade dysplasia. The most common subtype of bft gene was the bft1 gene, followed by the bft2 gene. An assessment of ETBF could represent a marker of CRC prognosis, especially in the precancerous lesions, and could be used for the screening of these conditions [104].
4.4. Enterococcus faecalis
E. faecalis is a Gram-positive commensal bacterium, that may be responsible for human disease through translocation from intestinal wall, oral cavity, and genito-urinary mucosa, leading to a systemic infection [119]. E. faecalis represents one of the most frequent causes of infection in older adults, and some studies underlined its importance for the development of cancer [120]. It has also been reported an association between enterococcal endocarditis and hidden CRC [119,121]. On the other hand, E. faecalis showed anti-inflammatory properties and probiotic activity, and is frequently administered in subjects with chronic sinusitis and bronchitis or in infant acute diarrhea [122].
Actually, there is no consensus on the role of E. faecalis in CRC: some studies highlighted its protective role or no role in CRC, whereas others reported potential pro-carcinogenic effects [123].
A study by Viljoen et al. carried out on 55 patients, did not highlight any significant clinical association between E. faecalis and CRC. However, the same study showed a relevant association bewteen clinicopathological features of CRC and Fusobacterium spp. and ETBF [105]. Miyamoto et al. observed that heat-killed E. faecalis strain EC-12 could suppress intestinal polyp development in Apc mutant Min mice. Administration of heat-killed EC-12 reduced the levels of c-Myc and cyclin D1 mRNA expression in intestinal polyps, by blocking the transcriptional activity of the T-cell factor/lymphoid enhancer factor [124].
E. faecalis could play a role in inducing CRC by activation of Wnt/β-catenin signaling and induction of pluripotent transcription factors linked to dedifferentiation. Indeed, exposure of murine primary colon epithelial cells to E. faecalis-infected macrophages contributed to CRC initiation through gene mutation, chromosomal instability, and endogenous cell transformation, which involved the transcription factors c-Myc, Klf4, Oct4, and Sox2i [125].
4.5. Escherichia coli
Classification of the Gram negative bacterium E. coli includes 8 phylogenetic groups (A, B1, B2, C, D, E, F and clade I). Commensal strains are commonly represented by A and B1 groups, being the largest part of the fecal flora of healthy individuals. Extraintestinal pathogenic strains (ExPEC) include mainly B2 and D groups, and may be responsible for many extraintestinal infections, due to the achievement of numerous virulence factors that potentially support the colonization of extraintestinal tissues [128]. However, both commensals and ExPEC are considered as a part of the normal gut microbiota in healthy subjects [129].
There is evidence that E. coli could play a role in the development of CRC [106,130]. Indeed, some patients with CRC may show an excessive growth of E. coli strains, mainly B2, characterized by high expression of virulence genes, including those encoding toxins and effectors that may induce carcinogenesis, such as colibactin, cytolethal distending toxins, cytotoxic necrotizing factors, and cycle-inhibiting factor [131,132]. In vitro studies showed that colibactin could be involved in DNA alkylation on adenine residues, leading to double-strand breaks [133,134]. Pleguezuelos-Manzano et al. demonstrated that exposure to genotoxic pks + E. coli, could be responsible for specific mutational signature in human intestinal organoids; indeed, an identical mutational signature was observed in 5876 human cancer genomes from two independent study cohorts, mostly in CRC [135].
Ambrosi et al. analyzed 272 E. coli isolates from colonoscopy biopsies, and showed that E. coli strains colonizing adenomatous polyps were characterized by specific phenotypes compared to those from normal mucosa, which included lack of motility, moderate to strong biofilm forming activity, and poor proteolytic capability [106].
In a study by Iyadorai et al. pks + E. coli was detected more frequently in CRC patients compared to healthy subjects. In vitro assays carried out on primary colon epithelial (PCE) and CRC (HCT116) cell lines, highlighted that the cytopathic effect of pks + E. coli strains could support the initiation and development of CRC [107].
5. Future Perspectives
Modulation of the gut microbiota, aiming to reverse microbial dysbiosis, could represent a new tool for prevention and treatment of CRC. The strategies could include the use of probiotics, prebiotics, postbiotics, antibiotics, and fecal microbiota transplantation (FMT) [136,137,138,139].
Overall, the effects of microbiota modulation on CRC prevention could be due to many mechanisms, such as the suppression of inflammatory state, stimulation of apoptosis of early cancer cells, re-establishment of intestinal barrier function and correction of microbiota composition [140,141]. Also, manipulation of the gut microbiota could alleviate chemotherapy-induced side effects, such as mucositis, as confirmed by a decreased incidence of diarrhea and weight loss after the administration of several probiotics strains in animal models [142,143].
There is growing evidence that modifications of microbial abundances in some pathological conditions could affect their co-abundance interactions; indeed, Chen et al. observed specific gut microbial co-abundance networks in patients with inflammatory bowel disease (IBD) and obesity. These findings underlined the importance of microbial dysbiosis in the pathogenesis of some diseases, and suggested that even the development of CRC could share similar mechanisms [144,145,146].
Promising preclinical studies suggested that modulation of gut microbiota could increase therapeutic efficacy of anticancer drugs. There is evidence that the administration of antibiotics could lead to clinical benefits to CRC patients by gut microbiota depletion and subsequent reduction of chemotherapeutic resistance. Indeed, a study by Geller et al. observed that intratumor bacteria could favor gemcitabine resistance through enzymatic inactivation, and therefore the administration of a gemcitabine-ciprofloxacin combination therapy could enhance the efficacy of chemotherapy [147].
Some studies demonstrated that the gut microbiota is also able to affect chemotherapy and/or immunotherapy efficacy by modulating immune response [148]. Oral administration of some probiotics, such as Bifidobacterium spp. and Akkermansia muciniphila, or FMT from treatment-responsive patients, stimulated the programmed cell death protein 1 ligand 1 (PD-L1)-based immunotherapy, thus blocking cancer development through the increase of dendritic cell and T cell response [149,150,151].
There is growing evidence that microbial shift markers could be used succesfully for non-invasive early diagnosis and/or prognostic assessment of CRC and advanced adenomas [81,152]. Mangifesta et al. performed a metataxonomic analysis based on 16S rRNA gene sequencing approach, and showed that some microbial taxa such as Bacteroides, Faecalibacterium, and Romboutsia, seem to be reduced in cancerogenic mucosa and in adenomatous polyps, thus representing potential new biomarkers of early carcinogenesis. Furthermore, the detection of high amounts of F. nucleatum in polyps, underlined the key role of this microorganism as a microbial biomarker for early diagnosis of CRC [153].
A study by Hale et al. showed that the composition of the gut microbiota in subjects with adenomas is significantly different from that of healthy subjects, and is similar to the microbiota of subjects with CRC. These changes could be a consequence of the Western diet and could result in metabolic changes leading to intestinal cellular damage and mutagenesis [81,154].
The combined assessment of heterogeneous CRC cohorts detected reproducible microbiota biomarkers and disease-predictive models that could represent useful tools for clinical prognostic tests and future research. A meta-analysis of 969 stool metagenomes carried out using data from five open access datasets and two new cohorts, showed that the gut microbiota in CRC was characterized by more richness than controls (p < 0.01), partly due to the growth of some species originating from the oral cavity. The results also highlighted an association between gluconeogenesis, putrefaction and fermentation processes with CRC, while the starch and stachyose degradation were associated with controls. A significant association between microbiota choline metabolism and CRC was also observed (p = 0.001) [155]. Another meta-analysis of eight stool metagenomic studies of CRC (n = 768) from different geographical areas, reported a significant enrichment in a group of 29 species in CRC metagenomes (FDR < 1 × 10−5). An elevated production of secondary bile acids from CRC metagenomes, higher expression of mucin and protein catabolism genes and reduction of carbohydrates degradation genes were observed, thus underlying a metabolic relationship between gut microbiota in CRC and a diet rich in meat and fat [156].
A study by Poore et al. carried out on The Cancer Genome Atlas (TCGA) detected specific microbial signatures in blood and tissue of different types of tumors, including CRC, which were predictive for patients with stage Ia-IIc tumor and tumors without any genomic modifications as detected by cell-free tumor DNA assessment. These findings could pave the way to a novel type of microbial-based CRC diagnostics [157].
Currently, there is a great limitation in availability of mouse models to study the interaction between gut microbiota and CRC. Zeb2IEC-Tg/+ (intestinal epithelial cell-specific transgenic expression of the epithelial-to-mesenchymal transition regulator Zeb2) mice represented the first and only microbiota-dependent CRC mouse model available so far. Specific characteristics of Zeb2IEC-Tg/+ mice included the presence of gut dysbiosis, and the preventive effect on carcinogenesis through the microbiota reduction by broad-spectrum antibiotics or germ-free rederivation [158].
6. Conclusions
In conclusion, detecting key relationships between diet, gut microbiota, and metabolites involved in the adenoma-carcinoma sequence could provide important basis for personalized medicine aimed at preventing and managing CRC. Secondary bile acids, H2S, and other bacterial metabolites could exert genotoxic activities and should be kept into account when investigating the adenoma and carcinoma development. Nonetheless, further studies are needed to evaluate the effects of diet, lifestyle, or medications on the gut metabolic environment and the microbiota. Finally, the identification of global microbiota signatures specific for CRC represents a promising tool in CRC diagnosis and therapy.
Author Contributions
Conceptualization, M.V. and A.B.; writing—original draft preparation, M.V.; writing—review and editing, M.V., R.C., F.B., and A.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The images used in the figure and graphical abstract are distributed under Creative Commons License and can be freely available at the following link: https://smart.servier.com/.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; Global Burden of Disease Cancer Collaboration; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived with Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2017: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2019, 5, 1749–1768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safiri, S.; Sepanlou, S.G.; Ikuta, K.S.; Bisignano, C.; Salimzadeh, H.; Delavari, A.; Ansari, R.; Roshandel, G.; Merat, S.; Fitzmaurice, C.; et al. The global, regional, and national burden of colorectal cancer and its attributable risk factors in 195 countries and territories, 1990–2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2019, 4, 913–933. [Google Scholar] [CrossRef] [Green Version]
- Hou, N.; Huo, D.; Dignam, J.J. Prevention of colorectal cancer and dietary management. Chin. Clin. Oncol. 2013, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Hughes, L.A.E.; Simons, C.C.J.M.; Brandt, P.A.; van den Engeland, M.; van Weijenberg, M.P. Lifestyle, Diet, and Colorectal Cancer Risk According to (Epi)genetic Instability: Current Evidence and Future Directions of Molecular Pathological Epidemiology. Curr. Colorectal Cancer Rep. 2017, 13, 455. [Google Scholar] [CrossRef] [Green Version]
- Schlesinger, S.; Lieb, W.; Koch, M.; Fedirko, V.; Dahm, C.C.; Pischon, T.; Nöthlings, U.; Boeing, H.; Aleksandrova, K. Body weight gain and risk of colorectal cancer: A systematic review and meta-analysis of observational studies. Obes Rev. 2015, 16, 607–619. [Google Scholar] [CrossRef]
- Shahjehan, F.; Merchea, A.; Cochuyt, J.J.; Li, Z.; Colibaseanu, D.T.; Kasi, P.M. Body Mass Index and Long-Term Outcomes in Patients with Colorectal Cancer. Front. Oncol. 2018, 8. [Google Scholar] [CrossRef]
- Bae, J.M.; Kim, J.H.; Kang, G.H. Molecular Subtypes of Colorectal Cancer and Their Clinicopathologic Features, with an Emphasis on the Serrated Neoplasia Pathway. Arch. Pathol. Lab. Med. 2016, 140, 406–412. [Google Scholar] [CrossRef] [Green Version]
- Vacante, M.; Borzì, A.M.; Basile, F.; Biondi, A. Biomarkers in colorectal cancer: Current clinical utility and future perspectives. World J. Clin. Cases 2018, 6, 869–881. [Google Scholar] [CrossRef]
- Sievers, C.K.; Grady, W.M.; Halberg, R.B.; Pickhardt, P.J. New insights into the earliest stages of colorectal tumorigenesis. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 723–729. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut Microbiota: Role in Pathogen Colonization, Immune Responses and Inflammatory Disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, K.; Hosomi, K.; Sawane, K.; Kunisawa, J. Metabolism of Dietary and Microbial Vitamin B Family in the Regulation of Host Immunity. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Tang, H.; Chen, P.; Xie, H.; Tao, Y. Demystifying the manipulation of host immunity, metabolism, and extraintestinal tumors by the gut microbiome. Signal Transduct. Target. Ther. 2019, 4, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Alcoholado, L.; Ramos-Molina, B.; Otero, A.; Laborda-Illanes, A.; Ordóñez, R.; Medina, J.A.; Gómez-Millán, J.; Queipo-Ortuño, M.I. The Role of the Gut Microbiome in Colorectal Cancer Development and Therapy Response. Cancers 2020, 12, 1406. [Google Scholar] [CrossRef] [PubMed]
- Alhinai, E.A.; Walton, G.E.; Commane, D.M. The Role of the Gut Microbiota in Colorectal Cancer Causation. Int. J. Mol. Sci. 2019, 20, 5295. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.H.; Yu, J. Gut microbiota in colorectal cancer: Mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 690–704. [Google Scholar] [CrossRef]
- Sun, T.; Liu, S.; Zhou, Y.; Yao, Z.; Zhang, D.; Cao, S.; Wei, Z.; Tan, B.; Li, Y.; Lian, Z.; et al. Evolutionary biologic changes of gut microbiota in an ‘adenoma-carcinoma sequence’ mouse colorectal cancer model induced by 1, 2-Dimethylhydrazine. Oncotarget 2016, 8, 444–457. [Google Scholar] [CrossRef] [Green Version]
- Arends, M.J. Pathways of colorectal carcinogenesis. Appl. Immunohistochem. Mol. Morphol. 2013, 21, 97–102. [Google Scholar] [CrossRef]
- Kidane, D.; Chae, W.J.; Czochor, J.; Eckert, K.A.; Glazer, P.M.; Bothwell, A.L.M.; Sweasy, J.B. Interplay between DNA repair and inflammation, and the link to cancer. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 116–139. [Google Scholar] [CrossRef]
- Saus, E.; Iraola-Guzmán, S.; Willis, J.R.; Brunet-Vega, A.; Gabaldón, T. Microbiome and colorectal cancer: Roles in carcinogenesis and clinical potential. Mol. Aspects Med. 2019, 69, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, Y.; Guo, H.; Yu, J.; Chen, Y.; Liu, Y.; Lan, L.; Li, J.; Wang, H.; Zhang, H. Molecular Alterations in Circulating Cell-Free DNA in Patients with Colorectal Adenoma or Carcinoma. Cancer Manag. Res. 2020, 12, 5159–5167. [Google Scholar] [CrossRef] [PubMed]
- Al-Sohaily, S.; Biankin, A.; Leong, R.; Kohonen-Corish, M.; Warusavitarne, J. Molecular pathways in colorectal cancer. J. Gastroenterol. Hepatol. 2012, 27, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Koveitypour, Z.; Panahi, F.; Vakilian, M.; Peymani, M.; Seyed Forootan, F.; Nasr Esfahani, M.H.; Ghaedi, K. Signaling pathways involved in colorectal cancer progression. Cell Biosci. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valli, A.; Harris, A.L.; Kessler, B.M. Hypoxia metabolism in ageing. Aging 2015, 7, 465–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henrikson, N.B.; Webber, E.M.; Goddard, K.A.; Scrol, A.; Piper, M.; Williams, M.S.; Zallen, D.T.; Calonge, N.; Ganiats, T.G.; Janssens, A.C.J.W.; et al. Family history and the natural history of colorectal cancer: Systematic review. Genet. Med. 2015, 17, 702–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rametta, S.; Grosso, G.; Galvano, F.; Mistretta, A.; Marventano, S.; Nolfo, F.; Buscemi, S.; Gangi, S.; Basile, F.; Biondi, A. Social disparities, health risk behaviors, and cancer. BMC Surg. 2013, 13 (Suppl. 2), S17. [Google Scholar] [CrossRef] [Green Version]
- Valle, L. Genetic predisposition to colorectal cancer: Where we stand and future perspectives. World J. Gastroenterol. 2014, 20, 9828–9849. [Google Scholar] [CrossRef]
- Schubert, S.A.; Morreau, H.; de Miranda, N.F.C.C.; van Wezel, T. The missing heritability of familial colorectal cancer. Mutagenesis 2020, 35, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Esteban-Jurado, C.; Garre, P.; Vila, M.; Lozano, J.J.; Pristoupilova, A.; Beltrán, S.; Abulí, A.; Muñoz, J.; Balaguer, F.; Ocaña, T.; et al. New genes emerging for colorectal cancer predisposition. World J. Gastroenterol. 2014, 20, 1961–1971. [Google Scholar] [CrossRef]
- Pietrzyk, Ł. Food properties and dietary habits in colorectal cancer prevention and development. Int. J. Food Prop. 2017, 20, 2323–2343. [Google Scholar] [CrossRef] [Green Version]
- Quang, L.N.; Hien, N.Q.; Quang, N.T.; Chung, N.T. Active Lifestyle Patterns Reduce the Risk of Colorectal Cancer in the North of Vietnam: A Hospital-Based Case–Control Study. Cancer Control 2019, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolfo, F.; Rametta, S.; Marventano, S.; Grosso, G.; Mistretta, A.; Drago, F.; Gangi, S.; Basile, F.; Biondi, A. Pharmacological and dietary prevention for colorectal cancer. BMC Surg. 2013, 13, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, T.; Lyon, C.J.; Bergin, S.; Caligiuri, M.A.; Hsueh, W.A. Obesity, Inflammation, and Cancer. Annu. Rev. Pathol. 2016, 11, 421–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, G.; Buscemi, S.; Galvano, F.; Mistretta, A.; Marventano, S.; La Vela, V.; Drago, F.; Gangi, S.; Basile, F.; Biondi, A. Mediterranean diet and cancer: Epidemiological evidence and mechanism of selected aspects. BMC Surg. 2013, 13 (Suppl. 2), S14. [Google Scholar] [CrossRef] [Green Version]
- Zitvogel, L.; Pietrocola, F.; Kroemer, G. Nutrition, inflammation and cancer. Nat. Immunol. 2017, 18, 843–850. [Google Scholar] [CrossRef]
- De Almeida, C.V.; de Camargo, M.R.; Russo, E.; Amedei, A. Role of diet and gut microbiota on colorectal cancer immunomodulation. World J. Gastroenterol. 2019, 25, 151–162. [Google Scholar] [CrossRef]
- Grosso, G.; Biondi, A.; Galvano, F.; Mistretta, A.; Marventano, S.; Buscemi, S.; Drago, F.; Basile, F. Factors associated with colorectal cancer in the context of the mediterranean diet: A case-control study. Nutr. Cancer 2014, 66, 558–565. [Google Scholar] [CrossRef]
- Turner, N.D.; Lloyd, S.K. Association between red meat consumption and colon cancer: A systematic review of experimental results. Exp. Biol. Med. 2017, 242, 813–839. [Google Scholar] [CrossRef] [Green Version]
- Fung, T.T.; Brown, L.S. Dietary Patterns and the Risk of Colorectal Cancer. Curr. Nutr. Rep. 2013, 2, 48–55. [Google Scholar] [CrossRef]
- Kantor, E.D.; Lampe, J.W.; Peters, U.; Vaughan, T.L.; White, E. Long-chain omega-3 polyunsaturated fatty acid intake and risk of colorectal cancer. Nutr. Cancer 2014, 66, 716–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Volpato, M.; Hull, M.A. Omega-3 polyunsaturated fatty acids as adjuvant therapy of colorectal cancer. Cancer Metastasis Rev. 2018, 37, 545–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borzì, A.M.; Biondi, A.; Basile, F.; Luca, S.; Vicari, E.S.D.; Vacante, M. Olive oil effects on colorectal cancer. Nutrients 2019, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pericleous, M.; Mandair, D.; Caplin, M.E. Diet and supplements and their impact on colorectal cancer. J. Gastrointest. Oncol. 2013, 4, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Low, E.E.; Demb, J.; Liu, L.; Earles, A.; Bustamante, R.; Williams, C.D.; Provenzale, D.; Kaltenbach, T.; Gawron, A.J.; Martinez, M.E.; et al. Risk Factors for Early-Onset Colorectal Cancer. Gastroenterology 2020, 159, 492–501.e7. [Google Scholar] [CrossRef] [PubMed]
- Jackson, C.S.; Oman, M.; Patel, A.M.; Vega, K.J. Health disparities in colorectal cancer among racial and ethnic minorities in the United States. J. Gastrointest. Oncol. 2016, 7, S32–S43. [Google Scholar] [CrossRef] [PubMed]
- Demb, J.; Earles, A.; Martínez, M.E.; Bustamante, R.; Bryant, A.K.; Murphy, J.D.; Liu, L.; Gupta, S. Risk factors for colorectal cancer significantly vary by anatomic site. BMJ Open Gastroenterol. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Vipperla, K.; O’Keefe, S.J. Diet, microbiota, and dysbiosis: A ‘recipe’ for colorectal cancer. Food Funct. 2016, 7, 1731–1740. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wu, Y.; He, L.; Wu, L.; Wang, X.; Liu, Z. Effects of the intestinal microbial metabolite butyrate on the development of colorectal cancer. J. Cancer 2018, 9, 2510–2517. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Yu, J. The association of diet, gut microbiota and colorectal cancer: What we eat may imply what we get. Protein Cell 2018, 9, 474–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Wu, K.; Meyerhardt, J.A.; Ogino, S.; Wang, M.; Fuchs, C.S.; Giovannucci, E.L.; Chan, A.T. Fiber intake and survival after colorectal cancer diagnosis. JAMA Oncol. 2018, 4, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.F.; Ibrahim, A.E.K.; Arends, M.J. Molecular pathological classification of colorectal cancer. Virchows Arch. 2016, 469, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhiqiang, F.; Jie, C.; Yuqiang, N.; Chenghua, G.; Hong, W.; Zheng, S.; Wanglin, L.; Yongjian, Z.; Liping, D.; Lizhong, Z.; et al. Analysis of population-based colorectal cancer screening in Guangzhou, 2011–2015. Cancer Med. 2019, 8, 2496–2502. [Google Scholar] [CrossRef] [PubMed]
- Conteduca, V.; Sansonno, D.; Russi, S.; Dammacco, F. Precancerous colorectal lesions (Review). Int. J. Oncol. 2013, 43, 973–984. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Shay, J.W. Multiple Roles of APC and its Therapeutic Implications in Colorectal Cancer. J. Natl. Cancer Inst. 2017, 109. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Zhang, R.; Shu, R.; Yu, J.; Li, H.; Long, H.; Jin, S.; Li, S.; Hu, Q.; Yao, F.; et al. Study of the Relationship between Microbiome and Colorectal Cancer Susceptibility Using 16SrRNA Sequencing. Biomed. Res. Int. 2020, 2020, 7828392. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Gao, Z.; Huang, L.; Qin, H. Gut microbiota and colorectal cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 757–769. [Google Scholar] [CrossRef]
- Liang, S.; Mao, Y.; Liao, M.; Xu, Y.; Chen, Y.; Huang, X.; Wei, C.; Wu, C.; Wang, Q.; Pan, X.; et al. Gut microbiome associated with APC gene mutation in patients with intestinal adenomatous polyps. Int. J. Biol. Sci. 2020, 16, 135–146. [Google Scholar] [CrossRef]
- Piotrowski, I.; Kulcenty, K.; Suchorska, W. Interplay between inflammation and cancer. Rep. Pract. Oncol. Radiother. 2020, 25, 422–427. [Google Scholar] [CrossRef]
- Lucas, C.; Barnich, N.; Nguyen, H.T.T. Microbiota, Inflammation and Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 1310. [Google Scholar] [CrossRef] [Green Version]
- Godos, J.; Biondi, A.; Galvano, F.; Basile, F.; Sciacca, S.; Giovannucci, E.L.; Grosso, G. Markers of systemic inflammation and colorectal adenoma risk: Meta-analysis of observational studies. World J. Gastroenterol. 2017, 23, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zheng, S.G. Hall of Fame among Pro-inflammatory Cytokines: Interleukin-6 Gene and Its Transcriptional Regulation Mechanisms. Front. Immunol. 2016, 7, 604. [Google Scholar] [CrossRef] [PubMed]
- Comstock, S.S.; Xu, D.; Hortos, K.; Kovan, B.; McCaskey, S.; Pathak, D.R.; Fenton, J.I. Association of serum cytokines with colorectal polyp number and type in adult males. Eur. J. Cancer Prev. 2016, 25, 173–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Tuddenham, S.; Sears, C.L. The intestinal microbiome and health. Curr. Opin. Infect. Dis. 2015, 28, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Chen, J.; Zheng, J.; Hu, G.; Wang, J.; Huang, C.; Lou, L.; Wang, X.; Zeng, Y. Mucosal adherent bacterial dysbiosis in patients with colorectal adenomas. Sci. Rep. 2016, 6, 26337. [Google Scholar] [CrossRef]
- DeGruttola, A.K.; Low, D.; Mizoguchi, A.; Mizoguchi, E. Current understanding of dysbiosis in disease in human and animal models. Inflamm. Bowel. Dis. 2016, 22, 1137–1150. [Google Scholar] [CrossRef] [Green Version]
- Keku, T.O.; Dulal, S.; Deveaux, A.; Jovov, B.; Han, X. The gastrointestinal microbiota and colorectal cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 2015, 308, G351–G363. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Hnatyszyn, A.; Hryhorowicz, S.; Kaczmarek-Ryś, M.; Lis, E.; Słomski, R.; Scott, R.J.; Pławski, A. Colorectal carcinoma in the course of inflammatory bowel diseases. Hered. Cancer Clin. Pract. 2019, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.T.; Kantilal, H.K.; Davamani, F. The Mechanism of Bacteroides fragilis Toxin Contributes to Colon Cancer Formation. Malays. J. Med. Sci. 2020, 27, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Molina, B.; Queipo-Ortuño, M.I.; Lambertos, A.; Tinahones, F.J.; Peñafiel, R. Dietary and Gut Microbiota Polyamines in Obesity- and Age-Related Diseases. Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, A.; Miao, J.; Sun, H.; Yan, G.; Wu, F.; Wang, X. Gut microbiota as important modulator of metabolism in health and disease. RSC Adv. 2018, 8, 42380–42389. [Google Scholar] [CrossRef] [Green Version]
- Singhal, R.; Shah, Y.M. Oxygen battle in the gut: Hypoxia and hypoxia-inducible factors in metabolic and inflammatory responses in the intestine. J. Biol. Chem. 2020, 295, 10493–10505. [Google Scholar] [CrossRef]
- Valli, A.; Morotti, M.; Zois, C.E.; Albers, P.K.; Soga, T.; Feldinger, K.; Fischer, R.; Frejno, M.; McIntyre, A.; Bridges, E.; et al. Adaptation to HIF1α Deletion in Hypoxic Cancer Cells by Upregulation of GLUT14 and Creatine Metabolism. Mol. Cancer Res. 2019, 17, 1531–1544. [Google Scholar] [CrossRef] [Green Version]
- Valli, A.; Rodriguez, M.; Moutsianas, L.; Fischer, R.; Fedele, V.; Huang, H.-L.; Van Stiphout, R.; Jones, D.; Mccarthy, M.; Vinaxia, M.; et al. Hypoxia induces a lipogenic cancer cell phenotype via HIF1α-dependent and -independent pathways. Oncotarget 2014, 6, 1920–1941. [Google Scholar] [CrossRef] [Green Version]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet–Microbe–Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Baxter, N.T.; Schmidt, A.W.; Venkataraman, A.; Kim, K.S.; Waldron, C.; Schmidt, T.M. Dynamics of Human Gut Microbiota and Short-Chain Fatty Acids in Response to Dietary Interventions with Three Fermentable Fibers. mBio 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Gaya, P.; Medina, M.; Sánchez-Jiménez, A.; Landete, J.M. Phytoestrogen Metabolism by Adult Human Gut Microbiota. Molecules 2016, 21, 1034. [Google Scholar] [CrossRef] [Green Version]
- Hale, V.L.; Chen, J.; Johnson, S.; Harrington, S.C.; Yab, T.C.; Smyrk, T.C.; Nelson, H.; Boardman, L.A.; Druliner, B.R.; Levin, T.R.; et al. Shifts in the fecal microbiota associated with adenomatous polyps. Cancer Epidemiol. Biomark. Prev. 2017, 26, 85–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, Y.; Stams, A.J.M.; de Vos, W.M.; Sánchez-Andrea, I. Enrichment of sulfidogenic bacteria from the human intestinal tract. FEMS Microbiol. Lett. 2017, 364. [Google Scholar] [CrossRef] [PubMed]
- Guo, F.-F.; Yu, T.-C.; Hong, J.; Fang, J.-Y. Emerging Roles of Hydrogen Sulfide in Inflammatory and Neoplastic Colonic Diseases. Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomasova, L.; Konopelski, P.; Ufnal, M. Gut Bacteria and Hydrogen Sulfide: The New Old Players in Circulatory System Homeostasis. Molecules 2016, 21, 1558. [Google Scholar] [CrossRef]
- Singh, S.B.; Lin, H.C. Hydrogen Sulfide in Physiology and Diseases of the Digestive Tract. Microorganisms 2015, 3, 866–889. [Google Scholar] [CrossRef] [Green Version]
- Herrmann, S.S.; Duedahl-Olesen, L.; Christensen, T.; Olesen, P.T.; Granby, K. Dietary exposure to volatile and non-volatile N-nitrosamines from processed meat products in Denmark. Food Chem. Toxicol. 2015, 80, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Steinberg, P. Red Meat-Derived Nitroso Compounds, Lipid Peroxidation Products and Colorectal Cancer. Foods 2019, 8, 252. [Google Scholar] [CrossRef] [Green Version]
- Wernke, K.M.; Xue, M.; Tirla, A.; Kim, C.S.; Crawford, J.M.; Herzon, S.B. Structure and bioactivity of colibactin. Bioorg. Med. Chem. Lett. 2020, 30, 127280. [Google Scholar] [CrossRef]
- Faïs, T.; Delmas, J.; Barnich, N.; Bonnet, R.; Dalmasso, G. Colibactin: More Than a New Bacterial Toxin. Toxins 2018, 10, 151. [Google Scholar] [CrossRef] [Green Version]
- Kasai, C.; Sugimoto, K.; Moritani, I.; Tanaka, J.; Oya, Y.; Inoue, H.; Tameda, M.; Shiraki, K.; Ito, M.; Takei, Y.; et al. Comparison of human gut microbiota in control subjects and patients with colorectal carcinoma in adenoma: Terminal restriction fragment length polymorphism and next-generation sequencing analyses. Oncol. Rep. 2016, 35, 325–333. [Google Scholar] [CrossRef] [Green Version]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma-carcinoma sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, B.A.; Dominianni, C.; Shapiro, J.A.; Church, T.R.; Wu, J.; Miller, G.; Yuen, E.; Freiman, H.; Lustbader, I.; Salik, J.; et al. The gut microbiota in conventional and serrated precursors of colorectal cancer. Microbiome 2016, 4, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.-Y.; Ge, Q.-X.; Cao, J.; Zhou, Y.-J.; Du, Y.-L.; Shen, B.; Wan, Y.-J.Y.; Nie, Y.-Q. Association of Fusobacterium nucleatum infection with colorectal cancer in Chinese patients. World J. Gastroenterol. 2016, 22, 3227–3233. [Google Scholar] [CrossRef] [PubMed]
- Fukugaiti, M.H.; Ignacio, A.; Fernandes, M.R.; Ribeiro Júnior, U.; Nakano, V.; Avila-Campos, M.J. High occurrence of Fusobacterium nucleatum and Clostridium difficile in the intestinal microbiota of colorectal carcinoma patients. Braz. J. Microbiol. 2015, 46, 1135–1140. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.-N.; Yu, T.-C.; Zhao, H.-J.; Sun, T.-T.; Chen, H.-M.; Chen, H.-Y.; An, H.-F.; Weng, Y.-R.; Yu, J.; Li, M.; et al. Berberine may rescue Fusobacterium nucleatum-induced colorectal tumorigenesis by modulating the tumor microenvironment. Oncotarget 2015, 6, 32013–32026. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chen, Y.; Fu, X.; Zhou, X.; Peng, Y.; Shi, L.; Chen, T.; Wu, Y. Invasive Fusobacterium nucleatum may play a role in the carcinogenesis of proximal colon cancer through the serrated neoplasia pathway. Int. J. Cancer 2016, 139, 1318–1326. [Google Scholar] [CrossRef] [Green Version]
- Mima, K.; Nishihara, R.; Qian, Z.R.; Cao, Y.; Sukawa, Y.; Nowak, J.A.; Yang, J.; Dou, R.; Masugi, Y.; Song, M.; et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut 2016, 65, 1973–1980. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Guo, F.; Yu, Y.; Sun, T.; Ma, D.; Han, J.; Qian, Y.; Kryczek, I.; Sun, D.; Nagarsheth, N.; et al. Fusobacterium nucleatum Promotes Chemoresistance to Colorectal Cancer by Modulating Autophagy. Cell 2017, 170, 548–563.e16. [Google Scholar] [CrossRef] [Green Version]
- Little, D.H.W.; Onizuka, K.M.; Khan, K.J. Referral for Colonoscopy in Patients with Streptococcus bovis Bacteremia and the Association with Colorectal Cancer and Adenomatous Polyps: A Quality Assurance Study. Gastrointest. Disord. 2019, 1, 385–390. [Google Scholar] [CrossRef] [Green Version]
- Corredoira-Sánchez, J.; García-Garrote, F.; Rabuñal, R.; López-Roses, L.; García-País, M.J.; Castro, E.; González-Soler, R.; Coira, A.; Pita, J.; López-Álvarez, M.J.; et al. Association between bacteremia due to Streptococcus gallolyticus subsp. gallolyticus (Streptococcus bovis I) and colorectal neoplasia: A case-control study. Clin. Infect. Dis. 2012, 55, 491–496. [Google Scholar] [CrossRef]
- Butt, J.; Romero-Hernández, B.; Pérez-Gómez, B.; Willhauck-Fleckenstein, M.; Holzinger, D.; Martin, V.; Moreno, V.; Linares, C.; Dierssen-Sotos, T.; Barricarte, A.; et al. Association of Streptococcus gallolyticus subspecies gallolyticus with colorectal cancer: Serological evidence. Int. J. Cancer 2016, 138, 1670–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purcell, R.V.; Pearson, J.; Aitchison, A.; Dixon, L.; Frizelle, F.A.; Keenan, J.I. Colonization with enterotoxigenic Bacteroides fragilis is associated with early-stage colorectal neoplasia. PLoS ONE 2017, 12, e0171602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, L.L.; Wu, N.; Zhu, Y.M.; Qiu, X.Y.; Chen, G.D.; Zhang, L.M.; Liu, Y.L. [Expression of enterotoxigenic Bacteroides fragilis and polyketide synthase gene-expressing Escherichia coli in colorectal adenoma patients]. Zhonghua Yi Xue Za Zhi 2016, 96, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Zamani, S.; Taslimi, R.; Sarabi, A.; Jasemi, S.; Sechi, L.A.; Feizabadi, M.M. Enterotoxigenic Bacteroides fragilis: A Possible Etiological Candidate for Bacterially-Induced Colorectal Precancerous and Cancerous Lesions. Front. Cell Infect Microbiol. 2020, 9. [Google Scholar] [CrossRef] [PubMed]
- Viljoen, K.S.; Dakshinamurthy, A.; Goldberg, P.; Blackburn, J.M. Quantitative profiling of colorectal cancer-associated bacteria reveals associations between fusobacterium spp., enterotoxigenic Bacteroides fragilis (ETBF) and clinicopathological features of colorectal cancer. PLoS ONE 2015, 10, e0119462. [Google Scholar] [CrossRef] [Green Version]
- Ambrosi, C.; Sarshar, M.; Aprea, M.R.; Pompilio, A.; Di Bonaventura, G.; Strati, F.; Pronio, A.; Nicoletti, M.; Zagaglia, C.; Palamara, A.T.; et al. Colonic adenoma-associated Escherichia coli express specific phenotypes. Microbes Infect. 2019, 21, 305–312. [Google Scholar] [CrossRef]
- Iyadorai, T.; Mariappan, V.; Vellasamy, K.M.; Wanyiri, J.W.; Roslani, A.C.; Lee, G.K.; Sears, C.; Vadivelu, J. Prevalence and association of pks+ Escherichia coli with colorectal cancer in patients at the University Malaya Medical Centre, Malaysia. PLoS ONE 2020, 15, e0228217. [Google Scholar] [CrossRef]
- IJspeert, J.E.G.; Vermeulen, L.; Meijer, G.A.; Dekker, E. Serrated neoplasia-role in colorectal carcinogenesis and clinical implications. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 401–409. [Google Scholar] [CrossRef]
- Güven, D.C.; Dizdar, O. Fusobacterium and colorectal carcinogenesis. Carcinogenesis 2018, 39, 84. [Google Scholar] [CrossRef]
- Rubinstein, M.R.; Wang, X.; Liu, W.; Hao, Y.; Cai, G.; Han, Y.W. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host. Microbe 2013, 14, 195–206. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.-T.; Luo, H.-S.; Gao, F.; Tang, Q.-C.; Chen, W. Fusobacterium nucleatum promotes the progression of colorectal cancer by interacting with E-cadherin. Oncol. Lett. 2018, 16, 2606–2612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abed, J.; Emgård, J.E.M.; Zamir, G.; Faroja, M.; Almogy, G.; Grenov, A.; Sol, A.; Naor, R.; Pikarsky, E.; Atlan, K.A.; et al. Fap2 Mediates Fusobacterium nucleatum Colorectal Adenocarcinoma Enrichment by Binding to Tumor-Expressed Gal-GalNAc. Cell Host. Microbe 2016, 20, 215–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Weng, W.; Peng, J.; Hong, L.; Yang, L.; Toiyama, Y.; Gao, R.; Liu, M.; Yin, M.; Pan, C.; et al. Fusobacterium nucleatum Increases Proliferation of Colorectal Cancer Cells and Tumor Development in Mice by Activating Toll-Like Receptor 4 Signaling to Nuclear Factor-κB, and Up-regulating Expression of MicroRNA-21. Gastroenterology 2017, 152, 851–866.e24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bullman, S.; Pedamallu, C.S.; Sicinska, E.; Clancy, T.E.; Zhang, X.; Cai, D.; Neuberg, D.; Huang, K.; Guevara, F.; Nelson, T.; et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science 2017, 358, 1443–1448. [Google Scholar] [CrossRef] [Green Version]
- Aymeric, L.; Donnadieu, F.; Mulet, C.; du Merle, L.; Nigro, G.; Saffarian, A.; Bérard, M.; Poyart, C.; Robine, S.; Regnault, B.; et al. Colorectal cancer specific conditions promote Streptococcus gallolyticus gut colonization. Proc. Natl. Acad. Sci. USA 2018, 115, E283–E291. [Google Scholar] [CrossRef] [Green Version]
- Baddour, L.M.; Wilson, W.R.; Bayer, A.S.; Fowler, V.G.; Tleyjeh, I.M.; Rybak, M.J.; Barsic, B.; Lockhart, P.B.; Gewitz, M.H.; Levison, M.E.; et al. Infective Endocarditis in Adults: Diagnosis, Antimicrobial Therapy, and Management of Complications: A Scientific Statement for Healthcare Professionals from the American Heart Association. Circulation 2015, 132, 1435–1486. [Google Scholar] [CrossRef]
- Habib, G.; Lancellotti, P.; Antunes, M.J.; Bongiorni, M.G.; Casalta, J.-P.; Del Zotti, F.; Dulgheru, R.; El Khoury, G.; Erba, P.A.; Iung, B.; et al. 2015 ESC Guidelines for the management of infective endocarditis: The Task Force for the Management of Infective Endocarditis of the European Society of Cardiology (ESC). Endorsed by: European Association for Cardio-Thoracic Surgery (EACTS), the European Association of Nuclear Medicine (EANM). Eur. Heart J. 2015, 36, 3075–3128. [Google Scholar] [CrossRef]
- Allen, J.; Hao, S.; Sears, C.L.; Timp, W. Epigenetic Changes Induced by Bacteroides fragilis Toxin. Infect. Immun. 2019, 87. [Google Scholar] [CrossRef] [Green Version]
- Khan, Z.; Siddiqui, N.; Saif, M.W. Enterococcus Faecalis Infective Endocarditis and Colorectal Carcinoma: Case of New Association Gaining Ground. Gastroenterol. Res. 2018, 11, 238–240. [Google Scholar] [CrossRef] [Green Version]
- Goh, H.M.S.; Yong, M.H.A.; Chong, K.K.L.; Kline, K.A. Model systems for the study of Enterococcal colonization and infection. Virulence 2017, 8, 1525–1562. [Google Scholar] [CrossRef] [Green Version]
- Alozie, A.; Köller, K.; Pose, L.; Raftis, M.; Steinhoff, G.; Westphal, B.; Lamprecht, G.; Podbielski, A. Streptococcus bovis infectious endocarditis and occult gastrointestinal neoplasia: Experience with 25 consecutive patients treated surgically. Gut. Pathog. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Bai, T.; Zhang, L.; Qian, W.; Song, J.; Hou, X. Inhibition effect of Bifidobacterium longum, Lactobacillus acidophilus, Streptococcus thermophilus and Enterococcus faecalis and their related products on human colonic smooth muscle in vitro. PLoS ONE 2017, 12, e0189257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Almeida, C.V.; Taddei, A.; Amedei, A. The controversial role of Enterococcus faecalis in colorectal cancer. Ther. Adv. Gastroenterol. 2018, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, S.; Komiya, M.; Fujii, G.; Hamoya, T.; Nakanishi, R.; Fujimoto, K.; Tamura, S.; Kurokawa, Y.; Takahashi, M.; Ijichi, T.; et al. Preventive Effects of Heat-Killed Enterococcus faecalis Strain EC-12 on Mouse Intestinal Tumor Development. Int. J. Mol. Sci. 2017, 18, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Yang, Y.; Huycke, M.M. Commensal-infected macrophages induce dedifferentiation and reprogramming of epithelial cells during colorectal carcinogenesis. Oncotarget 2017, 8, 102176–102190. [Google Scholar] [CrossRef] [PubMed]
- Iebba, V.; Totino, V.; Gagliardi, A.; Santangelo, F.; Cacciotti, F.; Trancassini, M.; Mancini, C.; Cicerone, C.; Corazziari, E.; Pantanella, F.; et al. Eubiosis and dysbiosis: The two sides of the microbiota. New Microbiol. 2016, 39, 1–12. [Google Scholar]
- Flandroy, L.; Poutahidis, T.; Berg, G.; Clarke, G.; Dao, M.-C.; Decaestecker, E.; Furman, E.; Haahtela, T.; Massart, S.; Plovier, H.; et al. The impact of human activities and lifestyles on the interlinked microbiota and health of humans and of ecosystems. Sci. Total Environ. 2018, 627, 1018–1038. [Google Scholar] [CrossRef]
- Vila, J.; Sáez-López, E.; Johnson, J.R.; Römling, U.; Dobrindt, U.; Cantón, R.; Giske, C.G.; Naas, T.; Carattoli, A.; Martínez-Medina, M.; et al. Escherichia coli: An old friend with new tidings. FEMS Microbiol. Rev. 2016, 40, 437–463. [Google Scholar] [CrossRef] [Green Version]
- Poolman, J.T.; Wacker, M. Extraintestinal Pathogenic Escherichia coli, a Common Human Pathogen: Challenges for Vaccine Development and Progress in the Field. J. Infect. Dis. 2016, 213, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Gagnière, J.; Raisch, J.; Veziant, J.; Barnich, N.; Bonnet, R.; Buc, E.; Bringer, M.-A.; Pezet, D.; Bonnet, M. Gut microbiota imbalance and colorectal cancer. World J. Gastroenterol. 2016, 22, 501–518. [Google Scholar] [CrossRef]
- Khan, A.A.; Khan, Z.; Malik, A.; Kalam, M.A.; Cash, P.; Ashraf, M.T.; Alshamsan, A. Colorectal cancer-inflammatory bowel disease nexus and felony of Escherichia coli. Life Sci. 2017, 180, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Bleich, R.M.; Arthur, J.C. Revealing a microbial carcinogen. Science 2019, 363, 689–690. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.R.; Jiang, Y.; Villalta, P.W.; Stornetta, A.; Boudreau, P.D.; Carrá, A.; Brennan, C.A.; Chun, E.; Ngo, L.; Samson, L.D.; et al. The human gut bacterial genotoxin colibactin alkylates DNA. Science 2019, 363. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Kim, C.; Healy, A.; Wernke, K.; Wang, Z.; Frischling, M.; Shine, E.; Wang, W.; Herzon, S.; Crawford, J. Structure elucidation of colibactin and its DNA cross-links. Science 2019, 365. [Google Scholar] [CrossRef] [PubMed]
- Pleguezuelos-Manzano, C.; Puschhof, J.; Rosendahl Huber, A.; van Hoeck, A.; Wood, H.M.; Nomburg, J.; Gurjao, C.; Manders, F.; Dalmasso, G.; Stege, P.B.; et al. Mutational signature in colorectal cancer caused by genotoxic pks + E. coli. Nature 2020, 580, 269–273. [Google Scholar] [CrossRef]
- Fong, W.; Li, Q.; Yu, J. Gut microbiota modulation: A novel strategy for prevention and treatment of colorectal cancer. Oncogene 2020, 39, 4925–4943. [Google Scholar] [CrossRef]
- Ambalam, P.; Raman, M.; Purama, R.K.; Doble, M. Probiotics, prebiotics and colorectal cancer prevention. Best Pract. Res. Clin. Gastroenterol. 2016, 30, 119–131. [Google Scholar] [CrossRef]
- Malaguarnera, G.; Leggio, F.; Vacante, M.; Motta, M.; Giordano, M.; Biondi, A.; Basile, F.; Mastrojeni, S.; Mistretta, A.; MalaGuarnera, M.; et al. Probiotics in the gastrointestinal diseases of the elderly. J. Nutr. Health Aging 2012, 16, 402–410. [Google Scholar] [CrossRef]
- Uccello, M.; Malaguarnera, G.; Basile, F.; Dagata, V.; Malaguarnera, M.; Bertino, G.; Vacante, M.; Drago, F.; Biondi, A. Potential role of probiotics on colorectal cancer prevention. BMC Surg. 2012, 12, S35. [Google Scholar] [CrossRef] [Green Version]
- Gamallat, Y.; Meyiah, A.; Kuugbee, E.D.; Hago, A.M.; Chiwala, G.; Awadasseid, A.; Bamba, D.; Zhang, X.; Shang, X.; Luo, F.; et al. Lactobacillus rhamnosus induced epithelial cell apoptosis, ameliorates inflammation and prevents colon cancer development in an animal model. Biomed. Pharmacother. 2016, 83, 536–541. [Google Scholar] [CrossRef]
- Kuugbee, E.D.; Shang, X.; Gamallat, Y.; Bamba, D.; Awadasseid, A.; Suliman, M.A.; Zang, S.; Ma, Y.; Chiwala, G.; Xin, Y.; et al. Structural Change in Microbiota by a Probiotic Cocktail Enhances the Gut Barrier and Reduces Cancer via TLR2 Signaling in a Rat Model of Colon Cancer. Dig. Dis. Sci. 2016, 61, 2908–2920. [Google Scholar] [CrossRef] [PubMed]
- Yeung, C.-Y.; Chan, W.-T.; Jiang, C.-B.; Cheng, M.-L.; Liu, C.-Y.; Chang, S.-W.; Chiang Chiau, J.-S.; Lee, H.-C. Amelioration of Chemotherapy-Induced Intestinal Mucositis by Orally Administered Probiotics in a Mouse Model. PLoS ONE 2015, 10, e0138746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, H.; Dong, Y.; Zhang, B.; Wang, H.; Peter, C.C.K.; Gao, P.; Fu, H.; Gao, Y. Bifidobacterium Infantis Ameliorates Chemotherapy-Induced Intestinal Mucositis Via Regulating T Cell Immunity in Colorectal Cancer Rats. Cell Physiol. Biochem. 2017, 42, 2330–2341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montalban-Arques, A.; Scharl, M. Intestinal microbiota and colorectal carcinoma: Implications for pathogenesis, diagnosis, and therapy. EBioMedicine 2019, 48, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Collij, V.; Jaeger, M.; van den Munckhof, I.C.L.; Vich Vila, A.; Kurilshikov, A.; Gacesa, R.; Sinha, T.; Oosting, M.; Joosten, L.A.B.; et al. Gut microbial co-abundance networks show specificity in inflammatory bowel disease and obesity. Nat. Commun. 2020, 11, 4018. [Google Scholar] [CrossRef]
- Flemer, B.; Lynch, D.B.; Brown, J.M.R.; Jeffery, I.B.; Ryan, F.J.; Claesson, M.J.; O’Riordain, M.; Shanahan, F.; O’Toole, P.W. Tumour-associated and non-tumour-associated microbiota in colorectal cancer. Gut 2017, 66, 633–643. [Google Scholar] [CrossRef]
- Geller, L.T.; Barzily-Rokni, M.; Danino, T.; Jonas, O.H.; Shental, N.; Nejman, D.; Gavert, N.; Zwang, Y.; Cooper, Z.A.; Shee, K.; et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science 2017, 357, 1156–1160. [Google Scholar] [CrossRef] [Green Version]
- Viaud, S.; Saccheri, F.; Mignot, G.; Yamazaki, T.; Daillère, R.; Hannani, D.; Enot, D.P.; Pfirschke, C.; Engblom, C.; Pittet, M.J.; et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science 2013, 342, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.-L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [Green Version]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Routy, B.; Le Chatelier, E.; Derosa, L.; Duong, C.P.M.; Alou, M.T.; Daillère, R.; Fluckiger, A.; Messaoudene, M.; Rauber, C.; Roberti, M.P.; et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science 2018, 359, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villéger, R.; Lopès, A.; Veziant, J.; Gagnière, J.; Barnich, N.; Billard, E.; Boucher, D.; Bonnet, M. Microbial markers in colorectal cancer detection and/or prognosis. World J. Gastroenterol. 2018, 24, 2327–2347. [Google Scholar] [CrossRef] [PubMed]
- Mangifesta, M.; Mancabelli, L.; Milani, C.; Gaiani, F.; de’Angelis, N.; de’Angelis, G.L.; van Sinderen, D.; Ventura, M.; Turroni, F. Mucosal microbiota of intestinal polyps reveals putative biomarkers of colorectal cancer. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, M.A.; Bird, A.R. The Impact of Diet and Lifestyle on Gut Microbiota and Human Health. Nutrients 2014, 7, 17–44. [Google Scholar] [CrossRef]
- Thomas, A.M.; Manghi, P.; Asnicar, F.; Pasolli, E.; Armanini, F.; Zolfo, M.; Beghini, F.; Manara, S.; Karcher, N.; Pozzi, C.; et al. Metagenomic analysis of colorectal cancer datasets identifies cross-cohort microbial diagnostic signatures and a link with choline degradation. Nat. Med. 2019, 25, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Wirbel, J.; Pyl, P.T.; Kartal, E.; Zych, K.; Kashani, A.; Milanese, A.; Fleck, J.S.; Voigt, A.Y.; Palleja, A.; Ponnudurai, R.; et al. Meta-analysis of fecal metagenomes reveals global microbial signatures that are specific for colorectal cancer. Nat. Med. 2019, 25, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Poore, G.D.; Kopylova, E.; Zhu, Q.; Carpenter, C.; Fraraccio, S.; Wandro, S.; Kosciolek, T.; Janssen, S.; Metcalf, J.; Song, S.J.; et al. Microbiome analyses of blood and tissues suggest cancer diagnostic approach. Nature 2020, 579, 567–574. [Google Scholar] [CrossRef]
- Slowicka, K.; Petta, I.; Blancke, G.; Hoste, E.; Dumas, E.; Sze, M.; Vikkula, H.; Radaelli, E.; Haigh, J.J.; Jonckheere, S.; et al. Zeb2 drives invasive and microbiota-dependent colon carcinoma. Nat. Cancer 2020, 1, 620–634. [Google Scholar] [CrossRef]
Figure 1.
Dysbiosis and other factors contributing to the adenoma-carcinoma progression. The adenoma-carcinoma progression may occur because of the genomic instability caused by alterations in the gut microbiota. These changes may be supported by diet and lifestyle, which promote dysbiosis, inflammatory state and epithelial DNA damage, thus contributing to CRC development. The carcinogenesis leads to gut niche changes, which may favor the proliferation of opportunistic pathogens.
Figure 1.
Dysbiosis and other factors contributing to the adenoma-carcinoma progression. The adenoma-carcinoma progression may occur because of the genomic instability caused by alterations in the gut microbiota. These changes may be supported by diet and lifestyle, which promote dysbiosis, inflammatory state and epithelial DNA damage, thus contributing to CRC development. The carcinogenesis leads to gut niche changes, which may favor the proliferation of opportunistic pathogens.


{kind=link}
Table 1.
Studies of gut bacteria associated with the development of adenoma and/or CRC
| Authors (Year). | Bacteria | Methods | Sample Size | Statistical Significance (p Value, Odds Ratio, and/or Hazard Ratio) | Clinical Evidence |
|---|---|---|---|---|---|
| Hale et al. (2017) [81] | Bilophila, Desulfovibrio, Mogibacterium, Bacteroidetes spp. | 16S rRNA gene sequencing | 233 adenomas, 547 controls | AUC of 0.6599, (p = 0.001) | Adenoma and CRC development |
| Kasai et al. (2016) [90] | Actinomyces, Atopobium, Fusobacterium, and Haemophilus spp. | T-RFLP and NGS | 49 controls, 50 adenomas, 9 CRC (3/9 invasive cancer and 6/9 carcinoma in adenoma | Actinomyces odontolyticus (p = 0.007), Bacteroides fragile (p = 0.004), Clostridium nexile (p = 0.036), Fusobacterium varium (p = 0.022), Haemophilus parainfluenzae (p = 0.020), Prevotella stercorea (p = 0.022), Streptococcus gordonii (p = 0.014), and Veillonella dispar (p = 0.042) | Association with CRC development |
| Feng et al. (2015) [91] | Bacteroides, Prevotella, and Parabacteroides spp. Alistipes putredinis, Bilophila wadsworthia, Lachnospiraceae bacterium, Fusobacterium, E. coli | MGWAS on stools | 55 controls, 42 advanced adenoma, 41 CRC | p = 0.005, p < 0.001 (among the groups respectively, Kruskal–Wallis test) | Development of advanced adenoma and CRC |
| Peters et al. (2016) [92] | Reduction in Clostridia (Clostridiaceae, and Lachnospiraceae), and enrichment in Bacilli and Gammaproteobacteria, (Enterobacteriales), Actinomyces and Streptococcus | 16S rRNA gene sequencing | 540 total: 144 CA, 73 serrated polyps, 323 polyp-free controls | CA p = 0.03; advanced CA p = 0.004. Distal or advanced CA vs. controls (p = 0.02 and p = 0.002) | Early stages of carcinogenesis and development of CAs |
| Li et al. (2016) [93] | F. nucleatum | FQ-PCR in CRC and normal tissues, FISH analysis (to confirm 22 cases) | 101 CRC | CRC vs. controls: 0.242 (95% C.I. 0.178–0.276) vs. 0.050 (95% C.I. 0.023–0.067), p < 0.001 | Association with CRC development and metastasis |
| Fukugaiti et al. (2015) [94] | F. nucleatum and Clostridium difficile | qRT-PCR | 17 total: 7 CRC | F. nucleatum (p < 0.01); Clostridium difficile (p < 0.04) | Possible role of in CRC carcinogenesis |
| Yu et al. (2015) [95] | Fusobacterium, Streptococcus and Enterococcus spp. | Pyrosequencing of the 16S ribosome RNA (rRNA) from fecal samples | 52 controls, 47 advanced adenoma, 42 CRC | Increase of the three bacteria groups during the adenoma-carcinoma sequence: p < 0.05. Increase of the Fusobacterial phylum: from normal (0.27%) to adenoma (0.61%) to CRC (1.69%) (p = 0.016) | F. nucleatum colonization in the gut may favor colorectal tumorigenesis |
| Yu et al. (2016) [96] | F. nucleatum | 16S rRNA FISH | 35 HPs, 33 SSAs, 48 proximal CRCs, and 10 matched metastatic lymph nodes | Higher Fusobacterium in proximal HPs and SSAs vs. proximal TAs and distal TAs (p < 0.05). Higher Fusobacterium in more proximal CRCs vs. distal CRCs (p < 0.05), and in matched metastatic lymph nodes vs. nonmetastatic lymph nodes (p < 0.001). | Carcinogenesis of proximal colon through the serrated neoplasia pathway. Less important role in the TA-carcinoma sequence. |
| Mima et al. (2016) [97] | F. nucleatum | Assessment of DNA in CRC tissue | 1069 CRC in the Nurses’ Health Study and the Health Professionals Follow-up Study | HRs for CRC-specific mortality in F. nucleatum-low cases and F. nucleatum-high cases:1.25 (95% C.I. 0.82 to 1.92) and 1.58 (95% C.I. 1.04 to 2.39), respectively, (p for trend = 0.020). Association with MSI-high OR 5.22 (95% C.I. 2.86 to 9.55) | Evidence of poorer survival, and potential use as prognostic biomarker |
| Yu et al. (2017) [98] | Fusobacterium, Anaerosporobacter, Parvimonas, Peptostreptococcus, and Prevotella | Pyrosequence (Roche 454 GS FLX) | Phase I: 16 CRC with recurrence and 15 CRC without recurrence Phase II: 48 CRC without recurrence and 44 CRC with recurrence | Recurrence rate in the high-risk vs. low-risk group (73.4% vs. 30.9%, p < 0.001) | High amount of F. nucleatum could favor CRC chemoresistance and predict potential CRC recurrence |
| Little et al. (2019) [99] | S. bovis | S. bovis-positive blood cultures | 86 patients with S. bovis bacteriemia | 30 patients underwent colonoscopy with 3 (10%) having adenocarcinoma and 11 (37%) having adenomatous polyps. Gastroenterology consultation was significantly associated with having a colonoscopy (p = 0.001). | Association between S. bovis bacteremia and CRC risk. |
| Corredoira-Sánchez et al. (2012) [100] | S. gallolyticus | S. gallolyticus positive blood cultures | 109 patients with S. gallolyticus bacteriemia and 196 controls | 98 patients underwent colonoscopy: 57 had adenomas (39 advanced adenomas) and 12 had invasive carcinomas. Total colorectal neoplasia in patients with S. gallolyticus bacteriemia vs. controls: 70% vs. 32%; OR 5.1, 95% C.I. 3.0–8.6). For advanced adenomas: 40% vs. 16%; OR 3.5; 95% C.I. 2.0–6.1. For invasive carcinomas: 12% vs. 5%; OR 2.9, 95% C.I. 1.2–6.9. | S. gallolyticus infection could represent a valuable marker for detection of occult CRC |
| Butt et al. (2016) [101] | S. gallolyticus | Antibody responses to recombinant affinity-purified S. gallolyticus pilus proteins Gallo1569, 2039, 2178 and 2179 were analysed by multiplex serology | 576 CRC and 576 controls | Antibody responses to Gallo2039 (OR 1.58, 95% C.I. 1.09–2.28), Gallo2178 (OR 1.58, 95% C.I. 1.09–2.30) and Gallo2179 (OR 1.45, 95% C.I. 1.00–2.11) were significantly associated with CRC risk. The association was stronger for positivity to two or more pilus proteins of Gallo1569, Gallo2178 and Gallo2179 (OR 1.93, 95% C.I. 1.04–3.56) and for double-positivity to Gallo2178 and Gallo2179 (OR 3.54, 95% C.I. 1.49–8.44) | Association between S. gallolyticus infection and CRC risk |
| Purcell et al. (2017) [102] | ETBF | Quantitative PCR | 150 consecutive patients who underwent colonoscopy | Associations with low-grade dysplasia (p = 0.007), tubular adenomas (p = 0.027), and serrated polyps (p = 0.007) | Potential marker of early colorectal carcinogenesis |
| Xie et al. (2016) [103] | ETBF and pks + E. coli | Quantitative real time PCR | 36 adenoma, 18 controls | Increase of toxin produced by ETBF in adenoma vs. controls (p = 0.003) and in pks + E. coli (p < 0.001) | Possible relationship with carcinogenesis in adenomas |
| Zamani et al. (2020) [104] | ETBF | Quantitative real-time PCR | 68 precancerous and CRC condition, 52 controls | Positivity of bft gene in patients vs. controls p = 0.00. OR 22.22 (95% C.I. 5–98.74) | Risk factor and screening marker for developing CRC |
| Viljoen, et al. (2015) [105] | Fusobacterium spp., Streptococcus gallolyticus, Enterococcus faecalis, ETBF, Enteropathogenic E. coli, and afaC− or pks + E. coli | Quantitative PCR | Paired tumor and normal tissue samples from 55 CRC | Fusobacterium was significantly higher in CRC vs. controls (p < 0.001). ETBF (FDR = 0.04 and 0.002 for controls and CRC, respectively) and Fusobacterium spp. (FDR = 0.03 CRC) levels were significantly higher in stage III/IV CRC | Associations with clinicopathological features, mainly for Fusobacterium and ETBF |
| Ambrosi et al. (2019) [106] | E. coli | 16S rRNA gene sequencing and PCR | Phase I: 20 adenomatous polyps, 20 polyps, 20 adjacent tissue close to polyps (5–7 cm), 10 controls Phase II: total 1500 biopsies, 600 adenomatous polyps, 600 adjacent non-adenomatous tissues, 300 controls | In polyps, prevalence of phylogroup A and B2, strong biofilm and poor protease producers (p < 0.05). Phylogroup B2 showed highest isolates with virulence factor score ≥10 (p = 0.0034). | Association of specific phenotypes of E. coli with adenomatous polyps |
| Iyadorai et al. (2020) [107] | Pks + E. coli | 16S rRNA gene sequencing and PCR | Phase I: Primary colon epithelial and CRC (HCT116) cell lines Phase II: 48 CRC (48 tumor and 48 matching non-malignant tissue), 23 controls (23 proximal and 23 distal biopsies) | 16.7% of CRC patients were positive for pks + E. coli vs. 4.35% of controls (p = 0.144). Pks + E. coli was observed in 1/26 colonoscopy biopsies from controls vs. 16/96 tissue samples from CRC (p = 0.01) | Initiation and development of CRC |
Abbreviations: T-RFLP: terminal restriction fragment length polymorphism, NGS: next-generation sequencing, MGWAS: metagenome-wide association study, CA: conventional adenoma, FQ-PCR: fluorescent quantitative polymerase chain reaction, FISH: fluorescence in situ hybridization, C.I.: confidence interval, qRT-PCR: real-time quantitative reverse transcription polymerase chain reaction, HP: proximal hyperplastic polyp, SSA: sessile serrated adenoma, TA: traditional adenoma, HR: hazard ratio, OR: odds ratio, ETBF: enterotoxigenic Bacteroides fragilis, afaC: afimbrial adhesin, pks: polyketide synthase, FDR: false discovery rate.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Vacante, M.; Ciuni, R.; Basile, F.; Biondi, A. Gut Microbiota and Colorectal Cancer Development: A Closer Look to the Adenoma-Carcinoma Sequence. Biomedicines 2020, 8, 489. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8110489
AMA Style
Vacante M, Ciuni R, Basile F, Biondi A. Gut Microbiota and Colorectal Cancer Development: A Closer Look to the Adenoma-Carcinoma Sequence. Biomedicines. 2020; 8(11):489. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8110489
Chicago/Turabian StyleVacante, Marco, Roberto Ciuni, Francesco Basile, and Antonio Biondi. 2020. "Gut Microbiota and Colorectal Cancer Development: A Closer Look to the Adenoma-Carcinoma Sequence" Biomedicines 8, no. 11: 489. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines8110489
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.