The Development of Brain Network in Males with Autism Spectrum Disorders from Childhood to Adolescence: Evidence from fNIRS Study

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
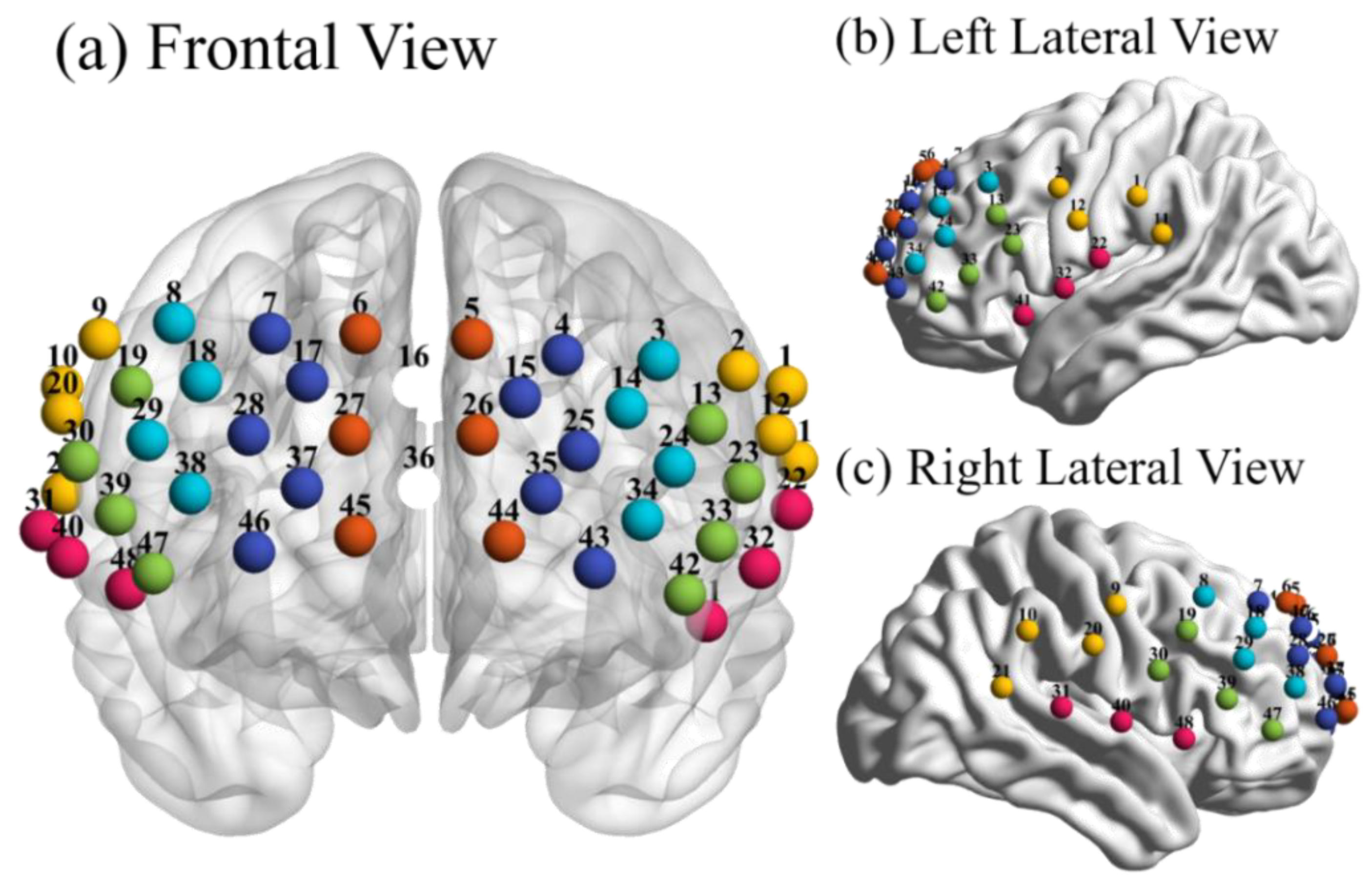
2.2. Data Acquisition
2.3. Data Preprocessing
2.4. Data Analysis
2.4.1. Network Construction
2.4.2. Network Analysis
2.4.3. Statistical Analysis
3. Results
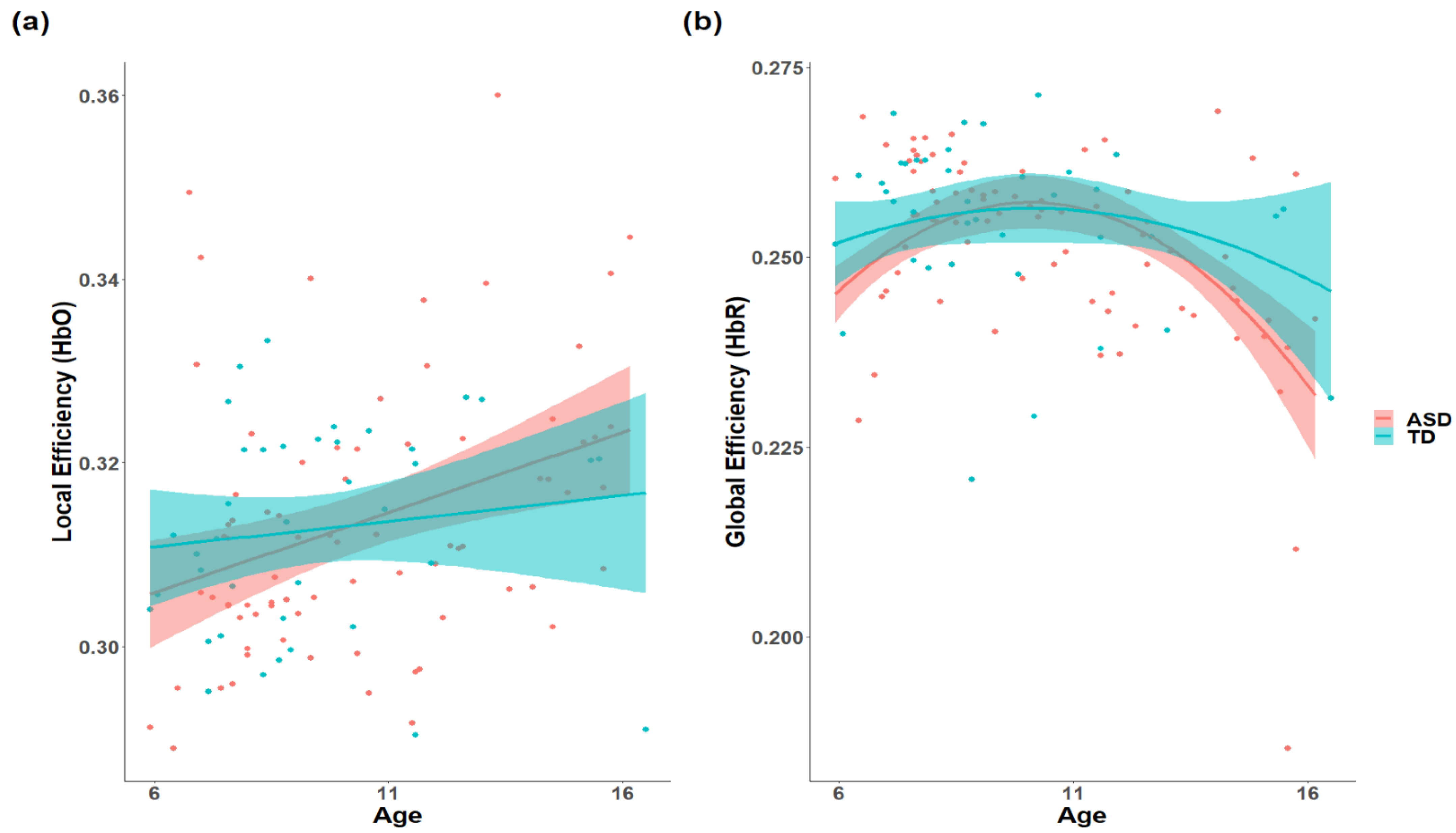
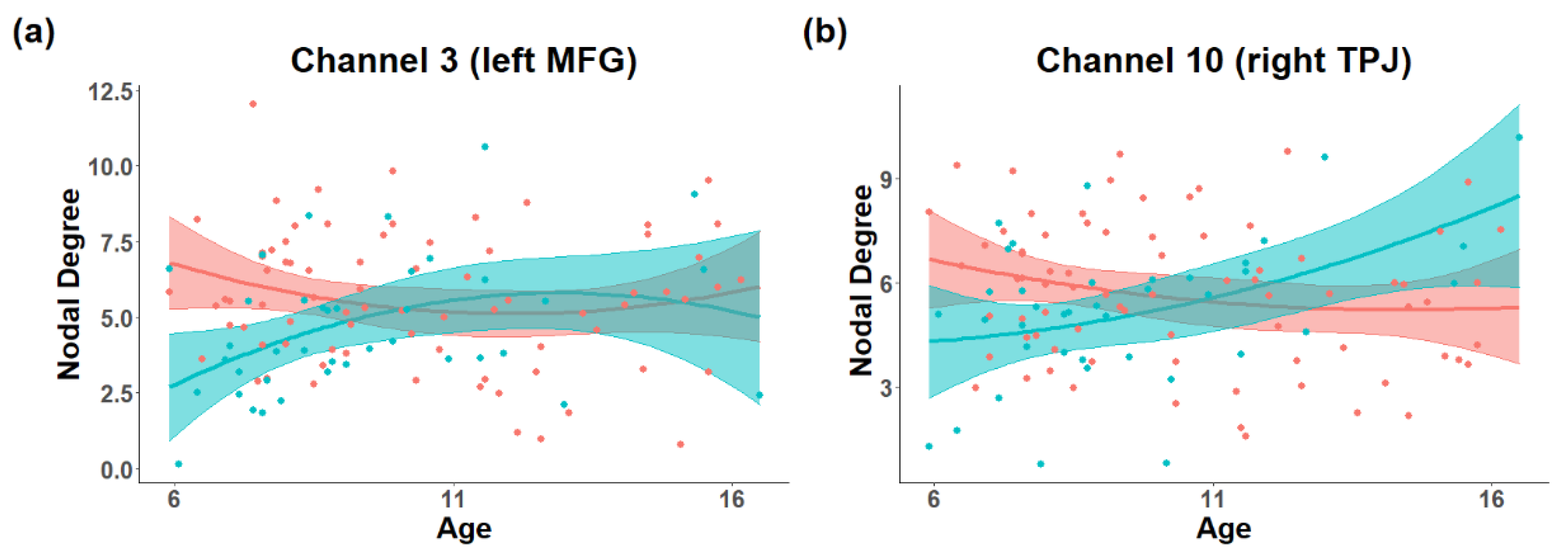
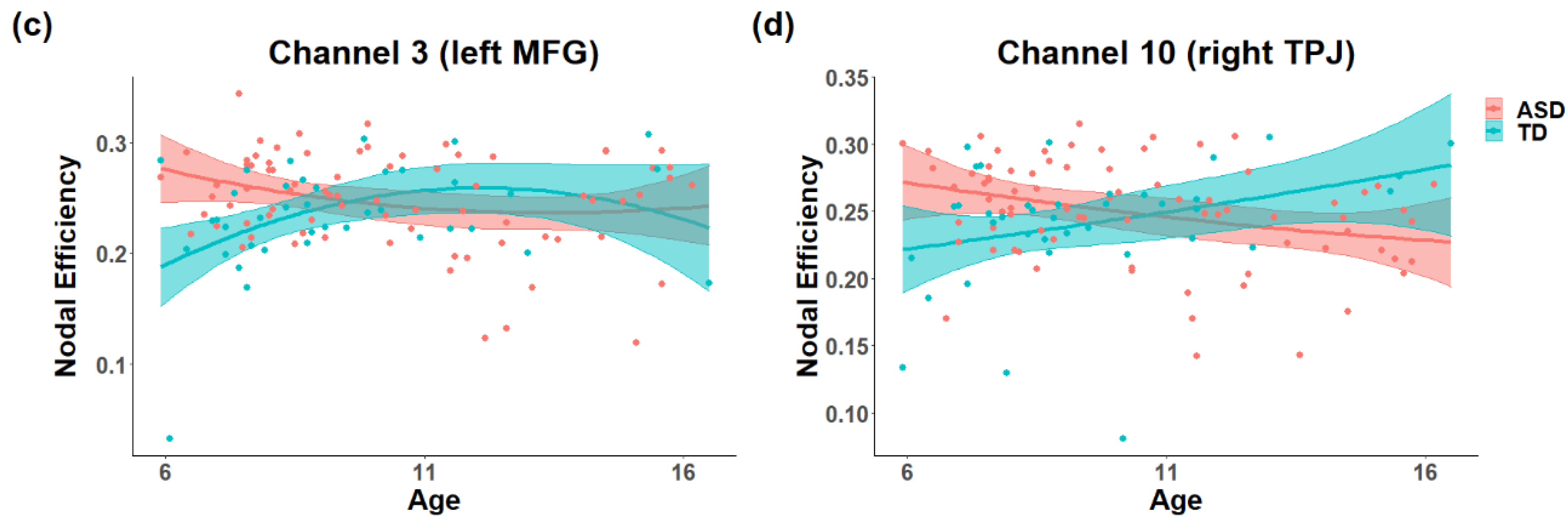
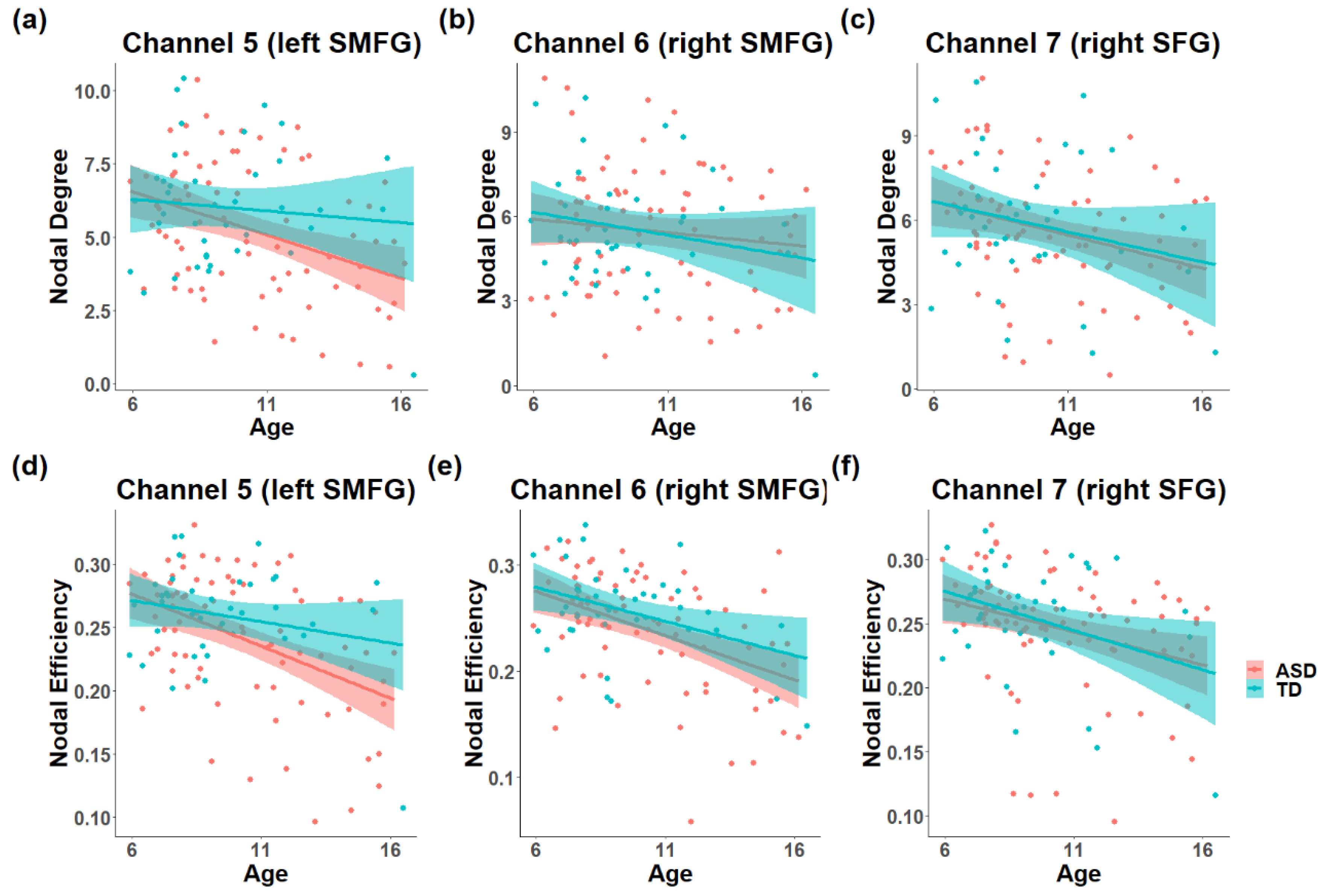
3.1. The Difference in the Development of Topology in the ToM Network
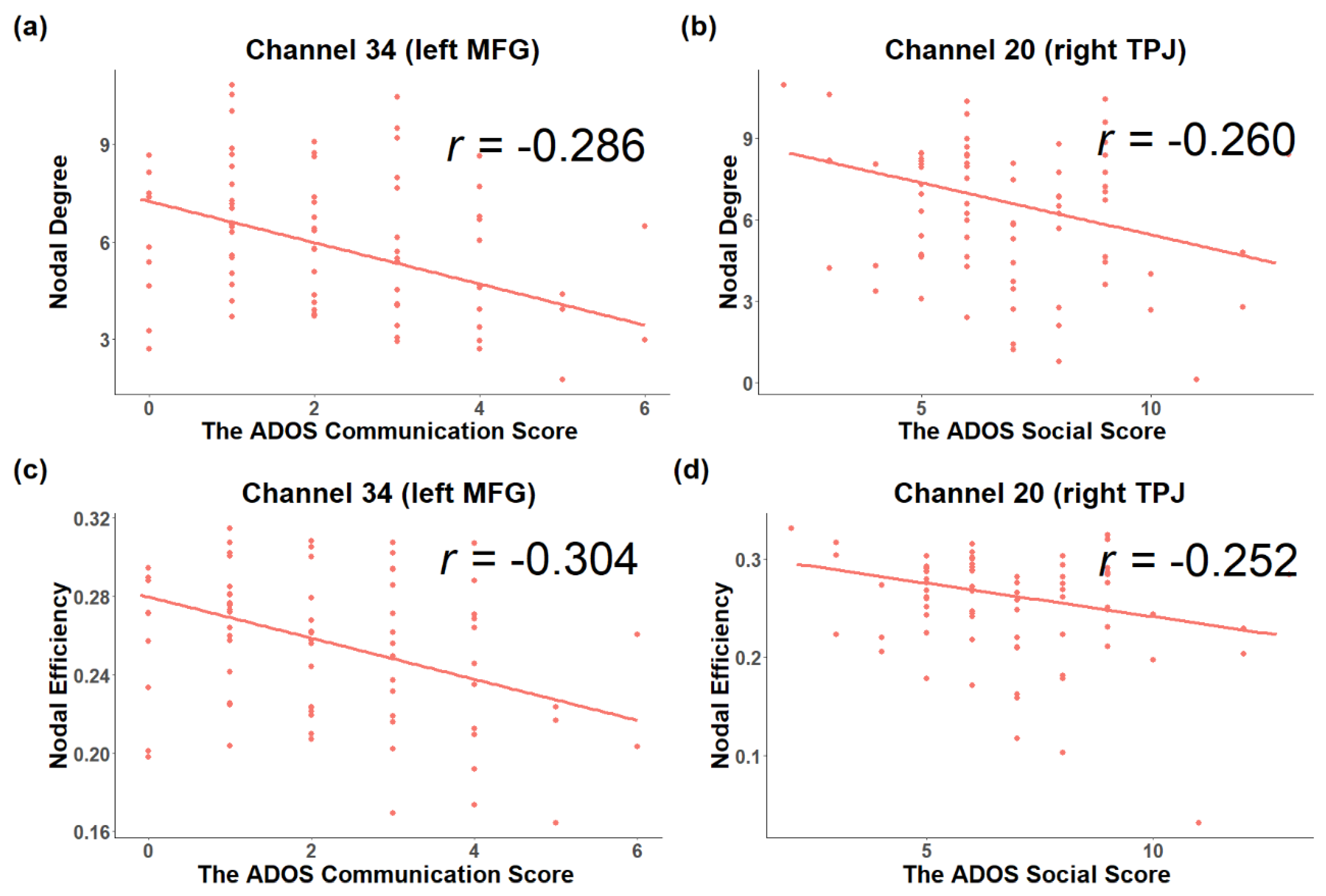
3.2. Associations between Topological Metrics and Severity of ASD Symptoms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Press: Washington, DC, USA, 2013. [Google Scholar]
- Blakemore, S.-J. Development of the Social Brain in Adolescence. J. R. Soc. Med. 2012, 105, 111–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picci, G.; Scherf, K.S. A Two-Hit Model of Autism: Adolescence as the Second Hit. Clin. Psychol. Sci. 2015, 3, 349–371. [Google Scholar] [CrossRef] [Green Version]
- Tunç, B.; Yankowitz, L.D.; Parker, D.; Alappatt, J.A.; Pandey, J.; Schultz, R.T.; Verma, R. Deviation from Normative Brain Development Is Associated with Symptom Severity in Autism Spectrum Disorder. Mol. Autism 2019, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Courchesne, E. Abnormal Early Brain Development in Autism. Mol. Psychiatry 2002, 7, S21–S23. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, E.; Pierce, K.; Schumann, C.M.; Redcay, E.; Buckwalter, J.A.; Kennedy, D.P.; Morgan, J. Mapping Early Brain Development in Autism. Neuron 2007, 56, 399–413. [Google Scholar] [CrossRef] [Green Version]
- Nomi, J.S.; Uddin, L.Q. Developmental Changes in Large-Scale Network Connectivity in Autism. NeuroImage Clin. 2015, 7, 732–741. [Google Scholar] [CrossRef] [Green Version]
- Abbott, A.E.; Nair, A.; Keown, C.L.; Datko, M.; Jahedi, A.; Fishman, I.; Müller, R.-A. Patterns of Atypical Functional Connectivity and Behavioral Links in Autism Differ Between Default, Salience, and Executive Networks. Cereb. Cortex 2016, 26, 4034–4045. [Google Scholar] [CrossRef] [PubMed]
- Rashid, B.; Blanken, L.M.E.; Muetzel, R.L.; Miller, R.; Damaraju, E.; Arbabshirani, M.R.; Erhardt, E.B.; Verhulst, F.C.; van der Lugt, A.; Jaddoe, V.W.V.; et al. Connectivity Dynamics in Typical Development and Its Relationship to Autistic Traits and Autism Spectrum Disorder. Hum. Brain Mapp. 2018, 39, 3127–3142. [Google Scholar] [CrossRef]
- Lynn, A.C.; Padmanabhan, A.; Simmonds, D.; Foran, W.; Hallquist, M.N.; Luna, B.; O’Hearn, K. Functional Connectivity Differences in Autism during Face and Car Recognition: Underconnectivity and Atypical Age-Related Changes. Dev. Sci. 2018, 21, e12508. [Google Scholar] [CrossRef] [Green Version]
- Lawrence, K.E.; Hernandez, L.M.; Bookheimer, S.Y.; Dapretto, M. Atypical Longitudinal Development of Functional Connectivity in Adolescents with Autism Spectrum Disorder. Autism Res. 2019, 12, 53–65. [Google Scholar] [CrossRef] [Green Version]
- Odriozola, P.; Dajani, D.R.; Burrows, C.A.; Gabard-Durnam, L.J.; Goodman, E.; Baez, A.C.; Tottenham, N.; Uddin, L.Q.; Gee, D.G. Atypical Frontoamygdala Functional Connectivity in Youth with Autism. Dev. Cogn. Neurosci. 2019, 37, 100603. [Google Scholar] [CrossRef]
- Dajani, D.R.; Uddin, L.Q. Local Brain Connectivity across Development in Autism Spectrum Disorder: A Cross-Sectional Investigation. Autism Res. 2016, 9, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Bullmore, E.; Sporns, O. Complex Brain Networks: Graph Theoretical Analysis of Structural and Functional Systems. Nat. Rev. Neurosci. 2009, 10, 312. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Evans, A. Graph Theoretical Modeling of Brain Connectivity. Curr. Opin. Neurol. 2010, 23, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Rudie, J.D.; Brown, J.A.; Beck-Pancer, D.; Hernandez, L.M.; Dennis, E.L.; Thompson, P.M.; Bookheimer, S.Y.; Dapretto, M. Altered Functional and Structural Brain Network Organization in Autism. NeuroImage Clin. 2013, 2, 79–94. [Google Scholar] [CrossRef] [Green Version]
- Riedel, B.C.; Jahanshad, N.; Thompson, P.M. Graph Theoretical Approaches towards Understanding Differences in Frontoparietal and Default Mode Networks in Autism. In Proceedings of the 2017 IEEE 14th International Symposium on Biomedical Imaging, Melbourne, VIC, Australia, 18–21 April 2017; pp. 460–463. [Google Scholar] [CrossRef]
- Lee, Y.; Park, B.; James, O.; Kim, S.-G.; Park, H. Autism Spectrum Disorder Related Functional Connectivity Changes in the Language Network in Children, Adolescents and Adults. Front. Hum. Neurosci. 2017, 11. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Zeng, K.; Kang, J.; Tong, Z.; Cai, E.; Chen, H.; Ding, M.; Gu, Y.; Ouyang, G.; Li, X. Development of Brain Network in Children with Autism from Early Childhood to Late Childhood. Neuroscience 2017, 367, 134–146. [Google Scholar] [CrossRef]
- Henry, T.R.; Dichter, G.S.; Gates, K. Age and Gender Effects on Intrinsic Connectivity in Autism Using Functional Integration and Segregation. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 414–422. [Google Scholar] [CrossRef]
- Chein, J.; Albert, D.; O’Brien, L.; Uckert, K.; Steinberg, L. Peers Increase Adolescent Risk Taking by Enhancing Activity in the Brain’s Reward Circuitry. Dev. Sci. 2011, 14, F1–F10. [Google Scholar] [CrossRef] [Green Version]
- Falk, E.B.; Cascio, C.N.; O’Donnell, M.B.; Carp, J.; Tinney, F.J., Jr.; Bingham, C.R.; Shope, J.T.; Ouimet, M.C.; Pradhan, A.K.; Simons-Morton, B.G. Neural Responses to Exclusion Predict Susceptibility to Social Influence. J. Adolesc. Health 2014, 54, S22–S31. [Google Scholar] [CrossRef] [Green Version]
- Baron-Cohen, S.; Leslie, A.M.; Frith, U. Does the Autistic Child Have a “Theory of Mind”? Cognition 1985, 21, 37–46. [Google Scholar] [CrossRef]
- Schaafsma, S.M.; Pfaff, D.W.; Spunt, R.P.; Adolphs, R. Deconstructing and Reconstructing Theory of Mind. Trends Cogn. Sci. 2015, 19, 65–72. [Google Scholar] [CrossRef] [Green Version]
- Fishman, I.; Keown, C.L.; Lincoln, A.J.; Pineda, J.A.; Müller, R.-A. Atypical Cross Talk between Mentalizing and Mirror Neuron Networks in Autism Spectrum Disorder. JAMA Psychiatry 2014, 71, 751–760. [Google Scholar] [CrossRef] [Green Version]
- White, S.J.; Frith, U.; Rellecke, J.; Al-Noor, Z.; Gilbert, S.J. Autistic Adolescents Show Atypical Activation of the Brain’s Mentalizing System Even without a Prior History of Mentalizing Problems. Neuropsychologia 2014, 56, 17–25. [Google Scholar] [CrossRef] [Green Version]
- Burrows, C.A.; Laird, A.R.; Uddin, L.Q. Functional Connectivity of Brain Regions for Self- and Other-Evaluation in Children, Adolescents and Adults with Autism. Dev. Sci. 2016, 19, 564–580. [Google Scholar] [CrossRef] [Green Version]
- Moessnang, C.; Baumeister, S.; Tillmann, J.; Goyard, D.; Charman, T.; Ambrosino, S.; Baron-Cohen, S.; Beckmann, C.; Bölte, S.; Bours, C.; et al. Social Brain Activation during Mentalizing in a Large Autism Cohort: The Longitudinal European Autism Project. Mol. Autism 2020, 11, 17. [Google Scholar] [CrossRef] [Green Version]
- Baron-Cohen, S.; Knickmeyer, R.C.; Belmonte, M.K. Sex Differences in the Brain: Implications for Explaining Autism. Science 2005, 310, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Kozhemiako, N.; Vakorin, V.; Nunes, A.S.; Iarocci, G.; Ribary, U.; Doesburg, S.M. Extreme Male Developmental Trajectories of Homotopic Brain Connectivity in Autism. Hum. Brain Mapp. 2019, 40, 987–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alaerts, K.; Swinnen, S.P.; Wenderoth, N. Sex Differences in Autism: A Resting-State FMRI Investigation of Functional Brain Connectivity in Males and Females. Soc. Cogn. Affect. Neurosci. 2016, 11, 1002–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholkmann, F.; Kleiser, S.; Metz, A.J.; Zimmermann, R.; Mata Pavia, J.; Wolf, U.; Wolf, M. A Review on Continuous Wave Functional Near-Infrared Spectroscopy and Imaging Instrumentation and Methodology. NeuroImage 2014, 85, 6–27. [Google Scholar] [CrossRef]
- Lord, C.; Risi, S.; Lambrecht, L.; Cook, E.H.; Leventhal, B.L.; DiLavore, P.C.; Pickles, A.; Rutter, M. The Autism Diagnostic Observation Schedule—Generic: A Standard Measure of Social and Communication Deficits Associated with the Spectrum of Autism. J. Autism Dev. Disord. 2000, 30, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Wang, J.; He, Y. BrainNet Viewer: A Network Visualization Tool for Human Brain Connectomics. PLoS ONE 2013, 8, e68910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholkmann, F.; Spichtig, S.; Muehlemann, T.; Wolf, M. How to Detect and Reduce Movement Artifacts in Near-Infrared Imaging Using Moving Standard Deviation and Spline Interpolation. Physiol. Meas. 2010, 31, 649–662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molavi, B.; Dumont, G.A. Wavelet Based Motion Artifact Removal for Functional Near Infrared Spectroscopy. In Proceedings of the 2010 Annual International Conference of the IEEE Engineering in Medicine and Biology, Buenos Aires, Argentina, 31 August–4 September 2010; pp. 5–8. [Google Scholar]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A Review of Time-Series Analysis Methods for near-Infrared Spectroscopy of the Brain. Appl. Opt. 2009, 48, D280. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, R.; Pirazzoli, L.; Blasi, A.; Bulgarelli, C.; Hakuno, Y.; Minagawa, Y.; Brigadoi, S. Recommendations for Motion Correction of Infant FNIRS Data Applicable to Data Sets Acquired with a Variety of Experimental Designs and Acquisition Systems. NeuroImage 2019. [Google Scholar] [CrossRef]
- White, B.R.; Snyder, A.Z.; Cohen, A.L.; Petersen, S.E.; Raichle, M.E.; Schlaggar, B.L.; Culver, J.P. Resting-State Functional Connectivity in the Human Brain Revealed with Diffuse Optical Tomography. Neuroimage 2009, 47, 148–156. [Google Scholar] [CrossRef] [Green Version]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional Connectivity in the Motor Cortex of Resting Human Brain Using Echo-Planar MRI. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef]
- Cai, L.; Dong, Q.; Wang, M.; Niu, H. Functional Near-Infrared Spectroscopy Evidence for the Development of Topological Asymmetry between Hemispheric Brain Networks from Childhood to Adulthood. Neurophotonics 2019, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Niu, H.; Wang, J.; Zhao, T.; Shu, N.; He, Y. Revealing Topological Organization of Human Brain Functional Networks with Resting-State Functional near Infrared Spectroscopy. PLoS ONE 2012, 7, e45771. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wang, X.; Xia, M.; Liao, X.; Evans, A.; He, Y. GRETNA: A Graph Theoretical Network Analysis Toolbox for Imaging Connectomics. Front. Hum. Neurosci. 2015, 9, 386. [Google Scholar] [CrossRef] [Green Version]
- Niu, H.; Li, Z.; Liao, X.; Wang, J.; Zhao, T.; Shu, N.; Zhao, X.; He, Y. Test-Retest Reliability of Graph Metrics in Functional Brain Networks: A Resting-State FNIRS Study. PLoS ONE 2013, 8, e72425. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Duan, F.; Watanabe, K.; Yoshimura, Y.; Kikuchi, M.; Minabe, Y.; Aihara, K. Detection of Atypical Network Development Patterns in Children with Autism Spectrum Disorder Using Magnetoencephalography. PLoS ONE 2017, 12, e0184422. [Google Scholar] [CrossRef] [Green Version]
- McCormick, E.M.; van Hoorn, J.; Cohen, J.R.; Telzer, E.H. Functional Connectivity in the Social Brain across Childhood and Adolescence. Soc. Cogn. Affect. Neurosci. 2018, 13, 819–830. [Google Scholar] [CrossRef]
- Bernas, A.; Barendse, E.M.; Aldenkamp, A.P.; Backes, W.H.; Hofman, P.A.M.; Hendriks, M.P.H.; Kessels, R.P.C.; Willems, F.M.J.; de With, P.H.N.; Zinger, S.; et al. Brain Resting-state Networks in Adolescents with High-functioning Autism: Analysis of Spatial Connectivity and Temporal Neurodynamics. Brain Behav. 2018, 8, e00878. [Google Scholar] [CrossRef]
- Di Martino, A.; Zuo, X.-N.; Kelly, C.; Grzadzinski, R.; Mennes, M.; Schvarcz, A.; Rodman, J.; Lord, C.; Castellanos, F.X.; Milham, M.P. Shared and Distinct Intrinsic Functional Network Centrality in Autism and Attention-Deficit/Hyperactivity Disorder. Biol. Psychiatry 2013, 74, 623–632. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Xu, J.; Li, J.; Peng, H.; Cai, T.; Li, X.; Wu, S.; Cao, W.; He, S. Decreased Functional Connectivity and Disrupted Neural Network in the Prefrontal Cortex of Affective Disorders: A Resting-State FNIRS Study. J. Affect. Disord. 2017, 221, 132–144. [Google Scholar] [CrossRef]
- Park, M.T.M.; Raznahan, A.; Shaw, P.; Gogtay, N.; Lerch, J.P.; Chakravarty, M.M. Neuroanatomical Phenotypes in Mental Illness: Identifying Convergent and Divergent Cortical Phenotypes across Autism, ADHD and Schizophrenia. J. Psychiatry Neurosci. JPN 2018, 43, 21. [Google Scholar] [CrossRef] [Green Version]
- Dajani, D.R.; Burrows, C.A.; Odriozola, P.; Baez, A.; Nebel, M.B.; Mostofsky, S.H.; Uddin, L.Q. Investigating Functional Brain Network Integrity Using a Traditional and Novel Categorical Scheme for Neurodevelopmental Disorders. NeuroImage Clin. 2019, 21, 101678. [Google Scholar] [CrossRef]
- Molenberghs, P.; Johnson, H.; Henry, J.D.; Mattingley, J.B. Understanding the Minds of Others: A Neuroimaging Meta-Analysis. Neurosci. Biobehav. Rev. 2016, 65, 276–291. [Google Scholar] [CrossRef] [Green Version]
- Schurz, M.; Radua, J.; Aichhorn, M.; Richlan, F.; Perner, J. Fractionating Theory of Mind: A Meta-Analysis of Functional Brain Imaging Studies. Neurosci. Biobehav. Rev. 2014, 42, 9–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kana, R.K.; Libero, L.E.; Hu, C.P.; Deshpande, H.D.; Colburn, J.S. Functional Brain Networks and White Matter Underlying Theory-of-Mind in Autism. Soc. Cogn. Affect. Neurosci. 2014, 9, 98–105. [Google Scholar] [CrossRef] [Green Version]
- White, S.J.; Coniston, D.; Rogers, R.; Frith, U. Developing the Frith-Happé Animations: A Quick and Objective Test of Theory of Mind for Adults with Autism. Autism Res. 2011, 4, 149–154. [Google Scholar] [CrossRef]
- Lombardo, M.V.; Chakrabarti, B.; Bullmore, E.T.; Baron-Cohen, S. Specialization of Right Temporo-Parietal Junction for Mentalizing and Its Relation to Social Impairments in Autism. NeuroImage 2011, 56, 1832–1838. [Google Scholar] [CrossRef]
- Murdaugh, D.L.; Nadendla, K.D.; Kana, R.K. Differential Role of Temporoparietal Junction and Medial Prefrontal Cortex in Causal Inference in Autism: An Independent Component Analysis. Neurosci. Lett. 2014, 568, 50–55. [Google Scholar] [CrossRef]
- Blakemore, S.-J.; den Ouden, H.; Choudhury, S.; Frith, C. Adolescent Development of the Neural Circuitry for Thinking about Intentions. Soc. Cogn. Affect. Neurosci. 2007, 2, 130–139. [Google Scholar] [CrossRef] [Green Version]
- Otti, A.; Wohlschlaeger, A.M.; Noll-Hussong, M. Is the Medial Prefrontal Cortex Necessary for Theory of Mind? PLoS ONE 2015, 10, e0135912. [Google Scholar] [CrossRef] [Green Version]
- Boccadoro, S.; Cracco, E.; Hudson, A.R.; Bardi, L.; Nijhof, A.D.; Wiersema, J.R.; Brass, M.; Mueller, S.C. Defining the Neural Correlates of Spontaneous Theory of Mind (ToM): An FMRI Multi-Study Investigation. Neuroimage 2019, 203, 116193. [Google Scholar] [CrossRef]
- Hartwright, C.E.; Apperly, I.A.; Hansen, P.C. Representation, Control, or Reasoning? Distinct Functions for Theory of Mind within the Medial Prefrontal Cortex. J. Cogn. Neurosci. 2013, 26, 683–698. [Google Scholar] [CrossRef]
- Miao, B.; Guan, J.; Meng, Q.; Zhang, Y. Fractional Amplitude of Low-Frequency Fluctuation and Degree Centrality in Autistic Children: A Resting-State FMRI Study. In Proceedings of the Third International Workshop on Pattern Recognition, Jinan, China, 26 July 2018; Volume 10828, p. 1082812. [Google Scholar]
- Noonan, S.K.; Haist, F.; Müller, R.A. Aberrant Functional Connectivity in Autism: Evidence from Low-Frequency BOLD Signal Fluctuations. Brain Res. 2009. [Google Scholar] [CrossRef] [Green Version]
- Wen, J.; Yu, T.; Liu, L.; Hu, Z.; Yan, J.; Li, Y.; Li, X. Evaluating the Roles of Left Middle Frontal Gyrus in Word Production Using Electrocorticography. Neurocase 2017, 23, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.-C.; Lin, H.-Y.; Chen, Y.-C.; Tseng, W.-Y.I.; Gau, S.S.-F. Boys with Autism Spectrum Disorder Have Distinct Cortical Folding Patterns Underpinning Impaired Self-Regulation: A Surface-Based Morphometry Study. Brain Imaging Behav. 2019. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ASD (n = 77) | TD (n = 40) | t | p | |
|---|---|---|---|---|
| Age (years) | 10.51 (2.86) 6~16.25 | 9.54(2.58) 6~16.58 | 1.780 | 0.078 |
| VIQ | 105.79 (17.44) | 106.05 (13.72) | −0.081 | 0.936 |
| PIQ | 104.68 (14.36) | 108.85 (12.66) | −1.536 | 0.127 |
| FSIQ | 101.47 (14.85) | 105.10 (12.10) | −1.322 | 0.189 |
| ADOS communication | 2.21 (1.50) | |||
| ADOS social | 6.79 (2.15) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, W.; Zhu, H.; Li, Y.; Wang, Y.; Bai, W.; Lao, U.; Zhang, Y.; Ji, Y.; He, S.; Zou, X. The Development of Brain Network in Males with Autism Spectrum Disorders from Childhood to Adolescence: Evidence from fNIRS Study. Brain Sci. 2021, 11, 120. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11010120
Cao W, Zhu H, Li Y, Wang Y, Bai W, Lao U, Zhang Y, Ji Y, He S, Zou X. The Development of Brain Network in Males with Autism Spectrum Disorders from Childhood to Adolescence: Evidence from fNIRS Study. Brain Sciences. 2021; 11(1):120. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11010120
Chicago/Turabian StyleCao, Wei, Huilin Zhu, Yan Li, Yu Wang, Wuxia Bai, Uchong Lao, Yingying Zhang, Yan Ji, Sailing He, and Xiaobing Zou. 2021. "The Development of Brain Network in Males with Autism Spectrum Disorders from Childhood to Adolescence: Evidence from fNIRS Study" Brain Sciences 11, no. 1: 120. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci11010120