
IL-17A Damages the Blood–Retinal Barrier through Activating the Janus Kinase 1 Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Transwell Cell Culture
2.3. Cell Treatments
2.4. Transepithelial Electrical Resistance (TEER)
2.5. FITC-Permeability Assay
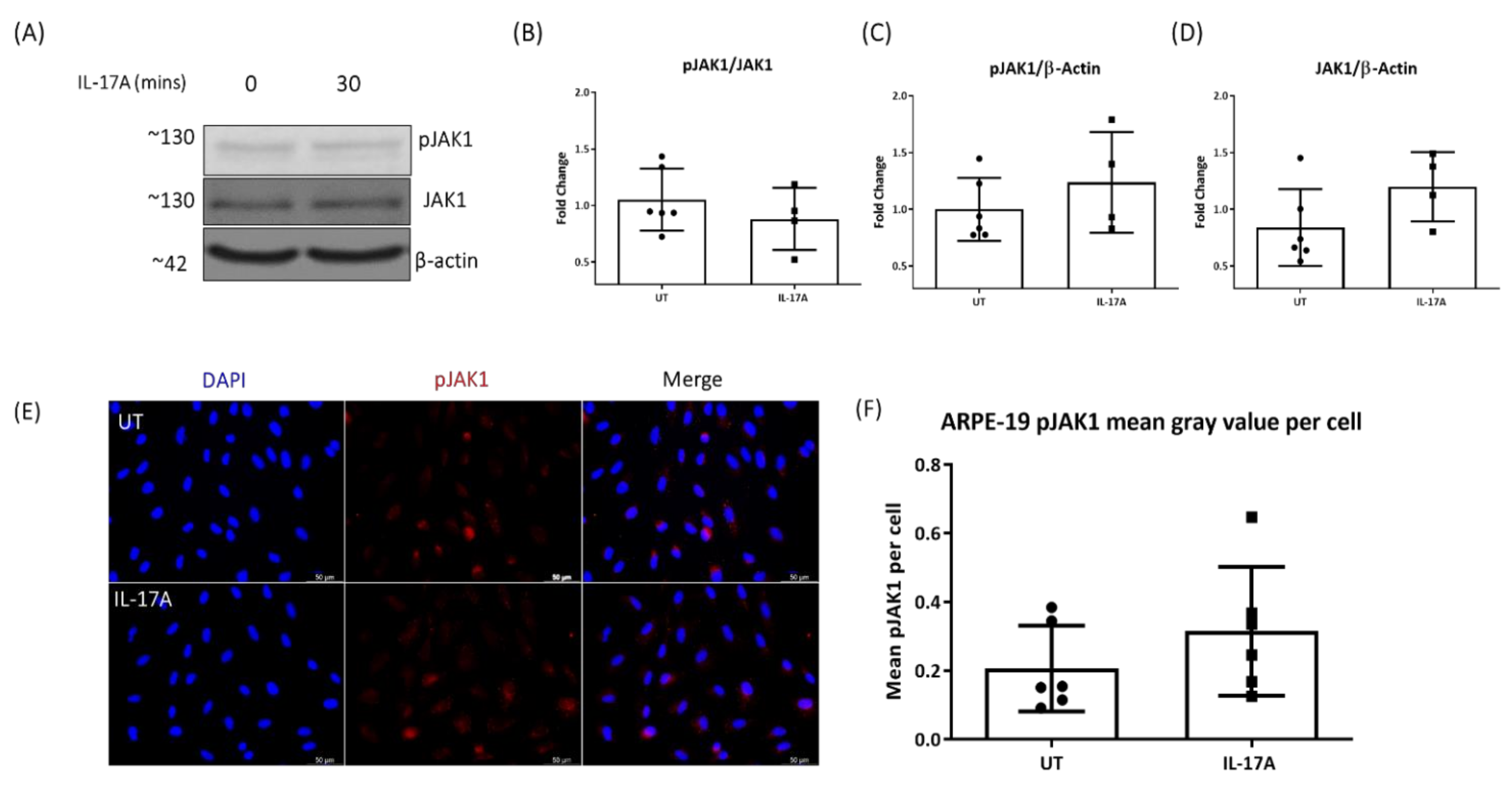
2.6. Immunocytochemistry
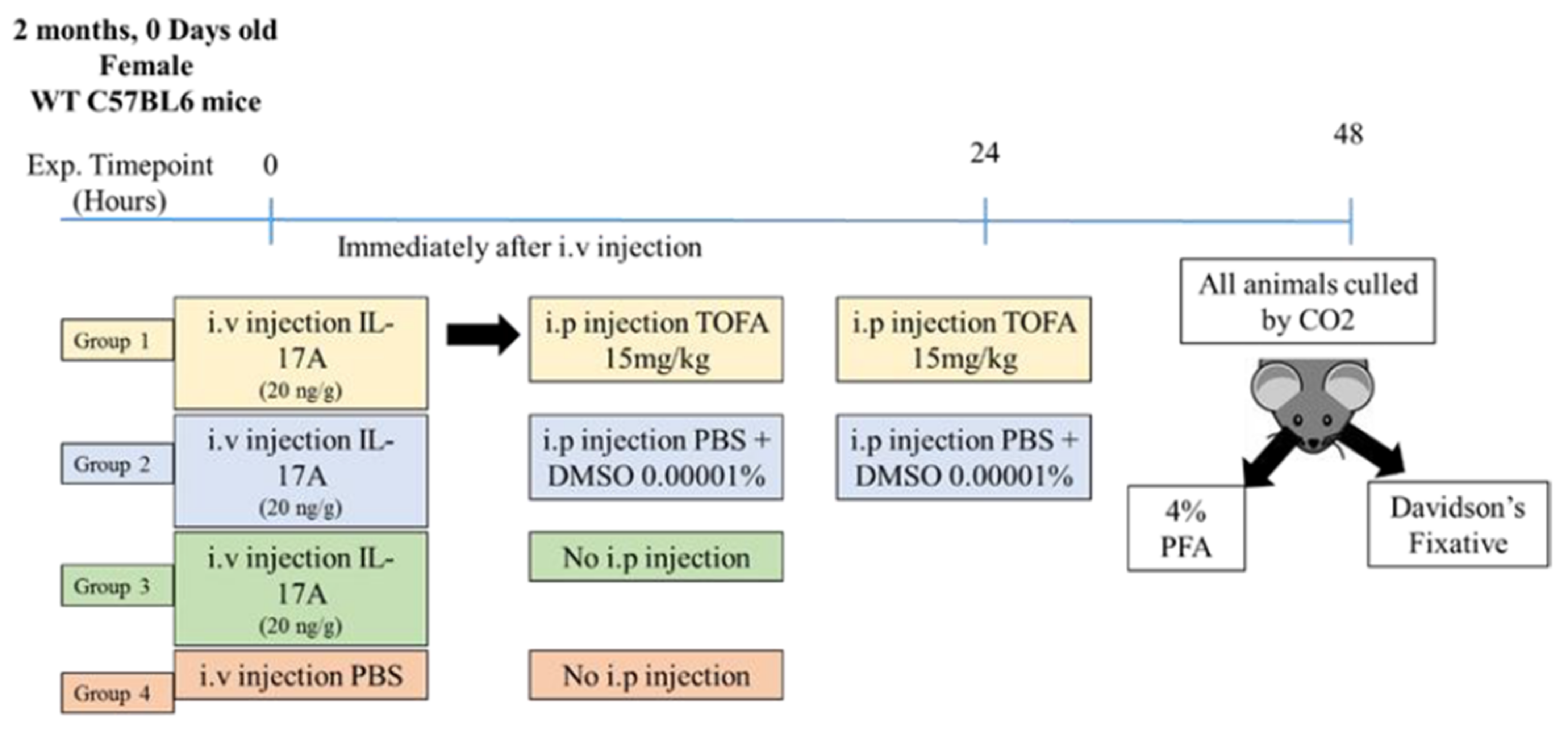
2.7. Animal Care and Housing
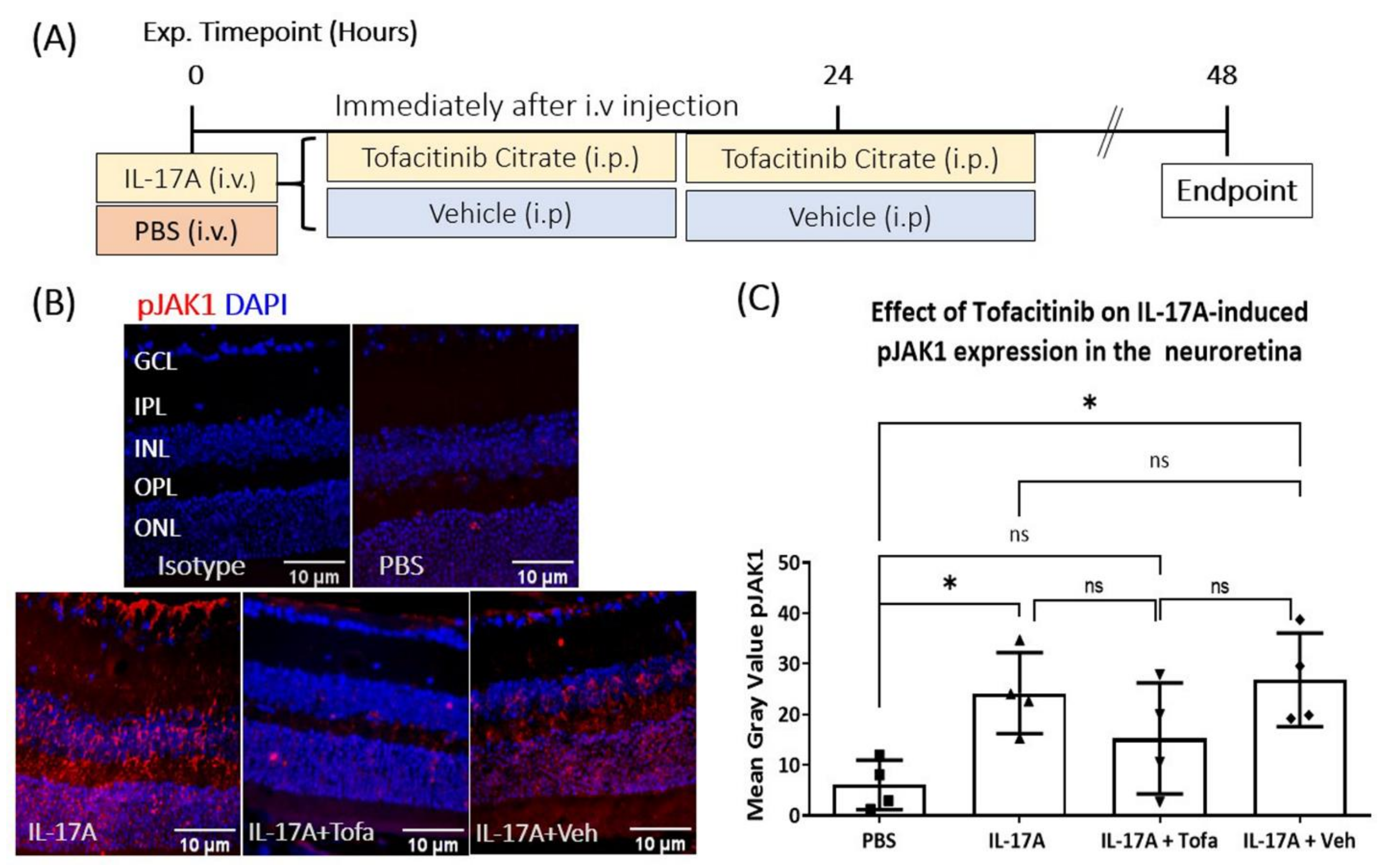
2.8. IL-17A Intravitreal and Intravenous Injection
2.9. In Vivo Fundus Imaging Using Micron IV
2.10. Albumin Leakage Quantification
2.11. pJAK1 Quantification Neuroretina
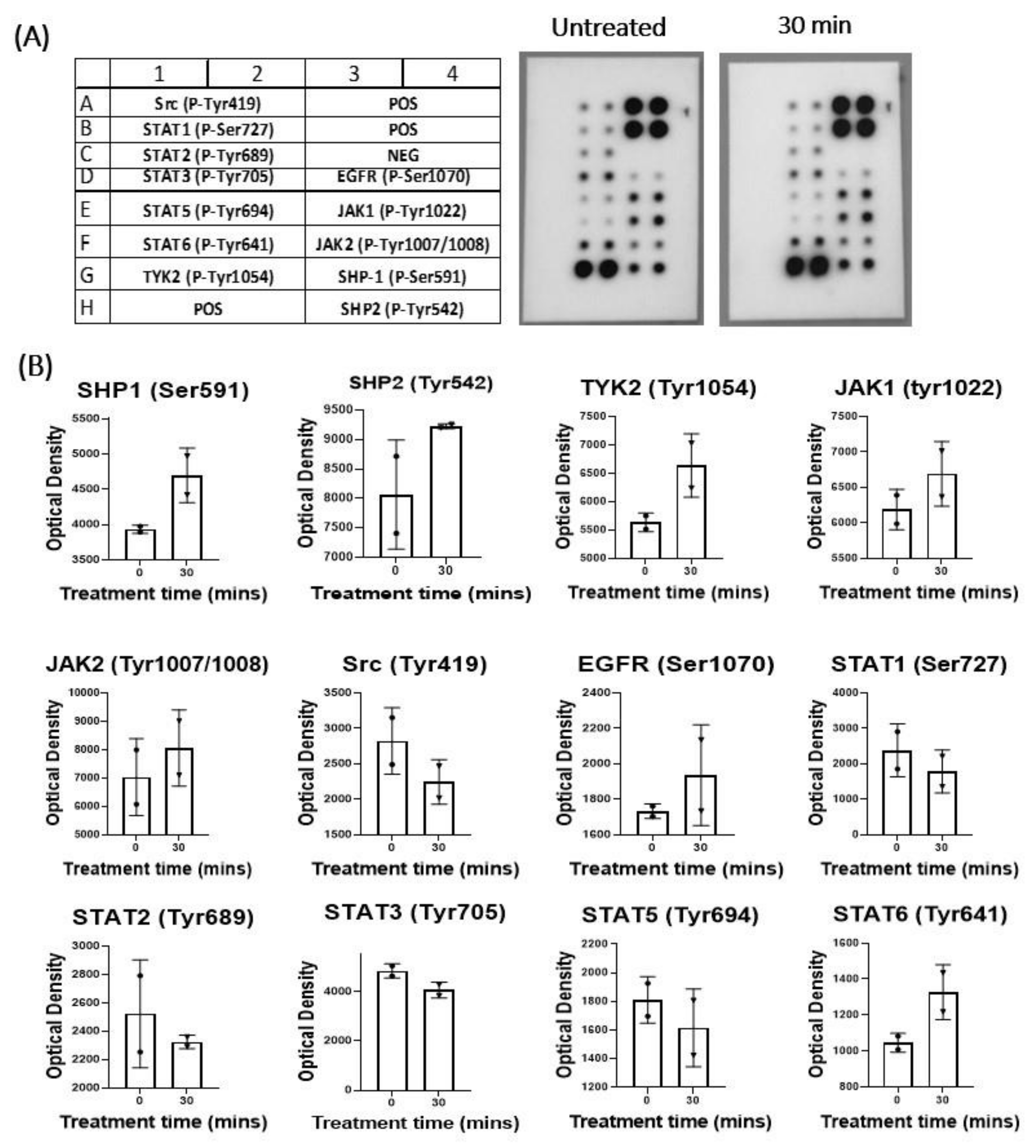
2.12. Western Blot
2.13. Statistical Analysis
3. Results
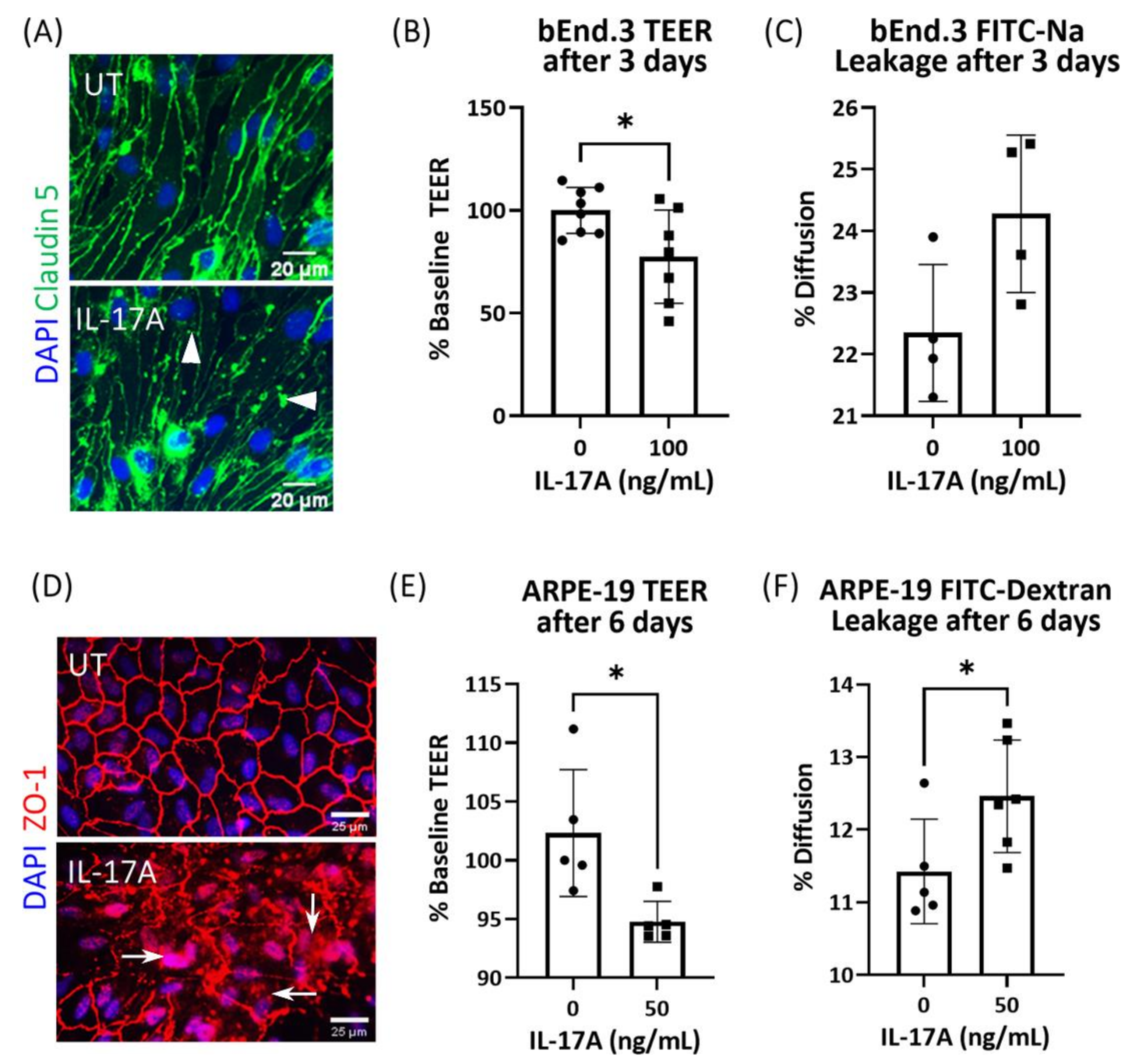
3.1. The Effect of IL-17A on bEnd3 Cell and ARPE19 Cell Tight Junctions and Barrier Function
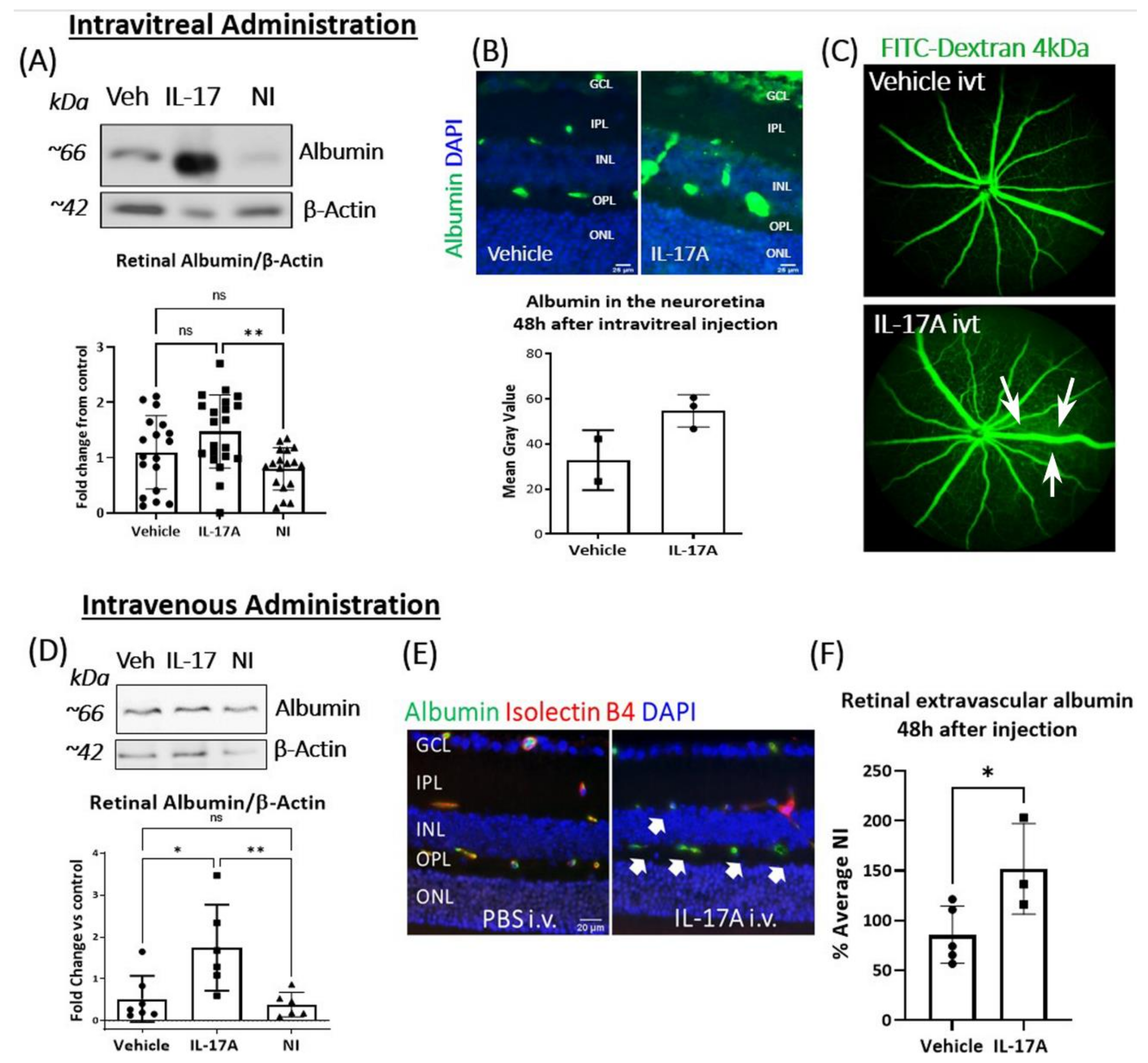
3.2. The Effect of IL-17A on BRB Integrity In Vivo
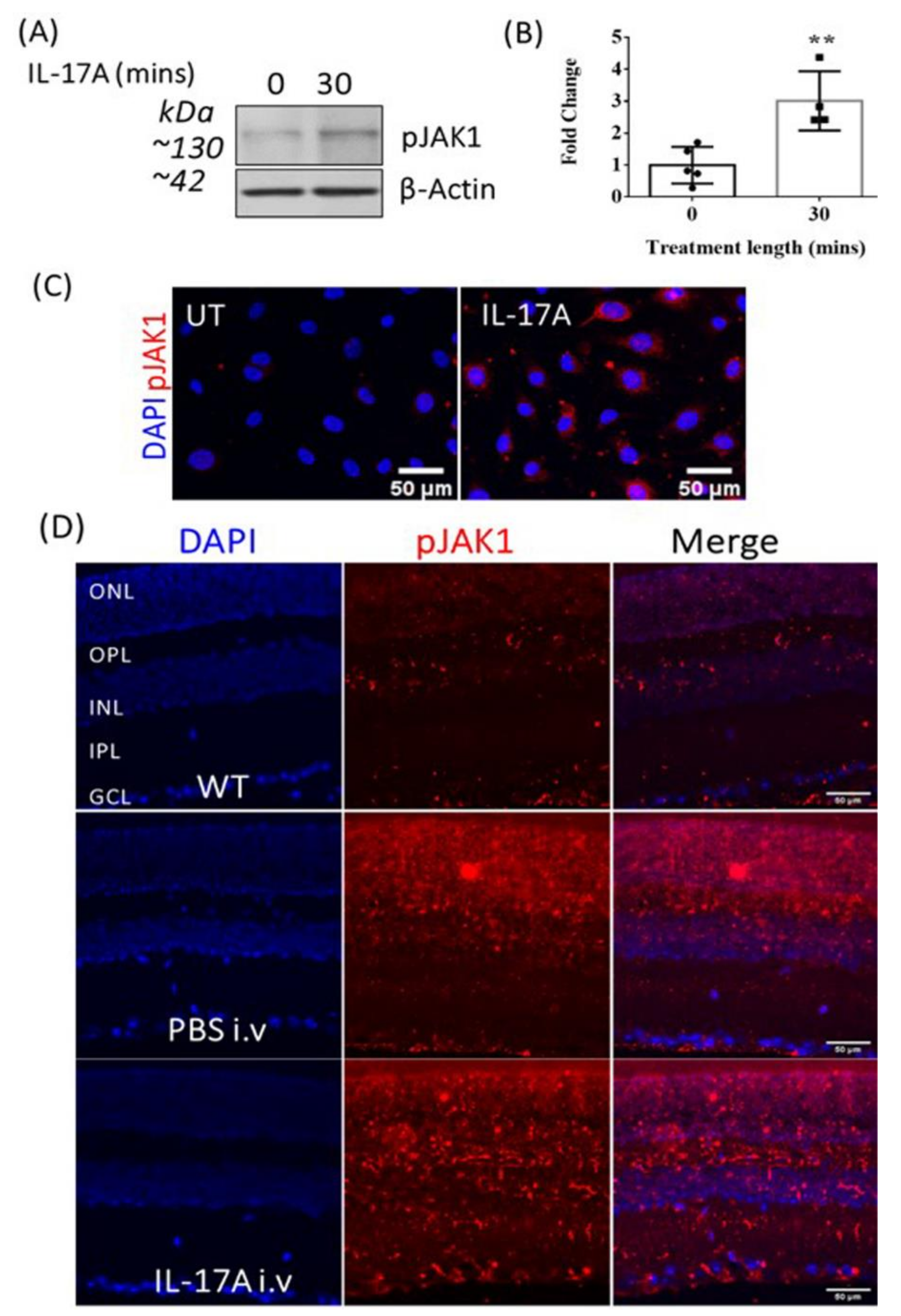
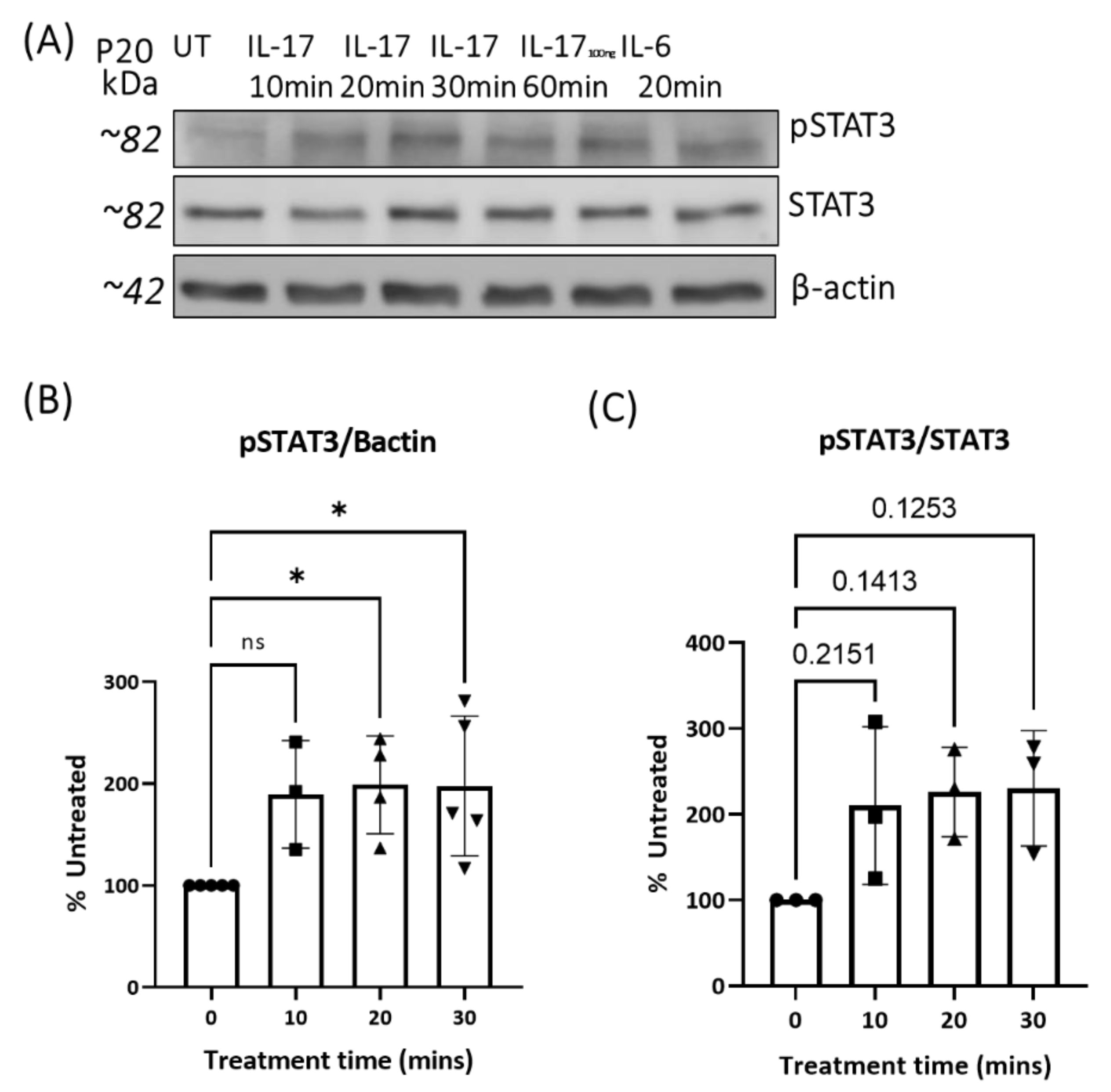
3.3. The Effect of IL-17A on pJAK1 Expression In Vitro and In Vivo
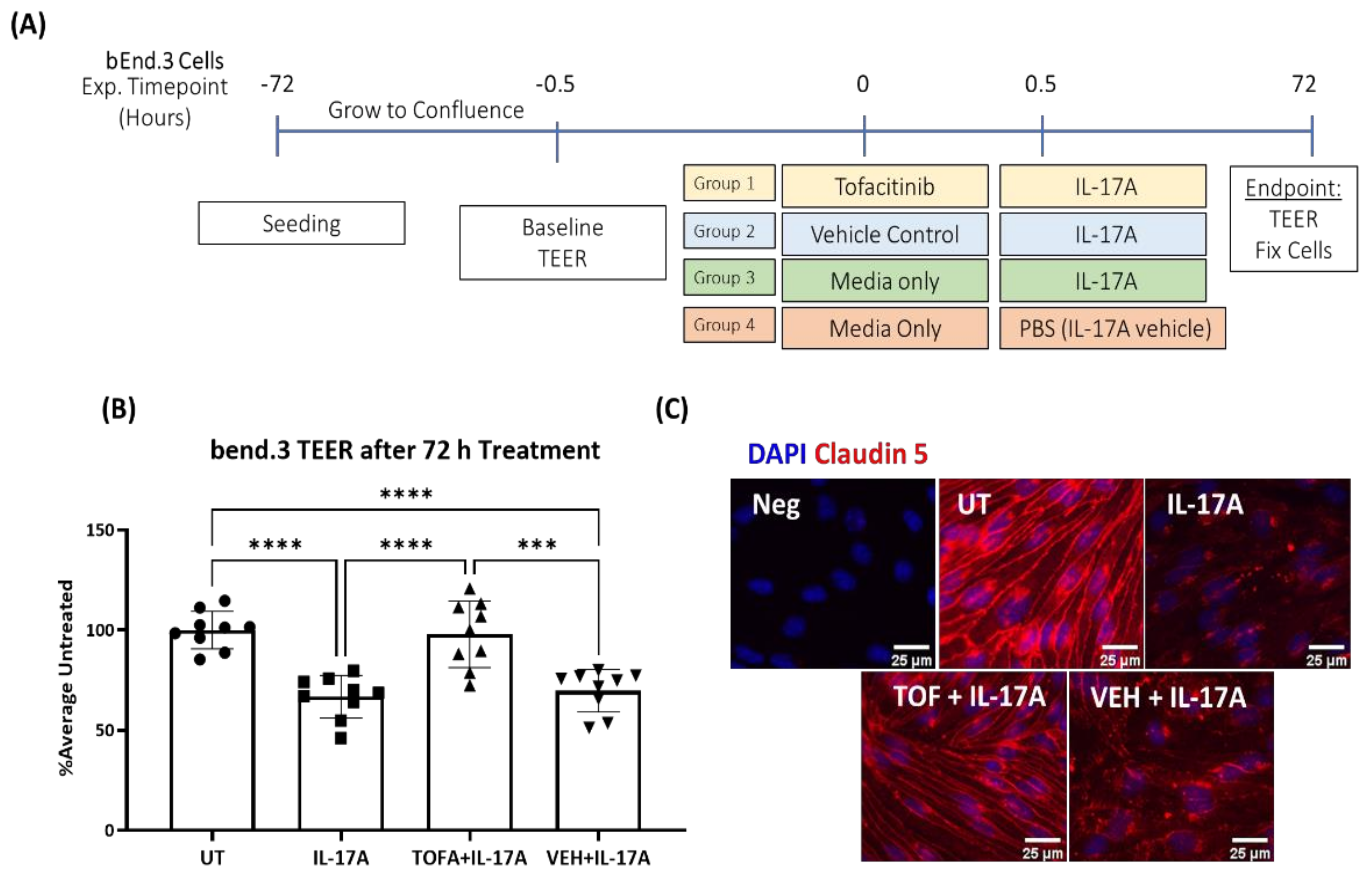
3.4. JAK1 Inhibitor Tofacitinib Citrate Ameliorates IL-17A-Mediated Leakage in an In Vitro Model of iBRB
3.5. The Effect of Tofacitinib Citrate in IL-17A-Mediated Retinal pJAK1 Expression
3.6. The Effect of Tofacitinib Citrate on IL-17A-Mediated BRB Leakage In Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A






References
- Bhagat, N.; Grigorian, R.A.; Tutela, A.; Zarbin, M.A. Diabetic Macular Edema: Pathogenesis and Treatment. Surv. Ophthalmol. 2009, 54, 1–32. [Google Scholar] [CrossRef] [PubMed]
- Hudson, N.; Cahill, M.; Campbell, M. Inner blood-retina barrier involvement in dry age-related macular degeneration (AMD) pathology. Neural Regen. Res. 2020, 15, 1656–1657. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yu, X.-W.; Zhang, D.-D.; Fan, Z.-G. Blood-retinal barrier as a converging pivot in understanding the initiation and development of retinal diseases. Chin. Med. J. 2020, 133, 2586–2594. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Forrester, J.V.; Liversidge, J.; Crane, I.J. Leukocyte trafficking in experimental autoimmune uveitis: Breakdown of blood-retinal barrier and upregulation of cellular adhesion molecules. Investig. Opthalmol. Vis. Sci. 2003, 44, 226–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forrester, J.V. Bowman lecture on the role of inflammation in degenerative disease of the eye. Eye 2013, 27, 340–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilger, B.C.; Abarca, E.M.; Salmon, J.H.; Patel, S. Treatment of Acute Posterior Uveitis in a Porcine Model by Injection of Triamcinolone Acetonide into the Suprachoroidal Space Using Microneedles. Investig. Opthalmol. Vis. Sci. 2013, 54, 2483–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elman, M.J.; Aiello, L.P.; Beck, R.W.; Bressler, N.M.; Bressler, S.B.; Edwards, A.R.; Ferris, F.; Friedman, S.M.; Glassman, A.R.; Miller, K.M.; et al. Randomized Trial Evaluating Ranibizumab Plus Prompt or Deferred Laser or Triamcinolone Plus Prompt Laser for Diabetic Macular Edema. Ophthalmology 2010, 117, 1064–1077.e35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ford, J.A.; Lois, N.; Royle, P.; Clar, C.; Shyangdan, D.; Waugh, N. Current treatments in diabetic macular oedema: Systematic review and meta-analysis. BMJ Open 2013, 3, e002269. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.G.; Scott, I.U.; Stewart, M.W.; Harry, W.F., Jr. Update on corticosteroids for diabetic macular edema. Clin. Ophthalmol. 2016, 10, 1723–1730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Navarro, V.; Cervera-Taulet, E.; Navarro-Palop, C.; Monferrer-Adsuara, C.; Hernández-Bel, L.; Montero-Hernández, J. Intravitreal dexamethasone implant Ozurdex® in naïve and refractory patients with different subtypes of diabetic macular edema. BMC Ophthalmol. 2019, 19, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leinonen, S.; Immonen, I.; Kotaniemi, K. Fluocinolone acetonide intravitreal implant (Retisert®) in the treatment of sight threatening macular oedema of juvenile idiopathic arthritis-related uveitis. Acta Ophthalmol. 2018, 96, 648–651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estebainha, R.; Goldhardt, R.; Falcão, M. A New Approach for Diabetic Macular Edema Treatment: Review of clinical practice results with 0.19 mg fluocinolone acetonide intravitreal implant including vitrectomized eyes. Curr. Ophthalmol. Rep. 2020, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Veritti, D.; Sarao, V.; Diplotti, L.; Samassa, F.; Lanzetta, P. Fluocinolone acetonide for the treatment of diabetic macular edema. Expert Opin. Pharmacother. 2017, 18, 1507–1516. [Google Scholar] [CrossRef] [PubMed]
- Phulke, S.; Kaushik, S.; Kaur, S.; Pandav, S. Steroid-induced Glaucoma: An Avoidable Irreversible Blindness. J. Curr. Glaucoma Prac. 2017, 11, 67–72. [Google Scholar] [CrossRef]
- Goñi, F.J.; Stalmans, I.; Denis, P.; Nordmann, J.-P.; Taylor, S.; Diestelhorst, M.; Figueiredo, A.R.; Garway-Heath, D.F. Elevated Intraocular Pressure After Intravitreal Steroid Injection in Diabetic Macular Edema: Monitoring and Management. Ophthalmol. Ther. 2016, 5, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Kersey, J.P.; Broadway, D.C. Corticosteroid-induced glaucoma: A review of the literature. Eye 2005, 20, 407–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, J.W.; Le Couter, J.; Strauss, E.C.; Ferrara, N. Vascular Endothelial Growth Factor A in Intraocular Vascular Disease. Ophthalmology 2013, 120, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Virgili, G.; Parravano, M.; Evans, J.R.; Gordon, I.; Lucenteforte, E. Anti-vascular endothelial growth factor for diabetic macular oedema: A network meta-analysis. Cochrane Database Syst. Rev. 2018, 10, CD007419. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Jiao, B.; Han, Q. Effect of Intravitreal Anti-Vascular Growth Factor Agents with or Without Macular Photocoagulation on Diabetic Macular Edema: A Systematic Review and Meta-Analysis. Diabetes Ther. 2019, 10, 1283–1296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, S.U.; Maturi, R.K. Therapeutic Options in Refractory Diabetic Macular Oedema. Drugs 2017, 77, 481–492. [Google Scholar] [CrossRef]
- Downey, L.; Acharya, N.; Devonport, H.; Gale, R.; Habib, M.; Manjunath, V.; Mukherjee, R.; Severn, P. Treatment choices for diabetic macular oedema: A guideline for when to consider an intravitreal corticosteroid, including adaptations for the COVID-19 era. BMJ Open Ophthalmol. 2021, 6, e000696. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Takamura, Y.; Morioka, M.; Gozawa, M.; Matsumura, T.; Inatani, M. Microaneurysm density in residual oedema after anti-vascular endothelial growth factor therapy for diabetic macular oedema. Acta Ophthalmol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Hombrebueno, J.R.; Ali, I.H.; Xu, H.; Chen, M. Sustained intraocular VEGF neutralization results in retinal neurodegeneration in the Ins2Akita diabetic mouse. Sci. Rep. 2015, 5, 18316. [Google Scholar] [CrossRef]
- Llorián-Salvador, M.; Barabas, P.; Byrne, E.M.; Lechner, J.; Augustine, J.; Curtis, T.M.; Chen, M.; Xu, H. VEGF-B Is an Autocrine Gliotrophic Factor for Müller Cells under Pathologic Conditions. Investig. Opthalmology Vis. Sci. 2020, 61, 35. [Google Scholar] [CrossRef]
- Rossino, M.G.; Dal Monte, M.; Casini, G. Relationships Between Neurodegeneration and Vascular Damage in Diabetic Retinopathy. Front. Neurosci. 2019, 13, 1172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Yu, H.; Yu, Y.; Geng, Y.; Li, D.; Yang, C.; Lv, Q.; Lu, L.; Liu, T.; Li, G.; et al. Levels of Inflammatory Cytokines IL-1β, IL-6, IL-8, IL-17A, and TNF-α in Aqueous Humour of Patients with Diabetic Retinopathy. J. Diabetes Res. 2018, 2018, 8546423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funk, M.; Schmidinger, G.; Maar, N.; Bolz, M.; Benesch, T.; Zlabinger, G.; Schmidt-Erfurth, U. Angiogenic and inflammatory markers in the intraocular fluid of eyes with diabetic macular edema and influence of therapy with Bevacizumab. Retina 2010, 30, 1412–1419. [Google Scholar] [CrossRef]
- Park, Y.-G.; Jee, D.; Kwon, J.-W. Aqueous Humor Cytokine Levels in Diabetic Macular Edema Patients with Cotton-Wool Spots. J. Diabetes Res. 2019, 2019, 8137417. [Google Scholar] [CrossRef]
- Litwińska, Z.; Sobuś, A.; Łuczkowska, K.; Grabowicz, A.; Mozolewska-Piotrowska, K.; Safranow, K.; Kawa, M.P.; Machalinski, B.; Machalińska, A. The Interplay Between Systemic Inflammatory Factors and MicroRNAs in Age-Related Macular Degeneration. Front. Aging Neurosci. 2019, 11, 286. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, S.; Takahashi, H.; Tan, X.; Inoue, Y.; Nomura, Y.; Arai, Y.; Fujino, Y.; Kawashima, H.; Yanagi, Y. Changes in multiple cytokine concentrations in the aqueous humour of neovascular age-related macular degeneration after 2 months of ranibizumab therapy. Br. J. Ophthalmol. 2018, 102, 448–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, S.; Yin, H.; Zheng, M.; Tang, Y.; Lu, B.; Chen, X.; Fu, Q.; Qin, Z.; Lyu, D.; Tang, Q.; et al. Cytokine profiling reveals increased serum inflammatory cytokines in idiopathic choroidal neovascularization. BMC Ophthalmol. 2019, 19, 94. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Shen, X.; Ding, C.; Qi, C.; Li, K.; Li, X.; Jala, V.R.; Zhang, H.-G.; Wang, T.; Zheng, J.; et al. Pivotal Role of Dermal IL-17-Producing γδ T Cells in Skin Inflammation. Immunity 2011, 35, 596–610. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.; Li, Y.; Xiao, Y.; Hu, W.; Wang, X.; Wang, P.; Zhang, X.; Yang, J.; Huang, Y.; He, W.; et al. Neutralization of interleukin-17A alleviates burn-induced intestinal barrier disruption via reducing pro-inflammatory cytokines in a mouse model. Burn. Trauma 2019, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- Setiadi, A.F.; Abbas, A.R.; Jeet, S.; Wong, K.; Bischof, A.; Peng, I.; Lee, J.; Bremer, M.; Eggers, E.L.; DeVoss, J.; et al. IL-17A is associated with the breakdown of the blood-brain barrier in relapsing-remitting multiple sclerosis. J. Neuroimmunol. 2019, 332, 147–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sigurdardottir, S.; Zapadka, T.E.; Lindstrom, S.I.; Liu, H.; Taylor, B.E.; Lee, C.A.; Kern, T.S.; Taylor, P.R. Diabetes-mediated IL-17A enhances retinal inflammation, oxidative stress, and vascular permeability. Cell Immunol. 2019, 341, 103921. [Google Scholar] [CrossRef]
- Xu, H.; Cai, M.; Zhang, X. Effect of the blockade of the IL-23-Th17-IL-17A pathway on streptozotocin-induced diabetic retinopathy in rats. Graefe’s Arch. Clin. Exp. Ophthalmol. 2015, 253, 1485–1492. [Google Scholar] [CrossRef]
- Qiu, A.-W.; Liu, Q.-H.; Wang, J.-L. Blocking IL-17A Alleviates Diabetic Retinopathy in Rodents. Cell. Physiol. Biochem. 2017, 41, 960–972. [Google Scholar] [CrossRef]
- Qiu, A.-W.; Bian, Z.; Mao, P.-A.; Liu, Q.-H. IL-17A exacerbates diabetic retinopathy by impairing Müller cell function via Act1 signaling. Exp. Mol. Med. 2016, 48, e280. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Liu, Y.; Lu, S.; Cai, X. Genetic Variants of Interleukin 17A Are Functionally Associated with Increased Risk of Age-Related Macular Degeneration. Inflammation 2014, 38, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.I.; Bayry, J. A role for IL-17 in age-related macular degeneration. Nat. Rev. Immunol. 2013, 13, 701. [Google Scholar] [CrossRef] [PubMed]
- Guedes, M.C.E.; Arroz, M.J.; Martins, C.; Angelo-Dias, M.; Proença, R.D.; Borrego, L.M. Regulatory T cells and IL-17A levels in noninfectious uveitis. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Jawad, S.; Liu, B.; Agron, E.; Nussenblatt, R.B.; Sen, H.N. Elevated Serum Levels of Interleukin-17A in Uveitis Patients. Ocul. Immunol. Inflamm. 2013, 21, 434–439. [Google Scholar] [CrossRef]
- Kezic, J.M.; Glant, T.T.; Rosenbaum, J.T.; Rosenzweig, H.L. Neutralization of IL-17 ameliorates uveitis but damages photoreceptors in a murine model of spondyloarthritis. Arthritis Res. Ther. 2012, 14, R18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Ren, X.; Liao, N.; Wen, F. Th17 cell frequency and IL-17A concentrations in peripheral blood mononuclear cells and vitreous fluid from patients with diabetic retinopathy. J. Int. Med. Res. 2016, 44, 1403–1413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimura, T.; Sonoda, K.-H.; Miyazaki, Y.; Iwakura, Y.; Ishibashi, T.; Yoshimura, A.; Yoshida, H. Differential roles for IFN- and IL-17 in experimental autoimmune uveoretinitis. Int. Immunol. 2008, 20, 209–214. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Wei, L.; Meyerle, C.; Tuo, J.; Sen, H.N.; Li, Z.; Chakrabarty, S.; Agrón, E.; Chan, C.-C.; Klein, M.L.; et al. Complement component C5a Promotes Expression of IL-22 and IL-17 from Human T cells and its Implication in Age-related Macular Degeneration. J. Transl. Med. 2011, 9, 111. [Google Scholar] [CrossRef] [Green Version]
- Ardeljan, D.; Wang, Y.; Park, S.; Shen, D.; Chu, X.K.; Yu, C.-R.; Abu-Asab, M.; Tuo, J.; Eberhart, C.G.; Olsen, T.W.; et al. Interleukin-17 Retinotoxicity Is Prevented by Gene Transfer of a Soluble Interleukin-17 Receptor Acting as a Cytokine Blocker: Implications for Age-Related Macular Degeneration. PLoS ONE 2014, 9, e95900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Yang, P.; Li, F.; Kijlstra, A. The Effects of Th17 Cytokines on the Inflammatory Mediator Production and Barrier Function of ARPE-19 Cells. PLoS ONE 2011, 6, e18139. [Google Scholar] [CrossRef] [Green Version]
- Huppert, J.; Closhen, D.; Croxford, A.; White, R.; Kulig, P.; Pietrowski, E.; Bechmann, I.; Becher, B.; Luhmann, H.J.; Waisman, A.; et al. Cellular mechanisms of IL-17-induced blood-brain barrier disruption. FASEB J. 2009, 24, 1023–1034. [Google Scholar] [CrossRef]
- Watanabe, T.; Dohgu, S.; Takata, F.; Nishioku, T.; Nakashima, A.; Futagami, K.; Yamauchi, A.; Kataoka, Y. Paracellular Barrier and Tight Junction Protein Expression in the Immortalized Brain Endothelial Cell Lines bEND.3, bEND.5 and Mouse Brain Endothelial Cell 4. Biol. Pharm. Bull. 2013, 36, 492–495. [Google Scholar] [CrossRef] [Green Version]
- Hodge, J.A.; Kawabata, T.T.; Krishnaswami, S.; Clark, J.D.; Telliez, J.-B.; Dowty, M.E.; Menon, S.; Lamba, M.; Zwillich, S. The mechanism of action of tofacitinib—An oral Janus kinase inhibitor for the treatment of rheumatoid arthritis. Clin. Exp. Rheumatol. 2016, 34, 318–328. [Google Scholar]
- Danese, S.; Grisham, M.; Hodge, J.; Telliez, J.-B. JAK inhibition using tofacitinib for inflammatory bowel disease treatment: A hub for multiple inflammatory cytokines. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, G155–G162. [Google Scholar] [CrossRef] [Green Version]
- Sandborn, W.J.; Ghosh, S.; Panes, J.; Vranic, I.; Su, C.; Rousell, S.; Niezychowski, W. Tofacitinib, an Oral Janus Kinase Inhibitor, in Active Ulcerative Colitis. New Engl. J. Med. 2012, 367, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Schön, M.P.; Erpenbeck, L. The Interleukin-23/Interleukin-17 Axis Links Adaptive and Innate Immunity in Psoriasis. Front. Immunol. 2018, 9, 1323. [Google Scholar] [CrossRef] [Green Version]
- Bunte, K.; Beikler, T. Th17 Cells and the IL-23/IL-17 Axis in the Pathogenesis of Periodontitis and Immune-Mediated Inflammatory Diseases. Int. J. Mol. Sci. 2019, 20, 3394. [Google Scholar] [CrossRef] [Green Version]
- Iwakura, Y.; Ishigame, H. The IL-23/IL-17 axis in inflammation. J. Clin. Investig. 2006, 116, 1218–1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Qian, J.; Guo, J.; Yuan, Y.-F.; Xue, K. Suppression of Experimental Autoimmune Uveoretinitis by Anti-IL-17 Antibody. Curr. Eye Res. 2009, 34, 297–303. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Huang, L.; Vergis, A.L.; Ye, H.; Bajwa, A.; Narayan, V.; Strieter, R.M.; Rosin, D.L.; Okusa, M.D. IL-17 produced by neutrophils regulates IFN-γ–mediated neutrophil migration in mouse kidney ischemia-reperfusion injury. J. Clin. Investig. 2010, 120, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Werner, J.L.; Gessner, M.A.; Lilly, L.M.; Nelson, M.P.; Metz, A.E.; Horn, D.; Dunaway, C.W.; Deshane, J.; Chaplin, D.D.; Weaver, C.T.; et al. Neutrophils Produce Interleukin 17A (IL-17A) in a Dectin-1- and IL-23-Dependent Manner during Invasive Fungal Infection. Infect. Immun. 2011, 79, 3966–3977. [Google Scholar] [CrossRef] [Green Version]
- Katayama, M.; Ohmura, K.; Yukawa, N.; Terao, C.; Hashimoto, M.; Yoshifuji, H.; Kawabata, D.; Fujii, T.; Iwakura, Y.; Mimori, T. Neutrophils Are Essential as A Source Of Il-17 In The Effector Phase Of Arthritis. PLoS ONE 2013, 8, e62231. [Google Scholar] [CrossRef]
- Akitsu, A.; Iwakura, Y. Interleukin-17-producing γδ T (γδ17) cells in inflammatory diseases. Immunology 2018, 155, 418–426. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, R.L.; Roark, C.L.; Born, W.K. IL-17-producing γδ T cells. Eur. J. Immunol. 2009, 39, 662–666. [Google Scholar] [CrossRef]
- Milovanovic, J.; Arsenijevic, A.; Stojanovic, B.; Kanjevac, T.; Arsenijevic, D.; Radosavljevic, G.; Milovanovic, M.; Arsenijevic, N. Interleukin-17 in Chronic Inflammatory Neurological Diseases. Front. Immunol. 2020, 11, 947. [Google Scholar] [CrossRef] [PubMed]
- Guedes, M.C.E.; Borrego, L.M.; Proença, R. Roles of interleukin-17 in uveitis. Indian J. Ophthalmol. 2016, 64, 628–634. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, W.; Li, Q. Increased Th1/Th17 Responses Contribute to Low-Grade Inflammation in Age-Related Macular Degeneration. Cell. Physiol. Biochem. 2017, 44, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Knickelbein, J.E.; Chan, C.-C.; Sen, H.N.; Ferris, F.L.; Nussenblatt, R.B. Inflammatory Mechanisms of Age-related Macular Degeneration. Int. Ophthalmol. Clin. 2015, 55, 63–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.-C.; Ardeljan, D. Molecular pathology of macrophages and interleukin-17 in age-related macular degeneration. Adv. Exp. Med. Biol. 2014, 801, 193–198. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Zhou, Y. Interleukin-17: The Role for Pathological Angiogenesis in Ocular Neovascular Diseases. Tohoku J. Exp. Med. 2019, 247, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Tong, Z.; Liu, W.; Yan, H.; Dong, C. Interleukin-17A deficiency ameliorates streptozotocin-induced diabetes. Immunology 2015, 146, 339–346. [Google Scholar] [CrossRef] [Green Version]
- Trunov, A.N.; Varvarinsky, E.V.; Chernykh, V.V.; Smirnov, E.V.; Chernyh, D.V. Proliferative and inflammatory factors in the vitreous of patients with proliferative diabetic retinopathy. Indian J. Ophthalmol. 2015, 63, 33–36. [Google Scholar] [CrossRef] [PubMed]
- You, T.; Bi, Y.; Li, J.; Zhang, M.; Chen, X.; Zhang, K. IL-17 induces reactive astrocytes and up-regulation of vascular endothelial growth factor (VEGF) through JAK/STAT signaling. Sci. Rep. 2017, 7, 41779. [Google Scholar] [CrossRef]
- Hu, B.; Wang, J.; Wu, X.; Chen, Y.; Yuan, W.; Chen, H. Interleukin-17 upregulates vascular endothelial growth factor by activating the JAK/STAT pathway in nucleus pulposus cells. Jt. Bone Spine 2017, 84, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Yun, J.H.; Park, S.W.; Kim, K.-J.; Bae, J.-S.; Lee, E.H.; Paek, S.H.; Kim, S.U.; Ye, S.; Kim, J.-H.; Cho, C.-H. Endothelial STAT3 Activation Increases Vascular Leakage Through Downregulating Tight Junction Proteins: Implications for Diabetic Retinopathy. J. Cell. Physiol. 2017, 232, 1123–1134. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Obasanmi, G.; Armstrong, D.; Lavery, N.-J.; Kissenpfennig, A.; Lois, N.; Xu, H. STAT3 activation in circulating myeloid-derived cells contributes to retinal microvascular dysfunction in diabetes. J. Neuroinflammation 2019, 16, 138. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Lechner, J.; Zhao, J.; Toth, L.; Hogg, R.; Silvestri, G.; Kissenpfennig, A.; Chakravarthy, U.; Xu, H. STAT3 Activation in Circulating Monocytes Contributes to Neovascular Age-Related Macular Degeneration. Curr. Mol. Med. 2016, 16, 412–423. [Google Scholar] [CrossRef]
- Li, X.; Cai, Y.; Wang, Y.-S.; Shi, Y.-Y.; Hou, W.; Xu, C.-S.; Wang, H.-Y.; Ye, Z.; Yao, L.-B.; Zhang, J. Hyperglycaemia Exacerbates Choroidal Neovascularisation in Mice via the Oxidative Stress-Induced Activation of STAT3 Signalling in RPE Cells. PLoS ONE 2012, 7, e47600. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Chen, M.; Kissenpfennig, A.; Xu, H. SOCS3 deletion in myeloid cells worsens retinal inflammation and increases angiogenesis in experimental autoimmune uveoretinitis. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3206. [Google Scholar]
- Chen, M.; Zhao, J.; Ali, I.H.; Marry, S.; Augustine, J.; Bhuckory, M.; Lynch, A.; Kissenpfennig, A.; Xu, H. Cytokine Signaling Protein 3 Deficiency in Myeloid Cells Promotes Retinal Degeneration and Angiogenesis through Arginase-1 Up-Regulation in Experimental Autoimmune Uveoretinitis. Am. J. Pathol. 2018, 188, 1007–1020. [Google Scholar] [CrossRef] [Green Version]
- Miserocchi, E.; Giuffrè, C.; Cornalba, M.; Pontikaki, I.; Cimaz, R. JAK inhibitors in refractory juvenile idiopathic arthritis-associated uveitis. Clin. Rheumatol. 2020, 39, 847–851. [Google Scholar] [CrossRef]
- Paley, M.A.; Karacal, H.; Rao, P.K.; Margolis, T.P.; Miner, J.J. Tofacitinib for refractory uveitis and scleritis. Am. J. Ophthalmol. Case Rep. 2019, 13, 53–55. [Google Scholar] [CrossRef]
- Sayoc-Becerra, A.; Krishnan, M.; Fan, S.; Jimenez, J.; Hernandez, R.; Gibson, K.; Preciado, R.; Butt, G.; McCole, D.F. The JAK-Inhibitor Tofacitinib Rescues Human Intestinal Epithelial Cells and Colonoids from Cytokine-Induced Barrier Dysfunction. Inflamm. Bowel Dis. 2020, 26, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Lauridsen, H.M.; Amezquita, R.A.; Pierce, R.; Jane-Wit, D.; Fang, C.; Pellowe, A.S.; Kirkiles-Smith, N.C.; Gonzalez, A.L.; Pober, J.S. IL-17 Promotes Neutrophil-Mediated Immunity by Activating Microvascular Pericytes and Not Endothelium. J. Immunol. 2016, 197, 2400–2408. [Google Scholar] [CrossRef] [PubMed]
- Lauridsen, H.M.; Pellowe, A.S.; Ramanathan, A.; Liu, R.; Miller-Jensen, K.; McNiff, J.M.; Pober, J.S.; Gonzalez, A.L. Tumor Necrosis Factor-α and IL-17A Activation Induces Pericyte-Mediated Basement Membrane Remodeling in Human Neutrophilic Dermatoses. Am. J. Pathol. 2017, 187, 1893–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Company, Product Number | Dilution Used |
|---|---|---|
| ZO-1 | Thermo Fisher, 61-7300 | 1:50 (IF) |
| Claudin-5 | Thermo Fisher, 34-1600 | 1:50 (IF) |
| Phospho-JAK1 (Tyr1034, Tyr1035) | Thermo Fisher, PA5-104554 | 1:50 (IF, IHC-P) |
| Albumin | Bethyl, a90-134a | 1:800 (IHC-p), 1:1000 (WB) |
| Biotinylated Isolectin B4 | Vector Labs, VEC.B-1205 | 1:50 (IHC-P) |
| Alexa Fluor® 594 AffiniPure Donkey Anti-Rabbit IgG (H+L) | Stratech, 711-585-152 | 1:300 (IF), 1:300 (IHC-p) |
| Donkey Anti-Rabbit 488 | Thermo Fisher, 34-1600 | 1:50 (IF) |
| Streptavidin, Alexa Fluor™ 594 conjugate | Thermo Fisher, S11227 | 1:300 (IHC-p) |
| Rabbit Anti-Mouse IgG H&L (HRP) | Abcam, ab6728 | 1:5000 (WB) |
| Alexa Fluor® 488 AffiniPure Donkey Anti-Goat IgG (H+L) | Stratech, 705-545-147 | 1:300 (IHC-p) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byrne, E.M.; Llorián-Salvador, M.; Tang, M.; Margariti, A.; Chen, M.; Xu, H. IL-17A Damages the Blood–Retinal Barrier through Activating the Janus Kinase 1 Pathway. Biomedicines 2021, 9, 831. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070831
Byrne EM, Llorián-Salvador M, Tang M, Margariti A, Chen M, Xu H. IL-17A Damages the Blood–Retinal Barrier through Activating the Janus Kinase 1 Pathway. Biomedicines. 2021; 9(7):831. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070831
Chicago/Turabian StyleByrne, Eimear M., María Llorián-Salvador, Miao Tang, Andriana Margariti, Mei Chen, and Heping Xu. 2021. "IL-17A Damages the Blood–Retinal Barrier through Activating the Janus Kinase 1 Pathway" Biomedicines 9, no. 7: 831. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070831