Upregulation of IL-1 Receptor Antagonist in a Mouse Model of Migraine

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Dataset Selection and Analysis
2.2. Statistical Analysis
3. Results
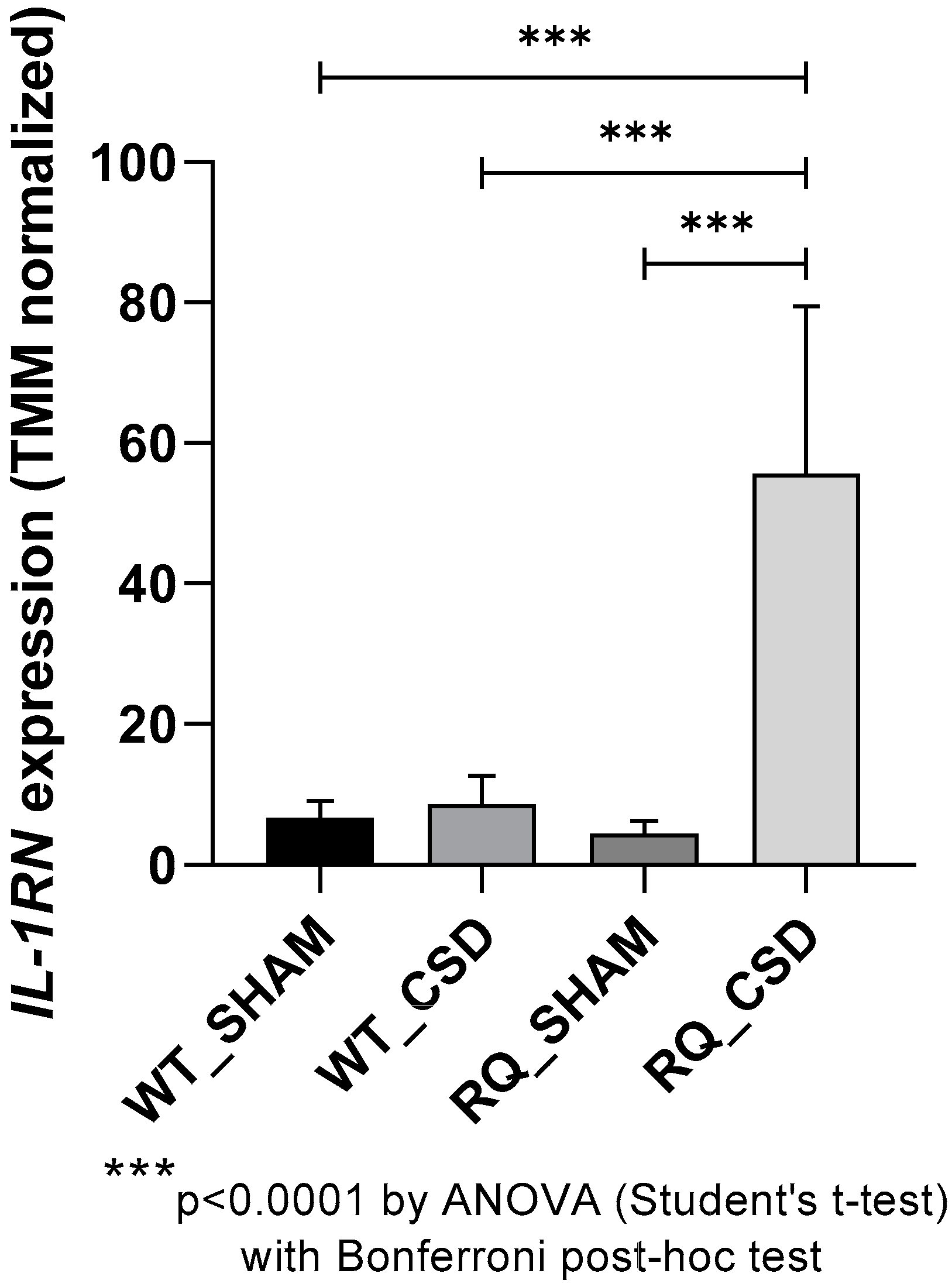
3.1. IL-1RN Expression after CSD
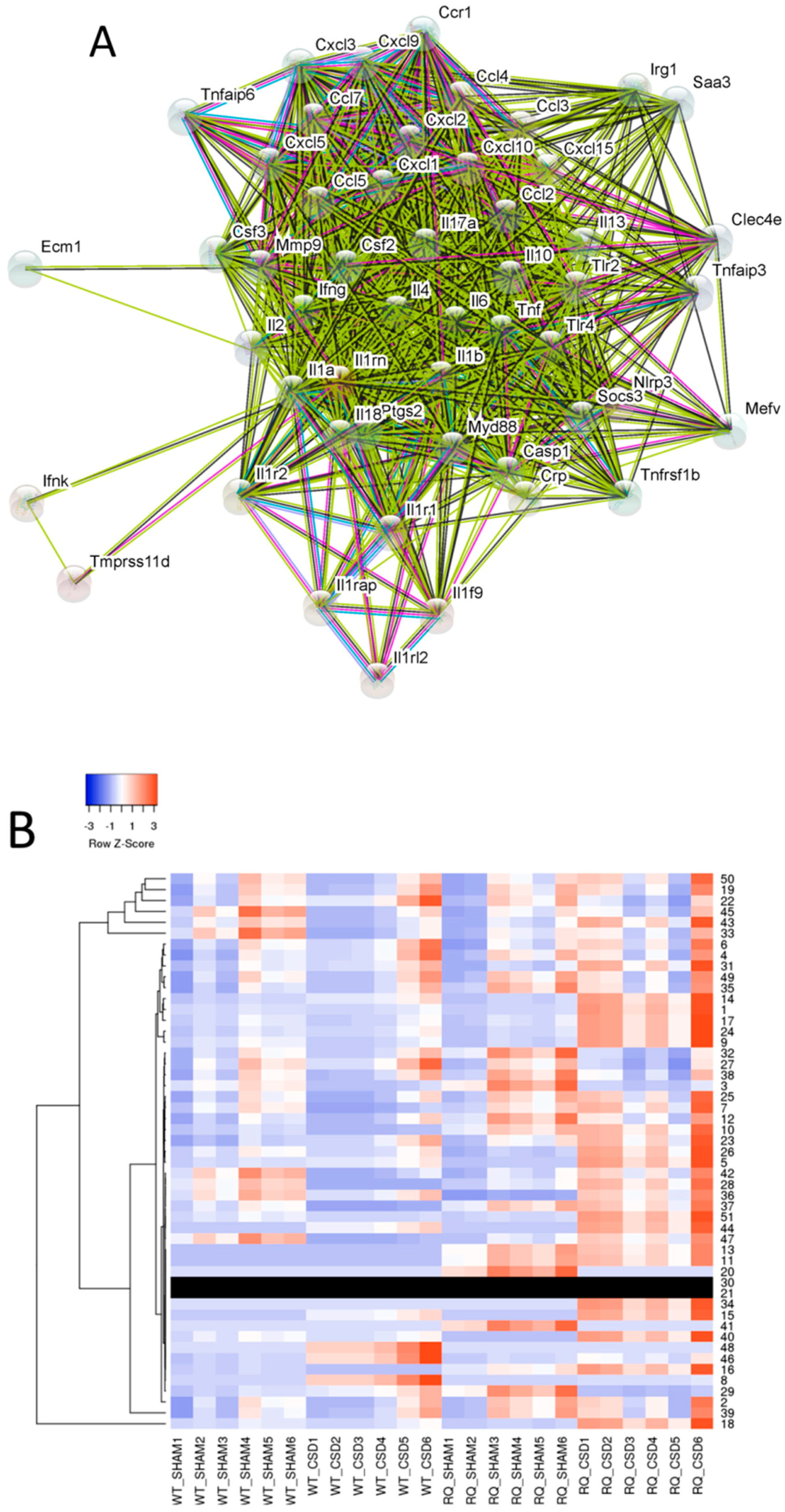
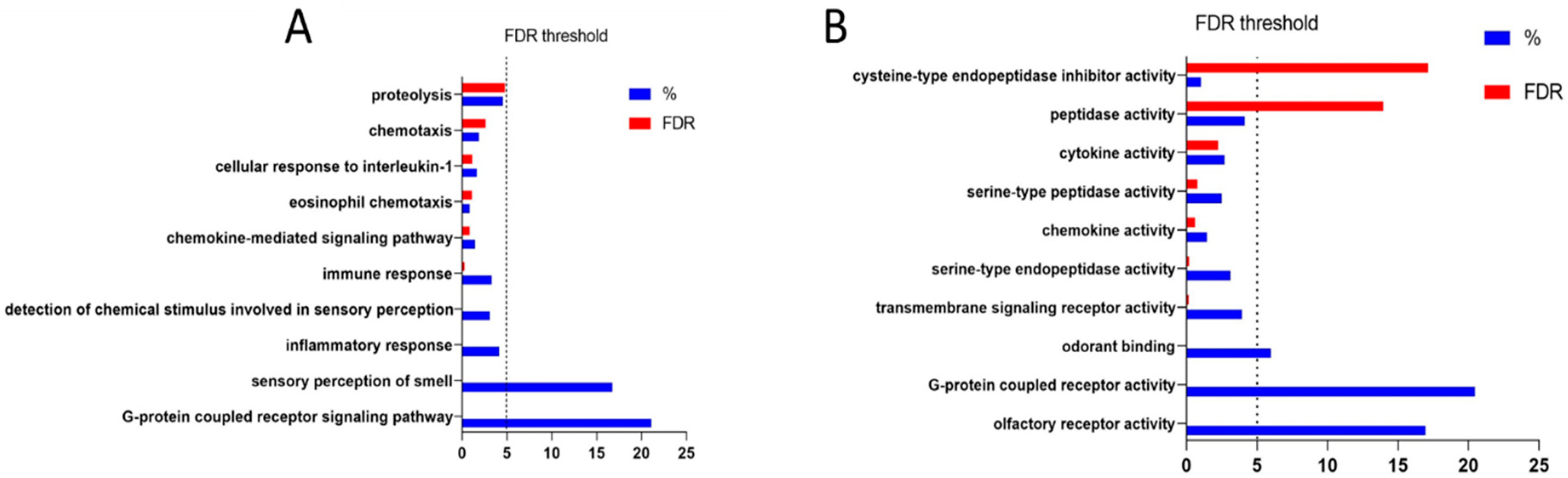
3.2. Identification of Genes Functionally Related to IL-1RN
3.3. Identification of Genes Statistically Correlated to IL-1RN
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Sample Availability
References
- Stovner, L.J.; Hagen, K.; Jensen, R.; Katsarava, Z.; Lipton, R.B.; Scher, A.I.; Steiner, T.J.; Zwart, J.A. The global burden of headache: A documentation of headache prevalence and disability worldwide. Cephalalgia 2007, 27, 193–210. [Google Scholar] [CrossRef] [PubMed]
- Vetvik, K.G.; MacGregor, E.A. Sex differences in the epidemiology, clinical features, and pathophysiology of migraine. Lancet Neurol. 2017, 16, 76–87. [Google Scholar] [CrossRef]
- Russell, M.B.; Rasmussen, B.K.; Thorvaldsen, P.; Olesen, J. Prevalence and sex-ratio of the subtypes of migraine. Int. J. Epidemiol. 1995. [Google Scholar] [CrossRef] [PubMed]
- Headache Classification Committee of the International Headache Society (IHS). The International Classification of Headache Disorders, 3rd edition (beta version). Cephalalgia 2013. [Google Scholar] [CrossRef]
- Lauritzen, M. Pathophysiology of the migraine aura: The spreading depression theory. Brain 1994, 117 Pt 1, 199–210. [Google Scholar] [CrossRef]
- Cui, Y.; Kataoka, Y.; Watanabe, Y. Role of cortical spreading depression in the pathophysiology of migraine. Neurosci. Bull. 2014, 30, 812–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longoni, M.; Ferrarese, C. Inflammation and excitotoxicity: Role in migraine pathogenesis. Neurol. Sci. 2006. [Google Scholar] [CrossRef]
- Edvinsson, L.; Haanes, K.A.; Warfvinge, K. Does inflammation have a role in migraine? Nat. Rev. Neurol. 2019. [Google Scholar] [CrossRef]
- Cacabelos, R.; Torrellas, C.; Fernández-Novoa, L.; López-Muñoz, F. Histamine and Immune Biomarkers in CNS Disorders. Mediat. Inflamm. 2016, 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Roeleveld, D.M.; Koenders, M.I. The role of the Th17 cytokines IL-17 and IL-22 in Rheumatoid Arthritis pathogenesis and developments in cytokine immunotherapy. Cytokine 2015, 74, 101–107. [Google Scholar] [CrossRef]
- Zhang, H.-L.; Zheng, X.-Y.; Zhu, J. Th1/Th2/Th17/Treg cytokines in Guillain–Barré syndrome and experimental autoimmune neuritis. Cytokine Growth Factor Rev. 2013, 24, 443–453. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Kubatzky, K.F.; Mitra, D.K. An Update on Interleukin-9: From Its Cellular Source and Signal Transduction to Its Role in Immunopathogenesis. Int. J. Mol. Sci. 2019, 20, 2113. [Google Scholar] [CrossRef] [PubMed]
- Dujmovic, I.; Mangano, K.; Pekmezovic, T.; Quattrocchi, C.; Mesaros, S.; Stojsavljevic, N.; Nicoletti, F.; Drulovic, J. The analysis of IL-1 beta and its naturally occurring inhibitors in multiple sclerosis: The elevation of IL-1 receptor antagonist and IL-1 receptor type II after steroid therapy. J. Neuroimmunol. 2009, 207, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Ayakannu, R.; Abdullah, N.A.; Radhakrishnan, A.K.; Lechimi Raj, V.; Liam, C.K. Relationship between various cytokines implicated in asthma. Hum. Immunol. 2019. [Google Scholar] [CrossRef]
- Nicoletti, F.; Mancuso, G.; Cusumano, V.; Di Marco, R.; Zaccone, P.; Bendtzen, K.; Teti, G. Prevention of endotoxin-induced lethality in neonatal mice by interleukin-13. Eur. J. Immunol. 1997, 27, 1580–1583. [Google Scholar] [CrossRef] [PubMed]
- Conway, T.F.; Hammer, L.; Furtado, S.; Mathiowitz, E.; Nicoletti, F.; Mangano, K.; Egilmez, N.K.; Auci, D.L. Oral Delivery of Particulate Transforming Growth Factor Beta 1 and All-Trans Retinoic Acid Reduces Gut Inflammation in Murine Models of Inflammatory Bowel Disease. J. Crohn’s Colitis 2015, 9, 647–658. [Google Scholar] [CrossRef]
- Nicoletti, F.; Di Marco, R.; Patti, F.; Reggio, E.; Nicoletti, A.; Zaccone, P.; Stivala, F.; Meroni, P.L.; Reggio, A. Blood levels of transforming growth factor-beta 1 (TGF-beta1) are elevated in both relapsing remitting and chronic progressive multiple sclerosis (MS) patients and are further augmented by treatment with interferon-beta 1b (IFN-beta1b). Clin. Exp. Immunol. 1998, 113, 96–99. [Google Scholar] [CrossRef]
- Barcellini, W.; Rizzardi, G.P.; Borghi, M.O.; Nicoletti, F.; Fain, C.; Del Papa, N.; Meroni, P.L. In vitro type-1 and type-2 cytokine production in systemic lupus erythematosus: Lack of relationship with clinical disease activity. Lupus 1996, 5, 139–145. [Google Scholar] [CrossRef]
- Fiotti, N.; Giansante, C.; Ponte, E.; Delbello, C.; Calabrese, S.; Zacchi, T.; Dobrina, A.; Guarnieri, G. Atherosclerosis and inflammation. Patterns of cytokine regulation in patients with peripheral arterial disease. Atherosclerosis 1999, 145, 51–60. [Google Scholar] [CrossRef]
- Nielsen, C.H.; Bendtzen, K. Immunoregulation by Naturally Occurring and Disease-Associated Autoantibodies: Binding to cytokines and their role in regulation of T-cell responses. Adv. Exp. Med. Biol. 2012, 750, 116–132. [Google Scholar] [CrossRef]
- Arend, W.P.; Malyak, M.; Guthridge, C.J.; Gabay, C. INTERLEUKIN-1 RECEPTOR ANTAGONIST: Role in Biology. Annu. Rev. Immunol. 2002. [Google Scholar] [CrossRef]
- Nicoletti, F.; Patti, F.; Di Marco, R.; Zaccone, P.; Nicoletti, A.; Meroni, P.L.; Reggio, A. Circulating serum levels of IL-1ra in patients with relapsing remitting multiple sclerosis are normal during remission phases but significantly increased either during exacerbations or in response to IFN-β treatment. Cytokine 1996. [Google Scholar] [CrossRef]
- Perini, F.; D’Andrea, G.; Galloni, E.; Pignatelli, F.; Billo, G.; Alba, S.; Bussone, G.; Toso, V. Plasma cytokine levels in migraineurs and controls. Headache 2005. [Google Scholar] [CrossRef] [PubMed]
- Empl, M.; Sostak, P.; Riedel, M.; Schwarz, M.; Müller, N.; Förderreuther, S.; Straube, A. Decreased sTNF-RI in migraine patients? Cephalalgia 2003. [Google Scholar] [CrossRef]
- Covelli, V.; Munno, I.; Pellegrino, N.M.; Di Venere, A.; Jirillo, E.; Buscaino, G.A. Exaggerated spontaneous release of tumor necrosis factor-alpha/cachectin in patients with migraine without aura. Acta Neurol. (Napoli) 1990, 12, 257–263. [Google Scholar]
- Yilmaz, I.A.; Özge, A.; Erdal, M.E.; Edgünlü, T.G.; Çakmak, S.E.; Yalin, O.Ö. Cytokine polymorphism in patients with migraine: Some suggestive clues of migraine and inflammation. Pain Med. 2010. [Google Scholar] [CrossRef] [PubMed]
- Skaper, S.D. Nerve growth factor: A neuroimmune crosstalk mediator for all seasons. Immunology 2017, 151, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, M.A. Neurogenic inflammation in the pathophysiology and treatment of migraine. Neurology 1993, 43 (Suppl. S3), S16–S20. [Google Scholar] [PubMed]
- Waeber, C.; Moskowitz, M.A. Migraine as an inflammatory disorder. Neurology 2012. [Google Scholar] [CrossRef]
- Vecchia, D.; Pietrobon, D. Migraine: A disorder of brain excitatory-inhibitory balance? Trends Neurosci. 2012, 35, 507–520. [Google Scholar] [CrossRef]
- Ceruti, S.; Villa, G.; Fumagalli, M.; Colombo, L.; Magni, G.; Zanardelli, M.; Fabbretti, E.; Verderio, C.; van den Maagdenberg, A.M.J.M.; Nistri, A.; et al. Calcitonin Gene-Related Peptide-Mediated Enhancement of Purinergic Neuron/Glia Communication by the Algogenic Factor Bradykinin in Mouse Trigeminal Ganglia from Wild-Type and R192Q Cav2.1 Knock-In Mice: Implications for Basic Mechanisms of Migraine Pain. J. Neurosci. 2011. [Google Scholar] [CrossRef]
- Friedman, M.H. Local inflammation as a mediator of migraine and tension-type headache. Headache 2004. [Google Scholar] [CrossRef]
- Rowe, B.H.; Colman, I.; Edmonds, M.L.; Blitz, S.; Walker, A.; Wiens, S. Randomized controlled trial of intravenous dexamethasone to prevent relapse in acute migraine headache. Headache 2008. [Google Scholar] [CrossRef]
- Van Den Maagdenberg, A.M.J.M.; Haan, J.; Terwindt, G.M.; Ferrari, M.D. Migraine: Gene mutations and functional consequences. Curr. Opin. Neurol. 2007, 20, 299–305. [Google Scholar] [CrossRef]
- Ophoff, R.A.; Terwindt, G.M.; Vergouwe, M.N.; Van Eijk, R.; Oefner, P.J.; Hoffman, S.M.G.; Lamerdin, J.E.; Mohrenweiser, H.W.; Bulman, D.E.; Ferrari, M.; et al. Familial hemiplegic migraine and episodic ataxia type-2 are caused by mutations in the Ca2+ channel gene CACNL1A4. Cell 1996. [Google Scholar] [CrossRef]
- Van Den Maagdenberg, A.M.J.M.; Pietrobon, D.; Pizzorusso, T.; Kaja, S.; Broos, L.A.M.; Cesetti, T.; Van De Ven, R.C.G.; Tottene, A.; Van Der Kaa, J.; Plomp, J.J.; et al. A Cacna1a knockin migraine mouse model with increased susceptibility to cortical spreading depression. Neuron 2004. [Google Scholar] [CrossRef]
- Jurkat-Rott, K.; Lerche, H.; Weber, Y.; Lehmann-Horn, F. Hereditary channelopathies in neurology. Adv. Exp. Med. Biol. 2010, 686, 305–334. [Google Scholar]
- Tfelt-Hansen, P.C.; Koehler, P.J. One hundred years of migraine research: Major clinical and scientific observations from 1910 to 2010. Headache 2011, 51, 752–778. [Google Scholar] [CrossRef]
- Tottene, A.; Conti, R.; Fabbro, A.; Vecchia, D.; Shapovalova, M.; Santello, M.; van den Maagdenberg, A.M.J.M.; Ferrari, M.D.; Pietrobon, D. Enhanced Excitatory Transmission at Cortical Synapses as the Basis for Facilitated Spreading Depression in CaV2.1 Knockin Migraine Mice. Neuron 2009. [Google Scholar] [CrossRef]
- Karatas, H.; Erdener, S.E.; Gursoy-Ozdemir, Y.; Lule, S.; Eren-Koçak, E.; Sen, Z.D.; Dalkara, T. Spreading depression triggers headache by activating neuronal Panx1 channels. Science 2013, 339, 1092–1095. [Google Scholar] [CrossRef]
- Noseda, R.; Burstein, R. Migraine pathophysiology: Anatomy of the trigeminovascular pathway and associated neurological symptoms, cortical spreading depression, sensitization, and modulation of pain. Pain 2013, 154 (Suppl. S1), S44–S53. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Levy, D.; Kainz, V.; Noseda, R.; Jakubowski, M.; Burstein, R. Activation of central trigeminovascular neurons by cortical spreading depression. Ann. Neurol. 2011. [Google Scholar] [CrossRef]
- GEO DataSets. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/gds (accessed on 15 April 2019).
- Eising, E.; Shyti, R.; ’t Hoen, P.A.C.; Vijfhuizen, L.S.; Huisman, S.M.H.; Broos, L.A.M.; Mahfouz, A.; Reinders, M.J.T.; Ferrari, M.D.; Tolner, E.A.; et al. Cortical Spreading Depression Causes Unique Dysregulation of Inflammatory Pathways in a Transgenic Mouse Model of Migraine. Mol. Neurobiol. 2017. [Google Scholar] [CrossRef]
- STRING. Available online: https://string-db.org/ (accessed on 15 April 2019).
- Heatmapper. Available online: http://www.heatmapper.ca/ (accessed on 15 April 2019).
- CoExpress software. Available online: http://www. bioinformatics.lu/coexpress/ (accessed on 15 April 2019).
- DAVID version 6.8. Available online: http://david.abcc.ncifcrf.gov/ (accessed on 15 April 2019).
- IBM SPSS Statistics 23. Available online: https://www.spss.it/ (accessed on 15 April 2019).
- GraphPad Prism 8.0.2. Available online: https://www.graphpad.com/ (accessed on 15 April 2019).
- Malhotra, R. Understanding migraine: Potential role of neurogenic inflammation. Ann. Indian Acad. Neurol. 2016. [Google Scholar] [CrossRef]
- Ramachandran, R. Neurogenic inflammation and its role in migraine. Semin. Immunopathol. 2018, 40, 301–314. [Google Scholar] [CrossRef]
- Johnston, M.M.; Rapoport, A.M. Triptans for the management of migraine. Drugs 2010, 70, 1505–1518. [Google Scholar] [CrossRef]
- Oliveira, A.B.; Bachi, A.L.L.; Ribeiro, R.T.; Mello, M.T.; Tufik, S.; Peres, M.F.P. Unbalanced plasma TNF-α and IL-12/IL-10 profile in women with migraine is associated with psychological and physiological outcomes. J. Neuroimmunol. 2017, 313, 138–144. [Google Scholar] [CrossRef]
- Costa, C.; Tozzi, A.; Rainero, I.; Cupini, L.M.; Calabresi, P.; Ayata, C.; Sarchielli, P. Cortical spreading depression as a target for anti-migraine agents. J. Headache Pain 2013. [Google Scholar] [CrossRef]
- Thompson, C.S.; Hakim, A.M. Cortical Spreading Depression Modifies Components of the Inflammatory Cascade. Mol. Neurobiol. 2005. [Google Scholar] [CrossRef]
- Choudhuri, R.; Cui, L.; Yong, C.; Bowyer, S.; Klein, R.M.; Welch, K.M.A.; Berman, N.E.J. Cortical spreading depression and gene regulation: Relevance to migraine. Ann. Neurol. 2002. [Google Scholar] [CrossRef]
- Urbach, A.; Bruehl, C.; Witte, O.W. Microarray-based long-term detection of genes differentially expressed after cortical spreading depression. Eur. J. Neurosci. 2006. [Google Scholar] [CrossRef]
- Fagone, P.; Mazzon, E.; Cavalli, E.; Bramanti, A.; Petralia, M.C.; Mangano, K.; Al-Abed, Y.; Bramati, P.; Nicoletti, F. Contribution of the macrophage migration inhibitory factor superfamily of cytokines in the pathogenesis of preclinical and human multiple sclerosis: In silico and in vivo evidences. J. Neuroimmunol. 2018, 322. [Google Scholar] [CrossRef]
- Fagone, P.; Mazzon, E.; Mammana, S.; Di Marco, R.; Spinasanta, F.; Basile, M.S.; Petralia, M.C.; Bramanti, P.; Nicoletti, F.; Mangano, K. Identification of CD4+ T cell biomarkers for predicting the response of patients with relapsing-remitting multiple sclerosis to natalizumab treatment. Mol. Med. Rep. 2019, 20, 678–684. [Google Scholar] [CrossRef]
- Mammana, S.; Bramanti, P.; Mazzon, E.; Cavalli, E.; Basile, M.S.; Fagone, P.; Petralia, M.C.; McCubrey, J.A.; Nicoletti, F.; Mangano, K. Preclinical evaluation of the PI3K/Akt/mTOR pathway in animal models of multiple sclerosis. Oncotarget 2018, 9, 8263–8277. [Google Scholar] [CrossRef] [Green Version]
- Fagone, P.; Muthumani, K.; Mangano, K.; Magro, G.; Meroni, P.L.; Kim, J.J.; Sardesai, N.Y.; Weiner, D.B.; Nicoletti, F. VGX-1027 modulates genes involved in lipopolysaccharide-induced Toll-like receptor 4 activation and in a murine model of systemic lupus erythematosus. Immunology 2014, 142. [Google Scholar] [CrossRef]
- Presti, M.; Mazzon, E.; Basile, M.S.; Petralia, M.C.; Bramanti, A.; Colletti, G.; Bramanti, P.; Nicoletti, F.; Fagone, P. Overexpression of macrophage migration inhibitory factor and functionally-related genes, D-DT, CD74, CD44, CXCR2 and CXCR4, in glioblastoma. Oncol. Lett. 2018, 16, 2881–2886. [Google Scholar] [CrossRef]
- Fagone, P.; Caltabiano, R.; Russo, A.; Lupo, G.; Anfuso, C.D.; Basile, M.S.; Longo, A.; Nicoletti, F.; De Pasquale, R.; Libra, M.; et al. Identification of novel chemotherapeutic strategies for metastatic uveal melanoma. Sci. Rep. 2017, 7, 44564. [Google Scholar] [CrossRef]
- Basile, M.S.; Mazzon, E.; Russo, A.; Mammana, S.; Longo, A.; Bonfiglio, V.; Fallico, M.; Caltabiano, R.; Fagone, P.; Nicoletti, F.; et al. Differential modulation and prognostic values of immune-escape genes in uveal melanoma. PLoS ONE 2019, 14, e0210276. [Google Scholar] [CrossRef]
- Mangano, K.; Mazzon, E.; Basile, M.S.; Di Marco, R.; Bramanti, P.; Mammana, S.; Petralia, M.C.; Fagone, P.; Nicoletti, F. Pathogenic role for macrophage migration inhibitory factor in glioblastoma and its targeting with specific inhibitors as novel tailored therapeutic approach. Oncotarget 2018, 9, 17951–17970. [Google Scholar] [CrossRef] [Green Version]
- Fagone, P.; Mangano, K.; Mammana, S.; Pesce, A.; Pesce, A.; Caltabiano, R.; Giorlandino, A.; Portale, T.R.; Cavalli, E.; Lombardo, G.A.G.; et al. Identification of novel targets for the diagnosis and treatment of liver fibrosis. Int. J. Mol. Med. 2015, 36, 747–752. [Google Scholar] [CrossRef]
- Mangano, K.; Cavalli, E.; Mammana, S.; Basile, M.S.; Caltabiano, R.; Pesce, A.; Puleo, S.; Atanasov, A.G.; Magro, G.; Nicoletti, F.; et al. Involvement of the Nrf2/HO-1/CO axis and therapeutic intervention with the CO-releasing molecule CORM-A1, in a murine model of autoimmune hepatitis. J. Cell. Physiol. 2018, 233, 4156–4165. [Google Scholar] [CrossRef]
- Fagone, P.; Nunnari, G.; Lazzara, F.; Longo, A.; Cambria, D.; Distefano, G.; Palumbo, M.; Nicoletti, F.; Malaguarnera, L.; Di Rosa, M. Induction of OAS gene family in HIV monocyte infected patients with high and low viral load. Antivir. Res. 2016, 131. [Google Scholar] [CrossRef]
- Arend, W.P.; Gabay, C. Physiologic role of interleukin-1 receptor antagonist. Arthritis Res. 2000, 2, 245–248. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.D.; Buchman, T.G. Interleukin-1 receptor antagonist as therapy for inflammatory disorders. Expert Opin. Biol. 2005. [Google Scholar] [CrossRef]
- Bø, S.H.; Davidsen, E.M.; Gulbrandsen, P.; Dietrichs, E.; Bovim, G.; Stovner, L.J.; White, L.R. Cerebrospinal fluid cytokine levels in migraine, tension-type headache and cervicogenic headache. Cephalalgia 2009. [Google Scholar] [CrossRef]
- Tilg, H.; Trehu, E.; Atkins, M.B.; Dinarello, C.A.; Mier, J.W. Interleukin-6 (IL-6) as an anti-inflammatory cytokine: Induction of circulating IL-1 receptor antagonist and soluble tumor necrosis factor receptor p55. Blood 1994, 83, 113–118. [Google Scholar]
- Tilg, H.; Mier, J.W.; Vogel, W.; Aulitzky, W.E.; Wiedermann, C.J.; Vannier, E.; Huber, C.; Dinarello, C.A. Induction of circulating IL-1 receptor antagonist by IFN treatment. J. Immunol. 1993, 150, 4678–4692. [Google Scholar]
- Arslan, H.H.; Tokgöz, E.; Yildizoʇlu, Ü.; Durmaz, A.; Bek, S.; Gerek, M. Evaluation of the changes in the nasal cavity during the migraine attack. J. Craniofac. Surg. 2014. [Google Scholar] [CrossRef]
- Aukerman, G.; Knutson, D.; Miser, W.F. Management of the acute migraine headache. Am. Fam. Physician 2002, 66, 2123–2130. [Google Scholar]
- Hsu, Y.C.; Lin, K.C.; Wang, S.J.; Wang, P.J.; Wang, Y.F.; Lee, L.H.; Wu, Z.A.; Shih, C.S.; Chen, Y.Y.; Chen, W.H.; et al. Medical treatment guidelines for acute migraine attacks. Acta Neurol. Taiwan. 2017, 26, 78–96. [Google Scholar]
- Raffaelli, B.; Mussetto, V.; Israel, H.; Neeb, L.; Reuter, U. Erenumab and galcanezumab in chronic migraine prevention: Effects after treatment termination. J. Headache Pain 2019, 20, 66. [Google Scholar] [CrossRef]
- Silberstein, S.D.; Stauffer, V.L.; Day, K.A.; Lipsius, S.; Wilson, M.-C. Galcanezumab in episodic migraine: Subgroup analyses of efficacy by high versus low frequency of migraine headaches in phase 3 studies (EVOLVE-1 & EVOLVE-2). J. Headache Pain 2019, 20, 75. [Google Scholar] [CrossRef]
- Dodick, D.W.; Lipton, R.B.; Silberstein, S.; Goadsby, P.J.; Biondi, D.; Hirman, J.; Cady, R.; Smith, J. Eptinezumab for prevention of chronic migraine: A randomized phase 2b clinical trial. Cephalalgia 2019. [Google Scholar] [CrossRef]
- Silberstein, S.D.; Cohen, J.M.; Yeung, P.P. Fremanezumab for the preventive treatment of migraine. Expert Opin. Biol. 2019, 1–9. [Google Scholar] [CrossRef]
- Neven, B.; Marvillet, I.; Terrada, C.; Ferster, A.; Boddaert, N.; Couloignier, V.; Pinto, G.; Pagnier, A.; Bodemer, C.; Tardieu, M.; et al. Long-term efficacy of the interleukin-1 receptor antagonist anakinra in ten patients with neonatal-onset multisystem inflammatory disease/chronic infantile neurologic, cutaneous, articular syndrome. Arthritis Rheum. 2010. [Google Scholar] [CrossRef]
- Landmann, E.C.; Walker, U.A. Pharmacological treatment options for cryopyrin-associated periodic syndromes. Expert Rev. Clin. Pharm. 2017, 10, 855–864. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Stable ID | Pearson r | 95% Confidence Interval | p-Value | R Squared |
|---|---|---|---|---|---|
| Cdhr5 | ENSMUSG00000025497 | 0.9996 | 0.9991 to 0.9998 | <0.0001 | 0.9992 |
| Gm8251 | ENSMUSG00000091844 | 0.9996 | 0.9991 to 0.9998 | <0.0001 | 0.9993 |
| Scarna3b | ENSMUSG00000088158 | 0.9993 | 0.9983 to 0.9997 | <0.0001 | 0.9986 |
| Nr0b1 | ENSMUSG00000025056 | 0.9991 | 0.9980 to 0.9996 | <0.0001 | 0.9983 |
| Lrrc15 | ENSMUSG00000052316 | 0.9987 | 0.9968 to 0.9994 | <0.0001 | 0.9973 |
| Ifi204 | ENSMUSG00000073489 | 0.9982 | 0.9959 to 0.9993 | <0.0001 | 0.9965 |
| Cd200r3 | ENSMUSG00000036172 | 0.9981 | 0.9955 to 0.9992 | <0.0001 | 0.9962 |
| Gm13389 | ENSMUSG00000087079 | 0.9977 | 0.9947 to 0.9990 | <0.0001 | 0.9955 |
| Hpx | ENSMUSG00000030895 | 0.9976 | 0.9944 to 0.9990 | <0.0001 | 0.9953 |
| Gm15941 | ENSMUSG00000086992 | 0.9974 | 0.9940 to 0.9989 | <0.0001 | 0.9949 |
| Zc3h12d | ENSMUSG00000039981 | 0.9973 | 0.9938 to 0.9989 | <0.0001 | 0.9947 |
| Gm49339 | ENSMUSG00000062593 | 0.9973 | 0.9936 to 0.9988 | <0.0001 | 0.9945 |
| Klre1 | ENSMUSG00000050241 | 0.9971 | 0.9933 to 0.9988 | <0.0001 | 0.9943 |
| Klk9 | ENSMUSG00000047884 | 0.9966 | 0.9921 to 0.9986 | <0.0001 | 0.9933 |
| Gm22486 | ENSMUSG00000080465 | 0.9966 | 0.9921 to 0.9986 | <0.0001 | 0.9933 |
| Abo | ENSMUSG00000015787 | 0.9965 | 0.9919 to 0.9985 | <0.0001 | 0.9931 |
| Cnga3 | ENSMUSG00000026114 | 0.9965 | 0.9917 to 0.9985 | <0.0001 | 0.9929 |
| Ccl4 | ENSMUSG00000018930 | 0.9960 | 0.9906 to 0.9983 | <0.0001 | 0.9920 |
| Snord66 | ENSMUSG00000077239 | 0.9958 | 0.9901 to 0.9982 | <0.0001 | 0.9916 |
| Gm13429 | ENSMUSG00000085141 | 0.9957 | 0.9898 to 0.9982 | <0.0001 | 0.9913 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lombardo, S.D.; Mazzon, E.; Basile, M.S.; Cavalli, E.; Bramanti, P.; Nania, R.; Fagone, P.; Nicoletti, F.; Petralia, M.C. Upregulation of IL-1 Receptor Antagonist in a Mouse Model of Migraine. Brain Sci. 2019, 9, 172. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9070172
Lombardo SD, Mazzon E, Basile MS, Cavalli E, Bramanti P, Nania R, Fagone P, Nicoletti F, Petralia MC. Upregulation of IL-1 Receptor Antagonist in a Mouse Model of Migraine. Brain Sciences. 2019; 9(7):172. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9070172
Chicago/Turabian StyleLombardo, Salvo Danilo, Emanuela Mazzon, Maria Sofia Basile, Eugenio Cavalli, Placido Bramanti, Riccardo Nania, Paolo Fagone, Ferdinando Nicoletti, and Maria Cristina Petralia. 2019. "Upregulation of IL-1 Receptor Antagonist in a Mouse Model of Migraine" Brain Sciences 9, no. 7: 172. https://0-doi-org.brum.beds.ac.uk/10.3390/brainsci9070172