
Serum miRNA Profiling for Early PDAC Diagnosis and Prognosis: A Retrospective Study

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. RNA Extraction
2.3. MicroRNA Expression Profiling
2.4. Statistical and Functional Analysis of miRNA Expression Data
2.5. Reverse Transcription and Quantitative PCR (qRT-PCR) of miRNAs
2.6. Statistical Analysis of Data
3. Results
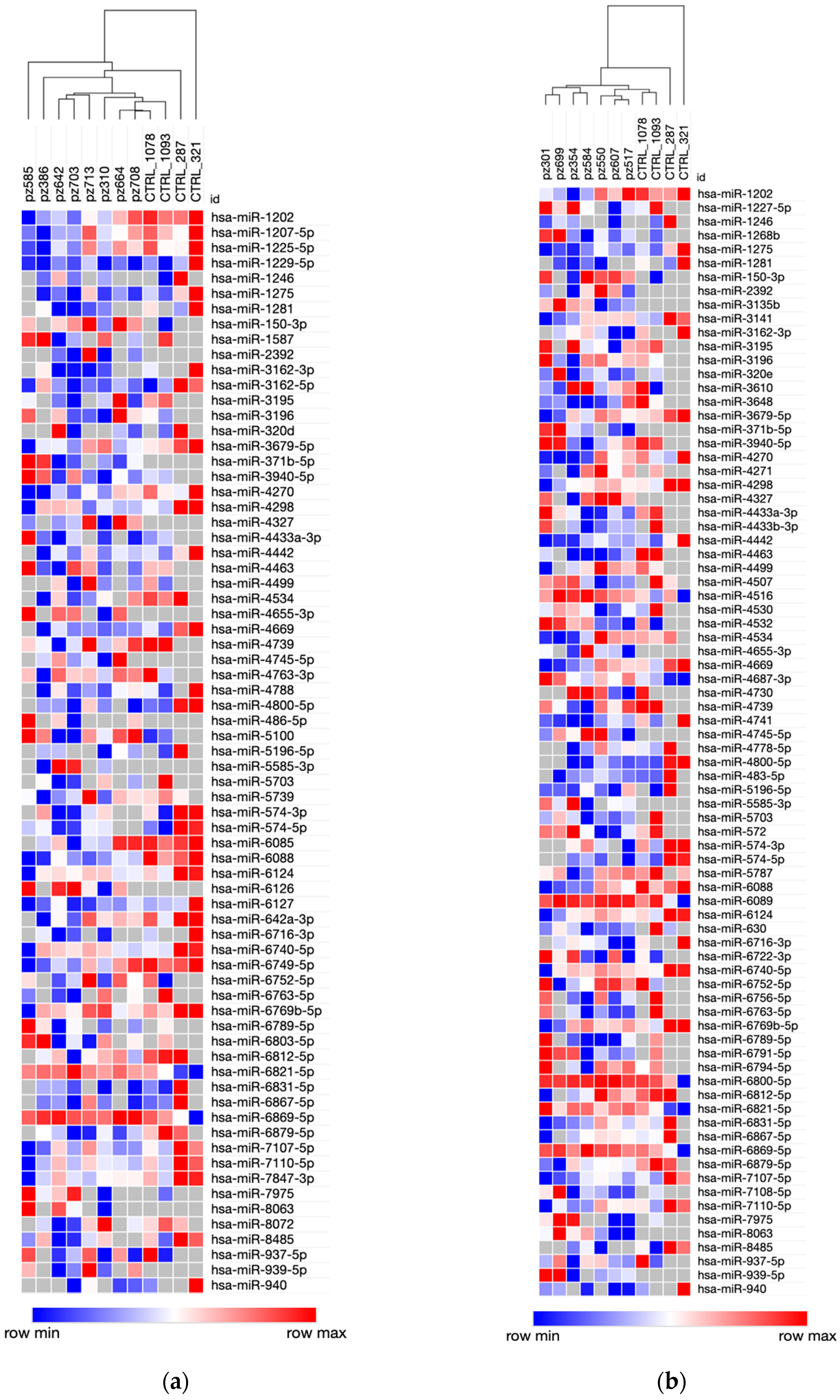
3.1. Serum miRNA Expression Signatures for Early PDAC Diagnosis
3.2. miRNA Expression Profiles and PDAC Survival Rates
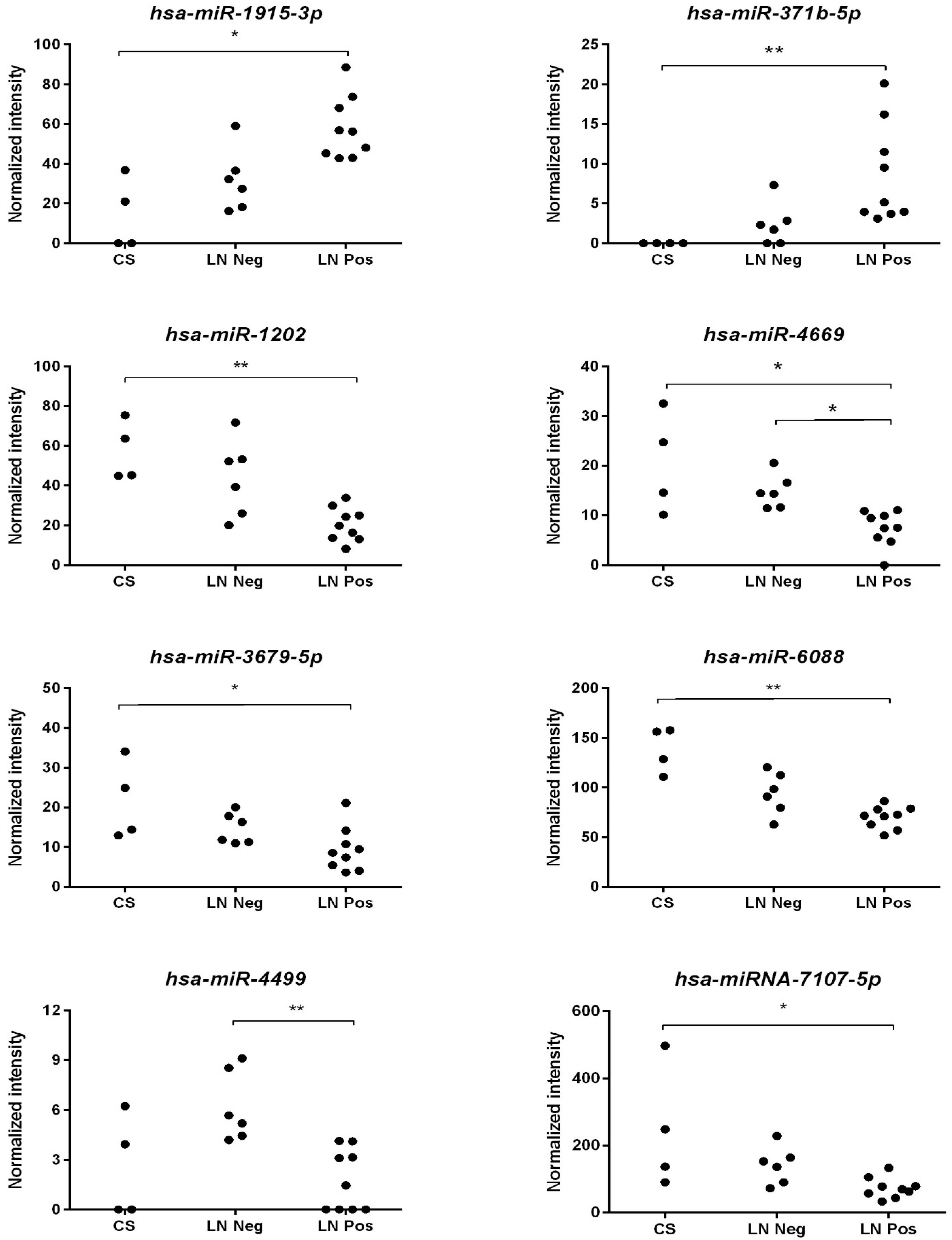
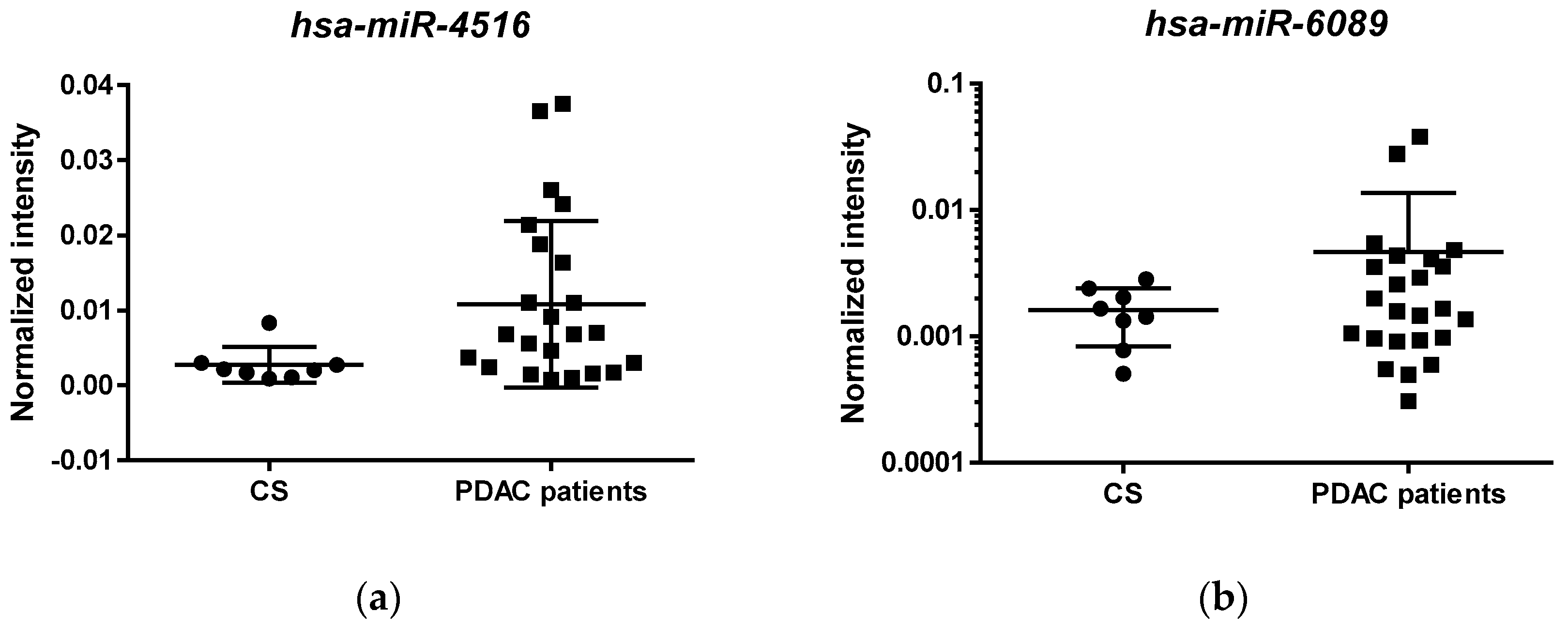
3.3. Validation of Selected miRNAs in an Independent Cohort of PDAC Patients Using qRT-PCR
3.4. Identification and Functional Analysis of Putative miRNAs Targets
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2021. CA Cancer J. Clin. 2021, 71, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Bardeesy, N.; Depinho, R.A. Pancreatic cancer biology and genetics. Nat. Rev. Cancer 2002, 2, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Ryan, D.P.; Hong, T.S.; Bardeesy, N. Pancreatic adenocarcinoma. N. Engl. J. Med. 2014, 371, 1039–1049. [Google Scholar] [CrossRef]
- Hezel, A.F.; Kimmelman, A.C.; Stanger, B.Z.; Bardeesy, N.; Depinho, R.A. Genetics and biology of pancreatic ductal adenocarcinoma. Genes Dev. 2006, 20, 1218–1249. [Google Scholar] [CrossRef] [PubMed]
- Al-Share, B.; Hammad, N.; Diab, M. Pancreatic adenocarcinoma: Molecular drivers and the role of targeted therapy. Cancer Metastasis Rev. 2021, 40, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, G.; Bungau, S.; Ilie, M.; Behl, T.; Vesa, C.M.; Brisc, C.; Bacalbasa, N.; Turi, V.; Costache, R.S.; Diaconu, C.C. Early diagnosis of pancreatic cancer: The key for survival. Diagnostics 2020, 10, 869. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, C.E.; Lin, C.C.; Mariotto, A.B.; Siegel, R.L.; Stein, K.D.; Kramer, J.L.; Alteri, R.; Robbins, A.S.; Jemal, A. Cancer treatment and survivorship statistics, 2014. CA Cancer J. Clin. 2014, 64, 252–271. [Google Scholar] [CrossRef] [PubMed]
- Cong, L.; Liu, Q.; Zhang, R.; Cui, M.; Zhang, X.; Gao, X.; Guo, J.; Dai, M.; Zhang, T.; Liao, Q.; et al. Tumor size classification of the 8th edition of TNM staging system is superior to that of the 7th edition in predicting the survival outcome of pancreatic cancer patients after radical resection and adjuvant chemotherapy. Sci. Rep. 2018, 8, 10383–10389. [Google Scholar] [CrossRef] [PubMed]
- Waddell, N.; Pajic, M.; Patch, A.-M.; Chang, D.K.; Kassahn, K.S.; Bailey, P.; Johns, A.L.; Miller, D.; Nones, K.; Quek, K.; et al. Whole genomes redefine the mutational landscape of pancreatic cancer. Nature 2015, 518, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Witkiewicz, A.K.; McMillan, E.A.; Balaji, U.; Baek, G.; Lin, W.-C.; Mansour, J.; Mollaee, M.; Wagner, K.-U.; Koduru, P.; Yopp, A.; et al. Whole-exome sequencing of pancreatic cancer defines genetic diversity and therapeutic targets. Nat. Commun. 2015, 6, 6744. [Google Scholar] [CrossRef]
- Bailey, P.; Chang, D.K.; Nones, K.; Johns, A.L.; Patch, A.-M.; Gingras, M.-C.; Miller, D.K.; Christ, A.N.; Bruxner, T.J.C.; Quinn, M.C.; et al. Genomic analyses identify molecular subtypes of pancreatic cancer. Nature 2016, 531, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Springer, S.; Wang, Y.; Molin, M.D.; Masica, D.L.; Jiao, Y.; Kinde, I.; Blackford, A.; Raman, S.P.; Wolfgang, C.L.; Tomita, T.; et al. A combination of molecular markers and clinical features improve the classification of pancreatic cysts. Gastroenterology 2015, 149, 1501–1510. [Google Scholar] [CrossRef]
- Knudsen, E.S.; O’Reilly, E.M.; Brody, J.R.; Witkiewicz, A.K. Genetic diversity of pancreatic ductal adenocarcinoma and opportunities for precision medicine. Gastroenterology 2016, 150, 48–63. [Google Scholar] [CrossRef] [PubMed]
- Oshima, M.; Okano, K.; Muraki, S.; Haba, R.; Maeba, T.; Suzuki, Y.; Yachida, S. Immunohistochemically detected expression of 3 major genes (CDKN2A/p16, TP53, and SMAD4/DPC4) strongly predicts survival in patients with resectable pancreatic cancer. Ann. Surg. 2013, 258, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Iacobuzio-Donahue, C.A.; Fu, B.; Yachida, S.; Luo, M.; Abe, H.; Henderson, C.M.; Vilardell, F.; Wang, Z.; Keller, J.W.; Banerjee, P.; et al. DPC4 gene status of the primary carcinoma correlates with patterns of failure in patients with pancreatic cancer. J. Clin. Oncol. 2009, 27, 1806–1813. [Google Scholar] [CrossRef]
- Berindan-Neagoe, I.; Monroig, P.D.C.; Pasculli, B.; Calin, G.A. MicroRNAome genome: A treasure for cancer diagnosis and therapy. CA Cancer J. Clin. 2014, 64, 311–336. [Google Scholar] [CrossRef]
- Sarwar, A.; Wang, B.; Su, Q.; Zhang, Y. MiRNAs directly targeting the key intermediates of biological pathways in pancreatic cancer. Biochem. Pharmacol. 2020, 70, 114357. [Google Scholar]
- Rong, Z.; Xu, J.; Shi, S.; Tan, Z.; Meng, Q.; Hua, J.; Liu, J.; Zhang, B.; Wang, W.; Yu, X.; et al. Circular RNA in pancreatic cancer: A novel avenue for the roles of diagnosis and treatment. Theranostics 2021, 11, 2755–2769. [Google Scholar] [CrossRef]
- Pandya, G.; Kirtonia, A.; Sethi, G.; Pandey, A.K.; Garg, M. The implication of long non-coding RNAs in the diagnosis, pathogenesis and drug resistance of pancreatic ductal adenocarcinoma and their possible therapeutic potential. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188423. [Google Scholar] [CrossRef]
- Terrinoni, A.; Calabrese, C.; Basso, D.; Aita, A.; Caporali, S.; Plebani, M.; Bernardini, S. The circulating miRNAs as diagnostic and prognostic markers. Clin. Chem. Lab. Med. 2019, 57, 932–953. [Google Scholar] [CrossRef]
- Hao, J.; Zhang, S.; Zhou, Y.; Hu, X.; Shao, C. MicroRNA 483-3p suppresses the expression of DPC4/Smad4 in pancreatic cancer. FEBS Lett. 2011, 585, 207–213. [Google Scholar] [CrossRef]
- Blahna, M.T.; Hata, A. Regulation of miRNA biogenesis as an integrated component of growth factor signaling. Curr. Opin. Cell Biol. 2013, 25, 233–240. [Google Scholar] [CrossRef]
- Yang, W.; Ju, H.-Y.; Tian, X.-F. Hsa-miR-4730 as a new and potential diagnostic and prognostic indicators for pancreatic cancer. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8801–8811. [Google Scholar]
- Hata, T.; Mizuma, M.; Masuda, K.; Chiba, K.; Ishida, M.; Ohtsuka, H.; Nakagawa, K.; Morikawa, T.; Kamei, T.; Unno, M. MicroRNA-593-3p expression in peritoneal lavage fluid as a prognostic marker for pancreatic cancer patients undergoing staging laparoscopy. Ann. Surg. Oncol. 2021, 70, 7–11. [Google Scholar]
- Wolfe, A.R.; Wald, P.; Webb, A.; Sebastian, N.; Walston, S.; Robb, R.; Chen, W.; Vedaie, M.; Dillhoff, M.; Frankel, W.L.; et al. A microRNA-based signature predicts local-regional failure and overall survival after pancreatic cancer resection. Oncotarget 2020, 11, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Huang, J.; Yang, Z.; Zhu, Y.; Zhang, Y.; Wang, J.; Yao, W. MicroRNA-221-3p is related to survival and promotes tumour progression in pancreatic cancer: A comprehensive study on functions and clinicopathological value. Cancer Cell Int. 2020, 20, 443. [Google Scholar] [CrossRef] [PubMed]
- Schultz, N.A.; Dehlendorff, C.; Jensen, B.V.; Bjerregaard, J.K.; Nielsen, K.R.; Bojesen, S.E.; Calatayud, D.; Nielsen, S.E.; Yilmaz, M.; Holländer, N.H.; et al. MicroRNA biomarkers in whole blood for detection of pancreatic cancer. JAMA 2014, 311, 392–404. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Jemal, A. The rise and fall of cancer mortality in the USA: Why does pancreatic cancer not follow the trend? Future Oncol. 2013, 9, 917–919. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ach, R.A.; Curry, B. Direct and sensitive miRNA profiling from low-input total RNA. RNA 2007, 13, 151–159. [Google Scholar] [CrossRef]
- Risso, D.; Massa, M.S.; Chiogna, M.; Romualdi, C. A modified LOESS normalization applied to microRNA arrays: A comparative evaluation. Bioinformatics 2009, 25, 2685–2691. [Google Scholar] [CrossRef]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. TM4 microarray software suite. Methods Enzymol. 2006, 411, 134–193. [Google Scholar]
- Tibshirani, R.; Hastie, T.; Narasimhan, B.; Chu, G. Diagnosis of multiple cancer types by shrunken centroids of gene expression. Proc. Natl. Acad. Sci. USA 2002, 99, 6567–6572. [Google Scholar] [CrossRef]
- Hsu, S.D.; Lin, F.M.; Wu, W.Y.; Liang, C.; Huang, W.C.; Chan, W.L.; Tsai, W.T.; Chen, G.Z.; Lee, C.J.; Chiu, C.M.; et al. miRTarBase: A database curates experimentally validated microRNA-target interactions. Nucleic Acids Res. 2011, 39, D163–D169. [Google Scholar] [CrossRef]
- Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar]
- Vega, E.A.; Kutlu, O.C.; Salehi, O.; James, D.; Alarcon, S.V.; Herrick, B.; Krishnan, S.; Kozyreva, O.; Conrad, C. Preoperative chemotherapy for pancreatic cancer improves survival and R0 rate even in early stage I. J. Gastrointest. Surg. 2020, 24, 2409–2415. [Google Scholar] [CrossRef]
- Shapiro, M.; Chen, Q.; Huang, Q.; Boosalis, V.A.; Yoon, C.H.; Saund, M.S.; Whang, E.E.; Gold, J.S. Associations of socioeconomic variables with resection, stage, and survival in patients with early-stage pancreatic cancer. JAMA Surg. 2016, 151, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Weiz, L.; Langheinz, A.; Burwinkel, B. Characterization of extracellular circulating microRNA. Nucleic Acids Res. 2011, 39, 7223–7233. [Google Scholar] [CrossRef]
- Endzeliņš, E.; Berger, A.; Melne, V.; Bajo-Santos, C.; Soboļevska, K.; Ābols, A.; Rodriguez, M.; Šantare, D.; Rudņickiha, A.; Lietuvietis, V.; et al. Detection of circulating miRNAs: Comparative analysis of extracellular vesicle-incorporated miRNAs and cell-free miRNAs in whole plasma of prostate cancer patients. BMC Cancer 2017, 17, 730. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.; Cheng, Y.Y.; Blenkiron, C.; Reid, G. Exploring mechanisms of microRNA downregulation in cancer. Microrna 2017, 6, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Mazza, T.; Gioffreda, D.; Fontana, A.; Biagini, T.; Carella, M.; Palumbo, O.; Maiello, E.; Bazzocchi, F.; Andriulli, A.; Tavano, F. Clinical significance of circulating miR-1273g-3p and miR-122-5p in pancreatic cancer. Front. Oncol. 2020, 10, 44. [Google Scholar] [CrossRef] [PubMed]
- Gungormez, C.; Gumushan Aktas, H.; Dilsiz, N.; Borazan, E. Novel miRNAs as potential biomarkers in stage II colon cancer: Microarray analysis. Mol. Biol. Rep. 2019, 46, 4175–4183. [Google Scholar] [CrossRef] [PubMed]
- Rani, S.; Gately, K.; Crown, J.; O’Byrne, K.; O’Driscoll, L. Global analysis of serum microRNAs as potential biomarkers for lung adenocarcinoma. Cancer Biol. Ther. 2013, 14, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Fornari, F.; Ferracin, M.; Trerè, D.; Milazzo, M.; Marinelli, S.; Galassi, M.; Venerandi, L.; Pollutri, D.; Patrizi, C.; Borghi, A.; et al. Circulating microRNAs, miR-939, miR-595, miR-519d and miR-494, identify cirrhotic patients with HCC. PLoS ONE 2015, 10, e0141448. [Google Scholar] [CrossRef]
- Chen, F.; Ni, X.; Chen, L.; Wang, X.; Xu, J. miR-939-3p promotes epithelial-mesenchymal transition and may be used as a prognostic marker in hepatocellular carcinoma. Oncol. Lett. 2020, 19, 2727–2732. [Google Scholar] [CrossRef] [PubMed]
- Ying, X.; Li-ya, Q.; Feng, Z.; Yin, W.; Ji-hong, L. MiR-939 promotes the proliferation of human ovarian cancer cells by repressing APC2 expression. Biomed Pharm. 2015, 71, 64–69. [Google Scholar] [CrossRef]
- Di Modica, M.; Regondi, V.; Sandri, M.; Iorio, M.V.; Zanetti, A.; Tagliabue, E.; Casalini, P.; Triulzi, T. Breast cancer-secreted miR-939 downregulates VE-cadherin and destroys the barrier function of endothelial monolayers. Cancer Lett. 2017, 384, 94–100. [Google Scholar] [CrossRef]
- Shen, Y.; Chen, G.; Gao, H.; Li, Y.; Zhuang, L.; Meng, Z.; Liu, L. miR-939-5p contributes to the migration and invasion of pancreatic cancer by targeting ARHGAP4. OncoTargets Ther. 2020, 13, 389–399. [Google Scholar] [CrossRef]
- Keller, A.; Fehlmann, T.; Backes, C.; Kern, F.; Gislefoss, R.; Langseth, H.; Rounge, T.B.; Ludwig, N.; Meese, E. Competitive learning suggests circulating miRNA profiles for cancers decades prior to diagnosis. RNA Biol. 2020, 17, 1416–1426. [Google Scholar] [CrossRef]
- Luo, G.; Jin, K.; Deng, S.; Cheng, H.; Fan, Z.; Gong, Y.; Qian, Y.; Huang, Q.; Ni, Q.; Liu, C.; et al. Roles of CA19-9 in pancreatic cancer: Biomarker, predictor and promoter. Biochim. Biophys. Acta Rev. Cancer 2020, 1875, 188409. [Google Scholar] [CrossRef]
- Pisano, A.; Griñan-Lison, C.; Farace, C.; Fiorito, G.; Fenu, G.; Jiménez, G.; Scognamillo, F.; Peña-Martin, J.; Naccarati, A.; Pröll, J.; et al. The inhibitory role of miR-486-5p on CSC phenotype has diagnostic and prognostic potential in colorectal cancer. Cancers 2020, 12, 3432. [Google Scholar] [CrossRef] [PubMed]
- Pederzoli, P.; Bassi, C.; Falconi, M.; Pedrazzoli, S. Does the extent of lymphatic resection affect the outcome in pancreatic cancer? Digestion 1997, 58, 536–541. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Yan, Z.; Yang, H.; Ni, H.; Zhang, L.; Wang, Y. Clinical value of combined detection of miR-1202 and miR-195 in early diagnosis of cervical cancer. Oncol. Lett. 2019, 17, 3387–3391. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Zhang, P.; Tan, Z.; Xu, J. MiR-1202 suppresses hepatocellular carcinoma cells migration and invasion by targeting cyclin dependent kinase 14. Biomed Pharm. 2017, 96, 1246–1252. [Google Scholar] [CrossRef]
- Wang, Y.N.; Chen, Z.H.; Chen, W.C. Novel circulating microRNAs expression profile in colon cancer: A pilot study. Eur. J. Med. Res. 2017, 22, 51. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, M.; Cascinu, S.; Kleeff, J.; Labianca, R.; Löhr, J.M.; Neoptolemos, J.; Real, F.X.; Van Laethem, J.L.; Heinemann, V. Addressing the challenges of pancreatic cancer: Future directions for improving outcomes. Pancreatology 2015, 15, 8–18. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Patient Nr. | Age | Gender | G | T | N | M | Stage | Survival (months) | Diabetes | CA 19-9kU/L | Total Bilirubin μmol/L | P-Glucose mmol/L | Survival |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 713 | 57 | M | 2 | 2 | 0 | 0 | IB | 113 | DM (C) | 131 | 10 | 5.0 | L |
| 708 | 70 | M | 1 | 3 | 0 | 0 | IIA | 86 | DM (P) | 1 | 8 | 10.4 | L |
| 386 | 59 | M | 2 | 3 | 1 | 0 | IIB | 75 | RGT (C) | 20 | 92 | 6.1 | L |
| 664 | 79 | F | 2 | 3 | 1 | 0 | IIB | 56 | RGT (C) | na | 39 | 4.7 | L |
| 642 | 79 | M | 2 | 3 | 0 | 0 | IIA | 45 | RGT (C) | 92 | 39 | 7.4 | L |
| 310 | 78 | F | 2 | 3 | 1 | 0 | IIB | 29 | DM (P) | 6969 | 135 | 8.8 | L |
| 703 | 72 | F | 2 | 3 | 1 | 0 | IIB | 25 | DM (P) | 5309 | 12 | 8.9 | L |
| 585 | 63 | M | 2 | 3 | 1 | 0 | IIB | 23 | DM (P) | 435 | 13 | 8.2 | L |
| 584 | 58 | F | 3 | 3 | 1 | 0 | IIB | 13 | RGT (C) | 106 | 81 | 5.2 | S |
| 354 | 63 | M | 2 | 4 | 1 | 0 | III | 11 | DM (C) | 519 | 94 | 5.9 | S |
| 301 | 65 | M | 3 | 3 | 1 | 0 | IIB | 11 | RGT (C) | 48 | 65 | 7.1 | S |
| 699 | 67 | F | 2 | 3 | 1 | 0 | IIB | 9 | DM (C) | 30 | 22 | 6.4 | S |
| 517 | 61 | F | 3 | 4 | 0 | 0 | III | 8 | DM (C) | 212 | 9 | 6.2 | S |
| 607 | 68 | M | 3 | 3 | 0 | 0 | IIA | 6 | RGT (C) | 29 | 7 | 5.3 | S |
| 550 | 84 | F | 2 | 3 | 0 | 0 | IIA | 2 | DM (P) | 105 | 39 | 8.1 | S |
| ROC | Asymptotic Normal | Increase/Decrease | ||||
|---|---|---|---|---|---|---|
| Obs | AUC | SE | 95% Conf. Interval | |||
| hsa-miR-7110-5p | 19 | 0.8167 | 0.1360 | 0.55014 | 1.00000 | Decrease |
| hsa-miR-3135-b | 19 | 0.8333 | 0.0630 | 0.70987 | 0.95680 | Increase |
| hsa-miR-4669 | 19 | 0.8333 | 0.1346 | 0.56959 | 1.00000 | Decrease |
| hsa-miR-7107-5p | 19 | 0.8333 | 0.1225 | 0.59329 | 1.00000 | Decrease |
| hsa-miR-574-3p | 19 | 0.8333 | 0.1225 | 0.59329 | 1.00000 | Decrease |
| hsa-miR-1275 | 19 | 0.8333 | 0.1188 | 0.60049 | 1.00000 | Decrease |
| hsa-miR-4466 | 19 | 0.8500 | 0.1351 | 0.58529 | 1.00000 | Increase |
| hsa-miR-3679-5p | 19 | 0.8500 | 0.1055 | 0.64316 | 1.00000 | Decrease |
| hsa-miR-2392 | 19 | 0.8667 | 0.0591 | 0.75085 | 0.98249 | Increase |
| hsa-miR-4655-3p | 19 | 0.8667 | 0.0591 | 0.75085 | 0.98249 | Increase |
| hsa-miR-6089 | 19 | 0.8667 | 0.1196 | 0.63230 | 1.00000 | Increase |
| hsa-miR-5100 | 19 | 0.8667 | 0.0970 | 0.67646 | 1.00000 | Increase |
| hsa-miR-6749-5p | 19 | 0.8667 | 0.0858 | 0.69858 | 1.00000 | Decrease |
| hsa-miR-4687-3p | 19 | 0.8833 | 0.0922 | 0.70258 | 1.00000 | Increase |
| hsa-miR-1915-3p | 19 | 0.8833 | 0.0922 | 0.70258 | 1.00000 | Increase |
| hsa-miR-6125 | 19 | 0.8833 | 0.1043 | 0.67896 | 1.00000 | Increase |
| hsa-miR-1202 | 19 | 0.8833 | 0.0812 | 0.72426 | 1.00000 | Decrease |
| hsa-miR-8485 | 19 | 0.8833 | 0.1043 | 0.67896 | 1.00000 | Decrease |
| hsa-miR-6126 | 19 | 0.9000 | 0.0535 | 0.79524 | 1.00000 | Increase |
| hsa-miR-939-5p | 19 | 0.9000 | 0.0535 | 0.79524 | 1.00000 | Increase |
| hsa-miR-6800-5p | 19 | 0.9000 | 0.0796 | 0.74399 | 1.00000 | Increase |
| hsa-miR-4516 | 19 | 0.9167 | 0.0684 | 0.78255 | 1.00000 | Increase |
| hsa-miR-6869-5p | 19 | 0.9167 | 0.0891 | 0.74206 | 1.00000 | Increase |
| hsa-miR-6850-5p | 19 | 0.9167 | 0.0746 | 0.77041 | 1.00000 | Increase |
| hsa-miR-4327 | 19 | 0.9333 | 0.0454 | 0.84430 | 1.00000 | Increase |
| hsa-miR-371b-5p | 19 | 0.9333 | 0.0454 | 0.84430 | 1.00000 | Increase |
| hsa-miR-6821-5p | 19 | 0.9667 | 0.0403 | 0.88761 | 1.00000 | Increase |
| χ2 | p | |
|---|---|---|
| hsa-miR-1202 | 10.861 | 0.0044 |
| hsa-miR-3679-5p | 8.061 | 0.0178 |
| hsa-miR-6088 | 11.782 | 0.0028 |
| hsa-miR-6791-5p | 6.027 | 0.0491 |
| hsa-miR-1915-3p | 10.861 | 0.0044 |
| hsa-miR-371b-5p | 11.982 | 0.0025 |
| hsa-miR-4669 | 12.430 | 0.0020 |
| hsa-miR-4499 | 9.682 | 0.0079 |
| hsa-miR-4442 | 8.219 | 0.0164 |
| hsa-miR-7107-5p | 9.482 | 0.0087 |
| hsa-miR-4800-5p | 6.685 | 0.0353 |
| Predictors | HR | SE | z | p | 95% CI | |
|---|---|---|---|---|---|---|
| Sex | 11.2116 | 15.25747 | 1.78 | 0.076 | 0.7785689 | 161.4499 |
| Age | 1.088652 | 0.0546704 | 1.69 | 0.091 | 0.9866048 | 1.201255 |
| hsa-miR-4516 | 1.59 × 10−46 | 7.36 × 10−45 | −2.28 | 0.023 | 6.58 × 10−86 | 3.85 × 10−07 |
| hsa-miR-6089 | 2.91 × 1034 | 1.24 × 1036 | 1.86 | 0.063 | 0.0129917 | 6.53 × 1070 |
| CA 19-9 | 1.000101 | 0.0000555 | 1.81 | 0.070 | 0.9999919 | 1.000209 |
| Hemoglobin | 0.9515861 | 0.0202986 | −2.33 | 0.020 | 0.9126218 | 0.9922141 |
| Stage | 20.64211 | 22.01492 | 2.84 | 0.005 | 2.552365 | 166.9418 |
| Entrez ID | Symbol | Gene Name | miRNA | Log2 (PDAC Patients/Control) | Experimental Evidence |
|---|---|---|---|---|---|
| 208 | AKT2 | AKT serine/threonine kinase 2 | hsa-miR-2861 | 0.60 | Luciferase reporter assay; Western blot |
| 9459 | ARHGEF6 | Rac/Cdc42 guanine nucleotide exchange factor 6 | hsa-miR-6127 | −1.90 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-6127 | −1.90 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-7110-5p | −1.11 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-4739 | −0.98 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-5787 | −0.74 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-6879-5p | −0.66 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-6756-5p | −0.30 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-6752-5p | 0.02 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-6791-5p | 0.09 | PAR-CLIP |
| 598 | BCL2L1 | BCL2 like 1 | hsa-miR-371b-5p | null | PAR-CLIP |
| 598 | BCL2L2 | BCL2 like 1 | hsa-miR-630 | −1.34 | Immunoblot;Luciferase reporter assay;qRT-PCR |
| 595 | CCND1 | cyclin D1 | hsa-miR-574-5p | −2.63 | PAR-CLIP |
| 595 | CCND1 | cyclin D1 | hsa-miR-3648 | −1.98 | PAR-CLIP |
| 595 | CCND1 | cyclin D1 | hsa-miR-5196-5p | −1.69 | PAR-CLIP |
| 595 | CCND1 | cyclin D1 | hsa-miR-7107-5p | −1.27 | PAR-CLIP |
| 595 | CCND1 | cyclin D1 | hsa-miR-6867-5p | −0.84 | PAR-CLIP |
| 595 | CCND1 | cyclin D1 | hsa-miR-3940-5p | 0.01 | qRT-PCR;Western blot |
| 595 | CCND1 | cyclin D1 | hsa-miR-2861 | 0.60 | Luciferase reporter assay;Western blot |
| 595 | CCND1 | cyclin D1 | hsa-miR-2392 | null | PAR-CLIP |
| 1019 | CDK4 | cyclin dependent kinase 4 | hsa-miR-3135b | null | HITS-CLIP |
| 1021 | CDK6 | cyclin dependent kinase 6 | hsa-miR-6716-3p | −2.51 | PAR-CLIP |
| 1021 | CDK6 | cyclin dependent kinase 6 | hsa-miR-8485 | −1.81 | HITS-CLIP |
| 1021 | CDK6 | cyclin dependent kinase 6 | hsa-miR-7847-3p | −0.82 | PAR-CLIP |
| 1021 | CDK6 | cyclin dependent kinase 6 | hsa-miR-4534 | −0.48 | PAR-CLIP |
| 1021 | CDK6 | cyclin dependent kinase 6 | hsa-miR-5739 | −0.14 | PAR-CLIP |
| 1021 | CDK6 | cyclin dependent kinase 6 | hsa-miR-371b-5p | null | PAR-CLIP |
| 1869 | E2F1 | E2F transcription factor 1 | hsa-miR-940 | −3.46 | PAR-CLIP |
| 1869 | E2F1 | E2F transcription factor 1 | hsa-miR-6763-5p | −1.09 | PAR-CLIP |
| 1869 | E2F1 | E2F transcription factor 1 | hsa-miR-939-5p | null | PAR-CLIP |
| 1870 | E2F2 | E2F transcription factor 2 | hsa-miR-4669 | −0.88 | HITS-CLIP |
| 1870 | E2F2 | E2F transcription factor 2 | hsa-miR-4433b-3p | −0.40 | PAR-CLIP |
| 1871 | E2F3 | E2F transcription factor 3 | hsa-miR-6124 | −0.94 | PAR-CLIP |
| 1871 | E2F3 | E2F transcription factor 3 | hsa-miR-6867-5p | −0.84 | PAR-CLIP |
| 1871 | E2F3 | E2F transcription factor 3 | hsa-miR-4778-5p | −0.33 | PAR-CLIP |
| 1871 | E2F3 | E2F transcription factor 3 | hsa-miR-4271 | −0.19 | PAR-CLIP |
| 1950 | EGF | epidermal growth factor | hsa-miR-940 | −3.46 | PAR-CLIP |
| 1950 | EGF | epidermal growth factor | hsa-miR-4433a-3p | −0.17 | PAR-CLIP |
| 1956 | EGFR | epidermal growth factor receptor | hsa-miR-574-3p | −1.57 | Luciferase reporter assay;Microarray;qRT-PCR;Western blot |
| 1956 | EGFR | epidermal growth factor receptor | hsa-miR-2861 | 0.60 | Luciferase reporter assay;Western blot |
| 2064 | ERBB2 | erb-b2 receptor tyrosine kinase 2 | hsa-miR-4270 | −0.97 | HITS-CLIP |
| 8517 | IKBKG | inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase gamma | hsa-miR-6127 | −1.90 | PAR-CLIP |
| 8517 | IKBKG | inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase gamma | hsa-miR-8485 | −1.81 | HITS-CLIP |
| 8517 | IKBKG | inhibitor of kappa light polypeptide gene enhancer in B-cells, kinase gamma | hsa-miR-4739 | −0.98 | HITS-CLIP |
| 3845 | KRAS | KRAS proto-oncogene, GTPase | hsa-miR-671-5p | −0.30 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-6763-5p | −1.09 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-6831-5p | −0.91 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-6088 | −0.80 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-4271 | −0.19 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-4433a-3p | −0.17 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-6869-5p | 1.18 | PAR-CLIP |
| 5594 | MAPK1 | mitogen-activated protein kinase 1 | hsa-miR-3135b | null | HITS-CLIP |
| 5602 | MAPK10 | mitogen-activated protein kinase 10 | hsa-miR-574-5p | −2.63 | HITS-CLIP |
| 5602 | MAPK10 | mitogen-activated protein kinase 10 | hsa-miR-6867-5p | −0.84 | HITS-CLIP |
| 5599 | MAPK8 | mitogen-activated protein kinase 8 | hsa-miR-371b-5p | null | PAR-CLIP |
| 5293 | PIK3CD | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta | hsa-miR-4433a-3p | −0.17 | PAR-CLIP |
| 5294 | PIK3CG | phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit gamma | hsa-miR-8485 | −1.81 | PAR-CLIP |
| 5295 | PIK3R1 | phosphoinositide-3-kinase regulatory subunit 1 | hsa-miR-8485 | −1.81 | PAR-CLIP |
| 5295 | PIK3R1 | phosphoinositide-3-kinase regulatory subunit 1 | hsa-miR-1202 | −0.94 | PAR-CLIP |
| 5296 | PIK3R2 | phosphoinositide-3-kinase regulatory subunit 2 | hsa-miR-3135b | null | PAR-CLIP |
| 5879 | RAC1 | ras-related C3 botulinum toxin substrate 1 | hsa-miR-574-3p | −1.57 | Luciferase reporter assay;Microarray;qRT-PCR;Western blot |
| 5879 | RAC1 | ras-related C3 botulinum toxin substrate 1 | hsa-miR-6763-5p | −1.09 | PAR-CLIP |
| 5879 | RAC1 | ras-related C3 botulinum toxin substrate 1 | hsa-miR-6124 | −0.94 | PAR-CLIP |
| 5879 | RAC1 | ras-related C3 botulinum toxin substrate 1 | hsa-miR-939-5p | null | PAR-CLIP |
| 5881 | RAC3 | ras-related C3 botulinum toxin substrate 3 | hsa-miR-5703 | −1.70 | PAR-CLIP |
| 5881 | RAC3 | ras-related C3 botulinum toxin substrate 3 | hsa-miR-4516 | 1.51 | PAR-CLIP |
| 5888 | RAD51 | RAD51 recombinase | hsa-miR-940 | −3.46 | PAR-CLIP |
| 5888 | RAD51 | RAD51 recombinase | hsa-miR-7847-3p | −0.82 | PAR-CLIP |
| 5888 | RAD51 | RAD51 recombinase | hsa-miR-1915-3p | 0.72 | HITS-CLIP |
| 5888 | RAD51 | RAD51 recombinase | hsa-miR-7975 | 1.09 | HITS-CLIP |
| 5888 | RAD51 | RAD51 recombinase | hsa-miR-371b-5p | null | HITS-CLIP |
| 5894 | RAF1 | Raf-1 proto-oncogene, serine/threonine kinase | hsa-miR-4534 | −0.48 | PAR-CLIP |
| 5894 | RAF1 | Raf-1 proto-oncogene, serine/threonine kinase | hsa-miR-6789-5p | −0.27 | PAR-CLIP |
| 5970 | RELA | RELA proto-oncogene, NF-kB subunit | hsa-miR-3162-3p | −2.72 | PAR-CLIP |
| 5970 | RELA | RELA proto-oncogene, NF-kB subunit | hsa-miR-4534 | −0.48 | PAR-CLIP |
| 4087 | SMAD2 | SMAD family member 2 | hsa-miR-8485 | −1.81 | HITS-CLIP |
| 4087 | SMAD2 | SMAD family member 2 | hsa-miR-937-5p | −0.38 | HITS-CLIP |
| 4089 | SMAD4 | SMAD family member 4 | hsa-miR-574-5p | −2.63 | HITS-CLIP |
| 4089 | SMAD4 | SMAD family member 4 | hsa-miR-574-3p | −1.57 | Luciferase reporter assay;qRT-PCR;Western blot |
| 4089 | SMAD4 | SMAD family member 4 | hsa-miR-6867-5p | −0.84 | HITS-CLIP |
| 4089 | SMAD4 | SMAD family member 4 | hsa-miR-371b-5p | null | PAR-CLIP |
| 6774 | STAT3 | signal transducer and activator of transcription 3 | hsa-miR-4270 | −0.97 | PAR-CLIP |
| 6774 | STAT3 | signal transducer and activator of transcription 3 | hsa-miR-4516 | 1.51 | Luciferase reporter assay;Microarray;qRT-PCR;Western blot |
| 7040 | TGFB1 | transforming growth factor beta 1 | hsa-miR-574-3p | −1.57 | Luciferase reporter assay;qRT-PCR;Western blot |
| 7046 | TGFBR1 | transforming growth factor beta receptor 1 | hsa-miR-6831-5p | −0.91 | HITS-CLIP |
| 7048 | TGFBR2 | transforming growth factor beta receptor 2 | hsa-miR-940 | −3.46 | PAR-CLIP |
| 7048 | TGFBR2 | transforming growth factor beta receptor 2 | hsa-miR-574-5p | −2.63 | HITS-CLIP |
| 7048 | TGFBR2 | transforming growth factor beta receptor 2 | hsa-miR-630 | −1.34 | Microarray |
| 7048 | TGFBR2 | transforming growth factor beta receptor 2 | hsa-miR-6867-5p | −0.84 | HITS-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-6127 | −1.90 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-5703 | −1.70 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-7110-5p | −1.11 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-937-5p | −0.38 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-6756-5p | −0.30 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-4271 | −0.19 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-6752-5p | 0.02 | PAR-CLIP |
| 7157 | TP53 | tumor protein p53 | hsa-miR-4516 | 1.51 | PAR-CLIP |
| 7422 | VEGFA | vascular endothelial growth factor A | hsa-miR-8485 | −1.81 | PAR-CLIP;HITS-CLIP |
| 7422 | VEGFA | vascular endothelial growth factor A | hsa-miR-6769b-5p | −0.70 | PAR-CLIP |
| 7422 | VEGFA | vascular endothelial growth factor A | hsa-miR-6756-5p | −0.30 | PAR-CLIP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aita, A.; Millino, C.; Sperti, C.; Pacchioni, B.; Plebani, M.; De Pittà, C.; Basso, D. Serum miRNA Profiling for Early PDAC Diagnosis and Prognosis: A Retrospective Study. Biomedicines 2021, 9, 845. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070845
Aita A, Millino C, Sperti C, Pacchioni B, Plebani M, De Pittà C, Basso D. Serum miRNA Profiling for Early PDAC Diagnosis and Prognosis: A Retrospective Study. Biomedicines. 2021; 9(7):845. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070845
Chicago/Turabian StyleAita, Ada, Caterina Millino, Cosimo Sperti, Beniamina Pacchioni, Mario Plebani, Cristiano De Pittà, and Daniela Basso. 2021. "Serum miRNA Profiling for Early PDAC Diagnosis and Prognosis: A Retrospective Study" Biomedicines 9, no. 7: 845. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9070845