Genetically Encoded Sensor Cells for the Screening of Glucocorticoid Receptor (GR) Effectors in Herbal Extracts

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. General Procedure
2.2. Preparation of Herbal and Plant Extracts
2.3. Sample Collection and Preparation to Detect Salivary Cortisol
2.4. Screening of GR Effectors Using Sensor Cells
2.5. Western Blotting
2.6. Competitive In Vitro GR Binding Assay
2.7. Statistics
2.8. Reporter Gene Assay
3. Results
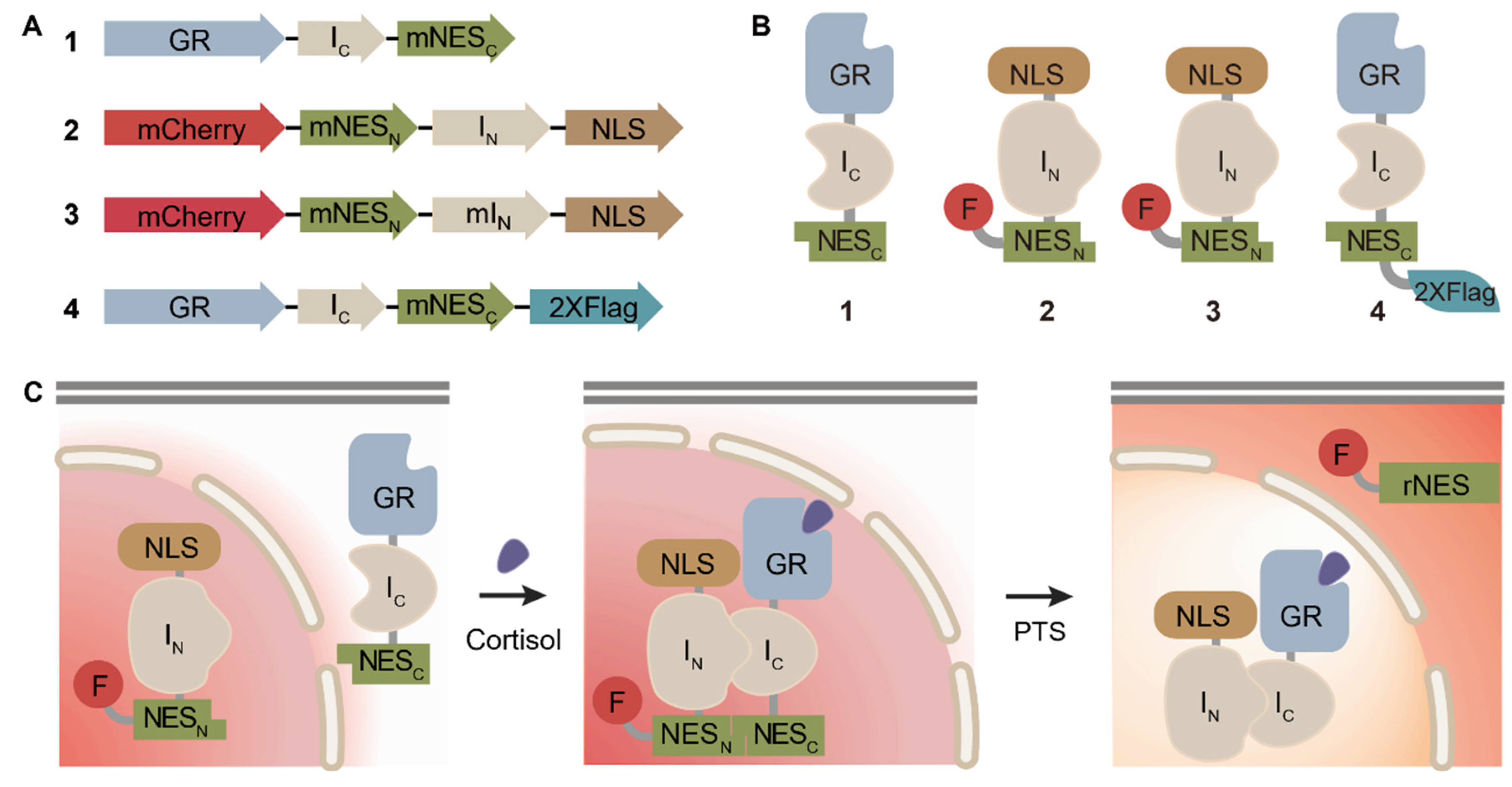
3.1. Fabrication and Characterization of Genetically Encoded Sensor Cells
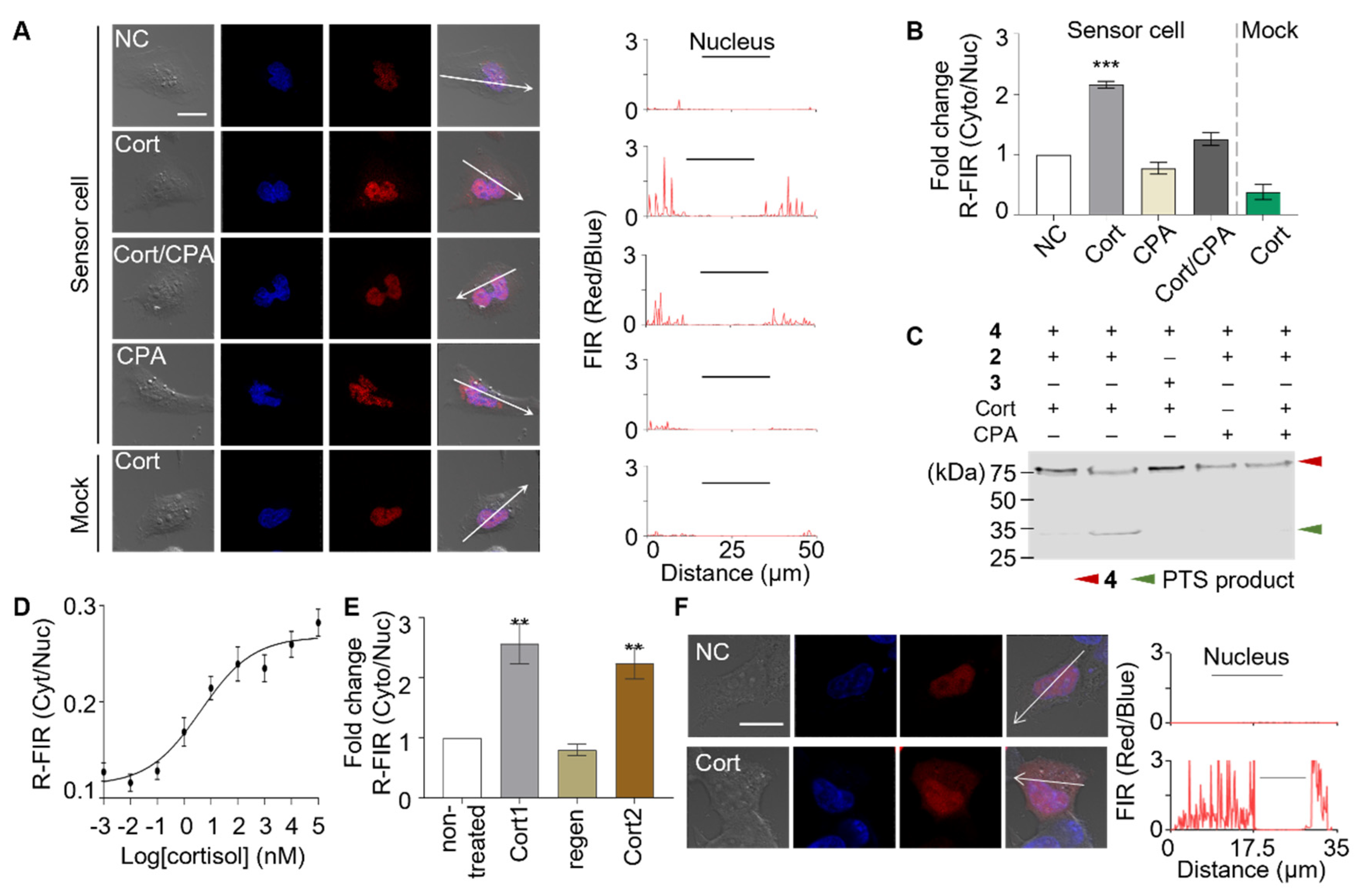
3.2. Evaluating the Performance of the Sensor Cells
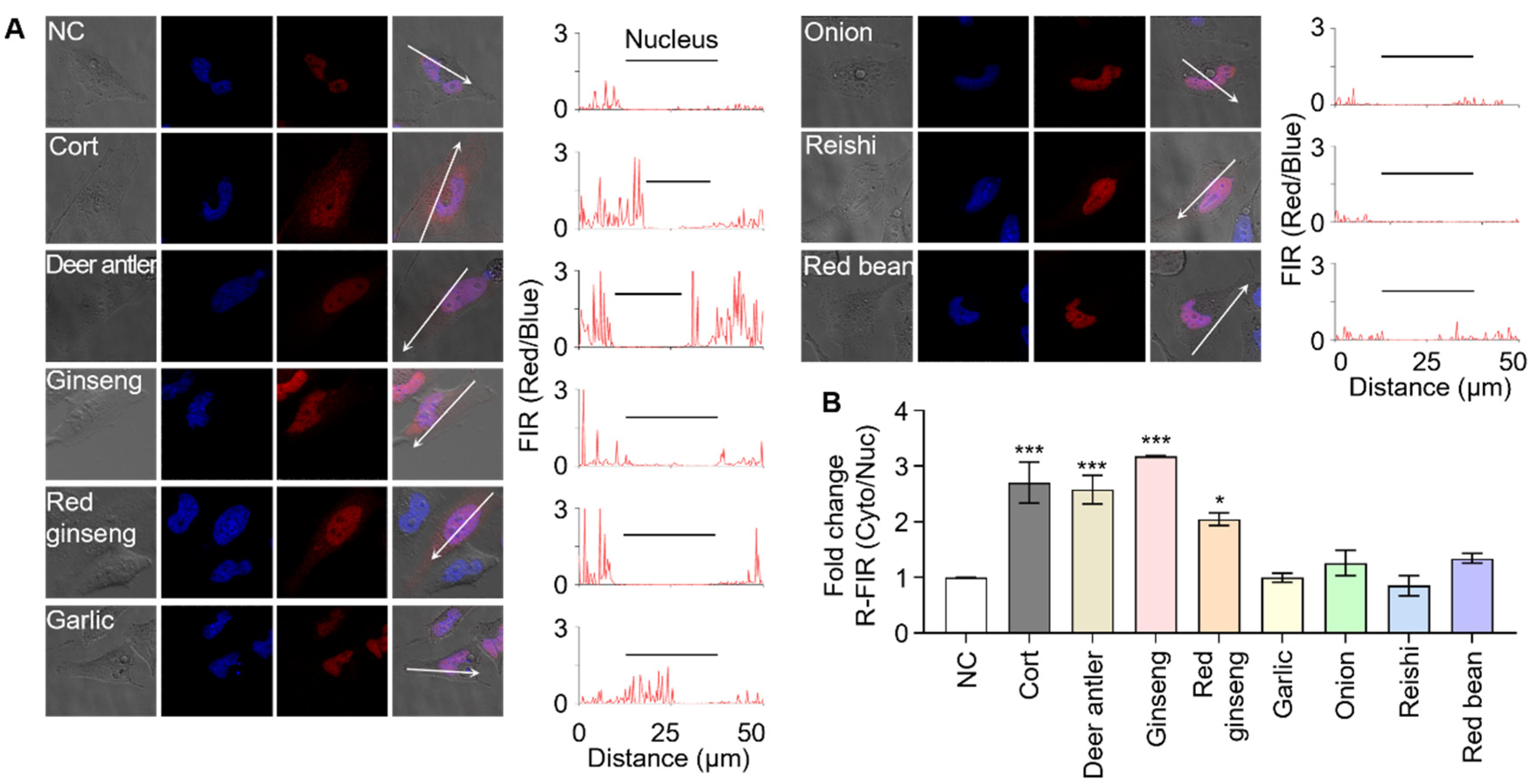
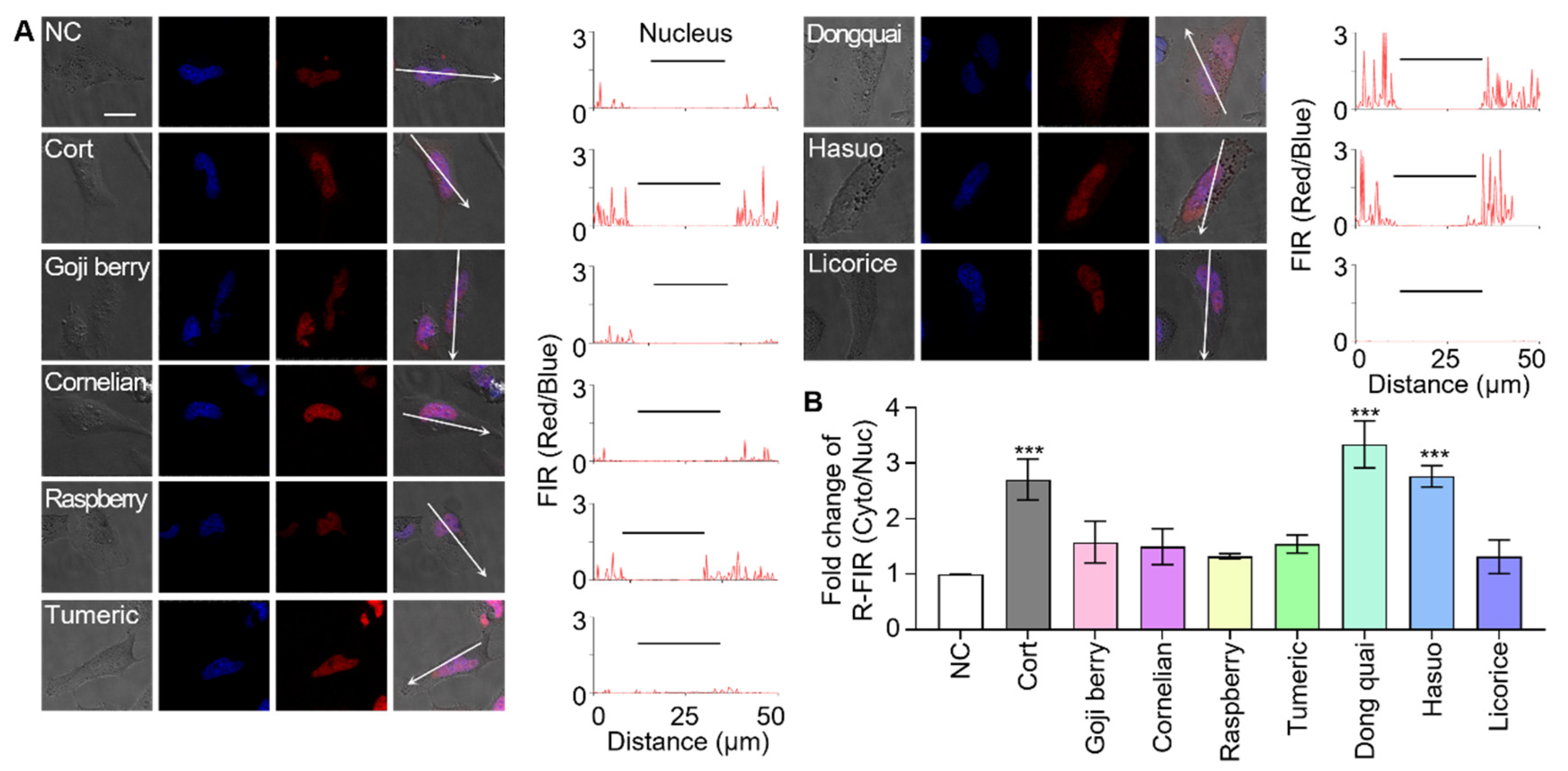
3.3. Screening Herbal Medicines to Identify GR Effectors
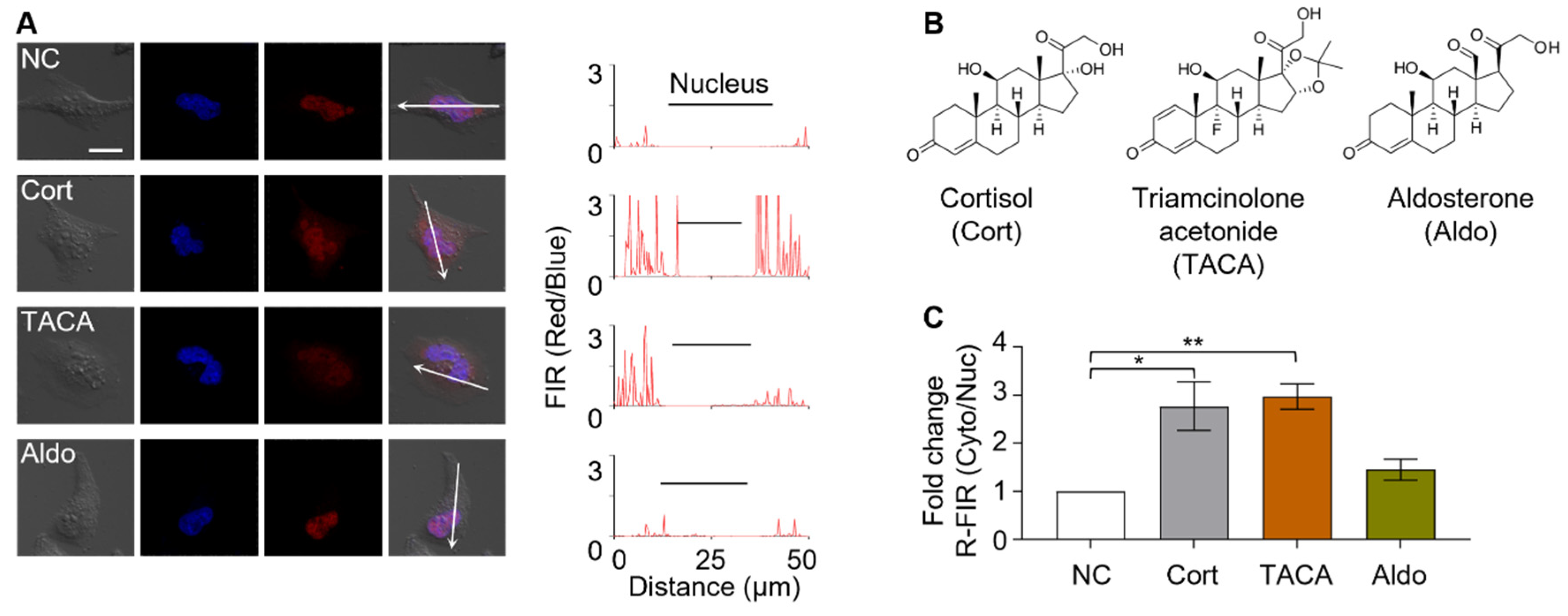
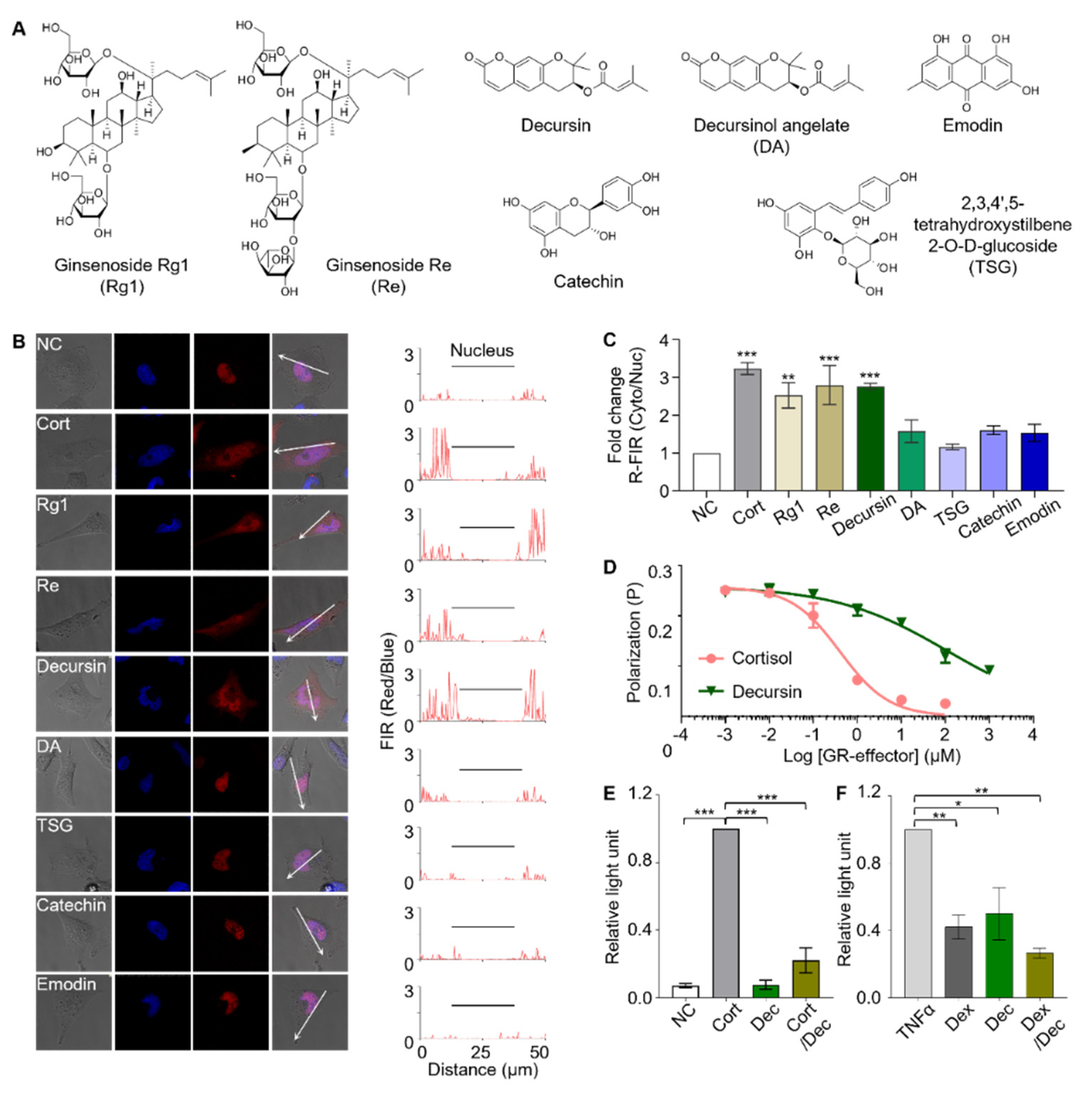
3.4. Identification of Active Components
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Lage, O.M.; Ramos, M.C.; Calisto, R.; Almeida, E.; Vasconcelos, V.; Vicente, F. Current Screening Methodologies in Drug Discovery for Selected Human Diseases. Mar. Drugs 2018, 16, 279. [Google Scholar] [CrossRef] [Green Version]
- Uri, A.; Nonga, O.E. What is the current value of fluorescence polarization assays in small molecule screening? Expert Opin. Drug Discov. 2020, 15, 131–133. [Google Scholar] [CrossRef] [Green Version]
- Khan, J.A.; Vulfson, E.N. Combinatorial Chemistry in Food Research. Comb. Chem. High Throughput Screen. 2003, 6, 569–574. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Choi, S.-J. Development of Tubing-based Stationary Liquid-phase Enzyme-linked Immunosorbent Assay. BioChip J. 2019, 13, 174–181. [Google Scholar] [CrossRef]
- Jung, J.; Bong, J.-H.; Kim, H.-R.; Park, J.-H.; Lee, C.K.; Kang, M.-J.; Kim, H.O.; Pyun, J.-C. Anti-SARS-CoV-2 Nucleoprotein Antibodies Derived from Pig Serum with a Controlled Specificity. BioChip J. 2021, 15, 195–203. [Google Scholar] [CrossRef]
- Banerjee, P.; Franz, B.; Bhunia, A.K. Mammalian cell-based sensor system. Adv. Biochem. Eng. Biotechnol. 2010, 117, 21–55. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Ching, H.; Yoon, J.-K.; Jeon, N.L.; Kim, Y. Microvascularized tumor organoids-on-chips: Advancing preclinical drug screening with pathophysiological relevance. Nano Converg. 2021, 8, 12. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.B.; Ozawa, T.; Umezawa, Y. A genetically encoded indicator for assaying bioactive chemicals that induce nuclear transport of glucocorticoid receptor. Anal. Biochem. 2005, 347, 213–220. [Google Scholar] [CrossRef]
- Liu, J.; Knappenberger, K.S.; Käck, H.; Andersson, G.; Nilsson, E.; Dartsch, C.; Scott, C.W. A Homogeneous in Vitro Functional Assay for Estrogen Receptors: Coactivator Recruitment. Mol. Endocrinol. 2003, 17, 346–355. [Google Scholar] [CrossRef] [Green Version]
- Weatherman, R.V.; Chang, C.Y.; Clegg, N.J.; Carroll, D.C.; Day, R.N.; Baxter, J.D.; McDonnell, D.P.; Scanlan, T.S.; Schaufele, F. Ligand-selective interactions of ER detected in living cells by fluorescence resonance energy transfer. Mol. Endocrinol. 2002, 16, 487–496. [Google Scholar] [CrossRef]
- Broussard, J.A.; Rappaz, B.; Webb, D.J.; Brown, C.M. Fluorescence resonance energy transfer microscopy as demonstrated by measuring the activation of the serine/threonine kinase Akt. Nat. Protoc. 2013, 8, 265–281. [Google Scholar] [CrossRef] [Green Version]
- Roszik, J.; Lisboa, D.; Szöllősi, J.; Vereb, G. Evaluation of intensity-based ratiometric FRET in image cytometry—Approaches and a software solution. Cytom. A 2009, 75, 761–767. [Google Scholar] [CrossRef]
- Berendzen, K.W.; Böhmer, M.; Wallmeroth, N.; Peter, S.; Vesić, M.; Zhou, Y.; Tiesler, F.K.; Schleifenbaum, F.; Harter, K. Screening for in planta protein-protein interactions combining bimolecular fluorescence complementation with flow cytometry. Plant Methods 2012, 8, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, K.; Kiuchi, T.; Shoji, K.; Sampei, K.; Mizuno, K. Visualization of cofilin-actin and Ras-Raf interactions by bimolecular fluorescence complementation assays using a new pair of split Venus fragments. Biotechniques 2012, 52, 45–50. [Google Scholar] [CrossRef]
- Magliery, T.J.; Wilson, C.G.M.; Pan, W.; Mishler, D.; Ghosh, I.; Hamilton, A.D.; Regan, L. Detecting Protein−Protein Interactions with a Green Fluorescent Protein Fragment Reassembly Trap: Scope and Mechanism. J. Am. Chem. Soc. 2005, 127, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.; Lee, E.; Kim, D.; Lee, M.; Ryu, J.; Kang, C.; Kim, S.; Kwon, Y. Cell-Based Biosensors Based on Intein-Mediated Protein Engineering for Detection of Biologically Active Signaling Molecules. Anal. Chem. 2018, 90, 9779–9786. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Jeon, H.; Ryu, J.; Kang, C.; Kim, S.; Park, S.; Kwon, Y. Genetically encoded biosensors for the detection of rapamycin: Toward the screening of agonists and antagonists. Analyst 2020, 145, 5571–5577. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.; Lee, E.; Kang, C.; Lee, M.; Kim, S.; Park, S.; Lee, D.; Kwon, Y. Rapid Screening of Glucocorticoid Receptor (GR) Effectors Using Cortisol-Detecting Sensor Cells. Int. J. Mol. Sci. 2021, 22, 4747. [Google Scholar] [CrossRef] [PubMed]
- Paulus, H. Inteins as Enzyme. Bioorganic Chem. 2001, 29, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.V.; Johnson, M.A.; Perler, F.B. Protein splicing: How inteins escape from precursor proteins. J. Biol. Chem. 2014, 289, 14498–14505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, T.P.; Marks, D.L. The regulation of muscle mass by endogenous glucocorticoids. Front. Physiol. 2015, 6, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niculet, E.; Bobeica, C.; Tatu, A.L. Glucocorticoid-Induced Skin Atrophy: The Old and the New. Clin. Cosmet. Investig. Dermatol. 2020, 13, 1041–1050. [Google Scholar] [CrossRef]
- Sari, E.; Üzek, R.; Duman, M.; Alagöz, H.Y.; Denizli, A. Prism coupler-based sensor system for simultaneous screening of synthetic glucocorticosteroid as doping control agent. Sens. Actuators B Chem. 2018, 260, 432–444. [Google Scholar] [CrossRef]
- Yadav, S.K.; Chandra, P.; Goyal, R.N.; Shim, Y.-B. A review on determination of steroids in biological samples exploiting nanobio-electroanalytical methods. Anal. Chim. Acta 2013, 762, 14–24. [Google Scholar] [CrossRef]
- Khan, M.S.; Dighe, K.; Wang, Z.; Srivastava, I.; Schwartz-Duval, A.S.; Misra, S.K.; Pan, D. Electrochemical-digital immunosensor with enhanced sensitivity for detecting human salivary glucocorticoid hormone. Analyst 2019, 144, 1448–1457. [Google Scholar] [CrossRef]
- Moshynskyy, I.; Viswanathan, S.; Vasilenko, N.; Lobanov, V.; Petric, M.; Babiuk, L.A.; Zakhartchouk, A.N. Intracellular localization of the SARS coronavirus protein 9b: Evidence of active export from the nucleus. Virus Res. 2007, 127, 116–121. [Google Scholar] [CrossRef]
- Laudat, M.H.; Cerdas, S.; Fournier, C.; Guiban, D.; Guilhaume, B.; Luton, J.P. Salivary Cortisol Measurement: A Practical Approach to Assess Pituitary-Adrenal Function. J. Clin. Endocrinol. Metabol. 1988, 66, 343–348. [Google Scholar] [CrossRef]
- Pancrazio, J.J.; Whelan, J.P.; Borkholder, D.A.; Ma, W.; Stenger, D.A. Development and Application of Cell-Based Biosensors. Ann. Biomed. Eng. 1999, 27, 697–711. [Google Scholar] [CrossRef] [PubMed]
- Paddle, B.M. Biosensors for chemical and biological agents of defence interest. Biosens. Bioelectron. 1996, 11, 1079–1113. [Google Scholar] [CrossRef]
- MS, U.; Ferdosh, S.; Haque Akanda, M.J.; Ghafoor, K.; AH, R.; Ali, M.E.; Kamaruzzaman, B.Y.; Fauzi, M.B.; Hadijah, S.; Shaarani, S.; et al. Techniques for the extraction of phytosterols and their benefits in human health: A review. Sep. Sci. Technol. 2018, 53, 2206–2223. [Google Scholar] [CrossRef]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Fung, F.Y.; Linn, Y.C. Steroids in traditional Chinese medicine: What is the evidence? Singap. Med. J. 2017, 58, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Wiseman, H. Phytochemicals: Health Effects. In Encyclopedia of Human Nutrition, 3rd ed.; Caballero, B., Ed.; Academic Press: Waltham, MA, USA, 2013; pp. 47–51. [Google Scholar]
- Gaspar-López, E.; Landete-Castillejos, T.; Estevez, J.A.; Ceacero, F.; Gallego, L.; García, A.J. Biometrics, testosterone, cortisol and antler growth cycle in Iberian red deer stags (Cervus elaphus hispanicus). Reprod. Domest. Anim. 2010, 45, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Francis, G.; Kerem, Z.; Makkar, H.P.S.; Becker, K. The biological action of saponins in animal systems: A review. Br. J. Nutr. 2002, 88, 587–605. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Chung, E.; Lee, K.Y.; Lee, Y.H.; Huh, B.; Lee, S.K. Ginsenoside-Rg1, one of the major active molecules from Panax ginseng, is a functional ligand of glucocorticoid receptor. Mol. Cell. Endocrinol. 1997, 133, 135–140. [Google Scholar] [CrossRef]
- Leung, K.W.; Wong, A.S.-T. Pharmacology of ginsenosides: A literature review. Chin. Med. 2010, 5, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, I.-W.; Cha, K.-M.; Wee, J.J.; Ye, M.B.; Kim, S.-K. A new validated analytical method for the quality control of red ginseng products. J. Ginseng. Res. 2013, 37, 475–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koh, E.; Jang, O.-H.; Hwang, K.-H.; An, Y.-N.; Moon, B. Effects of Steaming and Air-Drying on Ginsenoside Composition of Korean Ginseng (Panax ginseng C.A. Meyer). J. Food Process. Preserv. 2015, 39, 207–213. [Google Scholar] [CrossRef]
- Kao, T.-C.; Shyu, M.-H.; Yen, G.-C. Glycyrrhizic Acid and 18β-Glycyrrhetinic Acid Inhibit Inflammation via PI3K/Akt/GSK3β Signaling and Glucocorticoid Receptor Activation. J. Agric. Food Chem. 2010, 58, 8623–8629. [Google Scholar] [CrossRef]
- Kowalska, A.; Kalinowska-Lis, U. 18β-Glycyrrhetinic acid: Its core biological properties and dermatological applications. Int. J. Cosmet. Sci. 2019, 41, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.M.; Bae, B.-S.; Park, H.-W.; Ahn, N.-G.; Cho, B.-G.; Cho, Y.-L.; Kwak, Y.-S. Characterization of Korean Red Ginseng (Panax ginseng Meyer): History, preparation method, and chemical composition. J. Ginseng. Res. 2015, 39, 384–391. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.R.; Abd El-Aty, A.M.; Kim, I.S.; Shim, J.H. Determination of volatile flavor components in danggui cultivars by solvent free injection and hydrodistillation followed by gas chromatographic-mass spectrometric analysis. J. Chromatogr. A 2006, 1116, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Li, J.; Gao, X.M.; Amponsem, E.; Kang, L.Y.; Hu, L.M.; Zhang, B.L.; Chang, Y.X. Simultaneous determination of stilbenes, phenolic acids, flavonoids and anthraquinones in Radix polygoni multiflori by LC–MS/MS. J. Pharm. Biomed. Anal. 2012, 62, 162–166. [Google Scholar] [CrossRef]
- Shehzad, A.; Parveen, S.; Qureshi, M.; Subhan, F.; Lee, Y.S. Decursin and decursinol angelate: Molecular mechanism and therapeutic potential in inflammatory diseases. Inflamm. Res. 2018, 67, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shaik, A.A.; Xing, C.; Chai, Y.; Li, L.; Zhang, J.; Zhang, W.; Kim, S.-H.; Lü, J.; Jiang, C. A synthetic decursin analog with increased in vivo stability suppresses androgen receptor signaling in vitro and in vivo. Investig. New Drugs 2012, 30, 1820–1829. [Google Scholar] [CrossRef]
- Jiang, C.; Guo, J.; Wang, Z.; Xiao, B.; Lee, H.J.; Lee, E.O.; Kim, S.H.; Lu, J. Decursin and decursinol angelate inhibit estrogen-stimulated and estrogen-independent growth and survival of breast cancer cells. Breast Cancer Res. 2007, 9, R77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waddell, D.S.; Baehr, L.M.; van den Brandt, J.; Johnsen, S.A.; Reichardt, H.M.; Furlow, J.D.; Bodine, S.C. The glucocorticoid receptor and FOXO1 synergistically activate the skeletal muscle atrophy-associated MuRF1 gene. Am. J. Physiol. Endocrinol. Metabol. 2008, 295, E785–E797. [Google Scholar] [CrossRef]
- Galt, N.J.; McCormick, S.D.; Froehlich, J.M.; Biga, P.R. A comparative examination of cortisol effects on muscle myostatin and HSP90 gene expression in salmonids. Gen. Comp. Endocrinol. 2016, 237, 19–26. [Google Scholar] [CrossRef]
- Stoeber, M.; Jullié, D.; Lobingier, B.T.; Laeremans, T.; Steyaert, J.; Schiller, P.W.; Manglik, A.; von Zastrow, M. A Genetically Encoded Biosensor Reveals Location Bias of Opioid Drug Action. Neuron 2018, 98, 963–976.e965. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Mehta, S.; Zhang, J. Genetically encoded fluorescent biosensors illuminate kinase signaling in cancer. J. Biol. Chem. 2019, 294, 14814–14822. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, C.; Kim, S.; Lee, E.; Ryu, J.; Lee, M.; Kwon, Y. Genetically Encoded Sensor Cells for the Screening of Glucocorticoid Receptor (GR) Effectors in Herbal Extracts. Biosensors 2021, 11, 341. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11090341
Kang C, Kim S, Lee E, Ryu J, Lee M, Kwon Y. Genetically Encoded Sensor Cells for the Screening of Glucocorticoid Receptor (GR) Effectors in Herbal Extracts. Biosensors. 2021; 11(9):341. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11090341
Chicago/Turabian StyleKang, Chungwon, Soyoun Kim, Euiyeon Lee, Jeahee Ryu, Minhyeong Lee, and Youngeun Kwon. 2021. "Genetically Encoded Sensor Cells for the Screening of Glucocorticoid Receptor (GR) Effectors in Herbal Extracts" Biosensors 11, no. 9: 341. https://0-doi-org.brum.beds.ac.uk/10.3390/bios11090341