
Differential Long-Term Population Responses of Two Closely Related Human-Associated Sparrow Species with Respect to Urbanization


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Study Areas and Materials
2.3. Winter Bird Counts
2.4. Data Analyses
3. Results
3.1. House and Eurasian Tree Sparrow Trends in Europe and Finland
3.2. Winter Data and Sparrow Surveys
3.2.1. Background Variables
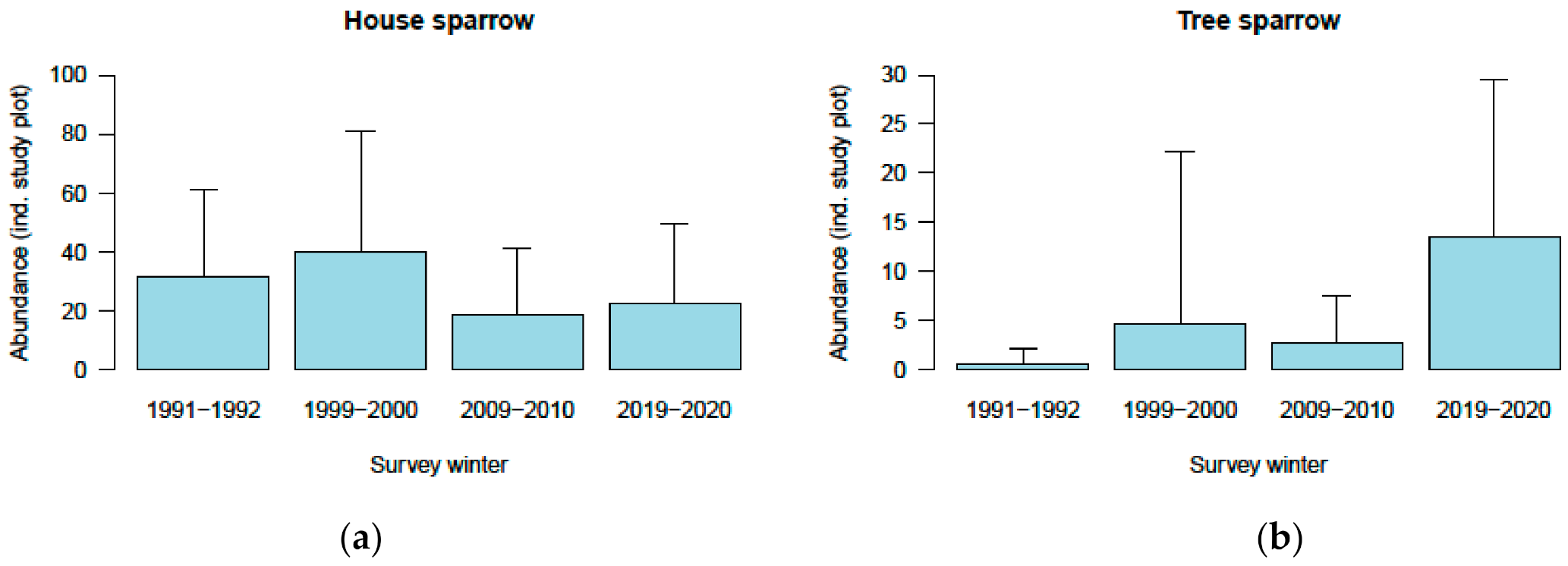
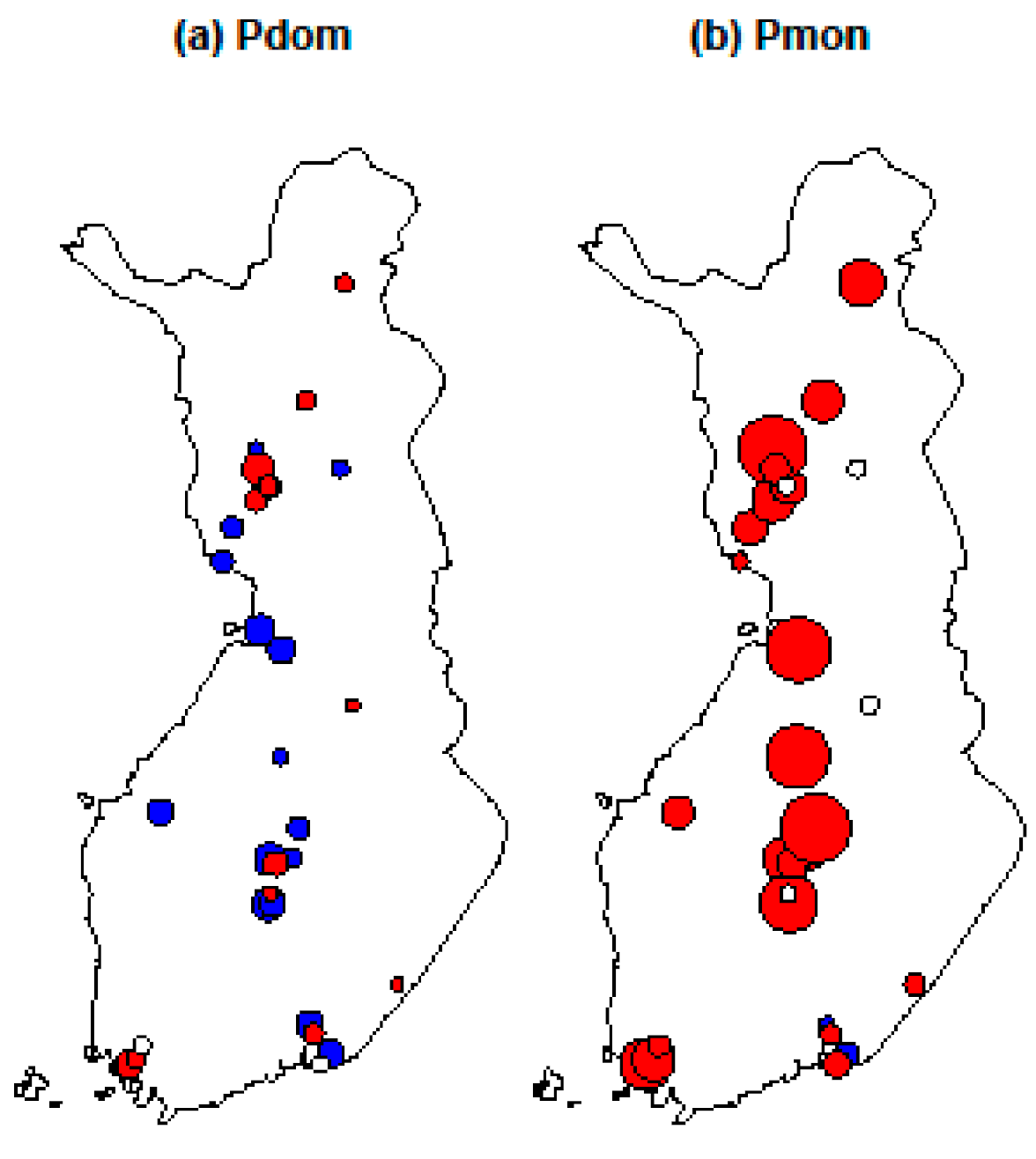
3.2.2. Population Trends during Winters
3.2.3. Factors Affecting Wintering Sparrows
4. Discussion
4.1. Sparrow Populations in Europe
4.2. Sparrow Populations in Finland
4.3. Sparrow Populations Growth Rates in Finland
4.4. Factors behind the Population Changes of the Two Sparrow Species
4.4.1. Latitude, Climate and Urbanization Level
4.4.2. Food Factor
4.4.3. Inter-Specific Competition
4.4.4. Predation and Changes in Predator Communities
4.4.5. Other Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marzluff, J.M. Worldwide urbanization and its effects on birds. In Avian Ecology and Conservation in an Urbanizing World; Marzluff, J.M., Bowman, R., Donelly, R., Eds.; Springer: Boston, MA, USA, 2001; pp. 19–47. [Google Scholar]
- Davey, C.M.; Chamberlain, D.E.; Newson, S.E.; Noble, D.G.; Johnston, A. Rise of the generalists: Evidence for climate driven homogenization in avian communities. Glob. Ecol. Biogeogr. 2012, 21, 568–578. [Google Scholar]
- Chace, J.F.; Walsh, J.J. Urban effects on native avifauna: A review. Landsc. Urban Plan. 2006, 74, 46–69. [Google Scholar]
- Tomiałojć, L. Quantitative studies on the synanthropic avifauna of Legnica and its environs. Acta Ornithol. 1970, 9, 293–392. [Google Scholar]
- Johnston, R.F. Synanthropic birds in North America. In Avian Ecology and Conservation in an Urbanizing World; Marzluff, J.M., Bowman, R., Donelly, R., Eds.; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2001; pp. 49–67. [Google Scholar]
- Jokimäki, J.; Suhonen, J. Distribution and habitat selection of wintering birds in urban environments. Landsc. Urban Plan. 1998, 39, 253–263. [Google Scholar]
- Fuller, R.A.; Warren, P.H.; Armsworth, P.R.; Barbosa, O.; Gaston, K.J. Garden bird feeding predicts the structure of urban avian assemblages. Divers. Distrib. 2008, 14, 131–137. [Google Scholar] [CrossRef]
- Robb, G.N.; McDonald, R.A.; Chamberlain, D.E.; Bearhop, S. Food for thought: Supplementary feeding as a driver of ecological change in avian populations. Front. Ecol. Environ. 2008, 6, 476–484. [Google Scholar]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L. Residential areas support overwintering possibilities of most bird species. Ann. Zool. Fenn. 2012, 49, 240–256. [Google Scholar]
- Galbraith, J.A.; Beggs, J.R.; Jones, D.N.; Stanley, M.C. Supplementary feeding restructures urban bird communities. Proc. Natl. Acad. Sci. USA 2015, 112, E2648–E2657. [Google Scholar]
- Blair, R.B. Land use and avian species diversity along an urban gradient. Ecol. Appl. 1996, 6, 506–519. [Google Scholar]
- Kekkonen, J. Pollutants in urbanized areas: Direct and indirect effects on bird populations. In Ecology and Conservation of Birds in Urban Environments; Murgui, E., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 227–250. [Google Scholar]
- Erskine, A.J. A ten-year urban winter bird count in Sackville, New Brunswick. Can. Field Nat. 1992, 106, 499–506. [Google Scholar]
- Lehikoinen, A.; Lehikoinen, E.; Valkama, J.; Väisänen, R.A.; Isomursu, M. Impacts of trichomonosis epidemics on Greenfinch Chloris chloris and Chaffinch Fringilla coelebs populations in Finland. Ibis 2013, 155, 357–366. [Google Scholar]
- Riley, S.P.; Serieys, L.E.; Moriarty, J.G. Infection disease and contaminants in urban wildlife: Unseen and often overlooked threats. In Urban Wildlife Conservation; McCleerly, R.A., Moorman, C.E., Peterson, N.M., Eds.; Springer: Boston, MA, USA, 2014; pp. 175–215. [Google Scholar]
- Walcott, C.F. Changes in bird life in Cambridge, Massachusetts from 1860 to 1964. Auk 1974, 91, 151–160. [Google Scholar]
- Batten, L.A. Population dynamics of suburban Blackbirds. Bird Study 1973, 20, 251–258. [Google Scholar]
- Sims, V.; Evans, K.L.; Newson, S.E.; Tratalos, J.A.; Gaston, K.J. Avian assemblage structure and domestic cat densities in urban environments. Divers. Distrib. 2008, 14, 387–399. [Google Scholar]
- van Heezik, Y.; Smyth, A.; Adams, A.; Gordon, J. Do domestic cats impose an unsustainable harvest on urban bird populations? Biol. Conserv. 2010, 143, 121–130. [Google Scholar]
- Lepczyk, C.A.; La Sorte, F.A.; Aronson, M.F.; Goddard, M.A.; MacGregor-Fors, I.; Nilon, C.H.; Warren, P.S. Global patterns and drivers of urban bird diversity. In Ecology and Conservation of Birds in Urban Environments; Murgui, E., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2017; pp. 13–33. [Google Scholar]
- Kark, S.; Iwaniuk, A.; Schalimtzek, A.; Banker, E. Living in the city: Can anyone become an ‘urban exploiter’? J. Biogeogr. 2007, 34, 638–651. [Google Scholar]
- Croci, S.; Butet, A.; Clergeau, P. Does urbanization filter birds on the basis of their biological traits. Condor 2008, 110, 223–240. [Google Scholar]
- Summers-Smith, D. The House Sparrow; Collins: London, UK, 1963; pp. 1–259. [Google Scholar]
- Avery, J.D.; Lockwood, J. Introduction history, impacts, and management of house sparrows in North America. In Ecology and Management of Terrestrial Vertebrate Invasive Species in the United States; Pitt, W.C., Beasley, J., Witmer, G.W., Eds.; CRC Press: Boca Raton, FL, USA, 2017; pp. 359–384. [Google Scholar]
- Summers-Smith, J.D. Current status of the house sparrow in Britain. Br. Wildl. 1999, 10, 381–386. [Google Scholar]
- Summers-Smith, J.D. The decline of the House Sparrow: A review. Br. Birds 2003, 96, 439–446. [Google Scholar]
- Summers-Smith, J.D. Changes in the house sparrow population in Britain. Int. Stud. Sparrows 2005, 30, 23–37. [Google Scholar]
- Siriwardena, G.M.; Robinson, R.A.; Crick, H.Q.P. Status and population trends of the House Sparrow Passer domesticus in Great Britain. In Investigation into the Causes of the Decline of Starlings and House Sparrows in Great Britain; BTO Research Report No 290; Crick, H.Q.P., Robinson, R.A., Appleton, G.F., Clark, N.A., Rickard, A.D., Eds.; DEFRA: Bristol, UK, 2002; pp. 33–51. [Google Scholar]
- Robinson, R.A.; Siriwardena, G.M.; Crick, H.Q. Size and trends of the House Sparrow Passer domesticus population in Great Britain. Ibis 2001, 147, 552–562. [Google Scholar]
- Berigan, L.A.; Greig, E.I.; Bonter, D.N. Urban House Sparrow (Passer domesticus) populations decline in North America. Wilson J. Ornithol. 2020, 132, 248–258. [Google Scholar]
- BirdLife International. European Birds of Conservation Concern: Populations, Trends and National Responsibilities; Birdlife International: Cambridge, UK, 2017; p. 172. [Google Scholar]
- De Laet, J.; Summers-Smith, J.D. The status of the urban house sparrow Passer domesticus in north-western Europe: A review. J. Ornithol. 2007, 148, 275–278. [Google Scholar]
- Burnett, J.L.; Roberts, C.P.; Allen, C.R.; Brown, M.B.; Moulton, M.P. Range expansion by Passer montanus in North America. Biol. Invasions 2017, 19, 5–9. [Google Scholar]
- Murgui, E.; Macias, A. Changes in the House Sparrow Passer domesticus population in Valencia (Spain) from 1998 to 2008. Bird Study 2010, 57, 281–288. [Google Scholar]
- Mueller, F. Attempt at a winter census of House Sparrows Passer domesticus in Plauen, Saxony (in Dutch with English abstract). Ornithol. Mitt. 2018, 70, 131–138. [Google Scholar]
- Zuckerberg, B.; Bonter, D.N.; Hochachka, W.M.; Koenig, W.D.; DeGaetano, A.T.; Dickinson, J.L. Climatic constraints on wintering bird distributions are modified by urbanization and weather. J. Anim. Ecol. 2011, 80, 403–413. [Google Scholar]
- Pakanen, V.M.; Ahonen, E.; Hohtola, E.; Rytkönen, S. Northward expanding resident species benefit from warming winters through increased foraging rates and predator vigilance. Oecologia 2018, 188, 991–999. [Google Scholar] [PubMed] [Green Version]
- Marzluff, J.M. A decadal review of urban ornithology and a prospectus for the future. Ibis 2017, 159, 1–13. [Google Scholar]
- Marzluff, J.M. A historical perspective on urban bird research: Trends, terms, and approaches. In Avian Ecology and Conservation in an Urbanizing World; Marzluff, J.M., Bowman, R., Donelly, R., Eds.; Springer: Boston, MA, USA; pp. 1–17.
- Fidino, M.; Magle, S.B. Trends in long-term urban bird research. In Ecology and Conservation of Birds in Urban Environments; Murgui, E., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 161–184. [Google Scholar]
- van Heezik, Y.; Seddon, P.J. Counting birds in urban areas: A review of methods for the estimation of abundance. In Ecology and Conservation of Birds in Urban Environments; Murgui, E., Hedblom, M., Eds.; Springer: Cham, Switzerland, 2018; pp. 185–207. [Google Scholar]
- Šálek, M.; Riegert, J.; Grill, S. House Sparrows Passer domesticus and Tree Sparrows Passer montanus: Fine-scale distribution, population densities, and habitat selection in a Central European city. Acta Ornithol. 2015, 50, 221–232. [Google Scholar]
- Tryjanowski, P.; Møller, A.P.; Morelli, F.; Indykiewicz, P.; Zduniak, P.; Myczko, Ł. Food preferences by birds using bird-feeders in winter: A large-scale experiment. Avian Res. 2018, 9, 16. [Google Scholar] [CrossRef] [Green Version]
- Väisänen, R.A. Changes in frequency and abundance of 63 bird species at winter feeding sites in Finland during 19 winters 1988/1989–2006/2007. Linnut Vuosik. 2008, 2007, 60–79, (In Finnish with English Summary). [Google Scholar]
- Cordero, P.J.; Senar, J.C. Persistent tree sparrows Passer montanus can counteract house sparrow P. domesticus competitive pressure. Bird Behav. 1994, 10, 7–13. [Google Scholar] [CrossRef]
- Vepsäläinen, V.; Pakkala, T.; Tiainen, J. Population increase and aspects of colonization of the Tree Sparrow Passer montanus, and its relationships with the House Sparrow Passer domesticus, in the agricultural landscapes of Southern Finland. Ornis Fenn. 2005, 82, 117–128. [Google Scholar]
- Węgrzynowicz, A. The use of nest–boxes by two species of Sparrows Passer domesticus and P. montanus with opposite trends of abundance—The study in Warsaw. Int. Stud. Sparrows 2012, 36, 18–29. [Google Scholar] [CrossRef] [Green Version]
- van Manen, W. House Sparrow Passer domesticus and Tree Sparrow P. montana in the Netherlands, 42 years of monitoring. Limosa 2020, 93, 49–58, (In Dutch with an English Abstract). [Google Scholar]
- IUCN Red List for Birds. Available online: http://www.birdlife.org (accessed on 26 January 2021).
- Valkama, J.; Vepsäläinen, V.; Lehikoinen, A. The Third Finnish Breeding Bird Atlas; Finnish Museum of Natural History and Ministry of Environment: Helsinki, Finland, 2011; Available online: http://atlas3.lintuatlas.fi/english (accessed on 18 November 2020)ISBN 978-952-10-7145-4.
- Lehikoinen, A.; Jukarainen, A.; Mikkola-Roos, M.; Below, A.; Lehtiniemi, T.; Pessa, J.; Rajasärkkä, A.; Rintala, J.; Rusanen, P.; Sirkiä, P.; et al. Birds. In The 2019 Red List of Finnish Species; Hyvärinen, E., Juslén, A., Kemppainen, E., Uddström, A., Liukko, U.-M., Eds.; Ministry of the Environment & Finnish Environment: Helsinki, Finland, 2019; pp. 560–570. [Google Scholar]
- BirdLife Finland 2020. Pihabongaus. Available online: https://www.birdlife.fi/tapahtumat/pihabongaus/ (accessed on 26 November 2020).
- BirdLife Finland 2020. Pönttöbongaus. Available online: https://www.birdlife.fi/tapahtumat/ponttobongaus/ (accessed on 26 November 2020).
- Fraixedas, S.; Lehikoinen, A.; Lindén, A. Impacts of climate and land-use change on wintering bird populations in Finland. J. Avian Biol. 2015, 46, 63–72. [Google Scholar]
- Jokimäki, J.; Suhonen, J.; Inki, K.; Jokinen, S. Biogeographical comparison of winter bird assemblages in urban environments in Finland. J. Biogeogr. 1996, 23, 379–386. [Google Scholar] [CrossRef]
- Suhonen, J.; Jokimäki, J. Temporally Stable Species Occupancy Frequency Distribution and Abundance–Occupancy Relationship Patterns in Urban Wintering Bird Assemblages. Front. Ecol. Evol. 2019, 7, 129. [Google Scholar] [CrossRef] [Green Version]
- Suhonen, J.; Jokimäki, J.; Kaisanlahti-Jokimäki, M.L.; Hakkarainen, H.; Huhta, E.; Inki, K.; Suorsa, P. Urbanization and stability of a bird community in winter. Ecoscience 2009, 16, 502–507. [Google Scholar] [CrossRef]
- Open data extraction. Finnish Meterological Insitute. Creative Commons 4.0 License (CC BY 4.0). Available online: https://www.ilmatieteenlaitos.fi/havaintojen-lataus (accessed on 20 December 2020).
- Bibby, C.J.; Burgess, N.D.; Hill, D.A.; Hillis, D.M.; Mustoe, S. Bird Census Techniques, 2nd ed.; Elsevier: Oxford, UK, 2000. [Google Scholar]
- Valkama, J.; Saurola, P.; Lehikoinen, A.; Lehikoinen, E.; Piha, M.V.; Sola, P.; Velmala, W. Suomen Rengastusatlas. Osa II; Luonnontieteellinen keskusmuseo: Helsinki, Finland, 2014; pp. 1–784. [Google Scholar]
- Tryjanowski, P.; Skórka, P.; Sparks, T.H.; Biaduń, W.; Brauze, T.; Hetmański, T.; Martyka, R.; Indykiewicz, P.; Myczko, Ł.; Kunysz, P.; et al. Urban and rural habitats differ in number and type of bird feeders and in bird species consuming supplementary food. Environ. Sci. Pollut. Res. 2015, 22, 15097–15103. [Google Scholar] [CrossRef] [Green Version]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L. Spatial similarity of urban bird communities: A multiscale approach. J. Biogeogr. 2003, 30, 1183–1193. [Google Scholar] [CrossRef]
- Morris, W.F.; Doak, D.F. Quantitative Conservation Biology; Sinauer: Sunderland, MA, USA, 2002. [Google Scholar]
- Keller, V.; Herrando, S.; Voříšek, P.; Franch, M.; Kipson, M.; Milanesi, P.; Marti, D.; Anton, M.; Klvaňová, A.; Kalyakin, M.V.; et al. European Breeding Bird Atlas 2: Distribution, Abundance and Change, Lynx ed.; European Bird Census Council: Beek, The Netherlands, 2020. [Google Scholar]
- Hole, D.G.; Whittingham, M.J.; Bradbury, R.B.; Anderson, G.Q.; Lee, P.L.; Wilson, J.D.; Krebs, J.R. Widespread local house-sparrow extinctions. Nature 2002, 418, 931–932. [Google Scholar] [CrossRef] [PubMed]
- Siriwardena, G.M.; Baillie, S.R.; Buckland, S.T.; Fewster, R.M.; Marchant, J.H.; Wilson, J.D. Trends in the abundance of farmland birds: A quantitative comparison of smoothed Common Birds Census indices. J. Appl. Ecol. 1998, 35, 24–43. [Google Scholar] [CrossRef]
- Gregory, R.D.; Noble, D.G.G.; Custance, J. The state of play of farmland birds: Population trends and conservation status of lowland farmland birds in the United Kingdom. Ibis 2004, 146, 1–13. [Google Scholar] [CrossRef]
- Węgrzynowicz, A. Changes in the House Sparrow Passer domesticus population in cities and towns of Poland in 1960–2010. Ornis Pol. 2013, 54, 225–236. [Google Scholar]
- Mohring, B.; Henry, P.Y.; Jiguet, F.; Malher, F.; Angelier, F. Investigating temporal and spatial correlates of the sharp decline of an urban exploiter bird in a large European city. Urban Ecosyst. 2021, 24, 501–513. [Google Scholar] [CrossRef]
- Summers-Smith, J.D. A history of the status of the Tree Sparrow Passer montanus in the British Isles. Bird Study 1989, 36, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Both, C.; Visser, M.E.; Van Balen, H. (The rise and fall of the Eurasian Tree Sparrow population). De opkomst en ondergang van een populatie Ringmussen Passer montanus. Limosa 2002, 75, 41–50. (In Dutch) [Google Scholar]
- Chamberlain, D.E.; Fuller, R.J. Contrasting patterns of change in the distribution and abundance of farmland birds in relation to farming system in lowland Britain. Glob. Ecol. Biogeogr. 2001, 10, 399–409. [Google Scholar] [CrossRef]
- Graham, J.; Jarnevich, C.; Young, N.; Newman, G.; Stohlgren, T. How will climate change affect the potential distribution of Eurasian tree sparrows Passer montanus in North America? Curr. Zool. 2011, 57, 648–654. [Google Scholar] [CrossRef] [Green Version]
- Węgrzynowicz, A. Importance of nest sites availability for abundance and changes in number of House and Tree Sparrow in Warsaw. Int. Stud. Sparrows 2012, 36, 56–65. [Google Scholar] [CrossRef]
- Virkkala, R.; Lehikoinen, A. Birds on the move in the face of climate change: High species turnover in northern Europe. Ecol. Evol. 2017, 7, 8201–8209. [Google Scholar] [CrossRef] [PubMed]
- Väisänen, R.A. Monitoring population changes of 86 land bird species breeding in Finland in 1983–2005. Linnut Vuosik. 2016, 2015, 83–98, (In Finnish with an English Summary). [Google Scholar]
- Lehikoinen, A.; Tirri, I. Changes in regional wintering abundances of birds in Finland. Linnut Vuosik. 2021, 2020, 18–29, (In Finnish with an English Summary). [Google Scholar]
- Väisänen, R.A. Long-term changes in frequency and abundance of bird species at winter feeding sites of Finland during 29 winters 1988/1989—2016/2017. Linnut Vuosik. 2018, 2017, 32–47, (In Finnish with English Summary). [Google Scholar]
- Väisänen, R.A. Frequency and abundance of 61 bird species at feeding sites in Finland in 1990/1991—2019/2020. Linnut Vuosik. 2021, 2020, 30–45, (In Finnish with English Summary). [Google Scholar]
- Lehikoinen, A.; Väisänen, R.A. Habitat-specific changes of Finnish winter bird populations in 1987–2014. Linnut Vuosik. 2014, 2013, 78–93, (In Finnish with English Summary). [Google Scholar]
- Wuczyński, A.; Wuczyński, M. Birds Wintering in Heterogeneous Farmland of Poland: Weather-Dependent Temporal Changes in Abundance and Habitat Associations. Acta Ornithol. 2019, 54, 105–124. [Google Scholar] [CrossRef]
- Murgui, E. Seasonal patterns of habitat selection of the House Sparrow Passer domesticus in the urban landscape of Valencia (Spain). J. Ornithol. 2009, 150, 85–94. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Toms, M.P.; Cleary-McHarg, R.; Banks, A.N. House sparrow (Passer domesticus) habitat use in urbanized landscapes. J. Ornithol. 2007, 148, 453–462. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, G. Effect of urbanization on the abundance and distribution of Tree Sparrows (Passer montanus) in Beijing. Chin. Birds 2010, 1, 188–197. [Google Scholar] [CrossRef]
- MacGregor-Fors, I.; Quesada, J.; Lee, J.G.; Yeh, P.J. Space invaders: House sparrow densities along three urban-agricultural landscapes. Avian Conserv. Ecol. 2017, 12, 11. [Google Scholar] [CrossRef]
- Nath, A.; Singha, H.; Haque, M.; Lahkar, B.P. Sparrows in urban complexity: Macro and micro-scale habitat use of sympatric sparrows in Guwahati City, India. Urban Ecosyst. 2019, 22, 1047–1060. [Google Scholar] [CrossRef]
- Bernat-Ponce, E.; Gil-Delgado, J.A.; López-Iborra, G.M. Replacement of semi-natural cover with artificial substrates in urban parks causes a decline of house sparrows Passer domesticus in Mediterranean towns. Urban Ecosyst. 2020, 23, 471–481. [Google Scholar] [CrossRef]
- Tomiałojć, L. Changes in tree sparrow Passer montanus populations from urban parks. Int. Stud. Sparrows 2012, 36, 4–17. [Google Scholar] [CrossRef] [Green Version]
- Plummer, K.E.; Risely, K.; Toms, M.P.; Siriwardena, G.M. The composition of British bird communities is associated with long-term garden bird feeding. Nat. Commun. 2019, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cowie, R.J.; Hinsley, S.A. The provision of food and the use of bird feeders in suburban gardens. Bird Study 1988, 35, 163–168. [Google Scholar] [CrossRef]
- Chosińska, K.; Duduś, L.; Jakubiec, Z. Supplemental feeding of birds in human settlements of western Poland. Int. Stud. Sparrows 2012, 36, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Anderies, J.M.; Katti, M.; Shochat, E. Living in the city: Resource availability, predation, and bird population dynamics in urban areas. J. Theor. Biol. 2007, 247, 36–49. [Google Scholar] [CrossRef]
- Chamberlain, D.E.; Cannon, A.R.; Toms, M.P.; Leech, D.I.; Hatchwell, B.J.; Gaston, K.J. Avian productivity in urban landscapes: A review and meta-analysis. Ibis 2009, 151, 1–18. [Google Scholar] [CrossRef]
- Oro, D.; Genovart, M.; Tavecchia, G.; Fowler, M.S.; Martínez-Abraín, A. Ecological and evolutionary implications of food subsidies from humans. Ecol. Lett. 2013, 16, 1501–1514. [Google Scholar] [CrossRef]
- Villén-Pérez, S.; Carrascal, L.M.; Seoane, J. Foraging patch selection in winter: A balance between predation risk and thermoregulation benefit. PLoS ONE 2013, 8, e68448. [Google Scholar]
- Siriwardena, G.M.; Baillie, S.R.; Wilson, J.D. Temporal variation in the annual survival rates of six granivorous birds with contrasting population trends. Ibis 1999, 141, 621–636. [Google Scholar] [CrossRef]
- Moudrá, L.; Zasadil, P.; Moudrý, V.; Šálek, M. What makes new housing development unsuitable for house sparrows (Passer domesticus)? Landsc. Urban Plan. 2018, 169, 124–130. [Google Scholar] [CrossRef]
- Rosin, Z.M.; Hiron, M.; Żmihorski, M.; Szymański, P.; Tobolka, M.; Pärt, T. Reduced biodiversity in modernized villages: A conflict between sustainable development goals. J. Appl. Ecol. 2020, 57, 467–475. [Google Scholar] [CrossRef]
- Jokimäki, J.; Kaisanlahti-Jokimäki, M.-L. (Eds.) Rovaniemen Pesimälinnusto. Lapin Yliopisto; Arktisen Keskuksen Tiedotteita 57; Joutsen Median Painotalo Oy: Oulu, Finland, 2012; (In Finnish with an English Summary). [Google Scholar]
- Antikainen, E. The vertical use of a city park by urban birds in Poland. Ornis Fenn. 1992, 69, 92–96. [Google Scholar]
- Ciach, M.; Fröhlich, A. Habitat type, food resources, noise and light pollution explain the species composition, abundance and stability of a winter bird assemblage in an urban environment. Urban Ecosyst. 2017, 20, 547–559. [Google Scholar] [CrossRef] [Green Version]
- McKinney, M.L. Urbanization, biodiversity and conservation. Biosciences 2002, 52, 883–890. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | Friedman Test | |||||||
|---|---|---|---|---|---|---|---|---|
| 1990 | 2000 | 2009 | 2019 | χ2 | df | p | ||
| Built area (ha) | Mean | 1.5A | 1.7B | 1.8C | 1.9C | 69.04 | 3 | <0.001 |
| SD | 1.9 | 2.2 | 2.4 | 2.4 | ||||
| Number of buildings | Mean | 16.9A | 17.4B | 17.5B | 17.5B | 7.90 | 3 | 0.043 |
| SD | 7.4 | 7.3 | 7.4 | 6.9 | ||||
| Number of inhabitants | Mean | 136.6 | 144.5 | 143.8 | 146.6 | 3.12 | 3 | 0.374 |
| SD | 121.3 | 137.7 | 142.9 | 152.5 | ||||
| Winter temperature (°C) | Mean | −4.4A | −6.8B | −11.8C | −2.5D | 91.58 | 3 | <0.001 |
| SD | 2.2 | 3.3 | 2.0 | 3.1 | ||||
| Snow arrival | Mean | 76.4A | 60.9B | 51.6B | 64.8B | 23.74 | 3 | <0.001 |
| SD | 17.8 | 16.0 | 35.7 | 49.5 | ||||
| Snow amount (cm) | Mean | 18.8 | 24.4 | 17.2 | 22.2 | 4.78 | 3 | 0.189 |
| SD | 11.7 | 16.3 | 7.3 | 22.9 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jokimäki, J.; Suhonen, J.; Kaisanlahti-Jokimäki, M.-L. Differential Long-Term Population Responses of Two Closely Related Human-Associated Sparrow Species with Respect to Urbanization. Birds 2021, 2, 230-249. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2030017
Jokimäki J, Suhonen J, Kaisanlahti-Jokimäki M-L. Differential Long-Term Population Responses of Two Closely Related Human-Associated Sparrow Species with Respect to Urbanization. Birds. 2021; 2(3):230-249. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2030017
Chicago/Turabian StyleJokimäki, Jukka, Jukka Suhonen, and Marja-Liisa Kaisanlahti-Jokimäki. 2021. "Differential Long-Term Population Responses of Two Closely Related Human-Associated Sparrow Species with Respect to Urbanization" Birds 2, no. 3: 230-249. https://0-doi-org.brum.beds.ac.uk/10.3390/birds2030017