
The Effects of Attention on the Syllable-Induced Prepulse Inhibition of the Startle Reflex and Cortical EEG Responses against Energetic or Informational Masking in Humans


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
2.1. Participants
2.2. Materials and Apparatus
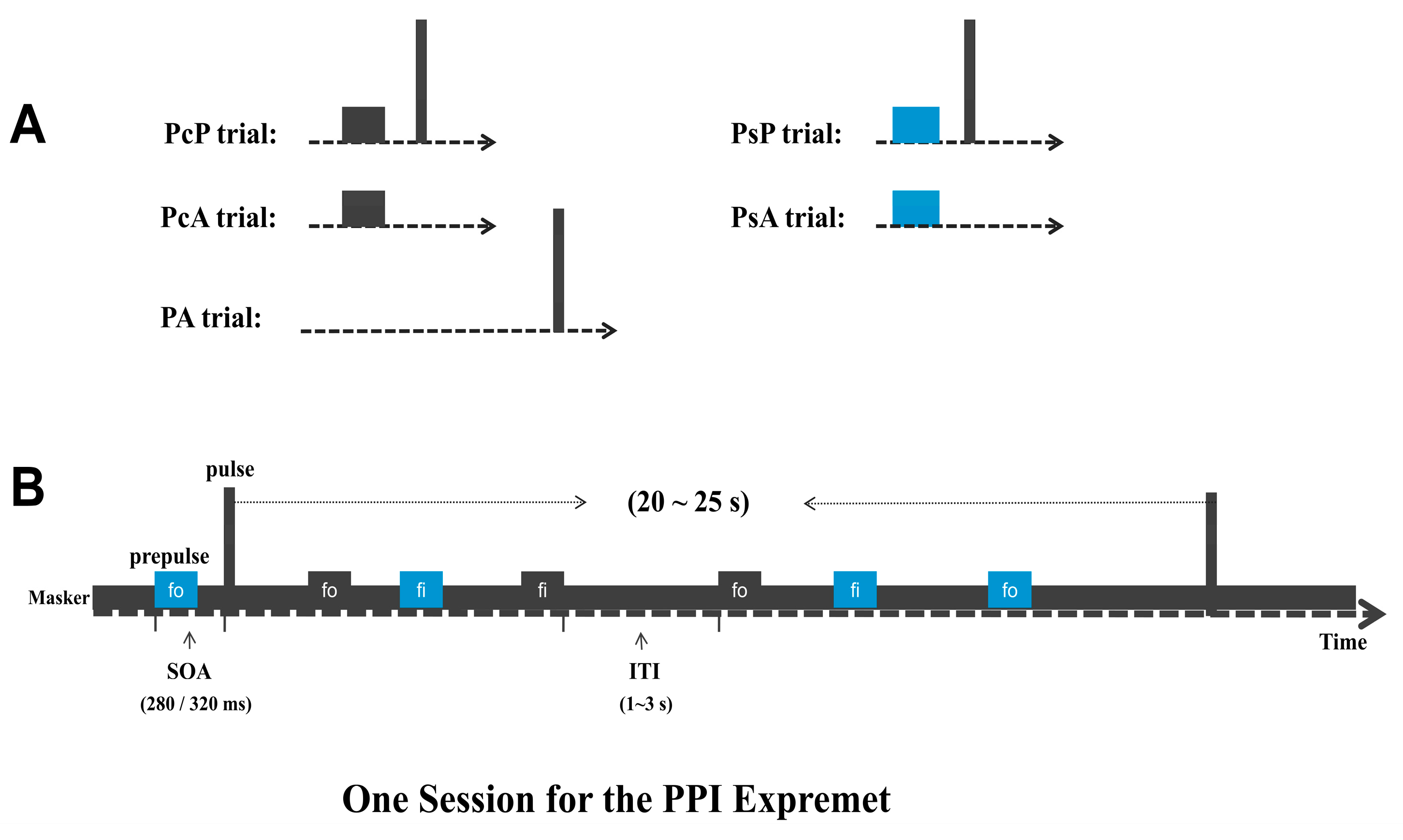
2.3. Procedures
2.4. Electroencephalogram (EEG) Data Acquisition and Processing
2.5. Startle Electromyogram (EMG) Signal Acquisition and Processing
3. Results
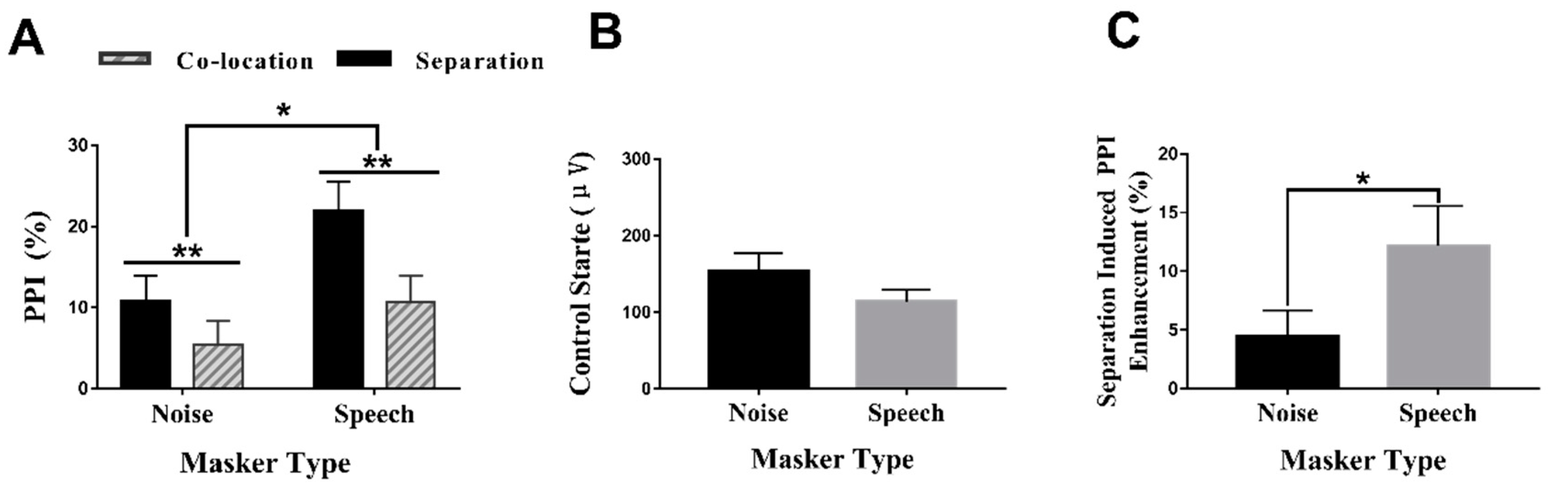
3.1. PPI under Different Conditions
3.2. Control (No Prepulse) Startle Reactivity and Separation-Induced PPI Enhancement (%) under Different Masker Types
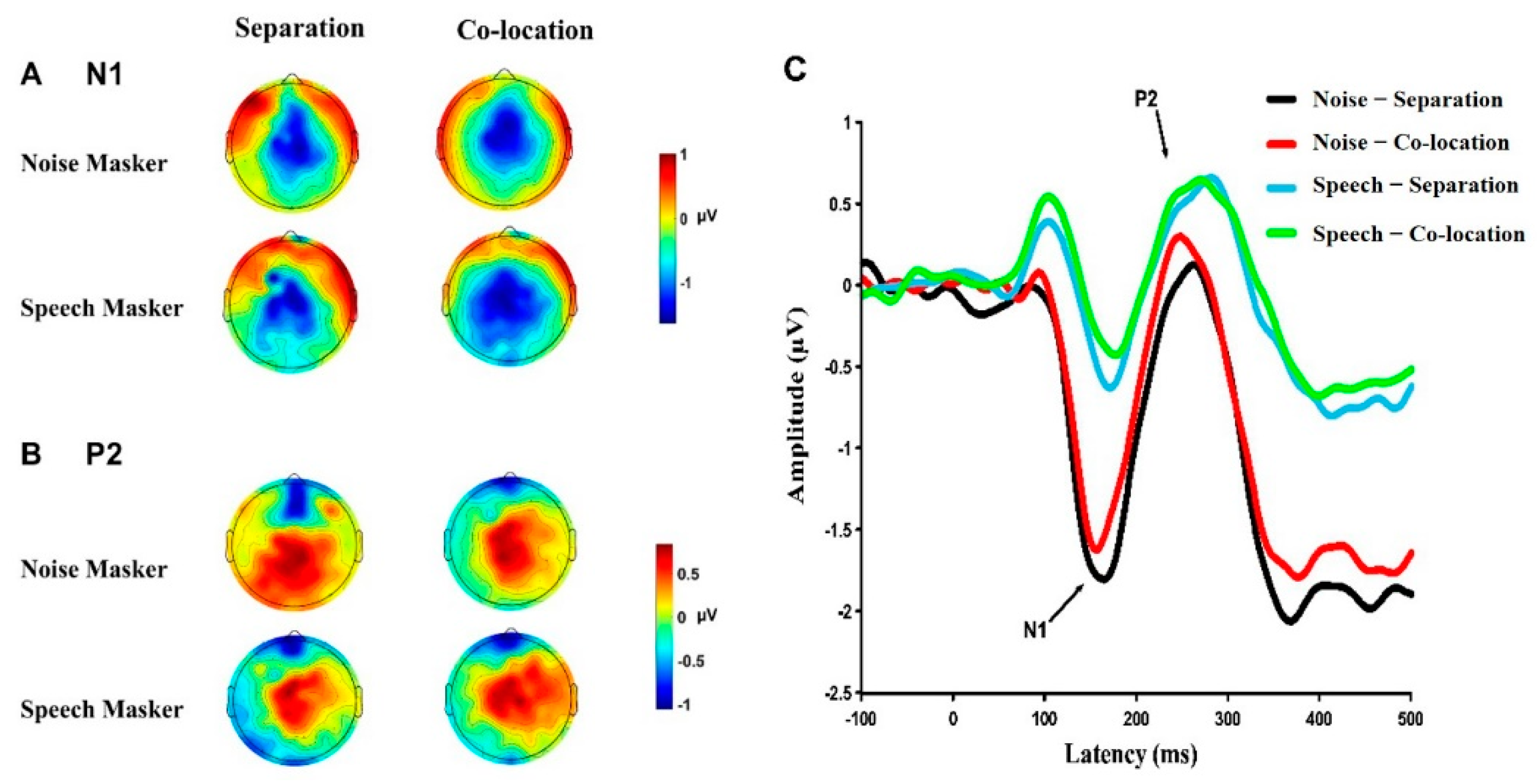
3.3. Cortical Responses under Different Conditions
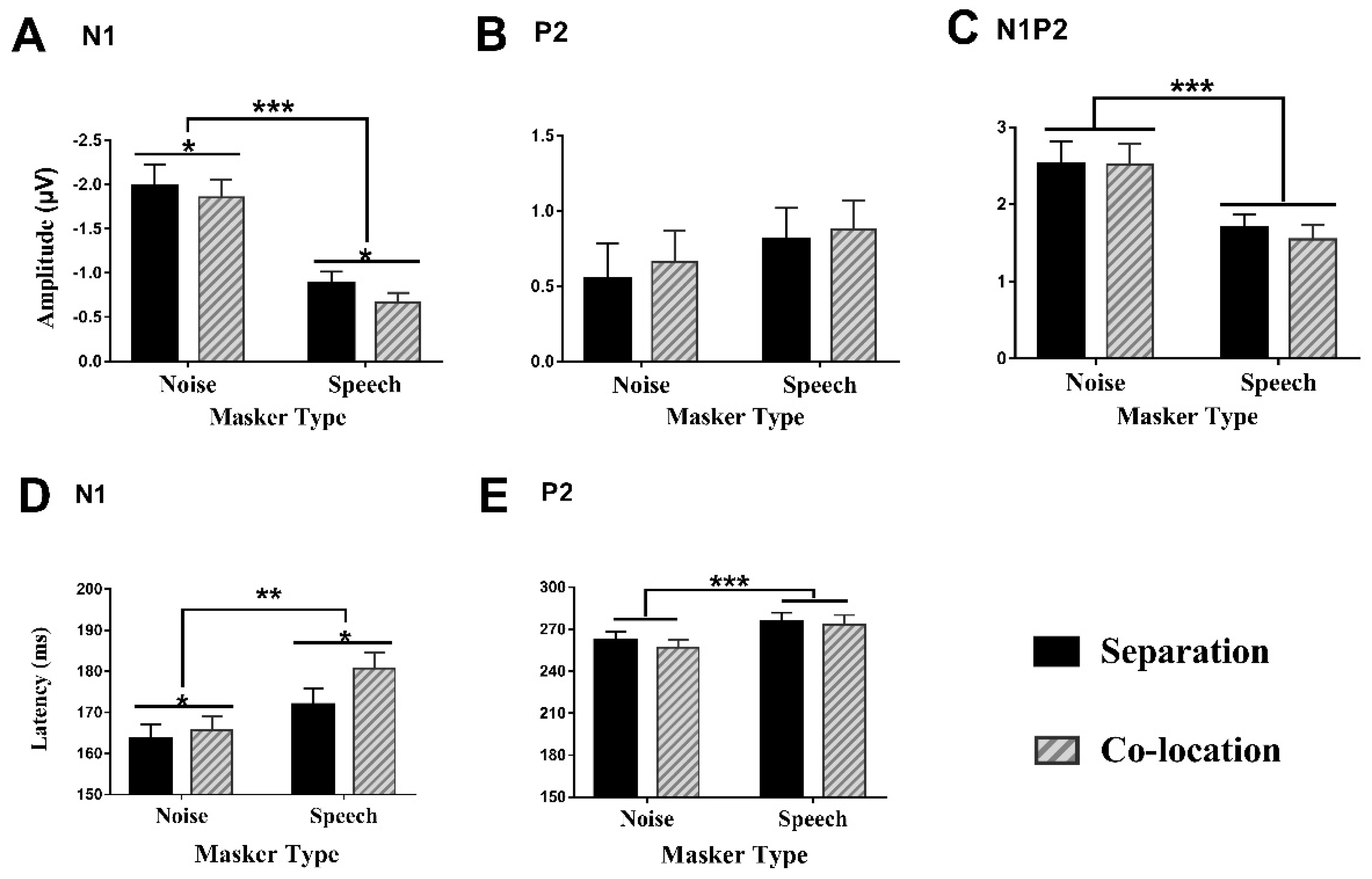
3.3.1. Amplitudes of the N1 Component
3.3.2. Amplitudes of the P2 Component
3.3.3. Amplitudes of the N1P2 Complex
3.3.4. Latencies of the N1 Component
3.3.5. Latencies of the P2 Component
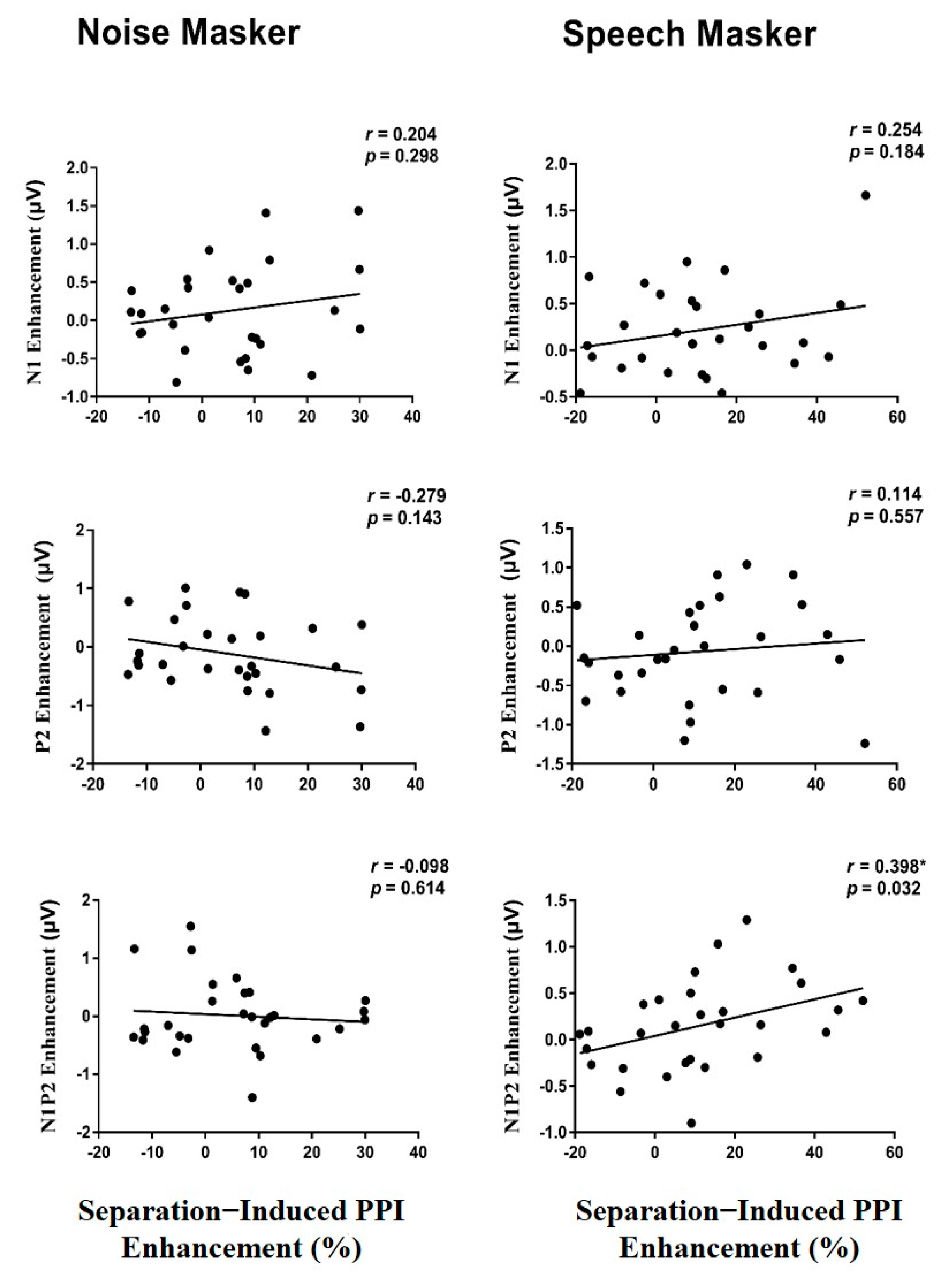
3.4. Correlation between PPI or Startle Reflex and ERP
3.5. Correlation between the PPI Enhancement and the ERP Enhancement
4. Discussion
4.1. Effects of Perceptual Separation
4.2. Effects of Masker Type
4.3. Relations between PPI and Cortical Responses to the Prepulse
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Li, L.; Du, Y.; Li, N.; Wu, X.; Wu, Y. Top-down modulation of prepulse inhibition of the startle reflex in humans and rats. Neurosci. Biobehav. Rev. 2009, 33, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, J.S.; Li, L.; Scott, B.W.; Frankland, P.W. Tactile, acoustic and vestibular systems sum to elicit the startle reflex. Neurosci. Biobehav. Rev. 2002, 26, 1–11. [Google Scholar] [CrossRef]
- Fendt, M.; Li, L.; Yeomans, J.S. Brain stem circuits mediating prepulse inhibition of the startle reflex. Psychopharmacology 2001, 156, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Swerdlow, N.R.; Sprock, J.; Light, G.A.; Cadenhead, K.; Calkins, M.E.; Dobie, D.J.; Freedman, R.; Green, M.F.; Greenwood, T.A.; Gur, R.E.; et al. Multi-site studies of acoustic startle and prepulse inhibition in humans: Initial experience and methodological considerations based on studies by the Consortium on the Genetics of Schizophrenia. Schizophr. Res. 2007, 92, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Dawson, M.E.; Schell, A.M.; Hazlett, E.A.; Nuechterlein, K.H.; Filion, D.L. On the clinical and cognitive meaning of impaired sensorimotor gating in schizophrenia. Psychiatry Res. 2000, 96, 187–197. [Google Scholar] [CrossRef]
- Dawson, M.E.; Hazlett, E.A.; Filion, D.L.; Nuechterlein, K.H.; Schell, A.M. Attention and schizophrenia: Impaired modulation of the startle reflex. J. Abnorm. Psychol. 1993, 102, 633–641. [Google Scholar] [CrossRef]
- Filion, D.L.; Poje, A.B. Selective and nonselective attention effects on prepulse inhibition of startle: A comparison of task and no-task protocols. Biol. Psychol. 2003, 64, 283–296. [Google Scholar] [CrossRef]
- Ding, Y.; Xu, N.; Gao, Y.; Wu, Z.; Li, L. The role of the deeper layers of the superior colliculus in attentional modulations of prepulse inhibition. Behav. Brain Res. 2019, 364, 106–113. [Google Scholar] [CrossRef]
- Lei, M.; Luo, L.; Qu, T.; Jia, H.; Li, L. Perceived location specificity in perceptual separation-induced but not fear conditioning-induced enhancement of prepulse inhibition in rats. Behav. Brain Res. 2014, 269, 87–94. [Google Scholar] [CrossRef]
- Lei, M.; Zhang, C.; Li, L. Neural correlates of perceptual separation-induced enhancement of prepulse inhibition of startle in humans. Sci. Rep. 2018, 8, 472. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Ding, Y.; Chen, B.; Gao, Y.; Wang, Q.; Wu, Z.; Lu, L.; Luo, L.; Zhang, C.; Bao, X.; et al. Both Val158Met Polymorphism of Catechol-O-Methyltransferase Gene and Menstrual Cycle Affect Prepulse Inhibition but Not Attentional Modulation of Prepulse Inhibition in Younger-Adult Females. Neuroscience 2019, 404, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.M.; Ding, Y.; Jia, H.X.; Li, L. Different effects of isolation-rearing and neonatal MK-801 treatment on attentional modulations of prepulse inhibition of startle in rats. Psychopharmacology 2016, 233, 3089–3102. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Wu, X.; Li, L. Emotional learning enhances stimulus-specific top-down modulation of sensorimotor gating in socially reared rats but not isolation-reared rats. Behav. Brain Res. 2010, 206, 192–201. [Google Scholar] [CrossRef] [PubMed]
- Brungart, D.S.; Simpson, B.D.; Freyman, R.L. Precedence-based speech segregation in a virtual auditory environment. J. Acoust. Soc. Am. 2005, 118, 3241–3251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Qi, J.G.; He, Y.; Alain, C.; Schneider, B.A. Attribute capture in the precedence effect for long-duration noise sounds. Hear. Res. 2005, 202, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Freyman, R.L.; Helfer, K.S.; McCall, D.D.; Clifton, R.K. The role of perceived spatial separation in the unmasking of speech. J. Acoust. Soc. Am. 1999, 106, 3578–3588. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Huang, Q.; Chen, X.; Qu, T.; Wu, X.; Li, L. Perceptual integration between target speech and target-speech reflection reduces masking for target-speech recognition in younger adults and older adults. Hear. Res. 2008, 244, 51–65. [Google Scholar] [CrossRef]
- Li, L.; Daneman, M.; Qi, J.G.; Schneider, B.A. Does the information content of an irrelevant source differentially affect spoken word recognition in younger and older adults? J. Exp. Psychol. Hum. Percept. Perform. 2004, 30, 1077–1091. [Google Scholar] [CrossRef] [Green Version]
- Schneider, B.A.; Li, L.; Daneman, M. How competing speech interferes with speech comprehension in everyday listening situations. J. Am. Acad. Audiol. 2007, 18, 559–572. [Google Scholar] [CrossRef] [Green Version]
- Brungart, D.S. Informational and energetic masking effects in the perception of two simultaneous talkers. J. Acoust. Soc. Am. 2001, 109, 1101–1109. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, C.; Chen, J.; Qu, H.; Li, W.; Wu, Y.; Schneider, B.A.; Li, L. The effect of perceived spatial separation on informational masking of Chinese speech. Hear. Res. 2005, 199, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Freyman, R.L.; Balakrishnan, U.; Helfer, K.S. Spatial release from informational masking in speech recognition. J. Acoust. Soc. Am. 2001, 109 Pt 1, 2112–2122. [Google Scholar] [CrossRef]
- Billings, C.J.; Bennett, K.O.; Molis, M.R.; Leek, M.R. Cortical encoding of signals in noise: Effects of stimulus type and recording paradigm. Ear Hear. 2011, 32, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Arnott, S.R.; Rabaglia, C.; Avivi-Reich, M.; Qi, J.; Wu, X.; Li, L.; Schneider, B.A. Attentional modulation of informational masking on early cortical representations of speech signals. Hear. Res. 2016, 331, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Parkkonen, L.; Wei, J.; Dong, J.R.; Ma, Y.; Carlson, S. Prepulse Inhibition of Auditory Cortical Responses in the Caudolateral Superior Temporal Gyrus in Macaca mulatta. Neurosci. Bull. 2018, 34, 291–302. [Google Scholar] [CrossRef]
- Inui, K.; Tsuruhara, A.; Kodaira, M.; Motomura, E.; Tanii, H.; Nishihara, M.; Keceli, S.; Kakigi, R. Prepulse inhibition of auditory change-related cortical responses. BMC Neurosci. 2012, 13, 135. [Google Scholar] [CrossRef] [Green Version]
- Ford, J.M.; Roth, W.T.; Menon, V.; Pfefferbaum, A. Failures of automatic and strategic processing in schizophrenia: Comparisons of event-related brain potential and startle blink modification. Schizophr. Res. 1999, 37, 149–163. [Google Scholar] [CrossRef]
- De Pascalis, V.; Cozzuto, G.; Russo, E. Effects of personality trait emotionality on acoustic startle response and prepulse inhibition including N100 and P200 event-related potential. Clin. Neurophysiol. 2013, 124, 292–305. [Google Scholar] [CrossRef]
- Inui, K.; Nakagawa, K.; Nishihara, M.; Motomura, E.; Kakigi, R. Inhibition in the Human Auditory Cortex. PLoS ONE 2016, 11, e0155972. [Google Scholar] [CrossRef]
- Kedzior, K.K.; Koch, M.; Basar-Eroglu, C. Prepulse inhibition (PPI) of auditory startle reflex is associated with PPI of auditory-evoked theta oscillations in healthy humans. Neurosci. Lett. 2006, 400, 246–251. [Google Scholar] [CrossRef]
- Zhang, C.; Lu, L.; Wu, X.; Li, L. Attentional modulation of the early cortical representation of speech signals in informational or energetic masking. Brain Lang. 2014, 135, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Fan, M.; Sun, B.; Wang, R.; Mo, L.; Zhang, J.X. Non-target language processing in Chinese-English bilinguals: A study of event-related potential. Psychol. Rep. 2012, 110, 935–945. [Google Scholar] [PubMed]
- Zhang, C.; Tao, R.; Zhao, H. Auditory spatial attention modulates the unmasking effect of perceptual separation in a “cocktail party” environment. Neuropsychologia 2019, 124, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, K.; Ross, B. Effects of age and age-related hearing loss on the brain. J. Commun. Disord. 2007, 40, 305–312. [Google Scholar] [CrossRef]
- Morales-Muñoz, I.; Jurado-Barba, R.; Fernández-Guinea, S.; Rodríguez-Jiménez, R.; Jiménez-Arriero, M.; Criado, J.R.; Rubio, G. Sensory Gating Deficits in First-Episode Psychosis: Evidence From Neurophysiology, Psychophysiology, and Neuropsychology. J. Nerv. Ment. Dis. 2016, 204, 877–884. [Google Scholar] [CrossRef]
- Annic, A.; Bocquillon, P.; Bourriez, J.L.; Derambure, P.; Dujardin, K. Effects of stimulus-driven and goal-directed attention on prepulse inhibition of the cortical responses to an auditory pulse. Clin. Neurophysiol. 2014, 125, 1576–1588. [Google Scholar] [CrossRef]
- Abduljawad, K.A.; Langley, R.W.; Bradshaw, C.M.; Szabadi, E. Effects of bromocriptine and haloperidol on prepulse inhibition: Comparison of the acoustic startle eyeblink response and the N1/P2 auditory-evoked response in man. J. Psychopharmacol. 1999, 13, 3–9. [Google Scholar] [CrossRef]
- San-Martin, R.; Zimiani, M.I.; Noya, C.; Ávila, M.A.V.; Shuhama, R.; Del-Ben, C.M.; Menezes, P.R.; Fraga, F.J.; Salum, C. A Method for Simultaneous Evaluation of Muscular and Neural Prepulse Inhibition. Front. Neurosci. 2018, 12, 654. [Google Scholar] [CrossRef] [Green Version]
- Graham, S.J.; Langley, R.W.; Bradshaw, C.M.; Szabadi, E. Effects of haloperidol and clozapine on prepulse inhibition of the acoustic startle response and the N1/P2 auditory evoked potential in man. J. Psychopharmacol. 2001, 15, 243–250. [Google Scholar] [CrossRef]
- Koch, M. The neurobiology of startle. Prog. Neurobiol. 1999, 59, 107–128. [Google Scholar] [CrossRef]
- Bendor, D.; Wang, X. The neuronal representation of pitch in primate auditory cortex. Nature 2005, 436, 1161–1165. [Google Scholar] [CrossRef] [PubMed]
- Theunissen, F.E.; Sen, K.; Doupe, A.J. Spectral-temporal receptive fields of nonlinear auditory neurons obtained using natural sounds. J. Neurosci. 2000, 20, 2315–2331. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Liu, L.; Yang, P.; Ding, Y.; Wang, C.; Li, L. The Effects of Attention on the Syllable-Induced Prepulse Inhibition of the Startle Reflex and Cortical EEG Responses against Energetic or Informational Masking in Humans. Brain Sci. 2022, 12, 660. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050660
Yang X, Liu L, Yang P, Ding Y, Wang C, Li L. The Effects of Attention on the Syllable-Induced Prepulse Inhibition of the Startle Reflex and Cortical EEG Responses against Energetic or Informational Masking in Humans. Brain Sciences. 2022; 12(5):660. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050660
Chicago/Turabian StyleYang, Xiaoqin, Lei Liu, Pengcheng Yang, Yu Ding, Changming Wang, and Liang Li. 2022. "The Effects of Attention on the Syllable-Induced Prepulse Inhibition of the Startle Reflex and Cortical EEG Responses against Energetic or Informational Masking in Humans" Brain Sciences 12, no. 5: 660. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci12050660