The Regulatory Functions of Circular RNAs in Digestive System Cancers
by
 and
and
Xiao Yuan
1,2,†, 
Ya Yuan
1,2,†,
Zhi He
1,
Diyan Li
1,2,
Bo Zeng
1,2,
Qingyong Ni
1,2,
Mingyao Yang
1,2,* and
Deying Yang
1,2,* 1
College of Animal Science and Technology, Sichuan Agricultural University, Chengdu, Sichuan 611130, China
2
Farm Animal Genetic Resources Exploration and Innovation Key Laboratory of Sichuan Province, Sichuan Agricultural University, Chengdu, Sichuan 611130, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Cancers 2020, 12(3), 770; https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030770
Submission received: 18 January 2020
/
Revised: 19 March 2020
/
Accepted: 20 March 2020
/
Published: 24 March 2020
Abstract
:Circular ribonucleic acids (circRNAs), which are a type of covalently closed circular RNA, are receiving increasing attention. An increasing amount of evidence suggests that circRNAs are involved in the biogenesis and development of multiple diseases such as digestive system cancers. Dysregulated circRNAs have been found to act as oncogenes or tumour suppressors in digestive system cancers. Moreover, circRNAs are related to ageing and a wide variety of processes in tumour cells, such as cell apoptosis, invasion, migration, and proliferation. Moreover, circRNAs can perform a remarkable multitude of biological functions, such as regulating splicing or transcription, binding RNA-binding proteins to enable function, acting as microRNA (miRNA) sponges, and undergoing translated into proteins. However, in digestive system cancers, circRNAs function mainly as miRNA sponges. Herein, we summarise the latest research progress on biological functions of circRNAs in digestive system cancers. This review serves as a synopsis of potential therapeutic targets and biological markers for digestive system cancer.
1. Introduction
Digestive system (oral cavity, oesophagus, stomach, colorectum, liver, and pancreas)-related cancers seriously affect human health. To date, multiple digestive system cancers have been identified. Oral cancer is a general term for malignant tumours that occur in the mouth, ninety percent of which are oral squamous cell carcinomas (OSCCs) [1]. Globally, a large number of new cases of OSCC are diagnosed every year [2]. Oesophageal cancer, mainly oesophageal squamous cell carcinoma (ESCC), is not only the eighth most common cancer but also the sixth most common cause of death from cancer throughout the world. There are geographical differences in the morbidity and mortality of ESCC. For example, China has a high incidence of oesophageal cancer [3]. Furthermore, gastric cancer (GC) is also commonly diagnosed and is identified as a cause of death from cancer [4]. Colorectal cancer (CRC) is the third and the second most frequently diagnosed cancer in males and females, respectively. In most parts of the world, the CRC incidence ratio is higher in men than in women [5]. Hepatocellular carcinoma (HCC), which is the most common malignant liver tumour, is the second primary cause of cancer-related death globally [6]. In addition, pancreatic cancer (PC) is a highly lethal disease in older adults, for which the mortality closely parallels the incidence [7]. Consequently, a more profound comprehension of the molecular mechanisms of cancer progression in the digestive system is indispensable, which provides new ideas for the identification of new prognostic and diagnostic biomarkers for their prevention and treatment.
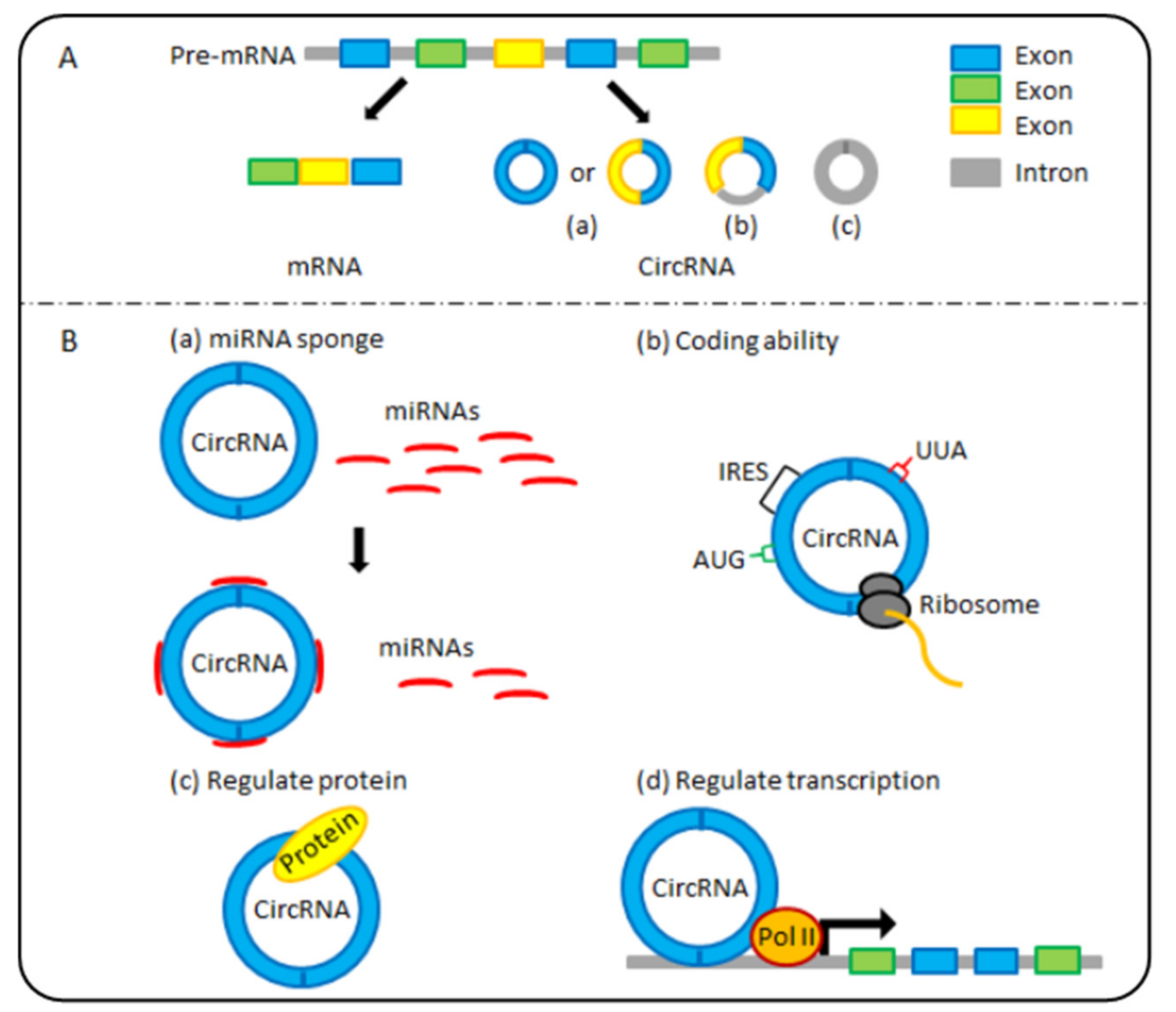
The term “circular ribonucleic acids (circRNA)” generally refers to a newly identified class of endogenous noncoding RNAs (ncRNAs). CircRNAs are covalent closed-loop structures without the 5′ caps or 3′ poly (A) tails that linear RNAs possess, which makes them more resistant to RNA exonucleases [8]. Furthermore, as shown in Figure 1A, circRNAs can originate from exons, introns (including intron lariats), or both exons and introns [9]. Importantly, different subclasses of circRNA molecules are produced by distinct biogenesis mechanisms [10,11]. The formation mechanism of exonic circRNAs (ecircRNAs) and exon-intron circRNAs (EIciRNAs) mainly includes three models: exon-skipping or lariat-driven circularisation, direct back-splicing or intron-pairing-driven circularisation, and RNA-binding-protein-driven circularisation [10]. The formation of intronic circRNAs (ciRNAs) depends on 2’,5’-phosphodiester formation from three different pathways [11]. In the 1970s, circRNAs were initially discovered as viroids in RNA viruses [12]. With the continuous improvement of high-throughput sequencing technology and bioinformatics, abundant and diverse circRNAs have been identified in various developmental stages and physiological conditions in numerous organisms, including humans [13], mice [13], and monkeys [14]. Many reports have revealed that circRNAs are indispensable in various physiological and pathological processes [15,16,17,18,19]. Recently, multiple possible biological functions of circRNAs have been discovered (Figure 1B). Typically, circRNAs function as efficient microRNA sponges or competing endogenous RNAs (ceRNAs) to affect gene expression [20]. In addition, circRNAs can regulate transcription [21] and protein functions [22]. Importantly, circRNAs are considered ncRNAs and have coding abilities [23]. In addition, circRNAs have been gradually confirmed to be related to ageing processes and various human diseases, including nervous system disorders, cardiovascular disorders, and cancers (e.g., gastrointestinal cancers) [24,25]. In this article, we highlight recent research on the latest progress in circRNA research on digestive system cancer and gut ageing. Finally, the future of circRNA research progress and directions for the diagnosis and targeted therapy of digestive system cancers are discussed.
2. CircRNAs Dysregulated in Digestive System Cancers
With the continuous improvement of high-throughput sequencing technology, numerous differentially expressed circRNAs have been identified in digestive system cancers (Table 1). For example, Sun et al. performed a comparative analysis of the global circRNA expression profiles between the malignantly transformed oesophageal carcinoma cell line SHEEC and the immortalised human oesophageal epithelial cell line SHEE [26]. They identified 813 significantly upregulated and 445 downregulated circRNAs [26]. Through differential gene expression analysis of ESCC and adjacent non-neoplastic tissues, 267 circRNAs—92 upregulated and 175 downregulated—were found to be dysregulated in ESCC tissues compared with non-neoplastic tissues [27]. Similarly, Shi et al. unearthed 744 dysregulated circRNAs in ESCC tissues [28]. The greater number of differentially expressed circRNAs found in the cell lines than in corresponding cancer tissue samples may be due to the differences between the simulated in vitro environment and the in vivo environment. Furthermore, different numbers of dysregulated circRNAs were found in the same kind of samples, possibly because of differences in the RNA quality, sequencing equipment, sequencing reagents, etc. The final results of high-throughput sequencing are affected by several factors, including the sample type, quantity, and quality; the sequencing equipment and reagents; and the subsequent data processing methodology. In conclusion, circRNAs might be implicated in the oncogenesis and evolution of digestive system cancers and could serve as new clinical diagnostic markers and therapeutic targets.
3. The Functions of CircRNAs in Digestive Tract Cancers
Recently, increasing evidence has demonstrated that many circRNAs potentially exhibit biological functions in digestive system cancers. Via influencing cell migration, proliferation, apoptosis, and invasion in digestive system cancer, circRNAs act as either oncogenes or tumour suppressors. Some mechanisms have been identified, such as serving as miRNA sponges and regulating protein functions. Among these, the principal mechanism of circRNAs unearthed in digestive system cancers is miRNA sponging. The mechanisms and functions of dysregulated circRNAs in digestive system cancers are displayed in Table 2.
3.1. Oral Cancer
CircDOCK1 (hsa_circ_100721) is associated with the apoptosis of OSCC cells, and the circDOCK1-miR-196a-5p-BIRC3 axis has great importance in OSCC [34]. Both circDOCK1 silencing and an increase in the miR-196a-5p mimic level led to increased apoptosis and decreased BIRC3 expression in OSCC cells [34]. In addition, in OSCC cell lines, hsa_circ_0008309 may exert an anti-cancer effect by manipulating the hsa_circ_0008309-miR-382-5p/miR-136-5p-ATXN1 pathway [35], and hsa_circ_0008309 overexpression inhibited miR-382-5p and miR-382-5p expression and increased ATXN1 expression. In summary, circDOCK1 and hsa_circ_0008309 act as tumour suppressors of OSCC and may be latent biomarkers for OSCC (Figure 2a).
3.2. Oesophageal Cancer
A previous study discovered that 3 circRNAs (hsa_circ_0067934, circ-TTC17, and circ-DLG1) act as oncogenes in ESCC (Figure 2b) [36,37,38]. In vitro, hsa_circ_0067934 silencing blocked cell cycle progression and inhibited the cell migration and proliferation of ESCC [36]. Similarly, circ-TTC17 knockdown significantly decreased the migration and proliferation of ESCC cells [37]. Furthermore, circ-DLG1 knockdown significantly reduced the proliferation of ESCC cells. Prediction and annotation analyses revealed that circ-DLG1 can sponge 20 miRNAs and affect 60 corresponding target mRNAs [38]. Importantly, 2 circRNAs (hsa_circ_0001946 and circ-SMAD7) were found to act as tumour suppressors in ESCC [37,39]. The overexpression of hsa_circ_0001946 restrained ESCC cell invasion, migration, and proliferation [39]. In addition, hsa_circ_0001946 was verified to predict disease-free survival (DFS), overall survival (OS) and recurrence in FFPE and frozen tissues. Furthermore, circ-SMAD7 knockdown promoted cell migration and proliferation ability, whereas circ-SMAD7 overexpression showed the opposite effect in ESCC [37]. Thus, these findings provide new ideas for screening for ESCC biomarkers.
3.3. Gastric Cancer
To date, 11 circRNAs—five upregulated (cancer-promoting) and six downregulated (cancer-suppressing)—have been found to be involved in GC (Figure 2c). The action mechanisms of six of these circRNAs have been described, revealing the pathological process of GC and providing new approaches for the prevention and therapy of GC.
3.3.1. CircRNAs that Act as Oncogenes in GC
By sponging members of the miR-125 family, the oncogenic circRNA circPVT1 may facilitate GC cell proliferation [15]. In addition, circRNA_001569 overexpression promoted cell viability and decreased GC apoptosis by suppressing miR-145 expression, which is mediated by NR4A2 [18]. Furthermore, inhibition of 3 circRNAs (circRNA0047905, circRNA0138960 and circRNA7690-15) restrained the cell invasion and proliferation of GC in vitro [19]. Future research should focus on the mechanism of action of these newly discovered circRNAs in GC.
3.3.2. CircRNAs that Act as Tumour Suppressors in GC
Six circRNAs—circYAP1, circLARP4, circRNA_100269, circPVRL3, circ-ZFR, and circ-104916—have been reported to act as tumour suppressors in GC (Table 2). Previous studies analysed the functional mechanisms of circYAP1, circLARP4, circRNA_100269, and circ-ZFR and found that they are ceRNAs that bind to miRNAs to inhibit GC. For example, circYAP1 restrained the invasion and proliferation of GC cells by binding miR-367-5p to inhibit p27Kip1 expression [40]. In addition, circLARP4 restrained the biological behaviours of GC cells through sponging miR-424 [41]. Furthermore, by targeting the LATS1 gene, miR-424 facilitated the cell invasion and proliferation of GC [41]. CircRNA_100269 was found to target miR-630, suppressing tumour cell growth [42]. In addition, miR-630 could obstruct the function of circRNA_100269 [42]. Circ-ZFR induced cell cycle arrest, suppressed GC cell proliferation, and promoted apoptosis through sponging miR-107/miR-130a and adjusting PTEN, and circ-ZFR regulated the expression level of p53 protein in vivo [43].
Moreover, these circRNAs act through other mechanisms. In particular, circPVRL3 may have protein-coding ability in view of the structure of its open reading frame, internal ribosomal entry sites, and m6A modification [44]. Knockdown of circPVRL3 promoted the cell migration and proliferation of GC [44]. Circ-104916 overexpression in vitro effectively suppressed the cell migration, proliferation and invasion abilities of GC [45]. In addition, circ-104916 downregulated N-cadherin, Vimentin, and Slug and upregulated E-cadherin, which suggests that circ-104916 may participate in epithelial–mesenchymal transition (EMT) [45]. Therefore, overexpression of circRNAs in GC could be an effective method to prevent the progression of GC.
3.4. Colorectal Cancer
Ten circRNAs—six upregulated in CRC (cancer-promoting) and four downregulated (cancer-suppressing) circRNAs—have been determined to be involved in the regulation of CRC (Table 2 and Figure 2d). These circRNAs are related to the cell migration, proliferation, and apoptosis of CRC and participate in the regulation of signalling pathways (e.g., the Ras signalling pathway and Wnt/β-catenin pathway). Knowledge about these circRNAs facilitates a deeper understanding of CRC pathogenesis.
3.4.1. CircRNAs that Act as Oncogenes in CRC
Six circRNAs have been found to be upregulated during the progression of CRC, and knockdown of circCCDC66 [46], circRNA-ACAP2 [47], and hsa_circ_0000069 [48] notably restrained CRC cell migration, invasion, and proliferation. In addition, circ-BANP [49] and hsa_circ_0007534 [50] participate in the regulation of CRC cell proliferation. Among these six circRNAs, circCCDC66 [46], circRNA-ACAP2 [47], and hsa_circ_0020397 [51] could be miRNA sponges that regulate the expression level of their targets.
The circRNA circCCDC66 was found to serve as a miRNA sponge to protect MYC mRNA from attack through miR-93 and miRNA-33b [46]. CircRNA-ACAP2 acted as a miRNA sponge to increase the expression of T lymphoma invasion and metastasis protein 1 (Tiam1) through abolishing the inhibitory function of miR-21-5p for Tiam1 expression, thereby promoting the invasion, migration, and proliferation of SW480 colon cancer cells [47]. Moreover, hsa_circ_0020397 inhibited apoptosis and promoted the cell invasion and viability of CRC through enhancing the expression of programmed death-ligand 1 (PD-L1) and telomerase reverse transcriptase (TERT), which are the target genes of miR-138, whereas miR-138 had the opposite effect [51]. Specifically, hsa_circ_0000069 knockdown caused G0/G1 period cell cycle detention in vitro [48]. These findings provide in-depth insights into CRC progression and a reference for the design of therapeutic targets.
3.4.2. CircRNAs that Act as Tumour Suppressors in CRC
Several circRNAs, including circITGA7, cir-ITCH, circ_0026344, and hsa_circ_0000567, have been found to be downregulated in CRC. CircITGA7 acted as a suppressor to repress the cell metastasis and proliferation of CRC by stimulating the transcription of its host gene ITGA7 and inhibiting the Ras signalling pathway [52]. Furthermore, through heightening the level of ITCH and restraining the Wnt/β-catenin pathway, cir-ITCH suppressed the proliferation of CRC cells [53]. In addition, circ_0026344 reduced the cell invasion and growth of CRC while accelerating apoptosis through sponging miR-31 and miR-21 in CRC [54]. Moreover, hsa_circ_0000567 knockdown promoted CRC cell migration and proliferation in vitro [55]. Therefore, overexpression of these circRNAs might regulate CRC.
4. CircRNAs Play Important Roles in Digestive Gland Cancers
Digestive gland cancers comprise HCC and PC. Previous studies have reported that circRNAs participate in the regulation of HCC cell migration, invasion, and proliferation and in pancreatic ductal adenocarcinoma (PDAC) cell invasion and proliferation [56,57,58] (Table 3). These circRNAs play a very important role in promoting or inhibiting the growth of digestive gland cancers.
4.1. Hepatocellular Carcinoma
A total of 18 circRNAs, 11 of which are upregulated in HCC (cancer-promoting) and the other 7 of which are downregulated (cancer-suppressing), have been found to be involved in HCC (Table 3 and Figure 2e).
4.1.1. CircRNAs that Act as Oncogenes in HCC
Eleven circRNAs have been found to participate in the regulation of HCC cell migration, proliferation, and invasion by two action mechanisms (as ceRNAs and by binding to proteins) (Table 3). CircFBLIM1 [59] and hsa_circ_0078710 [58] promoted HCC cell proliferation, invasion, and migration as ceRNA sponges. CircFBLIM1 was shown to regulate FBLIM1 expression by sponging miR-346 [59]. In addition, in HCC, hsa_circ_0078710 increased HDAC and CDK2 levels by sponging miR-31 and promoted HCC cell cycle progression, invasion, cell proliferation, and migration in vitro, as well as tumour formation by HCC cells in vivo [58]. Cullin 2 (Cul2) circular RNA (circ-10720) and circRNA_104075 have a functional mechanism similar to that of their upstream regulator. A recent study reported that through binding to the Cul2 promoter, Twist1 stimulates its transcription and selectively upregulates Cul2 circular RNA expression. These circRNAs can sponge miRNAs that target Vimentin, thus increasing the expression of Vimentin during EMT [60]. In addition, Cul2 circRNA stimulated the cell invasion, migration and proliferation of HCC [60], and circ_104075 acted as a ceRNA to encourage tumorigenesis by increasing the expression of YAP through sponging miR-582-3p [61]. Mechanistically, HNF4a binds to the -1409 to -1401 region of the circ_104075 promoter to increase the expression of circ_104075.
The remaining circRNAs act as oncogenes in HCC by binding to different miRNAs and influencing different signalling pathways (Table 3), which differentially affect the progression of HCC. Hsa_circ_101280 (hsa_circ_SLAIN1 and hsa_circ_0100929) greatly facilitated HCC tumorigenesis through upregulating JAK2 and adsorbing miR-375, which restrained apoptosis and enhanced proliferation of HCC cells [62]. Hsa_circ_0103809 promoted cell migration and proliferation and inhibited apoptosis in HCC through regulating the miR-490-5p/SOX2 signalling pathway [63]. These findings suggest that circRNAs play a pivotal role in HCC and could help us screen potential biomarkers and targets for the prevention and diagnosis of HCC.
4.1.2. CircRNAs that Act as Tumour Suppressors in HCC
Seven circRNAs have been found to serve as antitumour factors via multiple action mechanisms in HCC. CircARSP91 (hsa_circ_0085154) expression was influenced by the upstream regulator AR-ADAR1 [69]. The androgen receptor (AR) contributes to sex disparity in HCC. AR suppressed circARSP91 expression by upregulating ADAR1 p110 in HCC. In addition, circARSP91 repressed HCC tumour growth both in vivo and in vitro [69]. Similarly, DHX9 could induce downregulation of cSMARCA5 (hsa_circ_0001445) in HCC. cSMARCA5 overexpression facilitated apoptosis and repressed the invasion, migration, and proliferation of HCC cells, and this circRNA upregulated TIMP3 by adsorbing miR-181b-5p and miR-17-3p [70,71]. Furthermore, circMTO1 [72], circADAMTS14 [73], the circRNA SMAD2 [74], and circC3P1 (miR-4641/PCK1) [75] could serve as miRNA sponges to restrain HCC cell tumorigenic behaviours. Moreover, overexpression of both circZKSCAN1 and ZKSCAN1 mRNA repressed the invasion, migration, and proliferation of HCC cells; however, the mechanism of action of circZKSCAN1 was not related to its parental gene ZKSCAN1 (a zinc finger family gene). Overall, circARSP91 and miR-4641/PCK1 could be the most promising biomarkers because of their inhibitory effects on HCC cell migration, invasion, and proliferation and their clear mechanism of action.
4.2. Pancreatic Cancer
CircRNA_100782 and circ-PDE8A, which are upregulated in PC, are involved in the regulation of PDAC as miRNA sponges. Chen et al. found that circRNA_100782 knockdown inhibited the proliferation of PDAC cells by decreasing the expression of signal transducer and activator of transcription 3 (STAT3) and interleukin-6 receptor (IL6R), which are the target genes of microRNA-124 (miR-124) [57]. In addition, a study revealed that tumour-released exosomal circ-PDE8A serves as a miRNA sponge for miR-338 to adjust MACC1 and promotes the invasion of PDAC cells through the MACC/MET/ERK or AKT pathways [56]. These findings suggest that circRNA_100782 and circ-PDE8A act as oncogenes in PDAC and could be biomarkers for PDAC (Figure 2f).
5. Common CircRNAs Involved in Digestive System Cancers
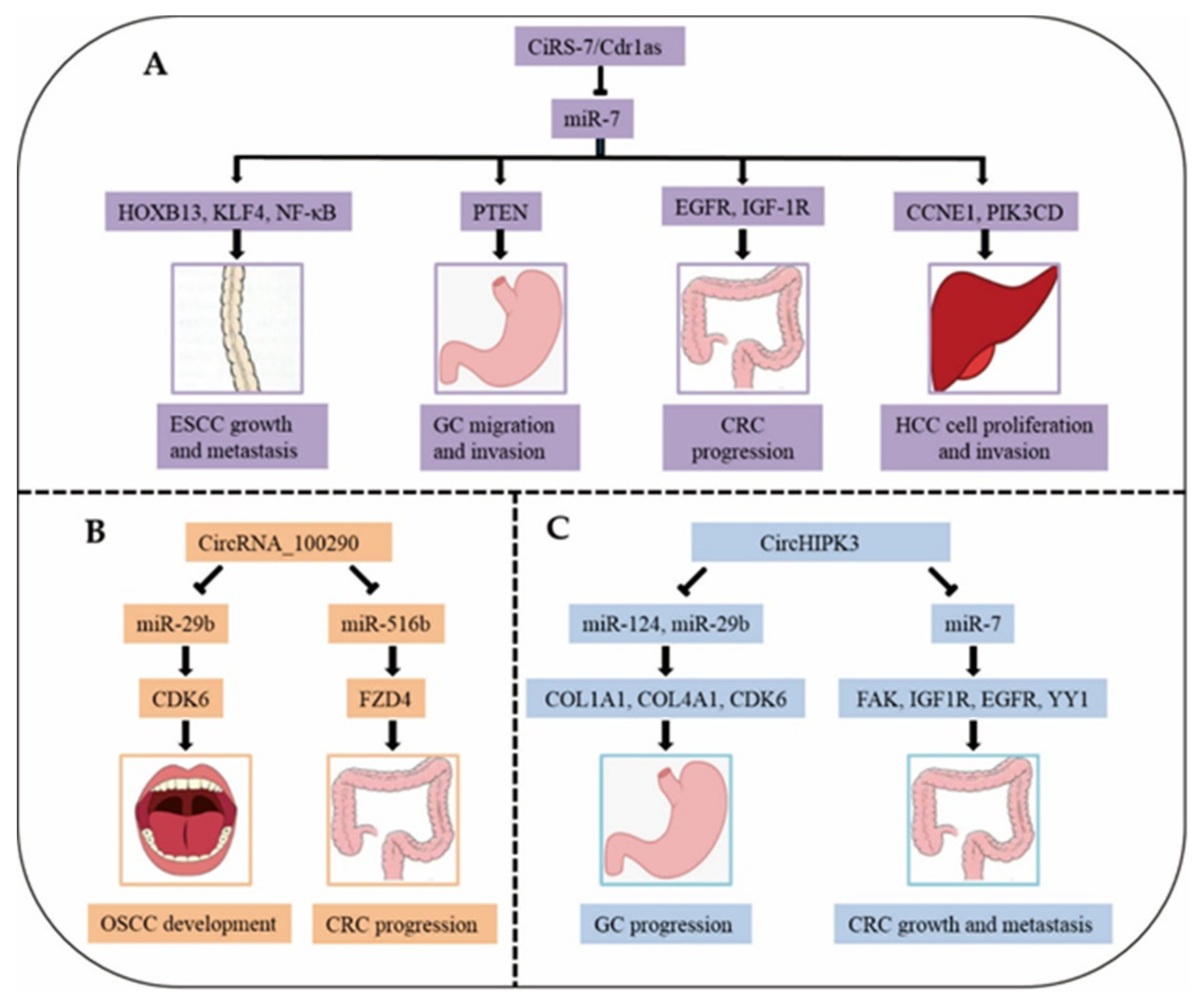
With increasing research on circRNAs in digestive system cancers, we found that six circRNAs—cerebellar degeneration-related protein 1 antisense RNA (ciRS-7 or Cdr1as), circRNA_100290, circHIPK3, circAGO2, hsa_circ_0014717, and hsa_circ_0001649—are dysregulated and perform similar functions in different cancers (Table 4). In digestive system cancers, the expression of ciRS-7, circRNA_100290, circHIPK3, and circAGO2 is upregulated, while that of hsa_circ_0014717 and hsa_circ_0001649 is downregulated. Among these six common circRNAs, ciRS-7 is involved in four digestive system cancers, namely, ESCC, GC, CRC, and HCC, through sponging miRNA-7 and different targets [77,78,79,80,81]. However, it is unclear how hsa_circ_0001649 affects the progression of CRC or HCC. Next, we explain the functions and mechanisms of these six circRNAs.
5.1. CiRS-7
CiRS-7, also called Cdr1as, serves as a super sponge for miR-7 and possesses over 60 common binding sites [20]. The overlapping co-expression of ciRS-7 and miR-7 was observed in the mouse brain, suggesting a high degree of endogenous interaction [91]. A study has found that ciRS-7 may affect the progress of sporadic Alzheimer’s disease (AD) through the miRNA-7/UBE2A signalling circuit [92]. In addition, ciRS-7 has been found to participate in the pathogenesis of multiple tumours (Figure 3A), which demonstrates that ciRS-7 may be a novel therapeutic target and prognostic marker for these deadly diseases.
Recent research revealed that ciRS-7 overexpression in ESCC encouraged cell invasion, migration and proliferation in vitro, as well as lung metastasis and tumour growth in vivo, by abolishing the tumour-suppressive effects of miR-7. Importantly, ciRS-7 served as a sponge for miR-7, resulting in the activation of the NF-κB/p65 pathway mediated by HOXB13 [77]. In addition, knockdown of the stem cell marker Kruppel-like factor-4 (KLF-4), which is a target gene of miR-7, attenuated cell invasion induced by ciRS-7 overexpression. Moreover, BAY 11–7082, an NF-κB inhibitor, also counteracted, to some extent, ciRS-7-mediated cell invasion [78]. Furthermore, ciRS-7 overexpression could block miR-7-induced tumour suppression, including reductions in cell migration and apoptosis, by counteracting the PTEN/PI3K/AKT pathway mediated by miR-7 in GC [79]. CiRS-7 overexpression in CRC cells dramatically reduced the tumour-suppressive activity (repression of cell invasion, migration and proliferation, and acceleration of apoptosis) of miR-7, and ciRS-7 regulated the EGFR/RAF1/MAPK pathway through suppressing the activity of miR-7 [80]. Intriguingly, ciRS-7 silencing inhibited CRC cell invasion and proliferation and decreased IGF-1R and EGFR expression, which could be rescued, in part, through a miR-7 inhibitor [81]. In HCC, ciRS-7 knockdown reduced the expression of PIK3CD and CCNE1, the direct targets of miR-7. In addition, ciRS-7 knockdown restrained HCC cell invasion and proliferation by adsorbing miR-7 [82]. Thus, ciRS-7 has a complex action mechanism in four digestive system cancers and could thus have great potential among the abovementioned six circRNAs as a therapeutic target for digestive system cancers.
5.2. CircRNA_100290
CircRNA_100290 (hsa_circRNA_100290) may serve as a ceRNA to adjust the expression of CDK6 by adsorbing miR-29b in OSCC [83] (Figure 3B). Knockdown of circRNA_100290 inhibited the proliferation of OSCC cells in vitro and in vivo [83]. Furthermore, silencing circRNA_100290 greatly decreased the rate of proliferation, restrained the invasion and migration abilities, and encouraged the apoptosis of CRC cells in vitro [84]. CircRNA_100290 is also a ceRNA for FZD4 that sponges miR-516b, resulting in Wnt/β-catenin pathway activation in CRC [84]. Therefore, circRNA_100290 may be an ideal biomarker for OSCC and CRC.
5.3. CircHIPK3
CircHIPK3 (hsa_circ_0000284) promoted cell proliferation through serving as a miRNA sponge for miR-124/miR-29b in GC. COL1A1, COL4A1 and CDK6 might play important functions mediated by the circHIPK3-miR-29b/miR-124 axes in GC [85] (Figure 3C). In addition, circHIPK3 knockdown markedly curbed cell invasion, migration, and proliferation, boosted the apoptosis of CRC in vitro, and inhibited CRC metastasis and growth in vivo. Furthermore, circHIPK3 overexpression significantly counteracted the miR-7-mediated repression of malignant phenotypes of CRC cells through upregulating the proto-oncogenes (FAK, IGF1R, EGFR, and YY1) targeted by miR-7. An upstream regulator of circHIPK3 expression in CRC is c-Myb, which is a transcription factor [86]. These findings suggest that circHIPK3 may be a therapeutic biomarker for GC and CRC. In addition, circHIPK3 regulates the proliferation, migration, invasion and apoptosis of non-small-cell lung cancer (NSCLC) cells through miR-149-mediated FOXM1 expression regulation, potentially providing novel insight into the pathogenesis of NSCLC [93].
5.4. CircAGO2
CircAGO2 (hsa_circ_0135889), an intronic circRNA produced from the AGO2 gene, was upregulated in both GC and CRC and promoted the invasion, metastasis and cell growth of cancer in vitro and in vivo [87]. In addition, circAGO2 facilitated its enrichment and activation on the 3’-untranslated region of target genes by interacting with human antigen R (HuR) protein, reducing argonaute 2 (AGO2) binding and repressing the silencing of AGO2/miRNA-mediated genes related to cancer progression [87]. Overall, circAGO2 might regulate cancer progression by facilitating the human antigen R (HuR)-inhibited effects of argonaute 2 (AGO2)-miRNA complexes. These results demonstrate that circAGO2 may be a potential therapeutic and diagnostic target for GC and CRC.
5.5. Hsa_circ_0014717
The overexpression of hsa_circ_0014717 induced G0/G1 period cell cycle arrest, restrained colony formation and CRC cell proliferation in vitro and inhibited xenograft tumour growth in vivo. Furthermore, hsa_circ_0014717 upregulated the expression of the cell cycle inhibitory protein p16 [94], and the knockdown of p16 reversed the inhibition of hsa_circ_0014717 on CRC cell growth [94]. Shao et al. discovered that hsa_circ_0014717 is stable in human gastric juice and that its characteristics make it suitable for use in clinical detection methodologies [88]. These results suggest that hsa_circ_0014717 may be a useful diagnostic marker for CRC.
5.6. Hsa_circ_0001649
Hsa_circ_0001649 overexpression inhibited cell invasion, migration, and proliferation and stimulated apoptosis in HCC [89]. In addition, the expression of hsa_circ_0001649 was heightened in serum samples from patients with CRC after surgery [90]. However, the concrete mechanism of action of hsa_circ_0001649 in HCC and CRC is not clear. Future studies should emphasise the function and mechanism of hsa_circ_0001649 and its potential as a biomarker in HCC and CRC.
6. The Clinical Significance of CircRNAs in Digestive System Cancers
6.1. Diagnostic Value Assessment
To date, circRNAs have been reported as potential diagnostic targets based on several efficient indicators in five digestive system cancers, such as receiver operating characteristic (ROC) curves, sensitivity and specificity (Table 5).
The diagnostic value of the combined circRNAs is higher than that of the single circRNAs in OSCC and PC. Three circRNAs have been identified as potential biomarkers for the diagnosis of OSCC, including hsa_circ_001242 [95], hsa_circ_0001874 and hsa_circ_0001971 [96]. Notably, the AUC (0.922) of the combination of hsa_circ_0001874 and hsa_circ_0001971 was larger than that of a single circRNA. A similar result was found in PC. Recent research revealed that circ-LDLRAD3 may be a new biomarker in the diagnosis of PC [97]. The expression of circ-LDLRAD3 in PC tissues was correlated with clinical stage, T classification, venous invasion, and lymphatic invasion, and in the plasma of patients with PC, it was significantly associated with CA19-9, N classification, venous invasion, clinical stage, metastasis, and lymphatic invasion [97]. Moreover, the AUC of circ-LDLRAD3 was 0.67. When combined with CA19-9, the AUC was increased to 0.87. These results suggest that the higher diagnostic value of the combination circRNAs could be an important choice for digestive system cancers.
The diagnostic values of circRNAs in different tissues differ in GC and PC. Six circRNAs—hsa_circ_002059 [98], hsa_circ_0000190 [99], hsa_circ_0000745 [100], hsa_circ_0000520 [26], hsa_circ_0000181 [101], and has_circ_0074362 [102]—may be potential novel biomarkers for the diagnosis of GC (Table 5). Of the six circRNAs, plasma hsa_circ_0000520 had the highest AUC (0.8967), sensitivity (0.8235), and specificity (0.8444) [26] and could therefore be a suitable diagnostic target for carcinoembryonic antigen (CEA) of GC patients’ plasma. Furthermore, hsa_circ_0000190 had a relatively higher diagnostic value for the combination of GC cancer tissue and patient plasma with AUC (0.775), sensitivity (0.712), and specificity (0.75) [103]. In PC patients, circRNA_104075 (hsa_circ_104075) in cancer tissues and patient serum had the highest AUC (0.973), sensitivity (0.96), and specificity (0.983) for diagnosis [61]. Thus, circRNAs can be selected as diagnostic targets for different digestive system cancer tissues.
6.2. Therapeutic Potential of CircRNAs
CircRNAs could have several advantages as potential targets for the treatment of digestive system cancers. First, circRNAs are stable molecules that may be relatively abundant in cells, tissues, and developmental stages. In addition, circRNAs are resistant to RNase R activity and exist in tissue [101], blood [104], saliva [96], and exosomes [105]. Moreover, circRNAs are highly conserved among species [106]. To date, multiple circRNAs involved in the regulation of digestive system cancer progression have been by in vivo and in vitro experiments [68,107]. A previous study suggested that circRNAs offer new opportunities to design innovative therapeutic strategies for cancer treatment [106]. Thus, circRNAs are very promising as treatment targets. For example, CiRS-7/Cdr1as is a miR sponge that is upregulated and involved in the regulation of ESCC [77], GC [78], CRC [79], and HCC [80] by different pathways, which suggests that the downregulation of ciRS-7/cdr1as expression might provide an effective treatment method. CircRNAs can act as sponges for miRNAs. Artificial circRNA sponges resistant to nuclease digestion could be synthesised using simple enzymatic ligation steps [108]. The synthetic circular miR-21 (scRNA21) sponge inhibited cancer cell proliferation and suppressed the activity of miR-21 on downstream protein targets, including the cancer protein DAXX [108]. Therefore, by studying the structure of endogenous circular RNA sponges to design and develop effective artificial sponges, we could eventually regulate the function of miRNAs in diseases.
7. Human Disease-Related CircRNA Databases
Following the application of high-throughput RNA sequencing and various modern computational approaches, a large number of human circRNA-related datasets have been established (Table 6). The circRNA basic information integration and analysis databases are circBase [111], CIRCpedia v2 [112], circView [113], and circBank [114]. It is worth noting that circBase and circBank provide the system nomenclature for circRNAs. However, circBase may fail to achieve accurate classification and positioning. CIRCpedia v2, circView, and circBank achieve more comprehensive and expert basic information integration regarding human circRNA. There are seven databases for the functional annotation of circRNAs (Table 6). The DeepBase v2.0 database focuses on the identification, expression, evolution, and function of circRNAs [115]. The CircInteractome and circNet databases emphasise circRNA-miRNA-gene and circRNA-protein regulatory networks [116]. The circFunBase database specifically provides the experimentally validated and computationally predicted functions of circRNA and circRNA-miRNA interaction networks [117]. The system nomenclature, function, and regulatory networks of human circRNAs (including digestive system cancer-related circRNAs) could be obtained from the above databases.
Moreover, the CSCD [118], circ2Traits [119], exoRBase [104], circRNA disease [120], circR2Disease [121], and somamiR 2.0 [105] databases integrate circRNA-human disease networks. Specifically, the circR2Disease (experimentally supported circular RNAs associated with various diseases) and somamiR 2.0 (miRNA-circRNA network) databases provide circRNA references in GC, CRC, HCC, and PC. The circRNA disease database presents experimentally supported circRNA-disease associations, including GC, CRC, and HCC [120]. Furthermore, a previous study demonstrated that exosomes can modulate the behaviour of recipient cells and may be used as circulating biomarkers for diseases [122]. ExoRBase aids researchers in identifying molecular signatures in blood exosomes and discovering new exosomal biomarkers and functional implications for human diseases, such as CRC, HCC, and PC [105].
In summary, these databases help researchers understand the essential information on circRNAs in digestive system cancers and other human diseases and promote understanding of the occurrence and development of disease as well as its diagnosis and prognosis.
8. Techniques and Methods of CircRNA Study in Digestive System Cancers
8.1. Identification of CircRNAs
Several techniques have been used to detect circRNAs in digestive system cancers from primary tumours and control tissues or cells [49,60], including microarray analysis [49] and high-throughput RNA sequencing [60]. Microarray analysis is suitable for screening known human circRNAs. High-throughput RNA sequencing is suitable for identifying new circRNAs and splice sites of circRNAs with sufficient underlying sequencing depth. Total RNA was treated with RNase R to degrade the linear RNA in library construction for screening circRNAs [43]. Then, circRNAs were experimentally validated by reverse transcription-quantitative polymerase chain reaction (RT-qPCR) with GAPDH as the reference gene [43]. Covalent loci of circRNA can be verified through Sanger sequencing. Furthermore, bioinformatics analysis is an accurate and convenient approach that can rapidly generate helpful information for further verification [127]. Thus, human disease-related circRNA databases could help us to analyse nomenclature, expression patterns, and evolution and predict the functions of human circRNAs.
8.2. Function and Mechanism of CircRNA
To provide the foundation for research on circRNA function, the subcellular location (nucleus or cytoplasm) of circRNAs first need to be analysed based on efficient nuclear/cytoplasmic RNA separation [44] and fluorescence in situ hybridisation (FISH) [48,52]. Then, gain-of-function (GOF) and loss-of-function (LOF) are employed to study circRNA functions in digestive system cancer cell lines and xenograft nude mouse models [46,67,72,75]. Human circRNA can be synthesised and cloned into the overexpression vector pcDNA3.1 [45]. The circRNA-pcDNA3.1 overexpression vector can then be transfected into digestive system cancer cell lines [40]. Next, those transfected cell lines are injected into a xenograft nude mouse model to test the GOF of circRNAs in an animal model [46,67,68,72,73,75]. On the other hand, small interfering RNAs (siRNAs) have been designed for the junction of circRNAs and transfected into digestive system cancer cell lines to examine the LOF of circRNAs [23]. Thus, multiple pathological processes can be researched with the aid of GOF and LOF of circRNAs in digestive system cancer, such as cell migration, proliferation, invasion, and anchorage-independent growth.
CircRNAs can act as a sponge binding to miRNA, thereby reducing the ability of miRNA to target mRNAs. Multiple databases can be used to analyse circRNA-miRNA-mRNA networks, such as circNet [123] and circFunBase [117] for known circRNAs and Targetscan [128] and RNAhybrid [129] for new circRNAs. A dual-luciferase assay (psiCHECK-2 vector) has been used to validate the relationship between circRNA and miRNA [42,51]. RNA immunoprecipitation (RIP) assays for AGO2 (argonaute 2) in digestive system cancer cell lines have been used to investigate the expression levels of endogenous circRNAs and miRNAs pulled down from AGO2-expressing cells by qPCR analysis [41]. Furthermore, Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Ontology (GO) analyses were performed to annotate genes targeted with the purpose of studying how circRNAs regulate parental gene expression [130]. On the other hand, circRNAs can bind to proteins and sequester them to particular subcellular locations [60,69]. Several techniques targeting this mechanism have been developed. To date, RNA pull-down assays and RIPs have been utilised to detect circRNA-protein interactions [131]. Following the discovery of new circRNA mechanisms, the development of new technology will help us to employ the function and regulatory mechanisms of circRNAs.
9. Conclusions
CircRNAs are a type of endogenous RNA that are stable and resist RNA nuclease digestion. CircRNAs have emerged as important regulators in multiple digestive system cancers. Dysregulated circRNAs were found to act as oncogenes or tumour suppressors in digestive system cancers (Figure 3). In the complex tumour microenvironment, overexpression of the downregulated circRNAs or depletion of the upregulated circRNAs in patients are the main approaches for circRNA-based therapy. According to current research data, the regulatory mechanism of circRNAs in digestive system cancers is very complex, and the underlying molecular mechanisms by which circRNAs cause and inhibit these cancers remain largely ambiguous. CircRNA transcriptome databases of OSCC, ESCC, GC, CRC, HCC, and PDAC have been reported and are convenient sources for selecting targets and biomarkers. The dysregulated circRNAs between digestive system cancer and normal cells/tissues may participate in regulating the inhibition and promotion of these digestive system cancers, suggesting that circRNAs could serve as therapeutic targets and biomarkers for clinical diagnosis. However, the above assumptions continue to present substantial difficulties and enormous challenges in clinical practice, and more studies on the significance, efficiency, generality, security, and reliability of these approaches are needed. Thus, we anticipate that this review will increase the comprehension of the principal functions of many circRNAs and their multiple regulatory hubs in the progression of digestive system cancers.
Author Contributions
Conceptualisation, D.Y. and M.Y.; writing—original draft preparation, X.Y. and Y.Y.; writing—review and editing: X.Y., Y.Y., Z.H., D.L., B.Z., Q.N., M.Y., D.Y.; funding acquisition, D.Y. and M.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the National Natural Science Foundation of China (31771338).
Acknowledgments
We thank the anonymous reviewers for their helpful and constructive comments that greatly contributed to improving the final version of this paper.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Yu, J.S.; Chen, Y.T.; Chiang, W.F.; Hsiao, Y.C.; Chu, L.J.; See, L.C.; Wu, C.S.; Tu, H.T.; Chen, H.W.; Chen, C.C.; et al. Saliva protein biomarkers to detect oral squamous cell carcinoma in a high-risk population in Taiwan. Proc. Natl. Acad. Sci. USA 2016, 113, 11549. [Google Scholar] [CrossRef] [Green Version]
- Irimie, A.I.; Braicu, C.; Sonea, L.; Zimta, A.A.; Cojocneanu-Petric, R.; Tonchev, K.; Mehterov, N.; Diudea, D.; Buduru, S.; Berindan-Neagoe, I. A Looking-Glass of Non-coding RNAs in oral cancer. Int. J. Mol. Sci. 2017, 18, 2620. [Google Scholar] [CrossRef] [Green Version]
- Lagergren, J.; Smyth, E.; Cunningham, D.; Lagergren, P. Oesophageal cancer. Lancet 2017, 390, 2383–2396. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Zheng, R.; Baade, P.D.; Zhang, S.; Zeng, H.; Bray, F.; Jemal, A.; Yu, X.Q.; He, J. Cancer statistics in China, 2015. CA Cancer J. Clin. 2016, 66, 115–132. [Google Scholar] [CrossRef] [Green Version]
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llovet, J.M.; Zucman-Rossi, J.; Pikarsky, E.; Sangro, B.; Schwartz, M.; Sherman, M.; Gores, G. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2016, 2, 16018. [Google Scholar] [CrossRef] [PubMed]
- Kamisawa, T.; Wood, L.D.; Itoi, T.; Takaori, K. Pancreatic cancer. Lancet 2016, 388, 73–85. [Google Scholar] [CrossRef]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floris, G.; Zhang, L.; Follesa, P.; Sun, T. Regulatory Role of Circular RNAs and Neurological Disorders. Mol. Neurobiol. 2017, 54, 5156–5165. [Google Scholar] [CrossRef]
- Meng, S.; Zhou, H.; Feng, Z.; Xu, Z.; Tang, Y.; Li, P.; Wu, M. CircRNA: Functions and properties of a novel potential biomarker for cancer. Mol Cancer 2017, 16, 94. [Google Scholar] [CrossRef]
- Petkovic, S.; Müller, S. RNA circularisation strategies in vivo and in vitro. Nucleic Acids Res. 2015, 43, 2454–2465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanger, H.L.; Klotz, G.; Riesner, D.; Gross, H.J.; Kleinschmidt, A.K. Viroids are single-stranded covalently closed circular RNA molecules existing as highly base-paired rod-like structures. Proc. Natl. Acad. Sci. USA 1976, 73, 3852–3856. [Google Scholar] [CrossRef] [Green Version]
- Werfel, S.; Nothjunge, S.; Schwarzmayr, T.; Strom, T.M.; Meitinger, T.; Engelhardt, S. Characterisation of circular RNAs in human, mouse and rat hearts. J. Mol. Cell. Cardiol. 2016, 98, 103–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.F.; Zhang, B.; Chen, X.J.; Wang, X.Y.; Zhang, B.W.; Ji, Y.Y.; Wu, K.C.; Wu, J.; Jin, Z.B. Circular RNAs in human and vertebrate neural retinas. RNA Biol. 2019, 16, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Li, Y.; Zheng, Q.; Bao, C.; He, J.; Chen, B.; Lyu, D.; Zheng, B.; Xu, Y.; Long, Z.; et al. Circular RNA profile identifies circPVT1 as a proliferative factor and prognostic marker in gastric cancer. Cancer Lett. 2017, 388, 208–219. [Google Scholar] [CrossRef] [PubMed]
- Drula, R.; Braicu, C.; Harangus, A.; Nabavi, S.M.; Trif, M.; Slaby, O.; Ionescu, C.; Irimie, A.; Berindan-Neagoe, I. Critical function of circular RNAs in lung cancer. Wiley Interdiscip. Rev. RNA 2020. [Google Scholar] [CrossRef]
- Cui, C.; Yang, J.; Li, X.; Liu, D.; Fu, L.; Wang, X. Functions and mechanisms of circular RNAs in cancer radiotherapy and chemotherapy resistance. Mol Cancer 2020, 19, 58. [Google Scholar] [CrossRef] [Green Version]
- Shen, F.; Liu, P.; Xu, Z.; Li, N.; Yi, Z.; Tie, X.; Zhang, Y.; Gao, L. CircRNA_001569 promotes cell proliferation through absorbing miR-145 in gastric cancer. J. Biochem. 2019, 165, 27–36. [Google Scholar] [CrossRef]
- Lai, Z.; Yang, Y.; Yan, Y.; Li, T.; Li, Y.; Wang, Z.; Shen, Z.; Ye, Y.; Jiang, K.; Wang, S. Analysis of co-expression networks for circular RNAs and mRNAs reveals that circular RNAs hsa_circ_0047905, hsa_circ_0138960 and has-circRNA7690-15 are candidate oncogenes in gastric cancer. Cell Cycle 2017, 16, 2301–2311. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256. [Google Scholar] [CrossRef] [PubMed]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Yuan, W.; Yang, X.; Li, P.; Wang, J.; Han, J.; Tao, J.; Li, P.; Yang, H.; Lv, Q.; et al. Circular RNA circ-ITCH inhibits bladder cancer progression by sponging miR-17/miR-224 and regulating p21, PTEN expression. Mol. Cancer 2018, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, B.; Chao, J.; Yao, H. Circular RNA and its mechanisms in disease: From the bench to the clinic. Pharmacol. Ther. 2018, 187, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Naeli, P.; Pourhanifeh, M.H.; Karimzadeh, M.R.; Shabaninejad, Z.; Movahedpour, A.; Tarrahimofrad, H.; Mirzaei, H.R.; Bafrani, H.H.; Savardashtaki, A.; Mirzaei, H.; et al. Circular RNAs and gastrointestinal cancers: Epigenetic regulators with a prognostic and therapeutic role. Crit. Rev. Oncol. Hematol. 2020, 145, 102854. [Google Scholar] [CrossRef]
- Sun, J.; Yuan, X.; Li, X.; Wang, D.; Shan, T.; Wang, W.; Wan, Q.; Wang, X.; Yan, J.; Gao, S. Comparative transcriptome analysis of the global circular RNAs expression profiles between SHEE and SHEEC cell lines. Am. J. Transl. Res. 2017, 9, 5169–5179. [Google Scholar]
- Chen, H.; Zheng, B.; Huang, L.; Zheng, W.; Chen, C. Microarray analysis reveals altered expression of multiple circular RNAs in the pathogenesis of esophageal squamous cell carcinoma. Transl. Cancer Res. Cancer Res. 2017, 6, 1248–1257. [Google Scholar] [CrossRef]
- Shi, P.; Sun, J.; He, B.; Song, H.; Li, Z.; Kong, W.; Wang, J.; Wang, J.; Xue, H. Profiles of differentially expressed circRNAs in esophageal and breast cancer. Cancer Manag. Res. 2018, 10, 2207–2221. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.F.; Li, B.W.; Sun, S.; Li, X.; Su, W.; Wang, Z.H.; Wang, F.; Zhang, W.; Yang, H.Y. Circular RNA Expression in Oral Squamous Cell Carcinoma. Front. Oncol. 2018, 8, 398. [Google Scholar] [CrossRef]
- Dang, Y.; Ouyang, X.; Zhang, F.; Wang, K.; Lin, Y.; Sun, B.; Wang, Y.; Wang, L.; Huang, Q. Circular RNAs expression profiles in human gastric cancer. Sci. Rep. 2017, 7, 9060. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, L.; Su, Y.; Zhang, X. Screening potential biomarkers for colorectal cancer based on circular RNA chips. Oncol. Rep. 2018, 39, 2499–2512. [Google Scholar] [CrossRef]
- Ren, S.; Xin, Z.; Xu, Y.; Xu, J.; Wang, G. Construction and analysis of circular RNA molecular regulatory networks in liver cancer. Cell Cycle 2017, 16, 2204–2211. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hao, X.; Wang, H.; Liu, Z.; He, Y.; Pu, M.; Zhang, H.; Yu, H.; Duan, J.; Qu, S. Circular RNA Expression Profile of Pancreatic Ductal Adenocarcinoma Revealed by Microarray. Cell. Physiol. Biochem. 2016, 40, 1334–1344. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wei, Y.; Yan, Y.; Wang, H.; Yang, J.; Zheng, Z.; Zha, J.; Bo, P.; Tang, Y.; Guo, X.; et al. CircDOCK1 suppresses cell apoptosis via inhibition of miR-196a-5p by targeting BIRC3 in OSCC. Oncol. Rep. 2018, 39, 951–966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, B.; Wang, F.; Li, X.; Sun, S.; Shen, Y.; Yang, H. Hsa_circ_0008309 May Be a Potential Biomarker for Oral Squamous Cell Carcinoma. Dis. Markers 2018, 2018, 7496890. [Google Scholar] [CrossRef] [Green Version]
- Xia, W.; Qiu, M.; Chen, R.; Wang, S.; Leng, X.; Wang, J.; Xu, Y.; Hu, J.; Dong, G.; Xu, P.L.; et al. Circular RNA has_circ_0067934 is upregulated in esophageal squamous cell carcinoma and promoted proliferation. Sci. Rep. 2016, 6, 35576. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, Q.; Sun, H.; Tang, W.; Yang, L.; Xu, Z.; Liu, Z.; Jin, H.; Cao, X. Circ-TTC17 Promotes Proliferation and Migration of Esophageal Squamous Cell Carcinoma. Dig. Dis. Sci. 2019, 64, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Rong, J.; Wang, Q.; Zhang, Y.; Zhu, D.; Sun, H.; Tang, W.; Wang, R.; Shi, W.; Cao, X.F. Circ-DLG1 promotes the proliferation of esophageal squamous cell carcinoma. Oncotargets Ther. 2018, 11, 6723–6730. [Google Scholar] [CrossRef] [Green Version]
- Fan, L.; Cao, Q.; Liu, J.; Zhang, J.; Li, B. Circular RNA profiling and its potential for esophageal squamous cell cancer diagnosis and prognosis. Mol. Cancer 2019, 18, 16. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Y.; Bian, Z.; Zhang, J.; Zhang, R.; Chen, X.; Huang, Y.; Wang, Y.; Zhu, J. Circular RNA YAP1 inhibits the proliferation and invasion of gastric cancer cells by regulating the miR-367-5p/p27 (Kip1) axis. Mol. Cancer 2018, 17, 151. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, H.; Hou, L.; Wang, G.; Zhang, R.; Huang, Y.; Chen, X.; Zhu, J. Circular RNA_LARP4 inhibits cell proliferation and invasion of gastric cancer by sponging miR-424-5p and regulating LATS1 expression. Mol. Cancer 2017, 16, 151. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, H.; Li, W.; Yu, J.; Li, J.; Shen, Z.; Ye, G.; Qi, X.; Li, G. CircRNA_100269 is downregulated in gastric cancer and suppresses tumor cell growth by targeting miR-630. Aging 2017, 9, 1585–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Liu, S.; Xu, Y.; Shu, R.; Wang, F.; Chen, C.; Zeng, Y.; Luo, H. Circular RNA-ZFR Inhibited Cell Proliferation and Promoted Apoptosis in Gastric Cancer by Sponging miR-130a/miR-107 and Modulating PTEN. Cancer Res. Treat. 2018, 50, 1396–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.D.; Xu, Z.P.; Sun, Z.Q.; Zhu, B.; Wang, Q.; Zhou, J.; Jin, H.; Zhao, A.; Tang, W.W.; Cao, X.F. Down-regulation of circPVRL3 promotes the proliferation and migration of gastric cancer cells. Sci. Rep. 2018, 8, 10111. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, F.; Lin, H.; Chen, Z.; Huang, Z.; Hu, J. The expression profile and clinical significance of circRNA0003906 in colorectal cancer. Oncotargets Ther. 2017, 10, 5187–5193. [Google Scholar] [CrossRef] [Green Version]
- Hsiao, K.Y.; Lin, Y.C.; Gupta, S.K.; Chang, N.; Yen, L.; Sun, H.S.; Tsai, S.J. Noncoding Effects of Circular RNA CCDC66 Promote Colon Cancer Growth and Metastasis. Cancer Res. 2017, 77, 2339–2350. [Google Scholar] [CrossRef] [Green Version]
- He, J.H.; Li, Y.G.; Han, Z.P.; Zhou, J.B.; Chen, W.M.; Lv, Y.B.; He, M.L.; Zuo, J.D.; Zheng, L. The CircRNA-ACAP2/Hsa-miR-21-5p/Tiam1 Regulatory Feedback Circuit Affects the Proliferation, Migration, and Invasion of Colon Cancer SW480 Cells. Cell. Physiol. Biochem. 2018, 49, 1539–1550. [Google Scholar] [CrossRef]
- Guo, J.N.; Li, J.; Zhu, C.L.; Feng, W.T.; Shao, J.X.; Wan, L.; Huang, M.D.; He, J.D. Comprehensive profile of differentially expressed circular RNAs reveals that hsa_circ_0000069 is upregulated and promotes cell proliferation, migration, and invasion in colorectal cancer. Onco Targets Ther. 2016, 9, 7451–7458. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Xu, Y.; Chen, Y.; Yan, F. Circular BANP, an upregulated circular RNA that modulates cell proliferation in colorectal cancer. Biomed. Pharmacother. 2017, 88, 138–144. [Google Scholar] [CrossRef]
- Zhang, R.; Xu, J.; Zhao, J.; Wang, X. Silencing of hsa-circ-0007534 suppresses proliferation and induces apoptosis in colorectal cancer cells. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 118–126. [Google Scholar]
- Zhang, X.L.; Xu, L.L.; Wang, F. Hsa_circ_0020397 regulates colorectal cancer cell viability, apoptosis and invasion by promoting the expression of the miR-138 targets TERT and PD-L1. Cell Biol. Int. 2017, 41, 1056–1064. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Zhang, C.; Lin, C.; Zhang, J.; Zhang, W.; Zhang, W.; Lu, Y.; Zheng, L.; Li, X. Circular RNA circITGA7 inhibits colorectal cancer growth and metastasis by modulating the Ras pathway and upregulating transcription of its host gene ITGA7. J. Pathol. 2018, 246, 166–179. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Zhu, H.; Shi, Y.; Wu, W.; Cai, H.; Chen, X. cir-ITCH plays an inhibitory role in colorectal cancer by regulating the Wnt/β-catenin pathway. PLoS ONE 2015, 10, e0131225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Y.; Liu, W.; Zhang, Y.; Zhang, Y.; Sun, S. CircRNA circ_0026344 as a prognostic biomarker suppresses colorectal cancer progression via microRNA-21 and microRNA-31. Biochem. Biophys. Res. Commun. 2018, 503, 870–875. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, X.; Lu, L.; He, L.; Hu, H.; Xu, Z. Circular RNA hsa_circ_0000567 can be used as a promising diagnostic biomarker for human colorectal cancer. J. Clin. Lab. Anal. 2018, 32, e22379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Yanfang, W.; Li, J.; Jiang, P.; Peng, T.; Chen, K.; Zhao, X.; Zhang, Y.; Zhen, P.; Zhu, J.; et al. Tumor-released exosomal circular RNA PDE8A promotes invasive growth via the miR-338/MACC1/MET pathway in pancreatic cancer. Cancer Lett. 2018, 432, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Shi, Y.; Zhang, Y.; Sun, J. CircRNA_100782 regulates pancreatic carcinoma proliferation through the IL6-STAT3 pathway. Oncotargets Ther. 2017, 10, 5783–5794. [Google Scholar] [CrossRef] [Green Version]
- Xie, B.; Zhao, Z.; Liu, Q.; Wang, X.; Ma, Z.; Li, H. CircRNA has_circ_0078710 acts as the sponge of microRNA-31 involved in hepatocellular carcinoma progression. Gene 2019, 683, 253–261. [Google Scholar] [CrossRef]
- Bai, N.; Peng, E.; Qiu, X.; Lyu, N.; Zhang, Z.; Tao, Y.; Li, X.; Wang, Z. circFBLIM1 act as a ceRNA to promote hepatocellular cancer progression by sponging miR-346. J. Exp. Clin. Cancer Res. 2018, 37, 172. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Chen, S.; Han, J.X.; Qian, B.; Wang, X.R.; Zhong, W.L.; Qin, Y.; Zhang, H.; Gao, W.F.; Lei, Y.Y.; et al. Twist1 Regulates Vimentin through Cul2 Circular RNA to Promote EMT in Hepatocellular Carcinoma. Cancer Res. 2018, 78, 4150–4162. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xu, Y.; Qian, Z.; Zheng, W.; Wu, Q.; Chen, Y.; Zhu, G.; Liu, Y.; Bian, Z.; Xu, W.; et al. circRNA_104075 stimulates YAP-dependent tumorigenesis through the regulation of HNF4a and may serve as a diagnostic marker in hepatocellular carcinoma. Cell Death Dis. 2018, 9, 1091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Wang, G.; Wang, J.; Li, C.; Zhang, L. Hsa_circ_101280 promotes hepatocellular carcinoma by regulating miR-375/JAK2. Immunol. Cell Biol. 2019, 97, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Cai, H.; Hu, B.; Ji, L.; Ruan, X.; Zheng, Z. Hsa_circ_0103809 promotes cell proliferation and inhibits apoptosis in hepatocellular carcinoma by targeting miR-490-5p/SOX2 signaling pathway. Am. J. Transl. Res. 2018, 10, 1690–1702. [Google Scholar] [PubMed]
- Gong, Y.; Mao, J.; Wu, D.; Wang, X.; Li, L.; Zhu, L.; Song, R. Circ-ZEB1.33 promotes the proliferation of human HCC by sponging miR-200a-3p and upregulating CDK6. Cancer Cell Int. 2018, 18, 116. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Huang, Z.L.; Xu, Y.H.; Zheng, Q.; Chen, Z.; Song, W.; Zhou, J.; Tang, Z.Y.; Huang, X.Y. Comprehensive circular RNA profiling reveals the regulatory role of the circRNA-100338/miR-141-3p pathway in hepatitis B-related hepatocellular carcinoma. Sci. Rep. 2017, 7, 5428. [Google Scholar] [CrossRef]
- Zhai, Z.; Fu, Q.; Liu, C.; Zhang, X.; Jia, P.; Xia, P.; Liu, P.; Liao, S.; Qin, T.; Zhang, H. Emerging Roles Of hsa-circ-0046600 Targeting The miR-640/HIF-1α Signalling Pathway In The Progression Of HCC. Onco Targets 2019, 12, 9291–9302. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.; Tan, J.; Gao, W.; Li, X.; Yang, Y.; Li, X.; Li, Y.; Wang, Q. Circular RNA hsa_circ_0016788 regulates hepatocellular carcinoma tumorigenesis through miR-486/CDK4 pathway. J. Cell. Physiol. 2018, 234, 500–508. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Wen, D.; Gong, L.; Wang, Y.; Liu, Z.; Yin, F. Circular RNA hsa_circ_0000673 promotes hepatocellular carcinoma malignance by decreasing miR-767-3p targeting SET. Biochem. Biophys. Res. Commun. 2018, 500, 211–216. [Google Scholar] [CrossRef]
- Shi, L.; Yan, P.; Liang, Y.; Sun, Y.; Shen, J.; Zhou, S.; Lin, H.; Liang, X.; Cai, X. Circular RNA expression is suppressed by androgen receptor (AR)-regulated adenosine deaminase that acts on RNA (ADAR1) in human hepatocellular carcinoma. Cell Death Dis. 2017, 8, e3171. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Xu, Q.G.; Wang, Z.G.; Yang, Y.; Zhang, L.; Ma, J.Z.; Sun, S.H.; Yang, F.; Zhou, W.P. Circular RNA cSMARCA5 inhibits growth and metastasis in hepatocellular carcinoma. J. Hepatol. 2018, 68, 1214–1227. [Google Scholar] [CrossRef]
- Zhang, X.; Zhou, H.; Jing, W.; Luo, P.; Qiu, S.; Liu, X.; Zhu, M.; Liang, C.; Yu, M.; Tu, J. The Circular RNA hsa_circ_0001445 Regulates the Proliferation and Migration of Hepatocellular Carcinoma and May Serve as a Diagnostic Biomarker. Dis. Markers 2018, 2018, 3073467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.; Li, J.; Wang, H.; Su, X.; Hou, J.; Gu, Y.; Qian, C.; Lin, Y.; Liu, X.; Huang, M.; et al. Circular RNA circMTO1 acts as the sponge of microRNA-9 to suppress hepatocellular carcinoma progression. Hepatology 2017, 66, 1151–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Li, D.; Liu, H.; Sun, H.; Liu, Z.; Zhang, L.; Hu, Y. The competing endogenous circular RNA ADAMTS14 suppressed hepatocellular carcinoma progression through regulating microRNA-572/regulator of calcineurin 1. J. Cell. Physiol. 2018, 234, 2460–2470. [Google Scholar] [CrossRef]
- Yang, Y.; Gao, X.; Zhang, M.; Yan, S.; Sun, C.; Xiao, F.; Huang, N.; Yang, X.; Zhao, K.; Zhou, H.; et al. Novel Role of FBXW7 Circular RNA in Repressing Glioma Tumorigenesis. JNCI 2018, 110, 304–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, L.; Wang, Y.; Cheng, Y.; Wang, W.; Lu, B.; Zhu, L.; Ma, Y. Circular RNA circC3P1 suppresses hepatocellular carcinoma growth and metastasis through miR-4641/PCK1 pathway. Biochem. Biophys. Res. Commun. 2018, 499, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Luo, J.; Hu, K.; Lin, J.; Huang, H.; Wang, Q.; Zhang, P.; Xiong, Z.; He, C.; Huang, Z.; et al. ZKSCAN1 gene and its related circular RNA (circZKSCAN1) both inhibit hepatocellular carcinoma cell growth, migration, and invasion but through different signaling pathways. Mol. Oncol. 2017, 11, 422–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.C.; Ke, S.; Meng, F.K.; Lu, J.; Zou, X.J.; He, Z.G.; Wang, W.F.; Fang, M.H. CiRS-7 promotes growth and metastasis of esophageal squamous cell carcinoma via regulation of miR-7/HOXB13. Cell Death Dis. 2018, 9, 838. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Wei, L.; Qin, T.; Yang, N.; Li, Z.; Xu, Z. Circular RNA ciRS-7 triggers the migration and invasion of esophageal squamous cell carcinoma via miR-7/KLF4 and NF-κB signals. Cancer Biol. Ther. 2019, 20, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Pan, H.; Li, T.; Jiang, Y.; Pan, C.; Ding, Y.; Huang, Z.; Yu, H.; Kong, D. Overexpression of Circular RNA ciRS-7 Abrogates the Tumor Suppressive Effect of miR-7 on Gastric Cancer via PTEN/PI3K/AKT Signaling Pathway. J. Cell. Biochem. 2017, 119, 440–446. [Google Scholar] [CrossRef]
- Weng, W.; Wei, Q.; Toden, S.; Yoshida, K.; Nagasaka, T.; Fujiwara, T.; Cai, S.; Qin, H.; Ma, Y.; Goel, A. Circular RNA ciRS-7-A Promising Prognostic Biomarker and a Potential Therapeutic Target in Colorectal Cancer. Clin. Cancer Res. 2017, 23, 3918–3928. [Google Scholar] [CrossRef] [Green Version]
- Tang, W.; Ji, M.; He, G.; Yang, L.; Niu, Z.; Jian, M.; Wei, Y.; Ren, L.; Xu, J. Silencing CDR1as inhibits colorectal cancer progression through regulating microRNA-7. Oncotargets Ther. 2017, 10, 2045–2056. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Gong, X.; Sun, L.; Zhou, Q.; Lu, B.; Zhu, L. The Circular RNA Cdr1as Act as an Oncogene in Hepatocellular Carcinoma through targeting miR-7 Expression. PLoS ONE 2016, 11, e0158347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Zhang, S.; Wu, J.; Cui, J.; Zhong, L.; Zeng, L.; Ge, S. circRNA_100290 plays a role in oral cancer by functioning as a sponge of the miR-29 family. Oncogene 2017, 36, 4551. [Google Scholar] [CrossRef] [PubMed]
- Fang, G.; Ye, B.L.; Hu, B.-R.; Ruan, X.J.; Shi, Y.X. CircRNA_100290 promotes colorectal cancer progression through miR-516b-induced downregulation of FZD4 expression and Wnt/β-catenin signaling. Biochem. Biophys. Res. Commun. 2018, 504, 184–189. [Google Scholar] [CrossRef]
- Cheng, J.; Zhuo, H.; Xu, M.; Wang, L.; Xu, H.; Peng, J.; Hou, J.; Lin, L.; Cai, J. Regulatory network of circRNA-miRNA-mRNA contributes to the histological classification and disease progression in gastric cancer. J. Transl. Med. 2018, 16, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, K.; Chen, X.; Xu, M.; Liu, X.; Hu, X.; Xu, T.; Sun, H.; Pan, Y.; He, B.; Wang, S. CircHIPK3 promotes colorectal cancer growth and metastasis by sponging miR-7. Cell Death Dis. 2018, 9, 417. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, F.; Fang, E.; Xiao, W.; Mei, H.; Li, H.; Li, D.; Song, H.; Wang, J.; Hong, M.; et al. Circular RNA circAGO2 drives cancer progression through facilitating HuR-repressed functions of AGO2-miRNA complexes. Cell Death Differ. 2019, 26, 1346–1364. [Google Scholar] [CrossRef]
- Shao, Y.; Li, J.; Lu, R.; Li, T.; Yang, Y.; Xiao, B.; Guo, J. Global circular RNA expression profile of human gastric cancer and its clinical significance. Cancer Med. 2017, 6, 1173–1180. [Google Scholar] [CrossRef]
- Zhang, X.; Qiu, S.; Luo, P.; Zhou, H.; Jing, W.; Liang, C.; Tu, J. Down-regulation of hsa_circ_0001649 in hepatocellular carcinoma predicts a poor prognosis. Cancer Biomark. 2018, 22, 135–142. [Google Scholar] [CrossRef]
- Ji, W.; Qiu, C.; Wang, M.; Mao, N.; Wu, S.; Dai, Y. Hsa_circ_0001649: A circular RNA and potential novel biomarker for colorectal cancer. Biochem. Biophys. Res. Commun. 2018, 497, 122–126. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Alexandrov, P.N.; Jaber, V.; Lukiw, W.J. Deficiency in the Ubiquitin Conjugating Enzyme UBE2A in Alzheimer’s Disease (AD) is Linked to Deficits in a Natural Circular miRNA-7 Sponge (circRNA; ciRS-7). Genes 2016, 7, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, H.; Han, X.; Ren, J.; Ren, K.; Li, Z.; Sun, Z. Circular RNA HIPK3 induces cell proliferation and inhibits apoptosis in non-small cell lung cancer through sponging miR-149. Cancer Biol. Ther. 2020, 21, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, J.; Cao, X.; Xu, L.; Chen, L. Hsa_circ_0014717 is downregulated in colorectal cancer and inhibits tumor growth by promoting p16 expression. Biomed. Pharmacother. 2018, 98, 775–782. [Google Scholar] [CrossRef]
- Sun, S.; Li, B.; Wang, Y.; Li, X.; Wang, P.; Wang, F.; Zhang, W.; Yang, H. Clinical Significance of the Decreased Expression of hsa_circ_001242 in Oral Squamous Cell Carcinoma. Dis. Markers 2018, 2018, 6514795. [Google Scholar] [CrossRef]
- Zhao, S.Y.; Wang, J.; Ouyang, S.B.; Huang, Z.K.; Liao, L. Salivary Circular RNAs Hsa_Circ_0001874 and Hsa_Circ_0001971 as Novel Biomarkers for the Diagnosis of Oral Squamous Cell Carcinoma. Cell. Physiol. Biochem. 2018, 47, 2511–2521. [Google Scholar] [CrossRef]
- Yang, F.; Liu, D.Y.; Guo, J.T.; Ge, N.; Zhu, P.; Liu, X.; Wang, S.; Wang, G.X.; Sun, S.Y. Circular RNA circ-LDLRAD3 as a biomarker in diagnosis of pancreatic cancer. World J. Gastroenterol. 2017, 23, 8345–8354. [Google Scholar] [CrossRef]
- Li, P.; Chen, S.; Chen, H.; Mo, X.; Li, T.; Shao, Y.; Xiao, B.; Guo, J. Using circular RNA as a novel type of biomarker in the screening of gastric cancer. Clin. Chim. Acta 2015, 444, 132–136. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, S.; Li, T.; Xiao, B.; Zhang, X. Clinical values of circular RNA 0000181 in the screening of gastric cancer. J. Clin. Lab. Anal. 2017, 32, e22333. [Google Scholar] [CrossRef] [Green Version]
- Huang, M.; He, Y.R.; Liang, L.C.; Huang, Q.; Zhu, Z.Q. Circular RNA hsa_circ_0000745 may serve as a diagnostic marker for gastric cancer. World J. Gastroenterol. 2017, 23, 6330–6338. [Google Scholar] [CrossRef]
- Sun, H.; Tang, W.; Rong, D.; Jin, H.; Fu, K.; Zhang, W.; Liu, Z.; Cao, H.; Cao, X. Hsa_circ_0000520, a potential new circular RNA biomarker, is involved in gastric carcinoma. Cancer Biomark. 2017, 21, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Shao, Y.; Sun, W.; Ye, G.; Zhang, X.; Xiao, B.; Guo, J. Downregulated expression of hsa_circ_0074362 in gastric cancer and its potential diagnostic values. Biomark. Med. 2017, 12, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, T.; Zhao, Q.; Xiao, B.; Guo, J. Using circular RNA hsa_circ_0000190 as a new biomarker in the diagnosis of gastric cancer. Clin. Chim. Acta 2017, 466, 167–171. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, Y.; Chen, B.; Zhao, J.; Yu, S.; Tang, Y.; Zheng, Q.; Li, Y.; Wang, P.; He, X.; et al. exoRBase: A database of circRNA, lncRNA and mRNA in human blood exosomes. Nucleic Acids Res. 2018, 46, D106–D112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, A.; Cui, Y. SomamiR 2.0: A database of cancer somatic mutations altering microRNA-ceRNA interactions. Nucleic Acids Res. 2016, 44, D1005–D1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verduci, L.; Strano, S.; Yarden, Y.; Blandino, G. The circRNA-microRNA code: Emerging implications for cancer diagnosis and treatment. Mol. Oncol. 2019, 13, 669–680. [Google Scholar] [PubMed] [Green Version]
- Zhu, P.; Ge, N.; Liu, D.; Yang, F.; Zhang, K.; Guo, J.; Liu, X.; Wang, S.; Wang, G.; Sun, S. Preliminary investigation of the function of hsa_circ_0006215 in pancreatic cancer. Oncol. Lett. 2018, 16, 603–611. [Google Scholar] [CrossRef]
- Liu, X.; Abraham, J.M.; Cheng, Y.; Wang, Z.; Wang, Z.; Zhang, G.; Ashktorab, H.; Smoot, D.T.; Cole, R.N.; Boronina, T.N.; et al. Synthetic Circular RNA Functions as a miR-21 Sponge to Suppress Gastric Carcinoma Cell Proliferation. Mol. Ther. Nucleic Acids 2018, 13, 312–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, Y.; Huang, L.; Zhang, J.; Pan, F.; Li, B.; Yan, Y.; Jia, B.; Liu, H.; Li, S.; et al. Decreased expression of hsa_circ_001988 in colorectal cancer and its clinical significances. Int. J. Clin. Exp. Pathol. 2015, 8, 16020–16025. [Google Scholar]
- Fu, L.; Wu, S.; Yao, T.; Chen, Q.; Xie, Y.; Ying, S.; Chen, Z.; Xiao, B.; Hu, Y. Decreased expression of hsa_circ_0003570 in hepatocellular carcinoma and its clinical significance. J. Clin. Lab. Anal. 2017, 32, e22239. [Google Scholar] [CrossRef]
- Glazar, P.; Papavasileiou, P.; Rajewsky, N. circBase: A database for circular RNAs. RNA 2014, 20, 1666–1670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Ma, X.K.; Li, G.W.; Yang, L. CIRCpedia v2: An Updated Database for Comprehensive Circular RNA Annotation and Expression Comparison. Genom. Proteom. Bioinform. 2018, 16, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xiang, Y.; Xia, S.; Liu, H.; Wang, J.; Ozguc, F.M.; Lei, L.; Kong, R.; Diao, L.; He, C.; et al. CircView: A visualisation and exploration tool for circular RNAs. Brief. Bioinform. 2018, 19, 1310–1316. [Google Scholar] [CrossRef]
- Liu, M.; Wang, Q.; Shen, J.; Yang, B.B.; Ding, X. Circbank: A comprehensive database for circRNA with standard nomenclature. RNA Biol. 2019, 16, 899–905. [Google Scholar] [CrossRef]
- Zheng, L.L.; Li, J.H.; Wu, J.; Sun, W.J.; Liu, S.; Wang, Z.L.; Zhou, H.; Yang, J.H.; Qu, L.H. deepBase v2.0: Identification, expression, evolution and function of small RNAs, LncRNAs and circular RNAs from deep-sequencing data. Nucleic Acids Res. 2016, 44, D196–D202. [Google Scholar] [CrossRef] [PubMed]
- Panda, A.C.; Dudekula, D.B.; Abdelmohsen, K.; Gorospe, M. Analysis of Circular RNAs Using the Web Tool CircInteractome. Methods Mol. Biol. 2018, 1724, 43–56. [Google Scholar] [PubMed] [Green Version]
- Meng, X.; Hu, D.; Zhang, P.; Chen, Q.; Chen, M. CircFunBase: A database for functional circular RNAs. Database 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Feng, J.; Chen, K.; Ma, Y.; Gong, J.; Cai, F.; Jin, Y.; Gao, Y.; Xia, L.; Chang, H.; et al. CSCD: A database for cancer-specific circular RNAs. Nucleic Acids Res. 2018, 46, D925–D929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosal, S.; Das, S.; Sen, R.; Basak, P.; Chakrabarti, J. Circ2Traits: A comprehensive database for circular RNA potentially associated with disease and traits. Front. Genet. 2013, 4, 283. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Wang, K.; Wu, F.; Wang, W.; Zhang, K.; Hu, H.; Liu, Y.; Jiang, T. circRNA disease: A manually curated database of experimentally supported circRNA-disease associations. Cell Death Dis. 2018, 9, 475. [Google Scholar] [CrossRef]
- Fan, C.; Lei, X.; Fang, Z.; Jiang, Q.; Wu, F.X. CircR2Disease: A manually curated database for experimentally supported circular RNAs associated with various diseases. Database 2018, 2018, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, M.; Gu, J.; Jiang, P.; Qian, H.; Xu, W.; Zhang, X. Exosomes in gastric cancer: Roles, mechanisms, and applications. Mol. Cancer 2019, 18, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.C.; Li, J.R.; Sun, C.H.; Andrews, E.; Chao, R.F.; Lin, F.M.; Weng, S.L.; Hsu, S.D.; Huang, C.C.; Cheng, C.; et al. CircNet: A database of circular RNAs derived from transcriptome sequencing data. Nucleic Acids Res. 2016, 44, D209–D215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Han, P.; Zhou, T.; Guo, X.; Song, X.; Li, Y. circRNADb: A comprehensive database for human circular RNAs with protein-coding annotations. Sci. Rep. 2016, 6, 34985. [Google Scholar] [CrossRef] [PubMed]
- Xia, S.; Feng, J.; Lei, L.; Hu, J.; Xia, L.; Wang, J.; Xiang, Y.; Liu, L.; Zhong, S.; Han, L.; et al. Comprehensive characterisation of tissue-specific circular RNAs in the human and mouse genomes. Brief. Bioinform. 2017, 18, 984–992. [Google Scholar] [PubMed] [Green Version]
- Wu, S.M.; Liu, H.; Huang, P.J.; Chang, I.Y.; Lee, C.C.; Yang, C.Y.; Tsai, W.S.; Tan, B.C. circlncRNAnet: An integrated web-based resource for mapping functional networks of long or circular forms of noncoding RNAs. Gigascience 2018, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yan, B.; Wang, Z.H.; Guo, J.T. The research strategies for probing the function of long noncoding RNAs. Genomics 2012, 99, 76–80. [Google Scholar] [CrossRef] [Green Version]
- Mon-Lopez, D.; Tejero-Gonzalez, C.M. Validity and reliability of the TargetScan ISSF Pistol & Rifle application for measuring shooting performance. Scand. J. Med. Sci. Sports 2019, 29, 1707–1712. [Google Scholar]
- Kruger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34, W451–W454. [Google Scholar] [CrossRef]
- Zhang, Z.; Song, N.; Wang, Y.; Zhong, J.; Gu, T.; Yang, L.; Shen, X.; Li, Y.; Yang, X.; Liu, X.; et al. Analysis of differentially expressed circular RNAs for the identification of a coexpression RNA network and signature in colorectal cancer. J. Cell. Biochem. 2019, 120, 6409–6419. [Google Scholar] [CrossRef]
- Du, W.W.; Zhang, C.; Yang, W.; Yong, T.; Awan, F.M.; Yang, B.B. Identifying and Characterizing circRNA-Protein Interaction. Theranostics 2017, 7, 4183–4191. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The classification and molecular functions of circular ribonucleic acids (circRNAs). (A) According to their sequences, circRNAs are divided into 3 main categories: (a) exonic circRNAs (ecircRNAs), (b) exon-intron circRNAs (EIciRNAs), and (c) intronic circRNAs (ciRNAs). (B) CircRNAs have four potential functions: (a) microRNA (miRNA) sponging: some circRNAs serve as efficient miRNA sponges, regulating the activity of miRNA target genes. (b) Coding ability: some circRNAs encode peptides or proteins and affect their biological function. (c) and (d) regulation: circRNAs affect protein function directly and regulate transcription.
Figure 1.
The classification and molecular functions of circular ribonucleic acids (circRNAs). (A) According to their sequences, circRNAs are divided into 3 main categories: (a) exonic circRNAs (ecircRNAs), (b) exon-intron circRNAs (EIciRNAs), and (c) intronic circRNAs (ciRNAs). (B) CircRNAs have four potential functions: (a) microRNA (miRNA) sponging: some circRNAs serve as efficient miRNA sponges, regulating the activity of miRNA target genes. (b) Coding ability: some circRNAs encode peptides or proteins and affect their biological function. (c) and (d) regulation: circRNAs affect protein function directly and regulate transcription.

Figure 2.
Dysregulated circRNAs act as oncogenes or tumour suppressors and have diagnostic value in digestive system cancers. (a) Oral cavity, (b) oesophagus, (c) stomach, (d) colorectum, (e) liver, and (f) pancreas. Green indicates circRNAs with potential diagnostic value in digestive system cancers.
Figure 2.
Dysregulated circRNAs act as oncogenes or tumour suppressors and have diagnostic value in digestive system cancers. (a) Oral cavity, (b) oesophagus, (c) stomach, (d) colorectum, (e) liver, and (f) pancreas. Green indicates circRNAs with potential diagnostic value in digestive system cancers.

Figure 3.
The mechanisms of three circRNAs (CiRS-7/Cdr1as, CircRNA_100290 and CircHIPK3) in different diseases. (A), Mechanism of ciRS-7 in ESCC, GC, CRC, and HCC. (B), Mechanism of circRNA_100290 in OSCC and CRC. (C), Mechanism of circHIPK3 in GC and CRC.
Figure 3.
The mechanisms of three circRNAs (CiRS-7/Cdr1as, CircRNA_100290 and CircHIPK3) in different diseases. (A), Mechanism of ciRS-7 in ESCC, GC, CRC, and HCC. (B), Mechanism of circRNA_100290 in OSCC and CRC. (C), Mechanism of circHIPK3 in GC and CRC.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Overview of differentially expressed circRNAs identified in digestive system cancers.
| Cancer Type | Sample | Number of Dysregulated CircRNAs (Up, Down) | References |
|---|---|---|---|
| OSCC | Cancer tissue and adjacent normal tissue | 16 (8, 8) | 2018 [29] |
| ESCC | SHEE and SHEEC cell lines | 1258 (813, 445) | 2017 [26] |
| ESCC | Cancer tissue and adjacent normal tissue | 267 (97, 175) | 2017 [27] |
| ESCC | Cancer tissue and adjacent normal tissue | 744 (469, 275) | 2018 [28] |
| GC | Cancer tissue and adjacent normal tissue | 713 (191, 522) | 2017 [30] |
| CRC | Cancer tissue and adjacent normal tissue | 10245 (6264, 3981) | 2018 [31] |
| HCC | Cancer tissue and adjacent normal tissue | 127 (113, 14) | 2017 [32] |
| PDAC | Cancer tissue and adjacent normal tissue | 351 (209, 142) | 2016 [33] |
Table 2.
CircRNAs in digestive tract cancers.
| Cancer Type | CircRNA | Dysregulation | MiRNA Sponge | Target Gene | Function | References |
|---|---|---|---|---|---|---|
| OSCC | CircDOCK1/hsa_circ_100721 | Down | miR-196a-5p | BIRC3 | Apoptosis (-) | [34] |
| Hsa_circ_0008309 | Down | miR-136-5p/miR-382-5p | ATXN1 | - | [35] | |
| ESCC | Hsa_circ_0067934 | Up | - | - | Proliferation (+), migration (+) | [36] |
| Circ-TTC17/hsa_circ_0021771 | Up | - | - | Proliferation (+), migration (+) | [37] | |
| Circ-DLG1/hsa_circ_0007203 | Up | - | - | Proliferation (+) | [38] | |
| Hsa_circ_0001946 | Down | - | - | Proliferation (−), invasion (−), migration (−) | [39] | |
| Circ-SMAD7/hsa_circ_0000848 | Down | - | - | Proliferation (−), migration (−) | [37] | |
| GC | CircPVT1 | Up | miR-125 | - | Proliferation (+) | [15] |
| CircRNA_001569/hsa_circ_001569 | Up | miR-145 | NR4A2 | Apoptosis (−) | [18] | |
| Hsa_circ_0047905 | Up | - | - | Proliferation (+), invasion (+) | [19] | |
| Hsa_circ_0138960 | Up | - | - | Proliferation (+), invasion (+) | [19] | |
| Hsa-circRNA7690-15 | Up | - | - | Proliferation (+), invasion (+) | [19] | |
| CircYAP1/has_circ_0002320 | Down | miR-367-5p | p27Kip1 | Proliferation (−), invasion (−) | [40] | |
| CircLARP4 | Down | miR-424 | LATS1 | Proliferation (−), invasion (−) | [41] | |
| CircRNA_100269/has_circ_100269 | Down | miR-630 | - | Proliferation (−) | [42] | |
| Circ-ZFR | Down | miR-130a/miR-107 | PTEN, p53 | Proliferation (−), apoptosis (+) | [43] | |
| CircPVRL3/has_circ_0066779 | Down | - | - | Proliferation (−), migration (−) | [44] | |
| Circ-104916/has_circ_104916 | Down | - | E-cadherin, N-cadherin, Vimentin and Slug | Proliferation (−), invasion (−), migration (−) | [45] | |
| CRC | CircCCDC66 | Up | miRNA-33b/miR-93 | MYC | Proliferation (+), invasion (+), migration (+) | [46] |
| CircRNA-ACAP2 | Up | miR-21-5p | Tiam1 | Proliferation (+), invasion (+), migration (+) | [47] | |
| Hsa_circ_0000069 | Up | - | - | Proliferation (+), invasion (+), migration (+) | [48] | |
| Circ-BANP | Up | - | - | Proliferation (+) | [49] | |
| Hsa_circ_0007534 | Up | - | - | Proliferation (+), apoptosis (−) | [50] | |
| Hsa_circ_0020397 | Up | miR-138 | TERT, PD-L1 | Invasion (+), apoptosis (−) | [51] | |
| CircITGA7/hsa_circ_0026782 | Down | - | Ras signalling pathway, ITGA7 | Proliferation (−), migration (−) | [52] | |
| Cir-ITCH | Down | - | Wnt/β-catenin pathway, ITCH | Proliferation (−) | [53] | |
| Circ_0026344/hsa_circ_0026344 | Down | miR-21/miR-31 | - | Proliferation (−), invasion (−) | [54] | |
| Hsa_circ_0000567 | Down | - | - | Proliferation (−), migration (−) | [55] |
Note: (+) means promotion and (−) means suppression.
Table 3.
CircRNAs in digestive gland cancers.
| Cancer Type | CircRNA | Dysregulation | Upstream Regulator | MiRNA Sponge | Target Gene | Function | References |
|---|---|---|---|---|---|---|---|
| HCC | Circ-10720/hsa_circ_10720 | Up | Twist1 | - | Vimentin | Proliferation (+), invasion (+), migration (+) | [60] |
| CircFBLIM1/hsa_circ_0010090 | Up | - | miR-346 | FBLIM1 | Proliferation (+), invasion (+), apoptosis (-) | [59] | |
| Circ-ZEB1.33 | Up | - | miR-200a-3p | CDK6 | Proliferation (+) | [64] | |
| CircRNA_104075/circ_104075 | Up | HNF4a | miR-582-3p | YAP | - | [61] | |
| CircRNA_100338/hsa_circRNA_100338 | Up | - | miR-141-3p | - | Migration (+) | [65] | |
| Hsa-circ-0046600 | Up | - | miR-640 | HIF-1α | Migration (+) | [66] | |
| Hsa_circ_0016788 | Up | - | miR-486 | CDK4 | Proliferation (+), invasion (+), apoptosis (−) | [67] | |
| Hsa_circ_101280/hsa_circ_SLAIN1/hsa_circ_0100929 | Up | - | miR-375 | JAK2 | Proliferation (+), apoptosis (−) | [62] | |
| Hsa_circ_0103809 | Up | - | miR-490-5p | SOX2 | Proliferation (+), migration (+), apoptosis (−) | [63] | |
| Hsa_circ_0000673 | Up | - | miR-767-3p | SET | Proliferation (+), invasion (+) | [68] | |
| Hsa_circ_0078710 | Up | - | miR-31 | HDAC, CDK2 | Proliferation (+), invasion (+), migration (+) | [58] | |
| CircARSP91/hsa_circ_0085154 | Down | AR-ADAR1 | - | - | - | [69] | |
| Hsa_circ_0001445/cSMARCA5 | Down | DHX9 | miR-17-3p/miR-181b-5p | TIMP3 | Proliferation (−), invasion (−), migration (−), apoptosis (+) | [70,71] | |
| CircMTO1/hsa_circRNA_0007874/hsa_circRNA_104135 | Down | - | miR-9 | p21 | Proliferation (−), invasion (−) | [72] | |
| CircADAMTS14/hsa_circ_0018665 | Down | - | miR-572 | RCAN1 | Proliferation (−), invasion (−), apoptosis (+) | [73] | |
| CircRNA SMAD2/hsa_circ_0000847 | Down | - | miR-629 | - | Invasion (−), migration (−) | [74] | |
| CircC3P1 | Down | - | miR-4641 | PCK1 | Proliferation (−), invasion (−), migration (−) | [75] | |
| CircZKSCAN1/hsa_circ_0001727 | Down | - | - | - | Proliferation (−), invasion (−), migration (−) | [76] | |
| PDAC | CircRNA_100782/hsa_circ_100782 | Up | - | miR-124 | IL6, STAT3 | Proliferation (+) | [57] |
| Circ-PDE8A/hsa_circ_0036627 | Up | - | miR-338 | MACC1, MET, ERK, AKT | Invasion (+) | [56] |
Note: (+) means promotion and (−) means suppression.
Table 4.
Common circRNAs in digestive system cancers.
| CircRNA | Cancer Type | Dysregulation | Upstream Regulator | MiRNA Sponge | Target Gene | Function | References |
|---|---|---|---|---|---|---|---|
| CiRS-7/Cdr1as | ESCC | Up | - | miR-7 | HOXB13, NF-κB/p65 | Proliferation (+), migration (+), invasion (+) | [77] |
| KLF4, NF-κB | Migration (+), invasion (+) | [78] | |||||
| GC | Up | - | miR-7 | PTEN/PI3K/AKT | Migration (+), apoptosis (-) | [79] | |
| CRC | Up | - | miR-7 | EGFR/RAF1/MAPK | Proliferation (+), migration (+), invasion (+), apoptosis (−) | [80] | |
| EGFR, IGF-1R | Proliferation (+), invasion (+) | [81] | |||||
| HCC | Up | - | miR-7 | CCNE1, PIK3CD | Proliferation (+), invasion (+) | [82] | |
| CircRNA_100290/hsa_circRNA_100290 | OSCC | Up | - | miR-29b | CDK6 | Proliferation (+) | [83] |
| CRC | Up | - | miR-516b | FZD4, Wnt/β-catenin | Proliferation (+), migration (+), invasion (+), apoptosis (−) | [84] | |
| CircHIPK3/hsa_circ_0000284 | GC | Up | - | miR-124/miR-29b | COL1A1, COL4A1, CDK6 | Proliferation (+) | [85] |
| CRC | Up | c-Myb | miR-7 | FAK, IGF1R, EGFR, YY1 | Proliferation (+), migration (+), invasion (+), apoptosis (−) | [86] | |
| CircAGO2/hsa_circ_0135889 | GC/CRC | Up | - | - | HuR | Proliferation (+), invasion (+), migration (+) | [87] |
| Hsa_circ_0014717 | GC | Down | - | - | - | - | [35] |
| CRC | Down | - | - | p16 | Proliferation (−) | [88] | |
| Hsa_circ_0001649 | CRC | Down | - | - | - | - | [89] |
| HCC | Down | - | - | - | Proliferation (−), migration (−), invasion (−), apoptosis (+) | [90] |
Note: (+) means promotion and (−) means suppression.
Table 5.
The diagnostic value of circRNAs in digestive system cancers.
| Cancer Type | CircRNA | Dysregulation | Sample | Clinicopathological Factors | AUC | Sensitivity | Specificity | References |
|---|---|---|---|---|---|---|---|---|
| OSCC | Hsa_circ_001242 | Down | Cancer tissue | Tumour size, T stage | 0.784 | 0.725 | 0.775 | [95] |
| Hsa_circ_0001874 | Up | Patients’ saliva | TNM stage, tumour grade | 0.863 | 0.7444 | 0.9024 | [96] | |
| Hsa_circ_0001971 | Up | Patients’ saliva | TNM stage | 0.845 | 0.7556 | 0.878 | [96] | |
| Hsa_circ_0001874 + hsa_circ_0001971 | - | - | - | 0.922 | 0.9268 | 0.7778 | [96] | |
| GC | Hsa_circ_002059 | Down | Cancer tissue | Distal metastasis, TNM stage, gender, age | 0.73 | 0.81 | 0.62 | [98] |
| Hsa_circ_0000190 | Down | Cancer tissue | Tumour diameter, lymphatic metastasis, distal metastasis, TNM stage, CA19-9 | 0.75 | 0.721 | 0.683 | [103] | |
| Patients’ plasma | CEA | 0.6 | 0.414 | 0.875 | ||||
| Cancer tissue + Patients’ plasma | - | 0.775 | 0.712 | 0.75 | ||||
| Hsa_circ_0000745 | Down | Cancer tissue | Tumour differentiation | - | - | - | [100] | |
| Patients’ plasma | TNM stage | 0.683 | 0.855 | 0.45 | ||||
| Patients’ plasma + CEA | - | 0.775 | 0.8 | 0.633 | ||||
| Hsa_circ_0000520 | Down | Cancer tissue | TNM stage | 0.6129 | 0.5357 | 0.8571 | [9] | |
| Patients’ plasma | CEA | 0.8967 | 0.8235 | 0.8444 | ||||
| Hsa_circ_0000181 | Down | Cancer tissue | Tumour diameter, lymphatic metastasis, distal metastasis, CA19-9 | 0.756 | 0.539 | 0.852 | [101] | |
| Patients’ plasma | Differentiation, CEA | 0.582 | 0.99 | 0.206 | ||||
| Hsa_circ_0074362 | Down | Cancer tissue | Lymphatic metastasis, CA19–9 | 0.63 | 0.362 | 0.843 | [102] | |
| CRC | Hsa_circ_001988 | Down | Cancer tissue | Differentiation, perineural invasion | 0.788 | 0.68 | 0.73 | [109] |
| Hsa_circ_0003906 | Down | Cancer tissue | Lymphatic metastasis, differentiation | 0.818 | 0.803 | 0.725 | [45] | |
| Hsa_circ_0000567 | Down | Cancer tissue | Tumour size, lymph metastasis, distal metastasis, TNM stage | 0.8653 | 0.8333 | 0.7647 | [55] | |
| Hsa_circ_0026782 | Down | Cancer tissue | Tumour size, lymph metastasis, distant metastasis, TNM stage | 0.8791 | 0.9275 | 0.6667 | [52] | |
| Hsa_circ_0000069 | Up | Cancer tissue | TNM stage, age | - | - | - | [48] | |
| HCC | Hsa_circ_0003570 | Down | Cancer tissue | Tumour diameter, differentiation, microvascular invasion, BCLC stages, TNM stages, serum AFP | 0.7 | 0.449 | 0.868 | [110] |
| Hsa_circ_0001445 | Down | Cancer tissue | The number of tumour foci | - | - | - | [71] | |
| Patients’ plasma | Serum AFP | 0.862 | 0.712 | 0.942 | ||||
| Hsa_circ_104075 | Up | Cancer tissue | - | - | - | - | [61] | |
| Patients’ serum | - | 0.973 | 0.96 | 0.983 | ||||
| Hsa_circ_0016788 | Up | Cancer tissue | - | 0.851 | - | - | [67] | |
| PC | Circ-LDLRAD3 | Up | Cancer tissue | Venous invasion, lymphatic invasion, clinical stage, T classification | 0.67 | 0.5738 | 0.7049 | [97] |
| Patients’ plasma | CA19-9, N classification, venous invasion, lymphatic invasion, clinical stage, metastasis | |||||||
| Circ-LDLRAD3 + CA19-9 | - | - | - | 0.87 | 0.8033 | 0.9355 | [97] |
Table 6.
Human disease circRNA-related databases.
| Name | Website | Description | References |
|---|---|---|---|
| CircRNA basic information integration and analysis databases | |||
| CircBase | http://www.circbase.org/ | CircRNA database for six species (human, mouse, worm, Latimeria chalumnae, Latimeria menadoensis) | [111] |
| CIRCpedia v2 | http://www.picb.ac.cn/rnomics/circpedia | Comprehensive circRNA annotation from over 180 RNA-seq datasets across six different species (human, mouse, rat, zebrafish, fly, worm) | [112] |
| CircView | http://gb.whu.edu.cn/CircView/ | A user-friendly visualisation tool for circRNAs detected by existing tools and regulatory elements, such as microRNA response elements and RNA-binding protein binding sites (human, mouse, zebrafish, fly, worm) | [113] |
| CircBank | http://www.circbank.cn/ | A comprehensive database for human circRNA with standard nomenclature, miRNA binding sites, conservation of circRNAs, m6A modification of circRNAs, mutation of circRNAs and protein-coding potential of circRNAs. | [114] |
| CircRNA-related function analysis database | |||
| DeepBase v2.0 | https://www.rna-seqblog.com/deepbase-v2-0-identification-expression-evolution-and-function-of-small-rnas-lncrnas-and-circular-rnas-from-deep-sequencing-data/ | Identification, expression, evolution and function of small RNAs, lncRNAs and circular RNAs from deep-sequencing data across 19 species | [115] |
| CircInteractome | https://omictools.com/circinteractome-tool | Exploring circular RNAs and their interacting proteins and microRNAs, identifying potential circRNAs which can act as RBP sponges, designing junction-spanning primers for specific detection of circRNAs of interest, siRNAs for circRNA silencing, and potential internal ribosomal entry sites (IRES). | [116] |
| CircNet | http://circnet.mbc.nctu.edu.tw/ | Providing the following resources: novel circRNAs, integrated miRNA-target networks, expression profiles of circRNA isoforms, genomic annotations of circRNA isoforms, sequences of circRNA isoforms, tissue-specific circRNA expression profiles, and circRNA-miRNA-gene regulatory networks | [123] |
| CircRNADb | http://reprod.njmu.edu.cn/circrnadb | Providing genomic information, exon splicing, genome sequences, internal ribosome entry sites (IRES), open reading frames (ORF) and references | [124] |
| TSCD | http://gb.whu.edu.cn/TSCD | Comprehensive characterisation of tissue-specific circular RNAs in the human and mouse genomes (including 26 different tissues) | [125] |
| CirclncRNAnet | http://app.cgu.edu.tw/circlnc/ | An integrated web-based resource for mapping functional networks of long or circular forms of noncoding RNAs | [126] |
| CircFunBase | http://bis.zju.edu.cn/CircFunBase/ | A database for functional circular RNAs (experimentally validated and computationally predicted functions and circRNA-miRNA interaction networks) | [117] |
| CircRNA databases related to human diseases | |||
| CSCD | http://gb.whu.edu.cn/CSCD | Comprehensive cancer-specific circRNA database | [118] |
| Circ2Traits | http://gyanxet-beta.com/circdb/ | Comprehensive knowledgebase of potential associations of circular RNAs with human diseases (including circRNAs with disease-miRNA and SNPs-circRNA loci) | [119] |
| ExoRBase | http://www.exoRBase.org | Database of circRNA, lncRNA and mRNA in human blood exosomes | [104] |
| CircRNA disease | http://cgga.org.cn:9091/circRNADisease/ | A manually curated database of experimentally supported circRNA-disease associations (48 kinds of disease) | [120] |
| CircR2Disease | http://bioinfo.snnu.edu.cn/CircR2Disease/ | A manually curated database for experimentally supported circular RNAs associated with various diseases (725 cases between 661 circRNAs and 100 human diseases) | [121] |
| SomamiR 2.0 | http://compbio.uthsc.edu/SomamiR | A database of cancer somatic mutations altering microRNA-ceRNA interactions (miRNA-circRNA) | [105] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yuan, X.; Yuan, Y.; He, Z.; Li, D.; Zeng, B.; Ni, Q.; Yang, M.; Yang, D. The Regulatory Functions of Circular RNAs in Digestive System Cancers. Cancers 2020, 12, 770. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030770
AMA Style
Yuan X, Yuan Y, He Z, Li D, Zeng B, Ni Q, Yang M, Yang D. The Regulatory Functions of Circular RNAs in Digestive System Cancers. Cancers. 2020; 12(3):770. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030770
Chicago/Turabian StyleYuan, Xiao, Ya Yuan, Zhi He, Diyan Li, Bo Zeng, Qingyong Ni, Mingyao Yang, and Deying Yang. 2020. "The Regulatory Functions of Circular RNAs in Digestive System Cancers" Cancers 12, no. 3: 770. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12030770
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.