Deregulated microRNAs Are Associated with Patient Survival and Predicted to Target Genes That Modulate Lung Cancer Signaling Pathways


 , , , , ,
, , , , ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
2.1. Patient Clinical and Histopathological Characteristics
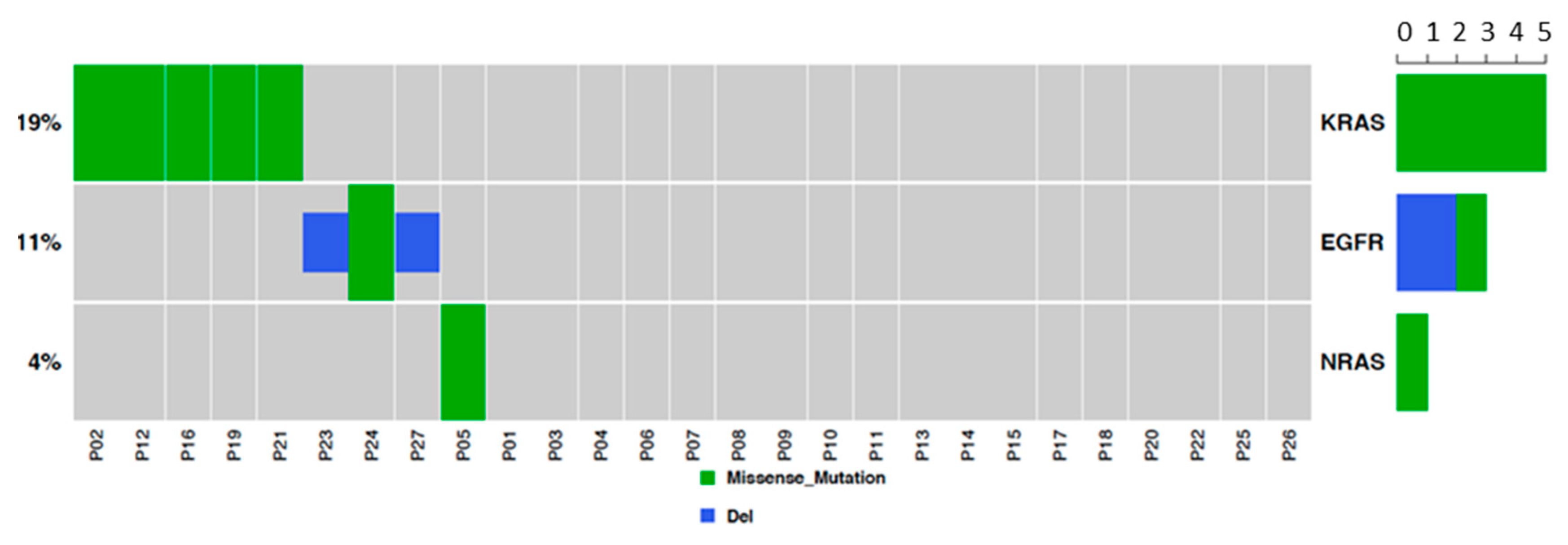
2.2. EGFR and RAS Driver Mutations in Lung AD and SCC
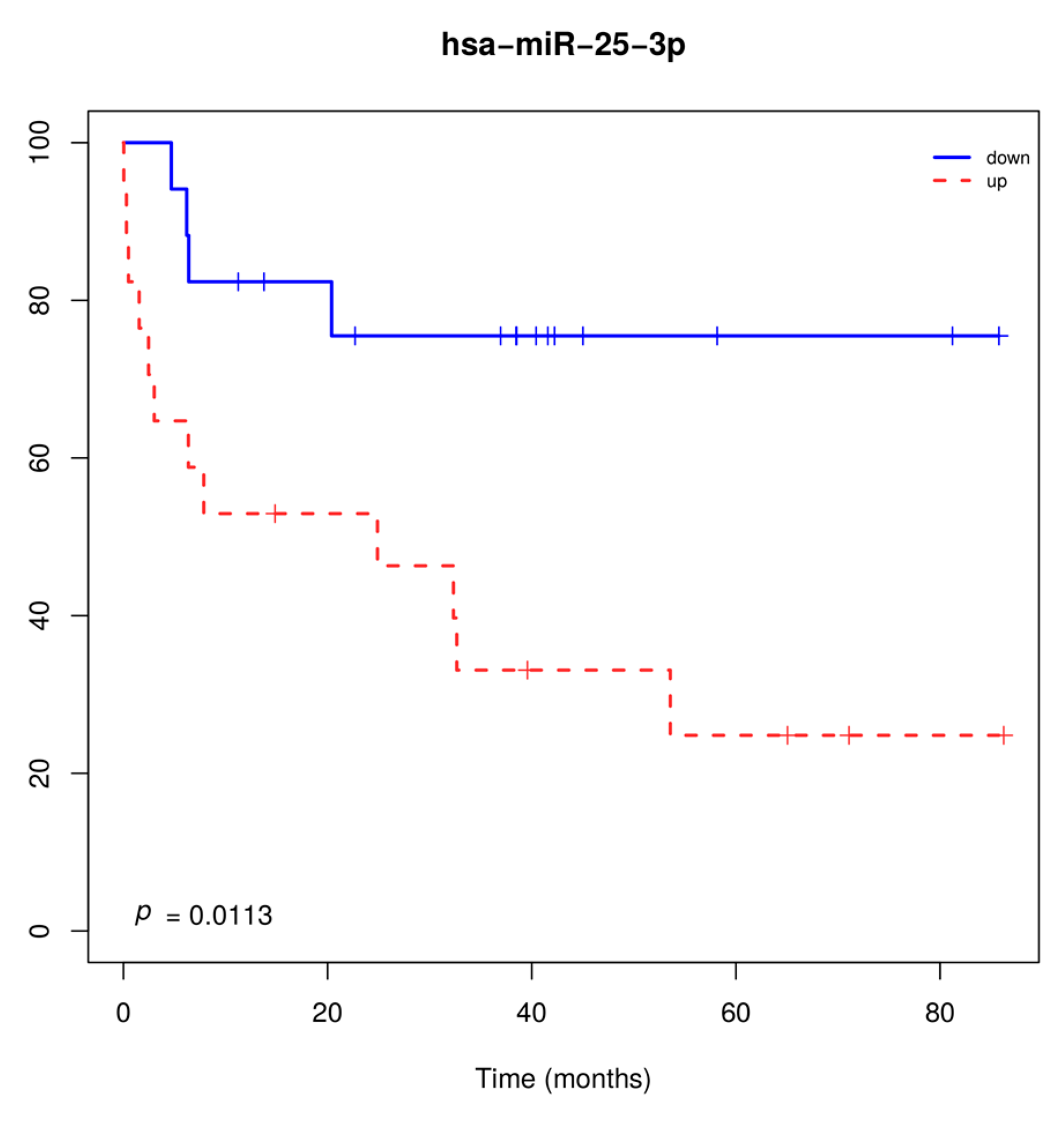
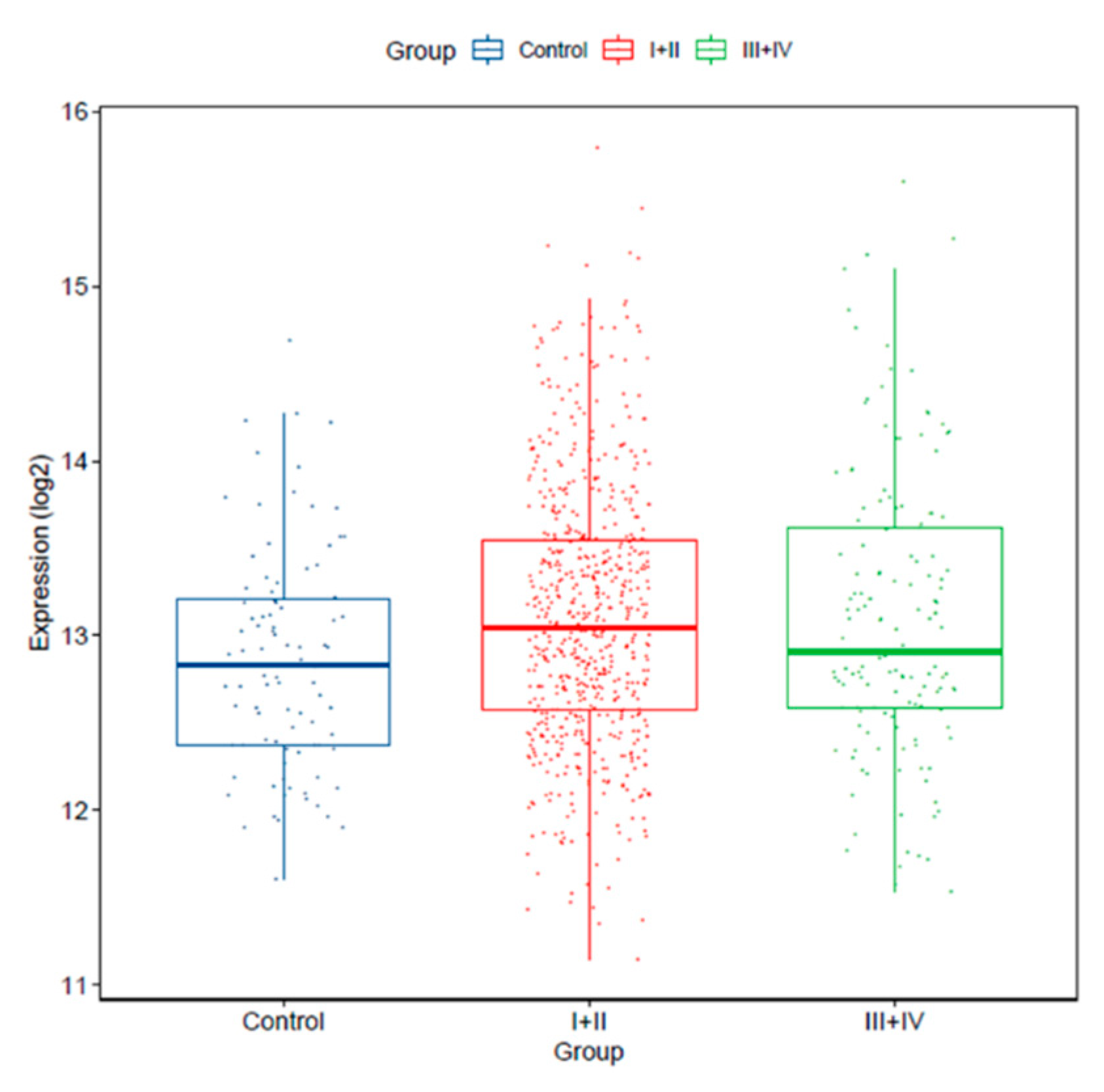
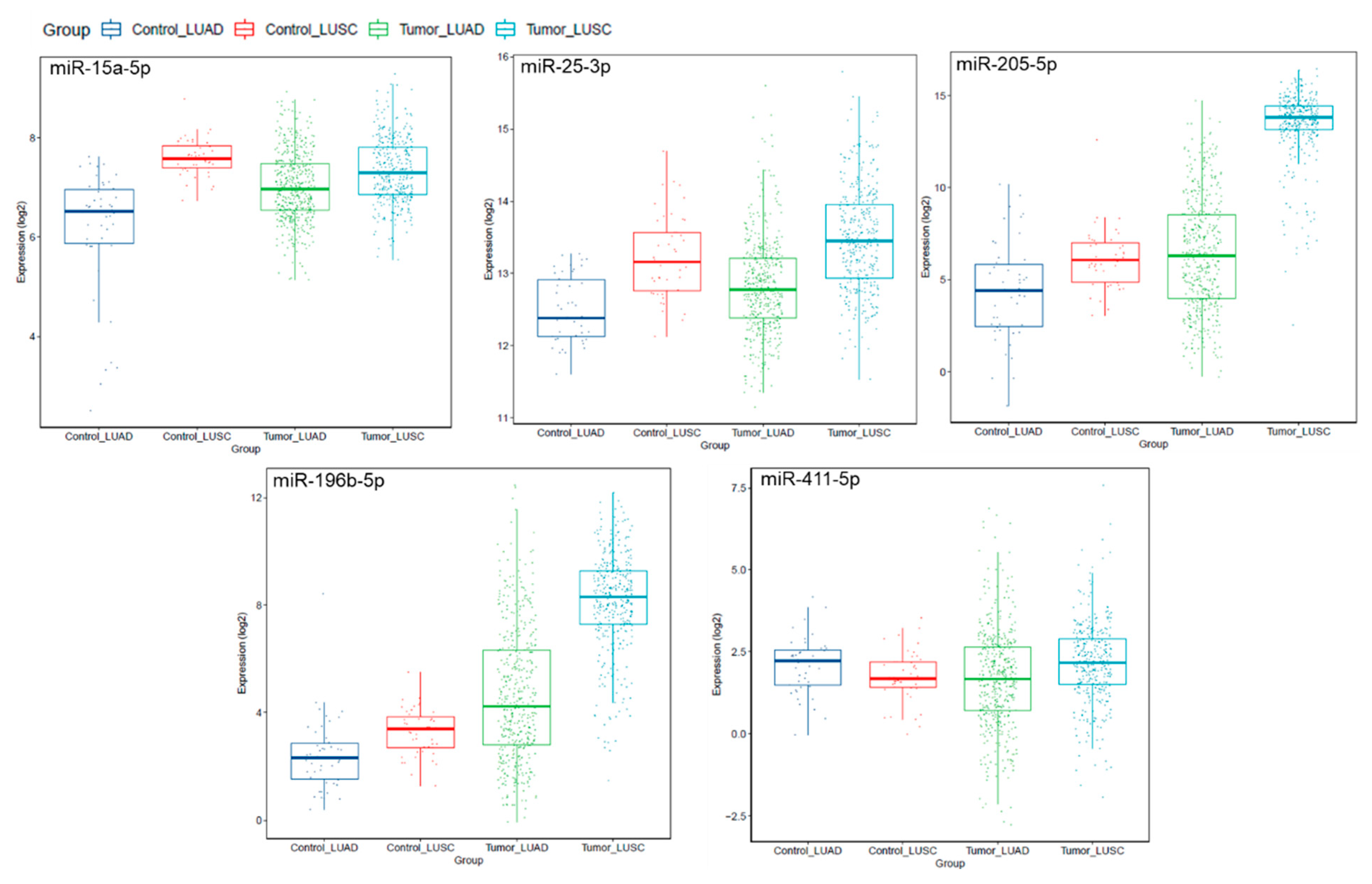
2.3. miRNAs Are Deregulated in Lung AD and SCC Compared to Normal Lung Tissues, and miR-25-3p Overexpression Is Significantly Associated with Poor Survival
2.4. miRNAs Are Predicted to Regulate Genes Abnormally Expressed in Lung AD and SCC, which Modulate Known Pathways of Lung Cancer
3. Discussion
4. Material and Methods
4.1. Ethics Statement
4.2. Patient Samples
4.3. RNA and DNA Extraction
4.4. SNaPShot Assay
4.5. EGFR Exon 19 Deletion Analysis
4.6. EGFR Exon 20 Insertion Analysis
4.7. Quantitative miRNA Expression Analysis by TaqMan Low Density Arrays
4.8. Computational and Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Estimativa 2020: Incidência de Câncer no Brasil INCA - Instituto Nacional de Câncer. Available online: https://www.inca.gov.br/publicacoes/livros/estimativa-2020-incidencia-de-cancer-no-brasil (accessed on 31 July 2020).
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Travis, W.D.; Brambilla, E.; Nicholson, A.G.; Yatabe, Y.; Austin, J.H.M.; Beasley, M.B.; Chirieac, L.R.; Dacic, S.; Duhig, E.; Flieder, U.B.; et al. The 2015 World Health Organization Classification of Lung Tumors. J. Thorac. Oncol. 2015, 10, 1243–1260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerman, P.S.; Voet, D.; Lawrence, M.S.; Voet, D.; Jing, R.; Cibulskis, K.; Sivachenko, A.; Stojanov, P.; McKenna, A.; Lander, E.S.; et al. Comprehensive genomic characterization of squamous cell lung cancers. Nature 2012, 489, 519–525. [Google Scholar]
- The Cancer Genome Atlas Research Network; Cancer Genome Atlas Research Network; Collisson, E.A.; Berger, A.H.; Beer, D.G.; Hammerman, P.S.; Herman, J.G.; Jurisica, I.; Kwiatkowski, D.; Robertson, G.; et al. Comprehensive molecular profiling of lung adenocarcinoma. Nature 2014, 511, 543–550. [Google Scholar] [CrossRef]
- Chang, J.T.-H.; Lee, Y.-M.; Huang, R.S. The impact of the Cancer Genome Atlas on lung cancer. Transl. Res. 2015, 166, 568–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kris, M.G.; Johnson, B.E.; Berry, L.D.; Kwiatkowski, D.J.; Iafrate, A.J.; Wistuba, I.I.; Varella-Garcia, M.; Franklin, W.A.; Aronson, S.L.; Su, P.-F.; et al. Using Multiplexed Assays of Oncogenic Drivers in Lung Cancers to Select Targeted Drugs. JAMA 2014, 311, 1998. [Google Scholar] [CrossRef]
- Leal, L.F.; De Paula, F.E.; De Marchi, P.; Viana, L.D.S.; Pinto, G.D.J.; Carlos, C.D.; Berardinelli, G.N.; Miziara, J.E.; Da Silva, C.M.; Silva, E.C.A.; et al. Mutational profile of Brazilian lung adenocarcinoma unveils association of EGFR mutations with high Asian ancestry and independent prognostic role of KRAS mutations. Sci. Rep. 2019, 9, 3209. [Google Scholar] [CrossRef]
- Andreis, T.F.; Correa, B.S.; Vianna, F.S.; De-Paris, F.; Siebert, M.; Leistner-Segal, S.; Hahn, E.C.; Ulbrich, J.M.; Rivero, L.F.; De Oliveira, F.H.; et al. Analysis of Predictive Biomarkers in Patients With Lung Adenocarcinoma From Southern Brazil Reveals a Distinct Profile From Other Regions of the Country. J. Glob. Oncol. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Palacio, S.; Pontes, L.; Prado, E.; Arshad, J.; Ali, R.; Piha, T.; Bacchi, C.E.; Mudad, R.; Lopes, G. EGFR Mutation Testing: Changing Patterns of Molecular Testing in Brazil. Oncol. 2018, 24. [Google Scholar] [CrossRef] [Green Version]
- Larsen, J.E.; Minna, J.D. Molecular Biology of Lung Cancer: Clinical Implications. Clin. Chest Med. 2011, 32, 703–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iorio, M.V.; Croce, C.M. MicroRNA dysregulation in cancer: Diagnostics, monitoring and therapeutics. A comprehensive review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.A.; Arora, S.; Prakasam, G.; Calin, G.A.; Syed, M.A. MicroRNA in lung cancer: Role, mechanisms, pathways and therapeutic relevance. Mol. Asp. Med. 2019, 70, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’Ayan, A. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y. An expression meta-analysis of predicted microRNA targets identifies a diagnostic signature for lung cancer. BMC Med. Genom. 2008, 1, 61. [Google Scholar] [CrossRef]
- Macfarlane, L.-A.; Murphy, P.R. MicroRNA: Biogenesis, Function and Role in Cancer. Curr. Genom. 2010, 11, 537–561. [Google Scholar] [CrossRef] [Green Version]
- Tsai, Y.-M.; Tsai, Y.-M.; Lien, C.-T.; Kuo, P.-L.; Hung, A.J.-Y.; Hung, J.-Y. The Roles of MicroRNA in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 1611. [Google Scholar] [CrossRef] [Green Version]
- Reis, P.; Drigo, S.A.; Carvalho, R.; Lapa, R.M.L.; Felix, T.F.; Patel, D.; Cheng, D.; Pintilie, M.; Liu, G.; Tsao, M.-S. Circulating miR-16-5p, miR-92a-3p, and miR-451a in Plasma from Lung Cancer Patients: Potential Application in Early Detection and a Regulatory Role in Tumorigenesis Pathways. Cancers 2020, 12, 2071. [Google Scholar] [CrossRef]
- Cinegaglia, N.C.; Andrade, S.C.S.; Tokar, T.; Pinheiro, M.; Severino, F.E.; Oliveira, R.A.; Hasimoto, E.N.; Cataneo, D.C.; Cataneo, A.J.M.; Defaveri, J.; et al. Integrative transcriptome analysis identifies deregulated microRNA-transcription factor networks in lung adenocarcinoma. Oncotarget 2016, 7, 28920–28934. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Ding, M.; Xia, M.; Chen, S.; Van Le, A.; Soto-Gil, R.W.; Shen, Y.; Wang, N.; Wang, J.; Gu, W.; et al. A Five-miRNA Panel Identified From a Multicentric Case-control Study Serves as a Novel Diagnostic Tool for Ethnically Diverse Non-small-cell Lung Cancer Patients. EBioMedicine 2015, 2, 1377–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caiazza, C.; Mallardo, M. The Roles of miR-25 and its Targeted Genes in Development of Human Cancer. MicroRNA 2016, 5, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Xiang, J.; Hang, J.-B.; Che, J.-M.; Li, H. miR-25 is up-regulated in non-small cell lung cancer and promotes cell proliferation and motility by targeting FBXW7. Int. J. Clin. Exp. Pathol. 2015, 8, 9147–9153. [Google Scholar] [PubMed]
- Ding, X.; Zhong, T.; Jiang, L.; Huang, J.; Xia, Y.; Hu, R. miR-25 enhances cell migration and invasion in non-small-cell lung cancer cells via ERK signaling pathway by inhibiting KLF4. Mol. Med. Rep. 2018, 17, 7005–7016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.-X.; Su, Y.; Zhang, H.; Kong, J.-Y.; Yu, H.; Qian, B.-Y. Prognostic Implications for High Expression of MiR-25 in Lung Adenocarcinomas of Female Non-smokers. Asian Pac. J. Cancer Prev. 2014, 15, 1197–1203. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Liu, Y.; Xiao, B.; Qian, X. miR-25 modulates NSCLC cell radio-sensitivity through directly inhibiting BTG2 expression. Biochem. Biophys. Res. Commun. 2015, 457, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Chen, T.; Li, Y.; Gao, L.; Zhang, S.; Wang, T.; Chen, M. Downregulation of miR-25 modulates non-small cell lung cancer cells by targeting CDC42. Tumor Boil. 2014, 36, 1903–1911. [Google Scholar] [CrossRef]
- Li, J.; Yu, M.; Liu, Z.; Liu, B. Clinical significance of serum miR-25 in non-small-cell lung cancer. Br. J. Biomed. Sci. 2019, 76, 111–116. [Google Scholar] [CrossRef]
- Peran, I.; Madhavan, S.; Byers, S.W.; McCoy, M.D. Curation of the Pancreatic Ductal Adenocarcinoma Subset of the Cancer Genome Atlas Is Essential for Accurate Conclusions about Survival-Related Molecular Mechanisms. Clin. Cancer Res. 2018, 24, 3813–3819. [Google Scholar] [CrossRef] [Green Version]
- Nicolle, R.; Raffenne, J.; Paradis, V.; Couvelard, A.; De Reyniès, A.; Blum, Y.; Cros, J. Prognostic Biomarkers in Pancreatic Cancer: Avoiding Errata When Using the TCGA Dataset. Cancers 2019, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Hamamoto, J.; Soejima, K.; Yoda, S.; Naoki, K.; Nakayama, S.; Satomi, R.; Terai, H.; Ikemura, S.; Sato, T.; Yasuda, H.; et al. Identification of microRNAs differentially expressed between lung squamous cell carcinoma and lung adenocarcinoma. Mol. Med. Rep. 2013, 8, 456–462. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.Y.; Yoon, S.O.; Seol, S.-Y.; Hong, S.W.; Kim, J.W.; Choi, S.H.; Lee, J.-S.; Cho, J.Y. Overexpression of miR-196b and HOXA10 characterize a poor-prognosis gastric cancer subtype. World J. Gastroenterol. 2013, 19, 7078–7088. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Yan, W.; Zhang, G.; Lv, H.; Liu, Z.; Fang, F.; Zhang, W.; Zhang, J.; Tao, T.; You, Y.; et al. Upregulation of miR-196b Confers a Poor Prognosis in Glioblastoma Patients via Inducing a Proliferative Phenotype. PLoS ONE 2012, 7, e38096. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-F.; Zhang, P.-F.; Li, M.-Y.; Li, Q.-Q.; Chen, Z.-C. Identification of annexin A1 as a proinvasive and prognostic factor for lung adenocarcinoma. Clin. Exp. Metastasis 2011, 28, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.-L.; Lee, D.C.; Sohn, H.A.; Lee, S.Y.; Jeon, H.S.; Lee, J.H.; Park, C.G.; Lee, H.Y.; Yeom, Y.I.; Son, J.W.; et al. Homeobox A9 directly targeted by miR-196b regulates aggressiveness through nuclear Factor-kappa B activity in non-small cell lung cancer cells. Mol. Carcinog. 2015, 55, 1915–1926. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Feng, C.; Shi, S. miR-196b promotes lung cancer cell migration and invasion through the targeting of GATA6. Oncol. Lett. 2018, 16, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Bishop, J.A.; Benjamin, H.; Cholakh, H.; Chajut, A.; Clark, D.P.; Westra, W.H. Accurate Classification of Non-Small Cell Lung Carcinoma Using a Novel MicroRNA-Based Approach. Clin. Cancer Res. 2010, 16, 610–619. [Google Scholar] [CrossRef] [Green Version]
- Patnaik, S.K.; Yendamuri, S.; Mallick, R.; Dhillon, S.S. Reply to “MiR-205 and miR-375 microRNA Assays to Distinguish Squamous Cell Carcinoma From Adenocarcinoma in Lung Cancer Biopsies”. J. Thorac. Oncol. 2015, 10, e53. [Google Scholar] [CrossRef] [Green Version]
- Solomides, C.; Evans, B.J.; Navenot, J.-M.; Vadigepalli, R.; Peiper, S.C.; Wang, Z. MicroRNA Profiling in Lung Cancer Reveals New Molecular Markers for Diagnosis. Acta Cytol. 2012, 56, 645–654. [Google Scholar] [CrossRef]
- Lebanony, D.; Benjamin, H.; Gilad, S.; Ezagouri, M.; Dov, A.; Ashkenazi, K.; Gefen, N.; Izraeli, S.; Rechavi, G.; Pass, H.; et al. Diagnostic Assay Based on hsa-miR-205 Expression Distinguishes Squamous From Nonsquamous Non–Small-Cell Lung Carcinoma. J. Clin. Oncol. 2009, 27, 2030–2037. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, P.; Hu, G.; Xiao, Z.; Xu, F.; Zhong, T.; Huang, F.; Kuang, H.-B.; Zhang, W. Relative expressions of miR-205-5p, miR-205-3p, and miR-21 in tissues and serum of non-small cell lung cancer patients. Mol. Cell. Biochem. 2013, 383, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Huang, Y.; Gong, W. miR-205 promotes the growth, metastasis and chemoresistance of NSCLC cells by targeting PTEN. Oncol. Rep. 2013, 30, 2897–2902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbst, R.S.; Heymach, J.V.; Lippman, S.M. Molecular origins of cancer: Lung cancer. N. Engl. J. Med. 2008, 359, 1367–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.; Dias-Santagata, D.; Duke, M.; Hutchinson, K.; Lin, Y.-L.; Borger, D.R.; Chung, C.H.; Massion, P.P.; Vnencak-Jones, C.L.; Iafrate, A.J.; et al. A Platform for Rapid Detection of Multiple Oncogenic Mutations with Relevance to Targeted Therapy in Non–Small-Cell Lung Cancer. J. Mol. Diagn. 2011, 13, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Goswami, R.S.; Waldron, L.; Machado, J.; Cervigne, N.K.; Xu, W.; Reis, P.; Bailey, D.; Jurisica, I.; Crump, M.; Kamel-Reid, S. Optimization and analysis of a quantitative real-time PCR-based technique to determine microRNA expression in formalin-fixed paraffin-embedded samples. BMC Biotechnol. 2010, 10, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Deng, M.; Brägelmann, J.; Kryukov, I.; Saraiva-Agostinho, N.; Perner, S. FirebrowseR: An R client to the Broad Institute’s Firehose Pipeline. Database 2017, 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Home—miRWalk. Available online: http://mirwalk.umm.uni-heidelberg.de/ (accessed on 20 August 2020).
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, B. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, 4. [Google Scholar] [CrossRef]
- Wong, N.; Wang, X. miRDB: An online resource for microRNA target prediction and functional annotations. Nucleic Acids Res. 2014, 43, D146–D152. [Google Scholar] [CrossRef]
- Chou, C.-H.; Shrestha, S.; Yang, C.-D.; Chang, N.-W.; Lin, Y.-L.; Liao, K.-W.; Huang, W.-C.; Sun, T.-H.; Tu, S.-J.; Lee, W.-H.; et al. miRTarBase update 2018: A resource for experimentally validated microRNA-target interactions. Nucleic Acids Res. 2018, 46, D296–D302. [Google Scholar] [CrossRef]
- Enrichr. Available online: https://maayanlab.cloud/Enrichr/ (accessed on 20 August 2020).
- Starruß, J.; De Back, W.; Brusch, L.; Deutsch, A. Morpheus: A user-friendly modeling environment for multiscale and multicellular systems biology. Bioinformatics 2014, 30, 1331–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morpheus. Available online: https://software.broadinstitute.org/morpheus/ (accessed on 20 August 2020).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | AD | SCC | p |
|---|---|---|---|
| Age | |||
| Mean (SD) | 60.91 (10.3) | 62 (7.1) | 0.725 |
| Range | 40–84 | 51–73 | |
| Sex | N (%) | N (%) | |
| Male | 13 (56.5) | 11 (73.4) | 0.329 |
| Female | 10 (43.5) | 4 (26.6) | |
| Smoking | N (%) | N (%) | |
| No | 7 (30.5) | 1 (6.7) | 0.114 |
| Yes | 16 (69.5) | 14 (93.3) | |
| Stage | N (%) | N (%) | |
| I | 9 (39.1) | 4 (26.7) | 0.651 |
| II | 6 (26.1) | 4 (26.7) | |
| III | 8 (34.8) | 6 (40) | |
| IV | 0 | 1 (6.6) | |
| Death | n (%) | n (%) | |
| Cancer-associated | 5 (62.5) | 4 (44.4) | 0.637 |
| Other causes | 3 (37.5) | 8 (56.4) |
| miRNA | FC | p Value of FC |
|---|---|---|
| miR-143-3p | 0.326 | <0.001 |
| miR-140-5p | 0.369 | 0.049 |
| miR-376c-3p | 2.057 | 0.001 |
| miR-141-3p | 2.060 | 0.004 |
| miR-20a-5p | 2.107 | 0.003 |
| miR-199a-3p | 2.199 | 0.003 |
| miR-374-5p | 2.260 | <0.001 |
| miR-130a-3p | 2.414 | < 0.001 |
| miR-29b-3p | 2.511 | <0.001 |
| let-7d-5p | 2.580 | <0.001 |
| miR-93-5p | 2.751 | <0.001 |
| miR-142-3p | 2.920 | 0.007 |
| miR-15a-5p | 3.016 | 0.017 |
| miR-155-5p | 3.056 | 0.005 |
| miR-25-3p * | 3.371 | <0.001 |
| miR-429 | 3.593 | <0.001 |
| miR-452-5p | 3.668 | 0.007 |
| miR-20b-5p | 3.917 | <0.001 |
| miR-135b-5p | 4.026 | <0.001 |
| miR-708-5p | 4.354 | <0.001 |
| miR-200b-3p | 4.446 | <0.001 |
| miR-340-5p | 5.345 | <0.001 |
| miR-744-5p | 5.649 | <0.001 |
| miR-365-3p | 5.766 | <0.001 |
| miR-205-5p | 5.856 | 0.001 |
| miR-590-5p | 6.112 | 0.001 |
| miR-224-5p | 6.692 | <0.001 |
| miR-15b-5p | 6.712 | <0.001 |
| miR-21-5p | 7.827 | <0.001 |
| miR-95-3p | 9.817 | <0.001 |
| miR-31-5p | 13.929 | 0.001 |
| miR-196b-5p | 16.525 | 0.001 |
| miR-411-5p | 25.909 | 0.033 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Souza, C.P.; Cinegaglia, N.C.; Felix, T.F.; Evangelista, A.F.; Oliveira, R.A.; Hasimoto, E.N.; Cataneo, D.C.; Cataneo, A.J.M.; Scapulatempo Neto, C.; Viana, C.R.; et al. Deregulated microRNAs Are Associated with Patient Survival and Predicted to Target Genes That Modulate Lung Cancer Signaling Pathways. Cancers 2020, 12, 2711. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12092711
Souza CP, Cinegaglia NC, Felix TF, Evangelista AF, Oliveira RA, Hasimoto EN, Cataneo DC, Cataneo AJM, Scapulatempo Neto C, Viana CR, et al. Deregulated microRNAs Are Associated with Patient Survival and Predicted to Target Genes That Modulate Lung Cancer Signaling Pathways. Cancers. 2020; 12(9):2711. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12092711
Chicago/Turabian StyleSouza, Cristiano P., Naiara C. Cinegaglia, Tainara F. Felix, Adriane F. Evangelista, Rogério A. Oliveira, Erica N. Hasimoto, Daniele C. Cataneo, Antônio J. M. Cataneo, Cristovam Scapulatempo Neto, Cristiano R. Viana, and et al. 2020. "Deregulated microRNAs Are Associated with Patient Survival and Predicted to Target Genes That Modulate Lung Cancer Signaling Pathways" Cancers 12, no. 9: 2711. https://0-doi-org.brum.beds.ac.uk/10.3390/cancers12092711