
Lobeglitazone Exerts Anti-Inflammatory Effect in Lipopolysaccharide-Induced Bone-Marrow Derived Macrophages

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of LOBE and LPS
2.2. Experimental Group
2.3. Cell Culture
2.4. BMDM Preparation
2.5. Cell Cytotoxicity Test
2.6. NO Quantification
2.7. RT-qPCR
2.8. Western Blot
2.9. Statistical Analysis
3. Results
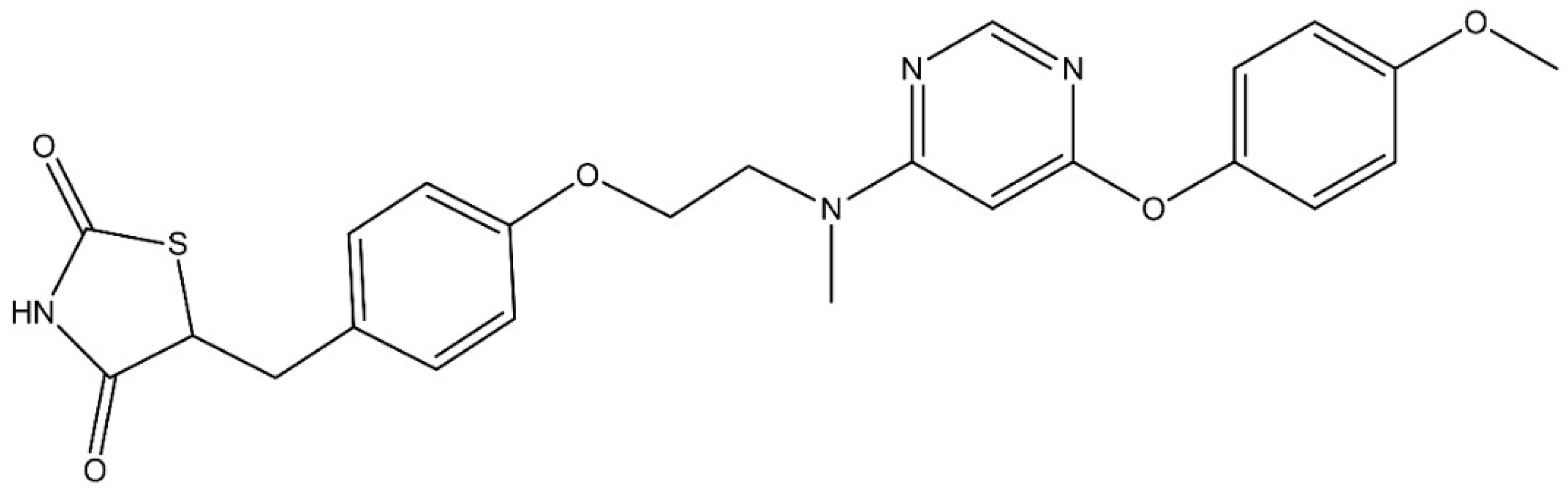
3.1. Molecular Structure of LOBE and Cytotoxicity of LOBE on BMDMs
3.2. Inhibitory Effect of NO Production by LOBE
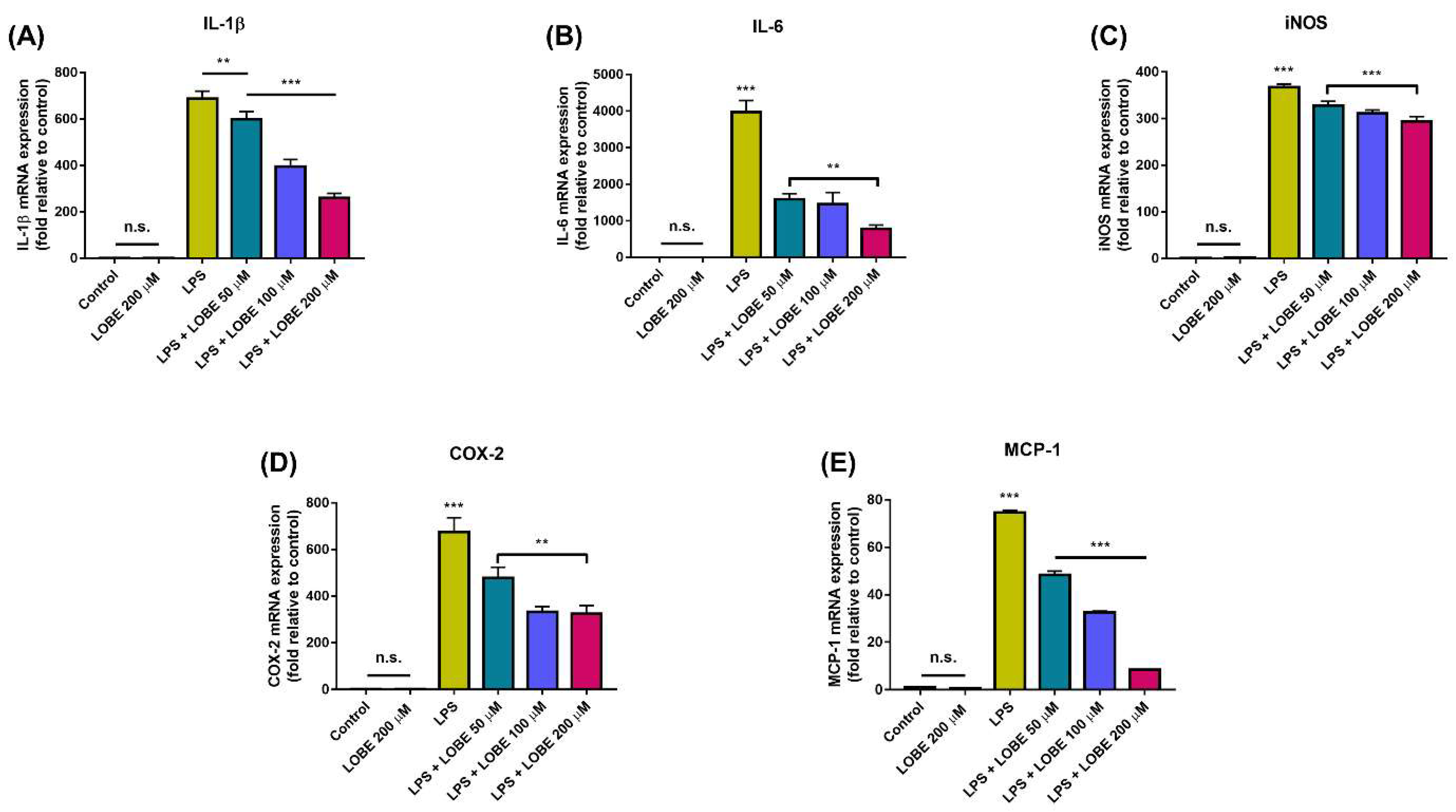
3.3. LOBE Decreased Pro-Inflammatory Gene Expressions in LPS-Induced BMDMs
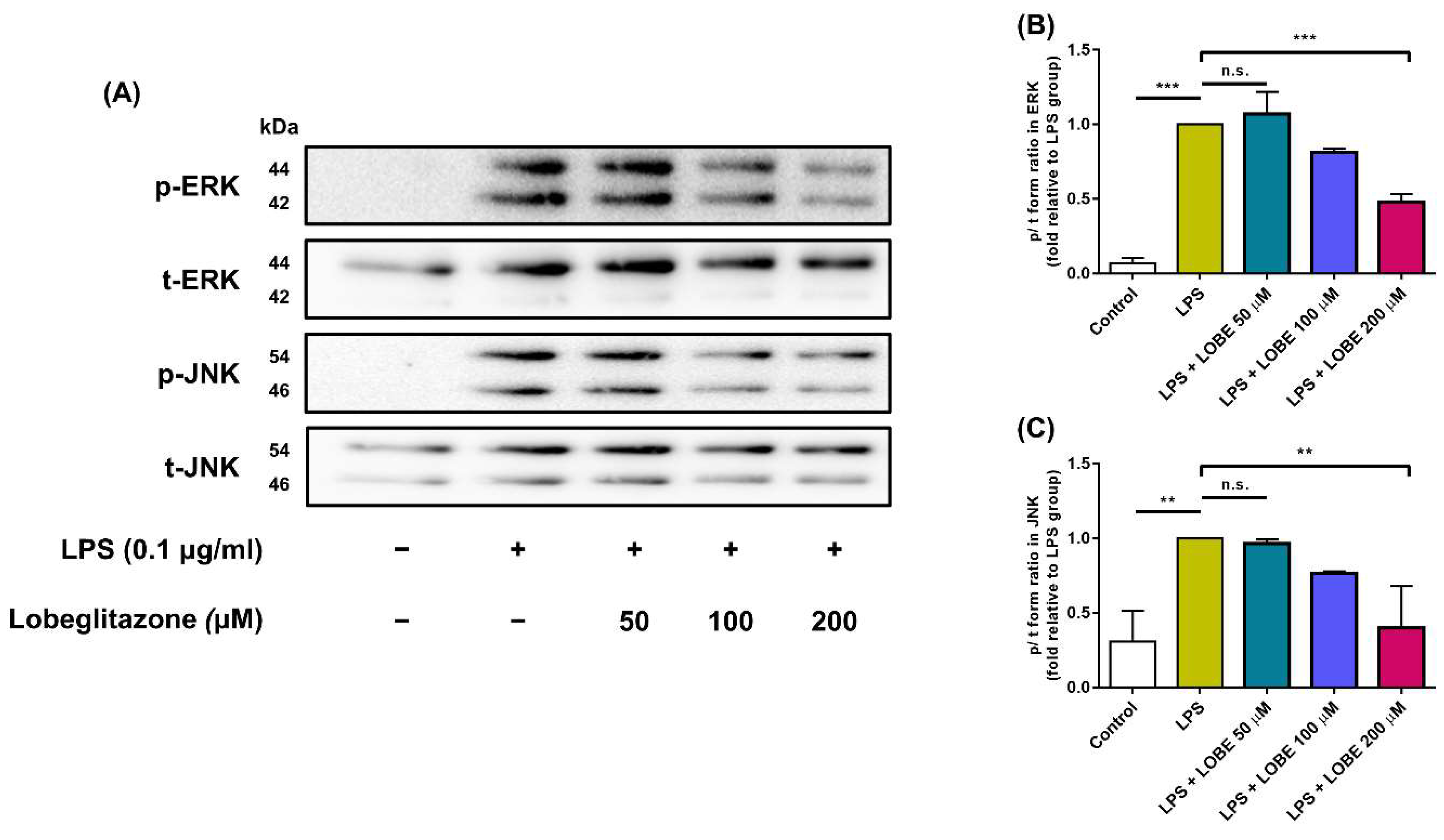
3.4. LOBE Acts through MAPK Signaling Pathway by Decreasing Phosphorylation of ERK and JNK Signaling
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Dinarello, C.A. Biologic basis for interleukin-1 in disease. Blood 1996, 87, 2095–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef] [PubMed]
- Giroux, M.; Descoteaux, A. Cyclooxygenase-2 expression in macrophages: Modulation by protein kinase C-alpha. J. Immunol. 2000, 165, 3985–3991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte chemoattractant protein-1 (MCP-1): An overview. J. Interferon Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef]
- Kushner, I. Regulation of the acute phase response by cytokines. Perspect. Biol. Med. 1993, 36, 611–622. [Google Scholar] [CrossRef]
- Ehses, J.A.; Perren, A.; Eppler, E.; Ribaux, P.; Pospisilik, J.A.; Maor-Cahn, R.; Gueripel, X.; Ellingsgaard, H.; Schneider, M.K.; Biollaz, G.; et al. Increased number of islet-associated macrophages in type 2 diabetes. Diabetes 2007, 56, 2356–2370. [Google Scholar] [CrossRef] [Green Version]
- Hansson, G.K.; Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef]
- David, S.; Kroner, A. Repertoire of microglial and macrophage responses after spinal cord injury. Nat. Rev. Neurosci. 2011, 12, 388–399. [Google Scholar] [CrossRef]
- Wyss-Coray, T.; Mucke, L. Ibuprofen, inflammation and Alzheimer disease. Nat. Med. 2000, 6, 973–974. [Google Scholar] [CrossRef]
- Klein, A.; Eliakim, R. Non Steroidal Anti-Inflammatory Drugs and Inflammatory Bowel Disease. Pharmaceuticals 2010, 3, 1084–1092. [Google Scholar] [CrossRef]
- Rudmann, D.G. On-target and off-target-based toxicologic effects. Toxicol. Pathol. 2013, 41, 310–314. [Google Scholar] [CrossRef]
- Bae, J.; Park, T.; Kim, H.; Lee, M.; Cha, B.S. Lobeglitazone: A Novel Thiazolidinedione for the Management of Type 2 Diabetes Mellitus. Diabetes Metab. J. 2021, 45, 326–336. [Google Scholar] [CrossRef] [PubMed]
- Yki-Jarvinen, H. Thiazolidinediones. N. Engl. J. Med. 2004, 351, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.H.; Jin, Z.; Yi, C.O.; Oh, J.; Jeong, E.A.; Lee, J.Y.; Park, K.A.; Kim, K.E.; Lee, J.E.; Kim, H.J.; et al. Effects of lobeglitazone on insulin resistance and hepatic steatosis in high-fat diet-fed mice. PLoS ONE 2018, 13, e0200336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.M.; Park, C.Y.; Cho, Y.M.; Ku, B.J.; Ahn, C.W.; Cha, B.S.; Min, K.W.; Sung, Y.A.; Baik, S.H.; Lee, K.W.; et al. Lobeglitazone and pioglitazone as add-ons to metformin for patients with type 2 diabetes: A 24-week, multicentre, randomized, double-blind, parallel-group, active-controlled, phase III clinical trial with a 28-week extension. Diabetes Obes. Metab. 2015, 17, 599–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.; Lee, Y.H.; Yun, M.R.; Lee, J.Y.; Shin, E.G.; Lee, B.W.; Kang, E.S.; Cha, B.S. Effects of lobeglitazone, a novel thiazolidinedione, on adipose tissue remodeling and brown and beige adipose tissue development in db/db mice. Int. J. Obes. 2018, 42, 542–551. [Google Scholar] [CrossRef]
- Kim, S.G.; Kim, D.M.; Woo, J.T.; Jang, H.C.; Chung, C.H.; Ko, K.S.; Park, J.H.; Park, Y.S.; Kim, S.J.; Choi, D.S. Efficacy and safety of lobeglitazone monotherapy in patients with type 2 diabetes mellitus over 24-weeks: A multicenter, randomized, double-blind, parallel-group, placebo controlled trial. PLoS ONE 2014, 9, e92843. [Google Scholar] [CrossRef]
- Lim, S.; Lee, K.S.; Lee, J.E.; Park, H.S.; Kim, K.M.; Moon, J.H.; Choi, S.H.; Park, K.S.; Kim, Y.B.; Jang, H.C. Effect of a new PPAR-gamma agonist, lobeglitazone, on neointimal formation after balloon injury in rats and the development of atherosclerosis. Atherosclerosis 2015, 243, 107–119. [Google Scholar] [CrossRef]
- Kim, S.J.; Ko, W.K.; Jo, M.J.; Arai, Y.; Choi, H.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effect of Tauroursodeoxycholic acid in RAW 264.7 macrophages, Bone marrow-derived macrophages, BV2 microglial cells, and spinal cord injury. Sci. Rep. 2018, 8, 3176. [Google Scholar] [CrossRef]
- Ko, W.K.; Lee, S.H.; Kim, S.J.; Jo, M.J.; Kumar, H.; Han, I.B.; Sohn, S. Anti-inflammatory effects of ursodeoxycholic acid by lipopolysaccharide-stimulated inflammatory responses in RAW 264.7 macrophages. PLoS ONE 2017, 12, e0180673. [Google Scholar] [CrossRef] [Green Version]
- Weischenfeldt, J.; Porse, B. Bone Marrow-Derived Macrophages (BMM): Isolation and Applications. Cold Spring Harb. Protoc. 2008, 2008, pdb–prot5080. [Google Scholar] [CrossRef] [Green Version]
- Toda, G.; Yamauchi, T.; Kadowaki, T.; Ueki, K. Preparation and culture of bone marrow-derived macrophages from mice for functional analysis. STAR Protoc. 2021, 2, 100246. [Google Scholar] [CrossRef]
- Lee, D.; Ko, W.K.; Kim, S.J.; Han, I.B.; Hong, J.B.; Sheen, S.H.; Sohn, S. Inhibitory Effects of Gold and Silver Nanoparticles on the Differentiation into Osteoclasts In Vitro. Pharmaceutics 2021, 13, 462. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Han, G.H.; Kim, S.J.; Ko, W.K.; Lee, D.; Lee, J.S.; Nah, H.; Han, I.B.; Sohn, S. Injectable Hydrogel Containing Tauroursodeoxycholic Acid for Anti-neuroinflammatory Therapy After Spinal Cord Injury in Rats. Mol. Neurobiol. 2020, 57, 4007–4017. [Google Scholar] [CrossRef]
- Bagasra, O.; Michaels, F.H.; Zheng, Y.M.; Bobroski, L.E.; Spitsin, S.V.; Fu, Z.F.; Tawadros, R.; Koprowski, H. Activation of the inducible form of nitric oxide synthase in the brains of patients with multiple sclerosis. Proc. Natl. Acad. Sci. USA 1995, 92, 12041–12045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuhlencordt, P.J.; Chen, J.; Han, F.; Astern, J.; Huang, P.L. Genetic deficiency of inducible nitric oxide synthase reduces atherosclerosis and lowers plasma lipid peroxides in apolipoprotein E-knockout mice. Circulation 2001, 103, 3099–3104. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R. Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs. Nat. New Biol. 1971, 231, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Minghetti, L. Cyclooxygenase-2 (COX-2) in inflammatory and degenerative brain diseases. J. Neuropathol. Exp. Neurol. 2004, 63, 901–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol. Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward (5′ → 3′) | Reverse (5′ → 3′) |
|---|---|---|
| IL-1β | CCCTGCAGCTGGAGAGTGTGG | TGTGCTCTGCTTCAGAGGTGCT |
| IL-6 | TTGTTGCTGTGGAGAAGCTGT | AACGTCACACACCAGCAGGTT |
| iNOS | CGGAGGAGAAGTGGGGTTTAGGAT | TGGGAGGCACTTGCGTTGATGG |
| COX-2 | GACCAGATAAGGGCAAGCAC | CTTGTCTTTGACCCAGTAGC |
| MCP-1 | ATGATTCTACCCACGGCAAG | CTGGAAGATGGTGATGGGTT |
| GAPDH | ATGATTCTACCCACGGCAAG | CTGGAAGATGGTGATGGGTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, D.; Ko, W.-K.; Kim, S.-J.; Han, G.-H.; Lee, D.; Sheen, S.-H.; Sohn, S. Lobeglitazone Exerts Anti-Inflammatory Effect in Lipopolysaccharide-Induced Bone-Marrow Derived Macrophages. Biomedicines 2021, 9, 1432. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9101432
Jeong D, Ko W-K, Kim S-J, Han G-H, Lee D, Sheen S-H, Sohn S. Lobeglitazone Exerts Anti-Inflammatory Effect in Lipopolysaccharide-Induced Bone-Marrow Derived Macrophages. Biomedicines. 2021; 9(10):1432. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9101432
Chicago/Turabian StyleJeong, Dabin, Wan-Kyu Ko, Seong-Jun Kim, Gong-Ho Han, Daye Lee, Seung-Hun Sheen, and Seil Sohn. 2021. "Lobeglitazone Exerts Anti-Inflammatory Effect in Lipopolysaccharide-Induced Bone-Marrow Derived Macrophages" Biomedicines 9, no. 10: 1432. https://0-doi-org.brum.beds.ac.uk/10.3390/biomedicines9101432