
State of the Art on the Microbial Production of Industrially Relevant Organic Acids
by
Raffaella Desirè Di Lorenzo
1,†, 
Immacolata Serra
1,†,
Danilo Porro
1,2,* and
Paola Branduardi
1,* 1
Department of Biotechnology and Biosciences, University of Milano Bicocca, Piazza della Scienza 2, 20126 Milano, Italy
2
Institute of Molecular Bioimaging and Physiology (IBFM), C.N.R. (National Research Council), Via F.lli Cervi 93, 20090 Segrate, Italy
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Catalysts 2022, 12(2), 234; https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020234
Submission received: 31 December 2021
/
Revised: 9 February 2022
/
Accepted: 15 February 2022
/
Published: 19 February 2022
(This article belongs to the Section Biocatalysis)
Abstract
:The industrial relevance of organic acids is high; because of their chemical properties, they can be used as building blocks as well as single-molecule agents with a huge annual market. Organic acid chemical platforms can derive from fossil sources by petrochemical refining processes, but most of them also represent natural metabolites produced by many cells. They are the products, by-products or co-products of many primary metabolic processes of microbial cells. Thanks to the potential of microbial cell factories and to the development of industrial biotechnology, from the last decades of the previous century, the microbial-based production of these molecules has started to approach the market. This was possible because of a joint effort of microbial biotechnologists and biochemical and process engineers that boosted natural production up to the titer, yield and productivity needed to be industrially competitive. More recently, the possibility to utilize renewable residual biomasses as feedstock not only for biofuels, but also for organic acids production is further augmenting the sustainability of their production, in a logic of circular bioeconomy. In this review, we briefly present the latest updates regarding the production of some industrially relevant organic acids (citric fumaric, itaconic, lactic and succinic acid), discussing the challenges and possible future developments of successful production.
1. Introduction
The current industrial system relying on fossil resources is putting at serious risk our living conditions on the planet. Globally, industry is responsible for over 30% of all greenhouse gas emissions, mainly represented by CO2, methane, nitrous oxide and fluorinated gasses (Sources of Greenhouse Gas Emissions—EPA). Most of these gasses arise from the production and use of bulk materials such as cement, metals, chemicals and petrochemical products, as well as from other human activities such as intensive farming. This description exemplifies the consequences of our model of growth, mainly based on a linear economy, which we thought to be sustainable when it was responding to the needs of a few billion people. Changing the paradigm from a linear to a circular economy and from the depletion of resources to exploitation of renewable biomasses is a promising alternative to reconnect human activities with biogeochemical cycles [1].
About 20 years ago, anticipating the current scenario, the U.S. Department of Energy (DOE) had identified twelve building-block chemicals [2]. Some years later, lactic acid was included in the list of the top-ten chemicals (Table 1) [3]. About 70% of the listed compounds are organic acids, which are good examples of building blocks. They find applications in the food & beverage industry, pharmaceutical and cosmetic segments, nutrition, and animal feed. In particular, chemical industries use organic acids as basic compounds for a wide variety of polymer and solvent production processes. In addition, they can be used as preservatives, as they can avoid the accumulation of reactive species, which are detrimental for cellular metabolism, if not for life [4,5]. It is not surprising, therefore, that the global annual organic acids market is projected to grow to about EUR 35 billion in 2027 [6].
Interestingly, organic acids can be produced by microbial metabolism, in some cases naturally, in some others after redirecting the carbon flux towards the desired target(s). Nonetheless, in terms of the production process, this implies non-trivial assumptions; the biocatalyst has to be able to transform a given biomass into a specific product, with maximized titer, yield and productivity. Furthermore, when commodities are produced, these parameters have to match with a final selling price of about or ideally below EUR 2 per kg [7]. Translating this concept into a feasible industrial process implies that the biocatalyst is forced to produce a non-natural product or, even if natural, to accumulate it up to non-physiological titer in a non-natural environment, possibly starting from a non-preferred substrate. In general, the engineered cell factories are proposed to overcome the main issues occurring during the determination of fermentation performances and, above all, during the scale-up process. Indeed, one of the main challenges remains the possibility to obtain strain(s) able to maintain the same performance of the lab-scale productions. The recent review by Wehrs et al. [8], provides a comprehensive analysis toward strains implementation. In particular, they underline the profound differences of the environment characterizing commercial-scale bioreactors if compared with laboratory-scale cultivation systems. The fluctuation and heterogeneity of chemical-physical parameters in large bioreactors and the genotypic instability or drift observed during long cultivations are prominent examples of issues that are often neglected or difficult to reproduce at lab-scale. This gap makes it extremely difficult to predict strains’ performances and consequently, optimal engineering design.
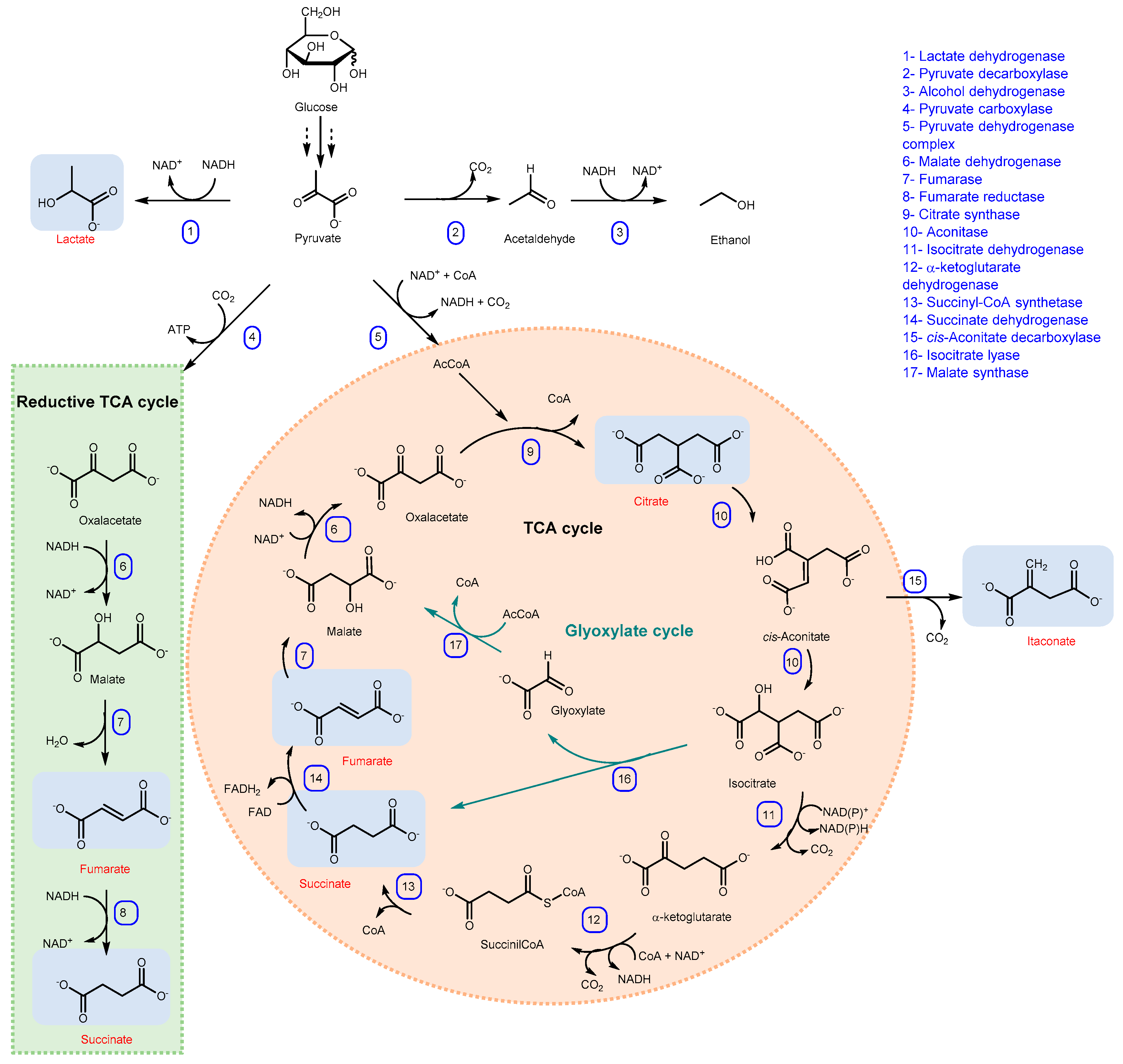
Despite the fact that these descriptions could lead to the conclusion of an unattainable goal, thanks to strain and process engineering, an increasing number of microbial processes are matching scale-up requirements and are emerging as real alternatives to chemical synthesis at industrial scale. Among the many organic acids that can be obtained by microorganisms, some have a prominent space in industry, both for tradition and in terms of competitiveness with traditional production: microbial citric, succinic, lactic, itaconic, lactobionic, gluconic, fumaric, propionic and acetic acid have approached the market, with different degrees of penetrance and success. From a chronological point of view, citric acid was the first organic acid industrially produced by a microbial cell factory, followed by lactic, fumaric, succinic and itaconic acid (Scheme 1).
In this review we present a state of the art on microbial processes for these organic acids which successfully reached the industrial, commercial, or piloting scale of production. Companies currently involved in this important challenge for our future will be listed and, when publicly available, information about the microbial strain currently in use will be given.
2. From First Evidence and Studies to Current Pilot or Commercial-Scale Production
2.1. Citric Acid (C6H8O7, 192.124 g·mol−1, pKa1 3.13, pKa2 4.76, pKa3 6.39)
Citric acid (CA, 2-hydroxy-propane-1,2,3-tricarboxylic acid) is an intermediate of the TCA cycle, and therefore present in virtually all living organisms.
CA was crystallized for the first time in 1784 by the Swedish chemist Karl Scheele; the procedure relied on treatment of the lemon juice with calcium hydroxide to obtain calcium citrate, subsequently treated with sulfuric acid to give citric acid [9].
Commercial production of citric acid started in England in 1826 from unripe Italian lemons and the Italian monopoly lasted until the late nineteenth century [10]. During World War I, export of lemons from Italy stopped and chemical production from glycerol (invented in 1880 by Grimoux and Adams [11]) took over. However, due to the increased importance of citric acid, fermentation became competitive enough and quickly replaced chemical synthesis. The first evidence of microbial production of CA dates back to 1893, when Wehmer observed that Penicillium glaucum could produce citric acid from sugars, and two years later, two strains of Penicillium, at that time named Citromyces spp., were isolated [12]. Unfortunately, production with Citromyces was never industrially significant because of the occurrence of contaminations and the long fermentation time. Later, Currie in 1917 found that, in various strains of Aspergillus niger, the production and secretion of CA was stimulated by growing them in a nutrient medium with an initial pH of 2.5–3.5 [13]. Afterwards, in 1919, the pharmaceutical company Pfizer, in cooperation with Currie, started the first pilot plant for CA production in New York using the fermentation process developed by the chemist and named SUCIAC “Sugar Under Conversion to Citric Acid”. Already in mid-1920 the fermentative production of CA surpassed the extractive production from lemons [14,15].
Citric acid is a colorless, odorless, and highly water-soluble compound. It is safe for human health and the environment and is considered almost indispensable and hard to substitute in light of the number of applications in very different sectors, such as food and beverages, the pharmaceutical, nutraceutical and cosmetics industries. The great majority of the produced citric acid (>65%) is addressed to the food and beverage (F&B) sector where it is employed both as a preservative, thanks to its antioxidant properties, and as flavor enhancer, used to balance the sweetness of juices, soft drinks and jams [16,17]. CA is certified as safe (Generally Regarded As Safe) by the Food and Drug Administration in the US and, together with its salts, is EU-approved as a food additive, being indicated as E330 (citric acid), E331 (sodium citrate), E332 (potassium citrate) and E333 (calcium citrate).
The second main user is the pharmaceutical sector, where CA is used in the preparation of effervescent tablets, as an anticoagulant in blood transfusions, as a crosslinking agent in films for controlled drug release, and as a palatability enhancer of unpleasant drugs. Moreover, many APIs are administered in the form of citrate salts [17]. Other uses deal with the detergent, cleaning and polymer industries and environmental remediation, and have been extensively reviewed by Ciriminna at al. [17]. More recently, the use of CA as one of the components of deep eutectic solvents, promising in the valorization of residual lignocellulosic biomasses, was proposed [18].
Many microorganisms have been found to produce and accumulate CA. For instance, some bacteria (such as Bacillus sp., Arthrobacter sp., Corynebacterium sp., Brevibacterium sp.), yeasts (such as Candida tropicalis, C. oleophila, Yarrowia lipolytica, Hansenula anomala) and filamentous fungi, mostly belonging to the Aspergillus genus, have been reported as CA producers [19,20]. Despite this wide portfolio, most of these microorganisms are not able to afford commercially exploitable amounts of CA, and thus, A. niger is still the preferred workhorse for productive purposes thanks to its robustness in industrial fermentation conditions. In fact, more than 80% of the world’s citric acid production is derived from A. niger, although some processes with the yeast Y. lipolytica have been reported [21]. The yeast-based process is, however, plagued by the significant production of by-products, such as isocitric acid, that inevitably lower the final product yield.
Citric acid is produced during the TCA cycle through the condensation of oxaloacetate and acetyl-CoA, resulting in 1 mol of CA from each mol of glucose with a maximum theoretical yield of 1.067 g/g glucose (Scheme 2). Although the enzymatic activities responsible for CA production have been identified and investigated, much less is known about the biochemical and physiological aspects of CA accumulation [22]. In fact, the exact mechanism underlying CA accumulation in A. niger has not been completely understood; several hypotheses have been proposed and the debate is still open. It is clear that CA overflow is the result of the synergistic effect of different imbalanced conditions, such as low pH, low iron concentration, limited availability of nitrogen and phosphate, and excess oxygen and carbon sources [10].
A. niger can use several substrates for its growth (fructose, glucose, galactose, mannose, glucose); however, glucose and sucrose, with a slight preference for the latter, are the preferred carbon sources for CA production [10]. Sucrose was demonstrated early on to be the best carbon source, and CA production still mainly relies on sucrose-based feedstocks. The use of molasses as residual carbon source substrate was reported already in 1948 at an industrial scale [23]. However, due to the impellent need to make production of CA more sustainable and less dependent on sucrose and sucrose-based feedstocks, the use of various carbon-rich residual biomasses has been evaluated. The exploitation of residual biomasses with different polymeric compositions is possible thanks to the presence in A. niger of a pool of extracellular hydrolyzing enzymes, such as invertase, glucoamylase, α-glucosidase and α-amylase. Several examples, both in scientific literature and patents, describe the use of corn, fruits, lignocellulosic and starchy materials [19,24,25,26,27,28] (Table 2).
The availability of high-producing strains is essential from an industrial point of view in order to develop highly efficient processes. Until recently, the main strategy for strain improvement was random chemical or physical mutagenesis followed by screening. These protocols, although time consuming, successfully allowed the improvement of CA yields. For instance, a combination of UV exposure, ethyl methane sulfonate (EMS) and acridine orange treatment to A. niger UMIP 2564 resulted in a 3.2-fold increase in product yield [29]. Adeoye et al. reported a 45.97-fold increase in citric acid production by A. niger, FUO 2 strain, subjected to UV radiation and cultivated on cassava peel substrate [30].
The release in 2007 of the first A. niger genome data (strain CBS 513.88) [31] paved the way to more targeted modifications such as those aimed at modifying carbon source utilization and uptake [32,33], enhancing product secretion [34], and modifying the mycelial morphology of the fungus, which is known to be critical for CA production in submerged fermentations [35]. Other attempts were made by improving the efficiency of the mitochondrial electron transport chain [36] or by inserting genes involved in the reductive branch of the tricarboxylic acid (rTCA) cycle [37].

{kind=link}
{kind=link}
Table 2.
Citric acid production with different substrates, microorganisms, and fermentation strategies.
Table 2.
Citric acid production with different substrates, microorganisms, and fermentation strategies.
| Microorganism | Substrate | Citric Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| A. niger MH 15-15 | Glucose | 31 | Shake flask | [38] |
| A. niger MH 15-15 | Sucrose | 53 | Shake flask | [38] |
| A. niger | Corn stover | 136.3 | Simultaneous saccharification and fermentation | [39] |
| A. niger GMCC 5751 | Liquefied corn | 151.7 | Batch | [40] |
| Adapted A. niger | Corn powder and citric acid effluent | 157.3 | Batch | [41] |
| Mutant A. niger MO-25 | Beet molasses and chicken feather peptone | 68.8 | Shake flask | [28] |
| Mutant A. niger UMIP 2564 | Sucrose | 96.3 | Shake flask | [29] |
| Mutant A. niger UMIP 2564 | Beet molasses | 98.3 | Shake flask | [29] |
| Mutant A. niger UMIP 2564 | Cane molasses | 91.2 | Shake flask | [29] |
| A. niger 831f | Liquefied cassava | 162.7 | Repeat-fed batch | [42] |
| Y. lipolytica NRRL YB-423 | Glycerol | 21.6 | Shake flask | [43] |
Production of CA with A. niger can be achieved by three different fermentation strategies: submerged, surface and solid-state fermentation, with each method having advantages and disadvantages.
Liquid surface fermentation (SF) was the original technique used for the first industrial manufacture since 1919, and until recently, some small- and medium-scale production plants adopted this method because of the low installation and operation costs. Typically, fermentation is carried out in trays with a capacity of 50–100 L, mounted one over another in aseptic chambers with controlled humidity and temperature. After spore germination, which is usually completed within the first 2 days, the fungus grows as mycelial mat. Finally, after 8–12 days of fermentation, the liquor can be separated from the mycelia and further processed for CA recovery [44].
Solid-state fermentation (SSF), also known as the Koji process, employs a solid insoluble material that acts both as substrate and support in a low-water activity environment. Different reactor configurations can be employed, such as flasks, trays, horizontal drums and glass columns [45]. SSF for CA production has several advantages over submerged and surface fermentation, since it allows the exploitation of agro-food waste materials [25] and is intrinsically correlated with lower energy and water consumption. Conversely, during SSF it is difficult to have full control of the process parameters; therefore, it is difficult to scale up and to standardize, requires longer process time and higher overall costs for product recovery compared to other strategies. For these reasons, the SSF production mode is still far from being industrially applicable [44].
Currently, most citric acid globally is produced by submerged fermentation (SmF) with either stirred tank or bubble-column reactors. Compared to surface fermentation, submerged fermentation requires more sophisticated equipment but, in turn, is characterized by higher yields, lower fermentation times, ease of process standardization and automation, and less risk of contamination [16,44].
A crucial step in CA production is represented by the downstream process, which has been estimated to account for 30–40% of the selling price [46]. It must be considered that the complexity of the procedure can be greatly influenced by the type of fermentation (SSF, SF or SmF) and by the fermentation substrate; for instance, the use of agro-food wastes may require additional steps to obtain a high-purity product. At the end of the fermentation, microbial biomass is removed by filtration, after which the broth can be subjected to different treatments to obtain the purified citric acid. The classical method for CA recovery, which remains the most used, relies on tri-calcium citrate salt precipitation caused by the addition of calcium oxide at 90 °C and pH 7, followed by treatment with sulfuric acid that results in soluble citric acid and calcium sulfate [16]. Calcium sulfate is filtered off, whereas citric acid undergoes further purification steps. An alternative method for CA recovery is based on liquid–liquid extraction with various solvents (such as aliphatic alcohols, ketones and tertiary amines) [47,48]. More recently, other methods have been proposed, such as supercritical CO2 [49], adsorption on cationic resins and electrodialysis [50,51].
A recent techno-economic and environmental analysis on CA production comparing recovery by precipitation, ion exchange and solvent extraction indicated that the ion exchange method is the most environmentally friendly, whereas solvent extraction, followed by precipitation, are the most economically convenient. Other factors having a large impact on the final selling price were electricity cost, by-product credit, feedstock cost, fermenter cost and wastewater treatment cost [52].
Global citric acid production rose from 0.5 million tons in 1992 [53] to more than 2.55 million tons in 2020, and a moderate growth is expected in 2021–2026 [54].
The fermentative process used nowadays for citric acid production is mainly via submerged fermentations of A. niger grown on molasses and/or other carbohydrates. The main producers (in alphabetic order) are: Archer Daniels Midland-ADM (Chicago, IL, USA), Cargill (Minneapolis, MN, USA), COFCO Biochemicals (Beijing, China), FoodChem (Shanghai, China), Jungbunzlauer (Basel, Switzerland) and Tate and Lyle (London, UK).
In the last two decades, production was mostly located in China [16,17], which is now acting as the major player in citric acid production, as can be easily inferred by patent analysis over the last two decades. In fact, China has >80% of the total applied patents regarding citric acid production and downstream [16].
2.2. Fumaric Acid (C4H4O4, 116.07 g·mol−1, pKa1 3.03, pKa2 4.44)
Fumaric acid (FA) is a trans four-carbon dicarboxylic acid, intermediate in the TCA cycle, isolated for the first time from the plant Fumaria officinalis.
Fumaric acid is a specialty chemical with a number of applications in very diverse fields. It is used as an additive and acidulant in food and beverages; being 1.5-fold more acidic than citric acid, lower amounts are required (0.91 g of fumaric acid vs 1.36 g of citric acid are required to achieve the same taste [55]). Moreover, it is supplemented into ruminal feed to decrease methanogenesis [56] and in quail’s feed has shown a number of benefits including improved growth, immune response and intestinal health [57]. Importantly, fumaric acid has two carboxylic groups and one double bond, characteristics that make it suitable for esterification and polymerization to produce, to cite some, paper resins, unsaturated polyester resins (UPR), biodegradable polymers, plasticizers and carboxylating agents for rubber [58]. Recently, its esters (FAEs) have shown important pharmaceutical applications in the treatment of sclerosis and psoriasis [59,60].
FA is mainly produced starting from petroleum-derived chemicals (mainly benzene, n-butane, n-butene) by conventional chemical processes based on the isomerization of maleic acid, obtained, in turn, from the hydrolysis of its anhydride [58]. Alternatively, enzymatic processes based on the activity of maleate isomerases (from Bacillus, Pseudomonas or Arthrobacter) on maleic acid have been developed [61,62]. However, this route had poor success as confirmed by the few available reports.
The first industrial production of FA was started by Pfizer using Rhizopus arrhizus in the 1940s, with a production of 4000 tons/year [58]. Later, the fermentative route was discontinued and replaced by the more economically convenient classical chemical route. However, recently, as for several other platform chemicals, the shortage of fossil resources and the need for green and sustainable processes gave a new boost to microbial production.
The most relevant species from a productive point of view are those belonging to the genus Rhizopus (arrhizus, oryzae, nigricans and formosa), which can accumulate and secrete large amounts of fumaric acid through the cytosolic reductive branch of the TCA (rTCA) pathway under aerobic conditions and limited nitrogen availability [58] (Scheme 2). However, not all R. oryzae strains are able to produce FA. In fact, some studies revealed that those strains able to produce FA did not belong to R. oryzae and should be reclassified as R. delemar [63], however, to avoid confusion, they are still named as R. oryzae. The rTCA cycle takes place in the cytosol starting from pyruvate and involves three reactions: (i) condensation, in presence of ATP, of pyruvate and carbon dioxide catalyzed by pyruvate carboxylase to produce oxaloacetic acid; (ii) conversion of oxaloacetic acid into malate by malate dehydrogenase; (iii) conversion of malate to fumarate catalyzed by fumarase. However, the rTCA cycle does not lead to the production of ATP, which must be produced by the oxidative branch of the TCA cycle in mitochondria or by ethanol formation. When nitrogen becomes limiting and cell growth stops, glucose metabolism and CO2 fixation continue, leading to the accumulation of fumarate [58]. The simultaneous occurrence of both TCA cycles was demonstrated in R. arrhizus also by carbon labeling studies [64]. The rTCA cycle has a theoretical maximum yield in non-growth conditions of 2 mol of FA per mol of glucose (1.3 g/g); however, experimental yields are much lower. This is mainly ascribed to the co-existence of the TCA cycle that fulfils the cell energy requirements and to the formation of fermentation by-products such as ethanol and lactic acid [65].
Although high concentrations of fumaric acid have been obtained using recombinant strains of R. oryzae [66,67], Escherichia coli [68,69], Saccharomyces cerevisiae [70,71], and Torulopsis glabrata [72], the major titer of fumaric acid is ascribable to the native fumaric acid-producing Rhizopus species [73], which are at present the main producers at industrial scale, and most of the research on fumaric acid is based on these microorganisms. So far, the highest titer of FA (>130 g/L) was obtained by Du Pont with R. arrhizus NRRL 1526 using glucose as the carbon source as claimed in the US patent US4877731A [74].
Glucose is indeed the most used carbon source for processes with Rhizopus species. However, to improve the sustainability of the process, several efforts were addressed to assess the use of alternative substrates and, in particular, of residual biomasses. Sebastian et al. reported the production of FA starting from perennial grasses hydrolysates using R. oryzae strains [75] with 0.3–0.5 g of FA per g of sugar. Production of FA was also achieved with R. oryzae via fed-batch SSF and SHF processes employing alkali-pretreated corncob [76]. Other examples deal with the use of food wastes [77,78], starchy materials [79,80,81], lignocellulosic materials [82,83], crude glycerol [84] and brewery wastewater [85] (Table 3). Two of the most relevant key factors for process optimization of FA production are the control of fungi morphology and of the pH of the production medium. Morphology of the fungi is considered one of the major technical challenges during fumaric acid production in submerged fermentations, and it is one of the most important variables that may hamper industrial scale production. In fact, during their growth, filamentous fungi can exhibit three different morphologies: pellets, filaments, and clumps. Filamentous mycelium is the most productive morphology; however, its formation causes operational problems due to the high viscosity of the broth. Clumps are the less productive form, because their inner part suffers from nutrient and oxygen limitations, leading to the production of ethanol and other fermentation by-products. Pellet morphology is preferred over the others since it allows efficient mass transfer while avoiding the increase in medium viscosity. Interestingly, it was observed that a lower pellet diameter corresponds to a more efficient mass transfer of both nutrients and oxygen, and thus increased yield [86]. Several reports showed how fungi morphology can be controlled by acting on different parameters, such as nitrogen source, spore concentration in the inoculum, pH, temperature and agitation rate [87,88,89,90]. Moreover, immobilization of cells onto a solid support was also proposed as a method to overcome morphology problems. Different metallic nano- and micro-particles were evaluated for their ability to influence R. oryzae 1526 morphology and pellet size. Microparticles of Fe3O4 at a concentration of 200 μg/mL were found to be the most effective providing the highest FA titer (42 g/L) [91]. Unusual materials were also exploited as immobilization supports; for instance, R. arrhizus RH 7-13-9 immobilized on loofah fibers allowed to achieve 30 g/L of fumaric acid when cultivated in a stirred tank reactor [92]. Another example by Naude et al. reports the use of R. oryzae cells attached to a polypropylene tube for continuous fermentation [93].
Fumaric acid production is strongly affected by the fermentation pH which, if not properly controlled through the addition of a neutralizing agent, quickly drops from 5.0 to 2.0 within the first 20 h of fermentation. The most frequently used neutralizing agent is CaCO3; however, its use is correlated with several issues. First, calcium fumarate is poorly soluble at 30 °C, and is consequently present as a precipitate that aggregates with cells, resulting in a high viscous suspension. Secondly, the conversion of the carboxylate salt to the free carboxylic acid requires the addition of H2SO4 which produces CaSO4 (gypsum), which is associated with several environmental concerns [65]. Unfortunately, the use of other neutralizing agents (i.e., Ca(OH)2, NaHCO3) is associated with lower yields and productivity [95]. In particular, sodium fumarate is highly soluble in water and high fumarate concentration may cause product inhibition [96]. Moreover, Na+ ions can in some cases negatively affect the cell metabolism [95].
An efficient strategy for pH control during fermentation is represented by the simultaneous fermentation-separation process (SFS), that, in addition, allows the alleviation of product inhibition phenomena, which cause reduced yields and productivity [94].
FA is produced in about 90,000 tons/y; MBI international (Lansing, MI, USA) uses the fungus R. oryzae for production [97], while for Myriant corporation (Quincy, MA, USA), there are indications about the development of different engineered strains of E. coli and S. cerevisiae for organic acids production, including fumaric acid, from specific carbon sources [98,99].
2.3. Itaconic Acid (C5H6O4, 130.10 g·mol−1, pKa1 3.85, pKa2 5.45)
Itaconic acid (IA) is an unsaturated dicarboxylic acid, also known as methylene succinic acid. It was firstly discovered in 1836 by Baup during the distillation of citric acid. However, the name itaconic acid its due to Crasso, who synthesized it through decarboxylation of cis-aconitic acid [100].
Itaconic acid can be considered mainly as a chemical building block, as it is exclusively used in non-food applications. It is used in styrene-butadiene rubber, synthetic latex, methyl methacrylate (MMA), unsaturated polyester resins (UPR), and as a chelant dispersant [101,102].
The production of IA by microbial fermentation is preferred to chemical synthesis, which is long, scarcely efficient and expensive because it is carried out starting from a substrate, citric acid, with the same cost of itaconic acid, and it requires high temperature, implying high energy consumption [103,104]. Itaconic acid is produced in about 41.400 tons/y. The first industry plant was founded by Pfizer Co. Inc (Brooklyn, 1955), which also developed a patented aerobic fermentation process to produce itaconic acid [101]. While in the past decades the majority of the itaconic acid production was ascribable to companies located in different parts of the world, such as Cargill, Iwata Chemicals and Melle, currently it is completely in the hands of Chinese industries [105,106].
The fungus Aspergillus terreus is known to be the best IA producer (about 160 g/L) [107], also at commercial level, for its extraordinary tolerance to low pH, high yield and titer of product, as described by [103,108]. In the last ten years, the basidiomycete Ustilago maydis, also known as a natural producer, due to its similarity to yeasts, gained attention as a candidate for large scale production, even if the IA yield on glucose is lower than that of A. terreus (0.2 g/g versus 0.48 g/g on glucose, respectively) [109,110,111] (Table 4).
Moreover, some other yeasts and fungi have been exploited as natural producers of itaconic acid, but have never reached the commercial level (i.e., Candida sp. and Pseudozyma antarctica) [112,113].
From a metabolic point of view, IA production from glucose in A. terreus mainly involves glycolysis and TCA (tricarboxylic acid cycle), where pyruvate produced by glycolysis is transported in the mitochondrial compartment to become acetil-CoA, which together with oxaloacetate, catalyzes the reaction to obtain citrate, and further into cis-aconinate, a substrate of cis-aconitate decarboxylase (CAD) responsible for the decarboxylation in itaconic acid (Scheme 2) [114]. IA is then transported outside the cell by a facilitator superfamily protein (mfsA gene) [115]. Differently, for the filamentous fungus Ustilago maydis, due to the lack of the CAD gene, the decarboxylation of cis-aconitate into itaconic acid occurs by the action of trans-aconitate decarboxylase (Tad1), preceded by isomerization through the cytosolic enzyme aconitate-D-isomerase (ADI). Afterwards, the membrane transport protein ITP1 allows the excretion of itaconic acid through the cell wall [116]. It is important to highlight that, since decarboxylation takes place in the cytosol, while cis-aconitate is produced in the mitochondrion, the action of a mitochondrial tricarboxylate transporter protein (Mtt) is necessary as well [102]. During the years, engineering strategy and/or mutagenesis techniques were developed to increase itaconic acid production [104]. The native producer of itaconic acid U. maydis was engineered by overexpressing the genes involved in itaconic acid production in A. terreus (rai1 and mttA) and deleting genes involved in the synthesis of by-products (cyp3, dgat, MEL and UA). The resulting strain, grown on glucose with a continuous feeding strategy, showed an itaconic acid titer and yield of 74.9 g/L and 0.54 g/g, respectively [117]. In addition, non-native producers of itaconic acid, due to some interesting characteristics for the fermentation (e.g., organic acid tolerance and production of a considerable amount of TCA intermediates), have also been considered for genetic modifications. The low pH-tolerant Pichia kudriavzevii, already engineered to produce some organic acids, has undergone multiple genetic modifications (introduction of CAD gene from A. terreus, overexpression of a native mitochondrial tricarboxylate transporter (mttA), and deletion of the gene encoding isocitrate dehydrogenase (ICD)), to establish itaconic acid production [118].
Table 4.
Itaconic acid production with different substrates, microorganisms, and fermentation strategies.
Table 4.
Itaconic acid production with different substrates, microorganisms, and fermentation strategies.
| Microorganism | Substrate | Itaconic Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| A. terreus DSM 23081 | Glucose | 160 | Fed-batch | [107] |
| A. terreus NRRL 1960 | d-Xylose | 53.97 | Batch | [119] |
| A. terreus NRRL 1960 | Bleached eucalyptus pulp | 37.5 | Shake flask | [120] |
| U. maydis MB215 | Glucose | 44.5 | Batch | [109] |
| Mutant U. maydis MB215 | Glucose | 80 | Fed-batch | [121] |
| Mutant U. maydis MB215 | Glucose | 74.9 | Continuous feeding | [117] |
| U. vetiveriae TZ1 | Glycerol | 34.7 | Batch | [122] |
| A. terreus CICC40205 | Wheat bran hydrolysate | 34.2 | Shake flask | [123] |
| A. terreus M69 | Corn stover hydrolysate | 33.6 | Shake flask | [124] |
Furthermore, the best citric acid producer A. niger was engineered by the introduction of cadA and acoA genes, two known key enzymes for itaconic acid production, which allowed IA production by an extension of the citric acid production pathway [125].
From an economical point of view, since the maximum yield of IA is reached by glucose fermentation of A. terreus, alternative carbon sources should be used to make the process competitive with petrochemical-derived products. Unfortunately, low-cost substrates (including potato starch, plant-derived hydrolysate, lignocellulosic biomasses) are related to nutrient deficiency and toxicity problems, which lead to poorly competitive yields compared to the ones obtained with pure sugars [126]. In addition, most residual biomasses require pre-treatment steps in order to be metabolized by the cells. A recent review about itaconic acid [101] reported different IA producer strains (native or not) modified in order to exploit some residual biomasses as alternative carbon sources.
2.4. Lactic Acid (C3H6O3, 90.07948 g·mol−1, pKa1 3.78)
Lactic acid (LA) was first isolated from sour milk by the Swedish chemist, Karl Wilhelm Scheele, in 1780, but only in 1857 did the chemist and microbiologist Louis Pasteur attribute its production to certain microorganisms [128]. LA appears colorless to yellow at 15 °C and at a pressure of 1 atm [129]. It is known to occur in two isomeric forms, (l(+)- or d(−)), due to the chiral carbon that confers optical activity to the monomer, as well as the racemic form, DL-Lactic acid [130].
The production of LA occurs by chemical synthesis or fermentation. Microbial fermentation is very attractive for lactic acid production since it allows to obtain pure isomers (l(+)- or d(−)-LA) instead of racemic mixture (dl-LA), with the latter being typical of the chemical synthesis [131]. Furthermore, the chemical synthesis involves the use of polluting substances (e.g., lactonitrile and the highly toxic hydrogen cyanide), which have high costs of production too [129]. These days, more than 90% of lactic acid is produced by fermentation processes [132]. Currently, many companies are involved in LA production at the commercial level: ADM (Chicago, IL, USA), Aldon Corporation (Avon, NY, USA), Anhui COFCO Biochemical and GALACTIC (Bengbu, China), Cargill (Minneapolis, MN, USA), ChonggingBofei Biochemical Products (Chongging, China), CorbionPurac (Amsterdam, The Netherlands), Galactic (New Orleans, LA, USA), Henan Jindan Lactic Acid Technology (Dancheng Henan, China), Musashino Chemical (Tokyo, Japan), Nature Works LLC (Minnetonka, MN, USA), Symbra Technology BV (Etten-Leur, The Netherlands), Tedia Company Inc (Fairfield, OH, USA), Yangtze Labre (Yangtze, China) [133]; among these, the biggest producers at industrial scale are Corbion and Cargill (producing about 240,000 and 180,000 tons, respectively) [134]. LA is mostly used as monomer of the biobased and biodegradable polylactic acid (PLA) for biodegradable plastics. PLA is today considered among the most promising polymers for bioplastics. PLA finds applications in packaging, agriculture, transport, electronics, textiles, and others. At present, the main suppliers of PLA are Cargill (Minneapolis, MN, USA) Galactic (Escanaffles, Belgium), Purac, (Amsterdam, The Netherlands), FKuR (Willich, Germany), and Shimadzu (Kyoto, Japan) [135,136].
The microorganisms used for microbial fermentation are different, depending on the isomer to be produced. Several microorganisms can produce pure L-LA, including bacteria, fungi, cyanobacteria, algae and yeasts [137,138]. Moreover, other strains are known for d-LA production, for instance Lactobacillus delbrueckii, Sporolactobacillus, Escherichia coli, and Bacillus coagulans [139] (Table 5).
Although other microorganisms have been identified as lactic acid producers, as described above, nowadays lactic acid bacteria (LABs) still represent the best producers at industrial scale [136]. They produce LA by glycolysis (Scheme 2) in anaerobic conditions, although they are micro-aerophilic or aerotolerant as well, at a temperature in the range of 25–45 °C (depending on the microorganism), and a pH between 5.5 and 6.5 [133,140]. It is possible to distinguish homofermentative LABs, belonging to the genera Streptococcus, Lactococcus, Enterococcus, Pediococcus and Lactobacillus, which produce lactic acid as main product, and heterofermentative LAB (as L. brevis, L. fermentum and L. reuteri) in which, together with lactic acid, there are other by-products such as ethanol, acetic acid and carbon dioxide. At the metabolic level, homofermentative LABs produce two molecules of LA per mole of glucose with a theoretical yield of 1 g/g through the Embden-Meyerhof-Parnas pathway, whereas the yield for heterofermentative LABs is 0.5 g/g or 0.6 g/g, depending on the pathway, phosphogluconate and phosphoketolase, respectively [136,141].
Generally, homofermentative strains use hexose sugars such as glucose to produce lactic acid as the only product, whereas pentose sugars such as xylose can enter in the phosphoketolase pathway to obtain lactic acid and other by-products, which leads to yield reduction and increase in the recovery cost [133]. Since homofermentative LABs can reach yield values very close to the maximum theoretical one, they were considered for industrial production.
In order to optimize lactic acid production, some aspects that can affect the fermentation process should be considered, including temperature, sugar concentration, effect of nutrients, mixed sugars, and by-product formation [133].
A critical role is held by the pH of the fermentation: the preferred form of the product is the undissociated one, which means that the final pH of the medium should be well below the pKa of LA, corresponding to 3.78. However, at this pH, most LABs show irreversible damage to their metabolic functions [142]. Therefore, several neutralizing agents are added to the fermentation broth to maintain the pH and protect the microbial cells. Unfortunately, the use of Ca(OH)2 or CaCO3 causes the formation of calcium lactate that must be removed through acid treatment (H2SO4), generating, as discussed above for other organic acids, calcium sulfate (gypsum), a waste product with serious disposal concerns [143,144].
For the successful production of LA, different modification technologies were considered. More recently, Tian et al. described the application of mutagenesis, adaptive evolution and metabolic engineering on lactic acid bacteria [145].
Along with LAB, other microorganisms have been evaluated as potential lactic acid producers. Some of the most interesting are the Bacillus strains which are able to grow at high temperatures (up to 55 °C), avoiding further sterilization of the culture media, and allow the exploitation of second-generation biomasses, since they may tolerate some by-products generated by pre-treatments [146,147,148].
In addition to bacteria, fungi, for example those belonging to the genus Rhizopus [141], and yeasts have attracted attention for their intrinsic ability to tolerate low pHs.
In respect to yeasts, while the initial preference was for the budding yeast Saccharomyces cerevisiae, which was the first engineered for lactic acid production [149], other yeasts have been exploited.
The natural fermentation occurring in yeasts leads, in most cases, to ethanol and CO2 formation, therefore there is a need to re-route pyruvate to the desired product. The engineering strategy most often used is the overexpression of a heterologous lactate dehydrogenase gene (LDH), coupled, in some cases, to the partial or complete deletion/inactivation of pyruvate decarboxylase (PDC) genes [150,151]. These modifications were often combined with further engineering or evolution to overcome poor growth on glucose related to the deficiency in 2C intermediates, usually deriving from acetaldehyde (as reviewed in [152]), and with the ability to tolerate low pH. Recently, Park at al. described the high titer of d-lactic acid (154 g/L) reached by the genetically modified yeast Pichia kudriavzevii (also known as Issatchenkia orientalis) [153], but the pH of the medium of 4.7 cannot assure the advantage of having the product in undissociated form.
Lactic acid production can be also improved by co-culture of strains with similar growth characteristics, allowing the cultivation of homofermentative strains which can take the advantage from heterofermentative ones, which normally produce low amounts of lactic acid due to the accumulation of other products [154]. Co-cultures are an interesting perspective also for the simultaneous saccharification and fermentation (SSF) strategy, and for the use of mixed sugars; both are typical conditions when raw materials are considered as fermentation substrate [133].
The increasing demand for lactic acid is due to the spread of end-products industries which include food & beverages, cosmetics, pharmaceuticals, polymers [133]. It is estimated that, by 2028, the market for lactic acid will reach USD 5.02 billion, and is expected to grow by about 8.0% from 2021 to 2028 [155].
For a commodity such as LA, it is crucial to consider all the factors which contribute to the final cost, and at the same time, it is relevant to take sustainability into account, as large production is expected. The use of cheap raw materials, which do not release dangerous by-products after pre-treatments, could be the ideal condition for economic lactic acid production [156]. Among the second-generation biomasses, the lignocellulosic ones are the most interesting for LA production due to their abundance [129]. However, they need pretreatments and enzymatic hydrolysis to release the sugars, toxic compounds can be released along with these steps, and the final purification of LA is more complex. All these elements need to be accurately considered. Recently, in order to analyze all the variables related to lignocellulosic biomasses pre-treatment and use for LA fermentation, Li et al. exploited the BioSTEAM platform to conclude that the optimization of separation process and fermentation is essential for a successful production process [157].
Indeed, the downstream affects the final cost of lactic acid, of which 50% is ascribable to recovery and purification [158]. As already discussed above, due to the decrease in fermentation pH as result of LA accumulation, neutralizing agents are used for LAB-based production [133]. This method is well known as precipitation and is still the conventional one for LA separation today [144]. Alternatively, MgO can substitute Ca(OH)2 or CaCO3, with the advantage that it can be reused after the lactic acid extraction. Li et al. recently described the pros and cons of the separation and purification techniques known today for LA [159]. Certainly, the recovery process is extremely complex considering the composition of the fermentation broth (cells, sugars, inorganic salts, and other organic acids) [160].
Table 5.
Lactic acid production with different substrates, microorganisms and fermentation strategies.
Table 5.
Lactic acid production with different substrates, microorganisms and fermentation strategies.
| Microorganism | Substrate | Lactic Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| Mutant P. kudriavzevii | Glucose | 154 (d-Lactic acid) | Fed-batch | [153] |
| Evolved L. delbrueckii S-NL31 | Soybean meal | 112.3 (d-Lactic acid) | Fed-batch | [161] |
| Evolved and mutated S. cerevisiae | Glucose | 82.6 (d-Lactic acid) | Fed-batch | [162] |
| Mutant S. cerevisiae | Glucose | 40 (d-Lactic acid) | Fed-batch | [163] |
| Mutant E. coli | Glycerol | 115 (d-Lactic acid) | Fed-batch | [164] |
| L. delbrueckii ssp. bulgaricus | Orange peel waste | 45 (d-Lactic acid) | Separate hydrolysis and fermentation | [165] |
| L. delbrueckii | Molasses and corn steep liquor | 162 (d-Lactic acid) | Fed-batch | [166] |
| B. coagulans A107 | Defatted rice bran (DRB) | 75.9 (l-Lactic acid) | Batch | [137] |
| L. brevis ATCC 367 and L. plantarum ATCC 21028 | Corn stover | 24 (l-Lactic acid) 31.2 (l-Lactic acid) | Simultaneous sequential | [167] |
| L. casei | Sugarcane molasses | 120.23 (NS) | Batch | [168] |
| Mutant S. cerevisiae | Glucose | 142 (l-Lactic acid) | Fed-batch | [169] |
| R. oryzae LA-UN-1 | Glucose | 162 (l-Lactic acid) | Fed-batch | [170] |
| L. rhamnosus DUT1908 | Starchy biomass | 108 (l-Lactic acid) | One step liquefaction, saccharification and fermentation | [171] |
| Mutated and evolved L. paracasei NCBIO01 | Glucose | 202 (l-Lactic acid) 9.06 (d-Lactic acid) | Open fermentation | [172] |
| B. coagulans and L. rhamnosus | Cassava bagasse | 113 (l-Lactic acid) | Simultaneous saccharification and co-fermentation | [173] |
| Mutant L. plantarum NCIMB 8826 | Raw corn starch | 50 (l-Lactic acid) | Batch | [174] |
| L. paracasei subsp. paracasei2 | Food waste | 34 (l-Lactic acid) | Batch | [175] |
NS: not specified.
2.5. Succinic Acid (C4H6O4, 118.09 g·mol−1, pKa1 4.2, pKa2 5.6)
Succinic acid (SA) is a dicarboxylic acid usually produced by hydrogenation of fossil-derived maleic anhydride, but due to the drawbacks associated with petroleum-chemical production (e.g., greenhouse gas emission and non-renewable energy consumption), nowadays the fermentative route is preferred for its production. Several studies have shown that the production of succinic acid by fermentation can also lead to a reduction in costs [176].
It is defined as “bio-succinic” acid, derived from plants, animals and microorganisms, although it is principally produced by anaerobic fermentation. In 1546 the German mineralist and doctor Georgius Agricola purified succinic acid for the first time from amber; afterwards, it was produced by microbial fermentation [177].
The first microorganisms recognized as succinic acid producers were bacteria isolated from rumen, such as Actinobacillus succinogenes, Anaerobiospirillum succiniciproducens, Mannheimia succiniciproducens, Basfia succiniciproducens, Bacillus fragilis and also some fungi such as Fusarium, Aspergillus and Penicillium species.
Among them, the most promising for industrial production is A. succinogenes; several fermentation aspects including carbon and nitrogen sources, as well as pH regulator, should be considered for a successful production process [178].
Unfortunately, the production of succinic acid by microbial fermentation is not competitive compared to petro-chemical products, as often happens for different organic acid production. The cost of the product per kg is USD 2.94 and USD 2.5, for bio-succinic acid and the combined bio- and fossil-based succinic acid, respectively [179]. Until now, the pivotal companies which produce succinic acid, according to the biorefinery concept, at commercial scale are Myriant (Quincy, MA, USA), Reverdia (Cassano Spinola, Italy) and Succinity (Montmelò, Spain) [180]. Succinic acid represents the key precursor for a wide range of bulk chemicals including 1,4-butanediol, gamma-butyrolactone, tetrahydrofuran, adipic acid, n-methylpyrrolidone, linear aliphatic esters, or biodegradable polymers (butylene succinic acid) (PBS) [181,182].
In most cases, succinic acid is produced by exploiting the reductive TCA cycle (Scheme 2) through anaerobic or micro-aerobic fermentation, principally by rumen bacteria [183]. In addition, there are two other pathways, the glyoxylate pathway and the oxidative TCA cycle, for succinate production [184].
In the last few years, several engineering strategies have been developed for the production of succinic acid by microbial fermentation. For this purpose, microorganisms such as S. cerevisiae, Y. lipolytica, E. coli, P. kudriavsevii, B. succiniciproducens have been considered, especially for their tolerance to low pH, essential to exceed the downstream costs, and for the many engineering tools available [176] (Table 6). The joint venture of DSM and Roquette, Reverdia, developed the modified strain of S. cerevisiae, SUC-297, for succinic acid production through the dual-phase fed-batch fermentation of glucose [185]. Likewise, other companies have worked to obtain mutated strains to increase their plant capacity, as reported by Ahn et al. [186].
At the same time, as for the other commodities before, low-cost feedstocks must be considered to obtain a sustainable bio-product. A two-stage pH fermentation, to promote the undissociated form of the acid, was proposed by Stylianou et al. [187] using as carbon sources the organic fraction of municipal solid waste (OFMSW) hydrolysates. In addition, other second-generation biomasses (e.g., mixed food, agricultural and textile waste as well as crude glycerol) have been successfully used as described by Li et al. [176].
The yield and efficiency of “bio-succinic” acid depends on the metabolic pathway involved. For example, the production of succinic acid by the reductive TCA cycle in anaerobic conditions can lead to defective cellular growth; Vemuri et al. proposed a dual-phase fermentation, which consists of an aerobic growth phase and an anaerobic production phase, combining glyoxylate and reductive TCA cycle, obtaining 99.2 g/L of succinic acid [188]. More recently, a mixed culture of two microorganisms, S. cerevisiae and A. succinogenes, was also exploited to create an efficient co-culture system for succinic acid production [189].
Table 6.
Succinic acid production with different substrates, microorganisms, and fermentation strategies.
Table 6.
Succinic acid production with different substrates, microorganisms, and fermentation strategies.
| Microorganism | Substrate | Succinic Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| Isolate AKR177 | Pure glycerol | 117 | Fed-batch | [190] |
| Isolate AKR177 | Crude glycerol | 86.9 | Fed-batch | [190] |
| A. succinogenes CCTCC M2012036 | Sugarcane bagasse | 39.9 70.81 | Batch Fed-batch | [191] |
| Mutant E. coli MH28 | Glycerol | 84.3 | Batch | [98] |
| Mutant S. cerevisiae SUC-297 | Glucose | 43 | Fed-batch | [185] |
| Mutant B. succiniciproducens LU15224 | Glycerol and maltose | 69.8 | Batch | [192] |
| Mutant Y. lipolytica | Crude Glycerol | 209.7 | Fed-batch | [193] |
| Mutant E. coli | Glucose | 99.2 | Dual-phase | [188] |
| Mutant Y. lipolytica | Glucose | 101.4 | Fed-batch | [194] |
| Mutant Y. lipolytica | Glycerol | 110.7 | Fed-batch | [195] |
| Evolved Y. lipolytica | Mixed food waste | 71.6 | Fed-batch | [196] |
3. Discussion and Conclusions
The examples reported are the best way of demonstrating that microbial-based production can at least compete, if still not replace, the fossil-based production of chemical platforms. Nonetheless, it was mentioned that many other commodity chemical platforms have been produced by microbial fermentation but have not reached the market yet.
Robustness of cell factories and scaling-up gaps are the prominent limitations. In respect to ameliorating the performances of cell factories, several aspects, often interconnected, need to be taken into account. The stoichiometry and thermodynamics of the native or heterologous pathways have to be balanced and considered, respectively, especially when the product of interest is implied to close or reroute other pathways as well. This also results in the possibility of calculating the theoretical yield and compensate for redox imbalance or availability of (micro)nutrients, to avoid by-product formation or/and energy overflow. Another relevant aspect is to evaluate the formation of oxidized products or free radicals, as a consequence of an energy imbalance or the fermentation settings [197].
In respect to the issue of scaling up, a promising support to fill the gap is offered by innovation in the field of scaling-down models, supported by the miniaturization of fermentation units, still fully equipped for online measurements. The very crucial point is to ask the system how to better simulate what happens in large-scale fermenters and how to prioritize the selected parameters, which can be decided only by sharing data and information and by having the whole process, including the final downstream, in mind [8]. In this respect, systems and synthetic biology can give a tremendous boost to our descriptive, predictive, and engineering capability, for example by including fluctuating cultivation conditions in quantitative descriptions and implementing cells with biosensors or molecular switchers tailored to compensate for stresses. These perspectives are particularly relevant if we consider, as recently reviewed, that the cost of bringing a process of commodity production to industrial scale can cost from USD 100 million to 1 billion [198].
As a final consideration, while it can be securely assessed that all the mentioned products are biobased, exactly as for biofuel production, the next question will be if they are really sustainable, as for most of them the feedstock is constituted by first-generation sugars. In order to exploit renewable substrates for the microbial production of organic acid, it is important to consider the difficulties related to the assimilation and metabolization by the cells. Recently, the genetic manipulation of membrane transporters involved in the import of second-generation biomasses has been summarized [199]. Nevertheless, there are different examples of improvements in the use of this kind of feedstock for organic acid production [200]. In parallel, great attention is more and more devoted to the utilization of CO2 as a carbon source, also for organic acid production [201,202], and not only in dependence on photosynthesis.
To conclude, thanks to experience in microbial biotechnology that has matured over the last decades, it is now very clear that obtaining the proof of concept for a process of production is no longer the limiting step. The main bottlenecks towards the development of profitable and competitive microbial and biobased processes are the scale-up parameters, which are very often difficult to replicate at laboratory-scale. The recent advances in technologies and in quantitative analysis, together with the tremendous advances in synthetic biology, will be crucial to boost research as well as the technology transfer of microbial organic acid production in the next few years.
Author Contributions
R.D.D.L. and I.S. Writing—Original Draft Preparation & Editing; P.B. and D.P. Conceptualization, Writing—Review & Editing. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge the support of FAR (Fondo di Ateneo per la Ricerca) of the University of Milano-Bicocca. The work was also partially supported by the SYSBIO Centre of Systems Biology. Studies on lactic acid production from agricultural renewable feedstock, ongoing in PB’s group, are supported by CARDoon valorization by InteGrAted biorefiNery (CARDIGAN) project, funded by the Italian Ministry of Education, University and Research (PRIN, 2017KBTK93).
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- Branduardi, P. Closing the loop: The power of microbial biotransformations from traditional bioprocesses to biorefineries, and beyond. Microb. Biotechnol. 2021, 14, 68–73. [Google Scholar] [CrossRef] [PubMed]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; National Renewable Energy Lab: Golden, CO, USA, 2004; Volume 2.
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—The US Department of Energy’s “top 10” revisited. Green Chem. 2010, 12, 539–555. [Google Scholar] [CrossRef]
- Becker, J.; Lange, A.; Fabarius, J.; Wittmann, C. Top value platform chemicals: Bio-based production of organic acids. Curr. Opin. Biotechnol. 2015, 36, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Song, C.W.; Shin, J.H.; Lee, S.Y. Biorefineries for the production of top building block chemicals and their derivatives. Metab. Eng. 2015, 28, 223–239. [Google Scholar] [CrossRef] [PubMed]
- Research, G.V. Organic Acids Market Analysis, by Product Type (Formic Acid, Citric Acid, Others), by Source Type (Biomass, Chemical Synthesis, Others), by Application (Food & Beverages Industry, Pharmaceutical Industry, Others), Forecasts To 2028. Available online: https://www.reportsanddata.com/report-detail/organic-acids-market (accessed on 14 December 2021).
- Van Dien, S. From the first drop to the first truckload: Commercialization of microbial processes for renewable chemicals. Curr. Opin. Biotechnol. 2013, 24, 1061–1068. [Google Scholar] [CrossRef]
- Wehrs, M.; Tanjore, D.; Eng, T.; Lievense, J.; Pray, T.R.; Mukhopadhyay, A. Engineering robust production microbes for large-scale cultivation. Trends Microbiol. 2019, 27, 524–537. [Google Scholar] [CrossRef] [Green Version]
- Scheele, C.W. Sammtliche Physische and Chemischewerke; Mayer & Müller: Berlin, Germany, 1784; Volume 2. [Google Scholar]
- Papagianni, M. Advances in citric acid fermentation by Aspergillus niger: Biochemical aspects, membrane transport and modeling. Biotechnol. Adv. 2007, 25, 244–263. [Google Scholar] [CrossRef]
- Grimoux, E.; Adams, P. Synthese de lácide citrique. C. R. Hebd. Seances Acad. Sci. 1880, 90, 1252. [Google Scholar]
- Wehmer, C.F.W. Beiträge zur Kenntnis Einheimischer Pilze; Hahn’sche Buchhandlung: Hannover/Jena, Germany, 1893. [Google Scholar]
- Currie, J.N. The citric acid fermentation of Aspergillus niger. J. Biol. Chem. 1917, 31, 15–37. [Google Scholar] [CrossRef]
- Apelblat, A. Citric Acid; Springer International Publishing: Cham, Switzerland, 2014; ISBN 978-3-319-11232-9. [Google Scholar]
- Judah, G. Pfizer’s Work on Penicillin for World War II becomes a National Historic Chemical Landmark. Available online: https://0-www-acs-org.brum.beds.ac.uk/content/acs/en/pressroom/newsreleases/2008/june/pfizers-work-on-penicillin-for-world-war-ii-becomes-a-national-historic-chemical-landmark.html (accessed on 13 December 2021).
- Mores, S.; de Souza Vandenberghe, L.P.; Júnior AI, M.; de Carvalho, J.C.; de Mello AF, M.; Pandey, A.; Soccol, C.R. Citric acid bioproduction and downstream processing: Status, opportunities, and challenges. Bioresour. Technol. 2021, 320, 124426. [Google Scholar] [CrossRef]
- Ciriminna, R.; Meneguzzo, F.; Delisi, R.; Pagliaro, M. Citric acid: Emerging applications of key biotechnology industrial product. Chem. Cent. J. 2017, 11, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Savi, L.K.; Dias, M.C.G.C.; Carpine, D.; Waszczynskyj, N.; Ribani, R.H.; Haminiuk, C.W.I. Natural deep eutectic solvents (NADES) based on citric acid and sucrose as a potential green technology: A comprehensive study of water inclusion and its effect on thermal, physical and rheological properties. Int. J. Food Sci. Technol. 2019, 54, 898–907. [Google Scholar] [CrossRef]
- Amato, A.; Becci, A.; Beolchini, F. Citric acid bioproduction: The technological innovation change. Crit. Rev. Biotechnol. 2020, 40, 199–212. [Google Scholar] [CrossRef]
- Behera, B.C.; Mishra, R.; Mohapatra, S. Microbial citric acid: Production, properties, application, and future perspectives. Food Front. 2021, 2, 62–76. [Google Scholar] [CrossRef]
- Erdem, C.; Papanikolaou, S.; Fickers, P.; Agirman, B.; Erten, H. Citric Acid Production by Yarrowia lipolytica. In Non-Conventional Yeasts: From Basic Research to Application; Sibirny, A., Ed.; Springer International Publishing: Berlin/Heidelberg, Germany, 2019; pp. 91–117. [Google Scholar]
- Yang, L.; Lübeck, M.; Lübeck, P.S. Aspergillus as a versatile cell factory for organic acid production. Fungal Biol. Rev. 2017, 31, 33–49. [Google Scholar] [CrossRef]
- Perlman, D. Mycological production of citric acid: The submerged culture method. Econ. Bot. 1949, 3, 360. [Google Scholar] [CrossRef]
- Gil, R.G.; Correa, H.S.; Contreras Larios, J.L.; González-Brambila, M.M. A biotechnological process for obtaining citric acid through paper cellulose aerobic bioreaction. Int. J. Chem. React. Eng. 2020, 18, 1–10. [Google Scholar] [CrossRef]
- Roukas, T.; Kotzekidou, P. Pomegranate peel waste: A new substrate for citric acid production by Aspergillus niger in solid-state fermentation under non-aseptic conditions. Environ. Sci. Pollut. Res. 2020, 27, 13105–13113. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, E.; Kontogiannopoulos, K.N.; Assimopoulou, A.N.; Mantzouridou, F.T. Feasibility of multi-hydrolytic enzymes production from optimized grape pomace residues and wheat bran mixture using Aspergillus niger in an integrated citric acid-enzymes production process. Bioresour. Technol. 2020, 309, 123317. [Google Scholar] [CrossRef]
- Papadaki, E.; Mantzouridou, F.T. Citric acid production from the integration of spanish-style green olive processing wastewaters with white grape pomace by Aspergillus niger. Bioresour. Technol. 2019, 280, 59–69. [Google Scholar] [CrossRef]
- Ozdal, M.; Kurbanoglu, E.B. Citric acid production by Aspergillus niger from agro-industrial by-products: Molasses and chicken feather peptone. Waste Biomass Valorization 2019, 10, 631–640. [Google Scholar] [CrossRef]
- Lotfy, W.A.; Ghanem, K.M.; El-Helow, E.R. Citric acid production by a novel Aspergillus niger isolate: I. Mutagenesis and cost reduction studies. Bioresour. Technol. 2007, 98, 3464–3469. [Google Scholar] [CrossRef] [PubMed]
- Adeoye, A.O.; Lateef, A.; Gueguim-Kana, E.B. Optimization of citric acid production using a mutant strain of Aspergillus niger on cassava peel substrate. Biocatal. Agric. Biotechnol. 2015, 4, 568–574. [Google Scholar] [CrossRef]
- Pel, H.J.; De Winde, J.H.; Archer, D.B.; Dyer, P.S.; Hofmann, G.; Schaap, P.J.; Turner, G.; De Vries, R.P.; Albang, R.; Albermann, K.; et al. Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat. Biotechnol. 2007, 25, 221–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Cao, Z.; Hou, L.; Yin, L.; Wang, D.; Gao, Q.; Wu, Z.; Wang, D. The opposite roles of agdA and glaA on citric acid production in Aspergillus niger. Appl. Microbiol. Biotechnol. 2016, 100, 5791–5803. [Google Scholar] [CrossRef]
- Xue, X.; Bi, F.; Liu, B.; Li, J.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Improving citric acid production of an industrial Aspergillus niger CGMCC 10142: Identification and overexpression of a high-affinity glucose transporter with different promoters. Microb. Cell Fact. 2021, 20, 1–13. [Google Scholar] [CrossRef]
- Steiger, M.G.; Rassinger, A.; Mattanovich, D.; Sauer, M. Engineering of the citrate exporter protein enables high citric acid production in Aspergillus niger. Metab. Eng. 2019, 52, 224–231. [Google Scholar] [CrossRef]
- Sun, X.; Wu, H.; Zhao, G.; Li, Z.; Wu, X.; Liu, H.; Zheng, Z. Morphological regulation of Aspergillus niger to improve citric acid production by chsC gene silencing. Bioprocess Biosyst. Eng. 2018, 41, 1029–1038. [Google Scholar] [CrossRef]
- Hou, L.; Liu, L.; Zhang, H.; Zhang, L.; Zhang, L.; Zhang, J.; Gao, Q.; Wang, D. Functional analysis of the mitochondrial alternative oxidase gene (aox1) from Aspergillus niger CGMCC 10142 and its effects on citric acid production. Appl. Microbiol. Biotechnol. 2018, 102, 7981–7995. [Google Scholar] [CrossRef] [PubMed]
- De Jongh, W.A.; Nielsen, J. Enhanced citrate production through gene insertion in Aspergillus niger. Metab. Eng. 2008, 10, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.; Brooks, J.D.; Maddox, I.S. The effect of the sugar source on citric acid production by Aspergillus niger. Appl. Microbiol. Biotechnol. 1984, 19, 393–397. [Google Scholar] [CrossRef]
- Hou, W.; Bao, J. Simultaneous saccharification and aerobic fermentation of high titer cellulosic citric acid by filamentous fungus Aspergillus niger. Bioresour. Technol. 2018, 253, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, J.; Cao, Z.; Wang, Y.; Gao, Q.; Zhang, J.; Wang, D. Inhibition of oxidative phosphorylation for enhancing citric acid production by Aspergillus niger. Microb. Cell Fact. 2015, 14, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.; Zhao, S.; Tan, F.; Li, H.; Chu, R.; Wang, X.; Sun, H.; Zhang, M. A novel green production process of citric acid on the pilot scale by directly recycling its extraction effluent. J. Clean. Prod. 2020, 277, 124068. [Google Scholar] [CrossRef]
- Yu, B.; Zhang, X.; Sun, W.; Xi, X.; Zhao, N.; Huang, Z.; Ying, Z.; Liu, L.; Liu, D.; Niu, H.; et al. Continuous citric acid production in repeated-fed batch fermentation by Aspergillus niger immobilized on a new porous foam. J. Biotechnol. 2018, 276–277, 1–9. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Characterization of Yarrowia lipolytica and related species for citric acid production from glycerol. Enzyme Microb. Technol. 2007, 41, 292–295. [Google Scholar] [CrossRef]
- Vandenberghe, L.P.S.; Rodrigues, C.; de Carvalho, J.C.; Medeiros, A.B.P.; Soccol, C.R. Production and application of citric acid. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 557–575. ISBN 9780444636621. [Google Scholar]
- Vandenberghe, L.P.S.; Soccol, C.R.; Prado, F.C.; Pandey, A. Comparison of citric acid production by solid-state fermentation in flask, column, tray, and drum bioreactors. Appl. Biochem. Biotechnol. 2004, 118, 293–304. [Google Scholar] [CrossRef]
- Straathof, A.J.J. The proportion of downstream costs in fermentative production processes. In Comprehensive Biotechnology; Elsevier: Amsterdam, The Netherlands, 2011; Volume 1, pp. 811–814. [Google Scholar]
- Kertes, A.S.; King, C.J. Extraction chemistry of fermentation product carboxylic acids. Biotechnol. Bioeng. 2009, 103, 431–445. [Google Scholar] [CrossRef] [PubMed]
- Araújo, E.M.R.; Coelho, F.E.B.; Balarini, J.C.; Miranda, T.L.S.; Salum, A. Solvent extraction of citric acid with different organic phases. Adv. Chem. Eng. Sci. 2017, 7, 304–324. [Google Scholar] [CrossRef] [Green Version]
- Djas, M.; Henczka, M. Reactive extraction of citric acid using supercritical carbon dioxide. J. Supercrit. Fluids 2016, 117, 59–63. [Google Scholar] [CrossRef]
- Pourcelly, G. Citric acid recovery by electrodialysis. In Encyclopedia of Membranes; Drioli, E., Giorno, L., Eds.; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Delgado Dobladez, J.A.; Águeda Maté, V.I.; Uribe Santos, D.L.; Torrellas, S.Á.; Larriba, M. Citric acid purification by simulated moving bed adsorption with methanol as desorbent. Sep. Sci. Technol. 2019, 54, 930–942. [Google Scholar] [CrossRef]
- Wang, J.; Cui, Z.; Li, Y.; Cao, L.; Lu, Z. Techno-economic analysis and environmental impact assessment of citric acid production through different recovery methods. J. Clean. Prod. 2020, 249, 119315. [Google Scholar] [CrossRef]
- Anastassiadis, S.; Morgunov, I.G.; Kamzolova, S.V.; Finogenova, T.V. Citric acid production patent review. Recent Pat. Biotechnol. 2008, 2, 107–123. [Google Scholar] [CrossRef] [PubMed]
- Citric Acid Market: Global Industry Trends, Share, Size, Growth, Opportunity and Forecast 2021–2026. Available online: https://www.imarcgroup.com/citric-acid-manufacturing-plant (accessed on 29 January 2022).
- Ilica, R.A.; Kloetzer, L.; Galaction, A.I.; Caşcaval, D. Fumaric acid: Production and separation. Biotechnol. Lett. 2019, 41, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Liu, N.; Cao, Y.; Jin, C.; Li, F.; Cai, C.; Yao, J. Effects of fumaric acid supplementation on methane production and rumen fermentation in goats fed diets varying in forage and concentrate particle size. J. Anim. Sci. Biotechnol. 2018, 9, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reda, F.M.; Ismail, I.E.; Attia, A.I.; Fikry, A.M.; Khalifa, E.; Alagawany, M. Use of fumaric acid as a feed additive in quail’s nutrition: Its effect on growth rate, carcass, nutrient digestibility, digestive enzymes, blood metabolites, and intestinal microbiota. Poult. Sci. 2021, 100, 101493. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Straathof, A.J.J.; Zijlmans, T.W.; Van Gulik, W.M.; Van Der Wielen, L.A.M. Fumaric acid production by fermentation. Appl. Microbiol. Biotechnol. 2008, 78, 379–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moharregh-Khiabani, D.; Linker, R.A.; Gold, R.; Stangel, M. Fumaric acid and its esters: An emerging treatment for multiple sclerosis. Curr. Neuropharmacol. 2009, 7, 60–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D. Fumaric acid esters for psoriasis: A systematic review. Ir. J. Med. Sci. 2017, 186, 161–177. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, S.; Iino, T.; Sato, S.; Nakahara, T.; Mukataka, S. Improvement of production rate and yield of fumaric acid from maleic acid by heat treatment of Pseudomonas alcaligenes strain XD-1. Biochem. Eng. J. 2003, 13, 7–13. [Google Scholar] [CrossRef]
- Goto, M.; Nara, T.; Tokumaru, I.; Fugono, N.; Uchida, Y.; Terasawa, M.; Yukawa, H. Method of Producing Fumaric Acid. U.S. Patent US5783428, 21 July 1998. [Google Scholar]
- Abe, A.; Oda, Y.; Asano, K.; Sone, T. Rhizopus delemar is the proper name for Rhizopus oryzae fumaric-malic acid producers. Mycologia 2007, 99, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Kenealy, W.; Zaady, E.; Du Preez, J.C. Biochemical aspects of fumaric acid accumulation by Rhizopus arrhizus. Appl. Environ. Microbiol. 1986, 52, 128–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, S.T.; Zhang, K.; Zhang, B.; Huang, H. Fumaric Acid. Compr. Biotechnol. Second Ed. 2011, 3, 163–177. [Google Scholar] [CrossRef]
- Zhang, B.; Skory, C.D.; Yang, S.T. Metabolic engineering of Rhizopus oryzae: Effects of overexpressing pyc and pepc genes on fumaric acid biosynthesis from glucose. Metab. Eng. 2012, 14, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Yang, S.T. Metabolic engineering of Rhizopus oryzae: Effects of overexpressing fumR gene on cell growth and fumaric acid biosynthesis from glucose. Process Biochem. 2012, 47, 2159–2165. [Google Scholar] [CrossRef]
- Song, C.W.; Kim, D.I.; Choi, S.; Jang, J.W.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of fumaric acid. Biotechnol. Bioeng. 2013, 110, 2025–2034. [Google Scholar] [CrossRef]
- Li, N.; Zhang, B.; Wang, Z.; Tang, Y.J.; Chen, T.; Zhao, X. Engineering Escherichia coli for fumaric acid production from glycerol. Bioresour. Technol. 2014, 174, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Chen, X.; Liu, L.; Jiang, L. Fumaric acid production in Saccharomyces cerevisiae by simultaneous use of oxidative and reductive routes. Bioresour. Technol. 2013, 148, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhu, P.; Liu, L. Modular optimization of multi-gene pathways for fumarate production. Metab. Eng. 2016, 33, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wu, J.; Song, W.; Zhang, L.; Wang, H.; Liu, L. Fumaric acid production by Torulopsis glabrata: Engineering the urea cycle and the purine nucleotide cycle. Biotechnol. Bioeng. 2015, 112, 156–167. [Google Scholar] [CrossRef]
- Sebastian, J.; Hegde, K.; Kumar, P.; Rouissi, T.; Brar, S.K. Bioproduction of fumaric acid: An insight into microbial strain improvement strategies. Crit. Rev. Biotechnol. 2019, 39, 817–834. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.B.; Ng, T.K. Fermentation Process for Carboxylic Acids. U.S. Patent US4877731A, 31 October 1989. [Google Scholar]
- Sebastian, J.; Rouissi, T.; Brar, S.K. Miscanthus sp. Perennial lignocellulosic biomass as feedstock for greener fumaric acid bioproduction. Ind. Crops Prod. 2022, 175, 114248. [Google Scholar] [CrossRef]
- Li, X.; Zhou, J.; Ouyang, S.; Ouyang, J.; Yong, Q. Fumaric acid production from alkali-pretreated corncob by fed-batch simultaneous saccharification and fermentation combined with separated hydrolysis and fermentation at high solids loading. Appl. Biochem. Biotechnol. 2017, 181, 573–583. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K.; Verma, M. A fermentative approach towards optimizing directed biosynthesis of fumaric acid by Rhizopus oryzae 1526 utilizing apple industry waste biomass. Fungal Biol. 2015, 119, 1279–1290. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Ma, J.; Wang, M.; Wang, W.; Deng, L.; Nie, K.; Yue, X.; Wang, F.; Tan, T. Food waste fermentation to fumaric acid by Rhizopus arrhizus rh7-13. Appl. Biochem. Biotechnol. 2016, 180, 1524–1533. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Li, S.; Xu, Q.; Gao, M.; Huang, H. Production of fumaric acid by simultaneous saccharification and fermentation of starchy materials with 2-deoxyglucose-resistant mutant strains of Rhizopus oryzae. Bioresour. Technol. 2012, 107, 363–367. [Google Scholar] [CrossRef]
- Wu, X.; Liu, Q.; Deng, Y.; Chen, X.; Zheng, Z.; Jiang, S.; Li, X. Production of fumaric acid by bioconversion of corncob hydrolytes using an improved Rhizopus oryzae strain. Appl. Biochem. Biotechnol. 2018, 184, 553–569. [Google Scholar] [CrossRef]
- Moresi, M.; Parente, E.; Petruccioli, M.; Federici, F. Fumaric acid production from hydrolysates of starch-based substrates. J. Chem. Technol. Biotechnol. 1992, 54, 283–290. [Google Scholar] [CrossRef]
- Deng, F.; Aita, G.M. fumaric acid production by Rhizopus oryzae ATCC®® 20344tm from lignocellulosic syrup. Bioenergy Res. 2018, 11, 330–340. [Google Scholar] [CrossRef]
- Scholz, S.A.; Graves, I.; Minty, J.J.; Lin, X.N. Production of cellulosic organic acids via synthetic fungal consortia. Biotechnol. Bioeng. 2017, 115, 1096–1100. [Google Scholar] [CrossRef]
- Zhou, Y.; Nie, K.; Zhang, X.; Liu, S.; Wang, M.; Deng, L.; Wang, F.; Tan, T. Production of fumaric acid from biodiesel-derived crude glycerol by Rhizopus arrhizus. Bioresour. Technol. 2014, 163, 48–53. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K. Enhanced fumaric acid production from brewery wastewater and insight into the morphology of Rhizopus oryzae 1526. Appl. Biochem. Biotechnol. 2014, 172, 2974–2988. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Du, G.; Hua, Z.; Zhou, J.; Chen, J. Optimization of fumaric acid production by Rhizopus delemar based on the morphology formation. Bioresour. Technol. 2011, 102, 9345–9349. [Google Scholar] [CrossRef] [PubMed]
- Papadaki, A.; Androutsopoulos, N.; Patsalou, M.; Koutinas, M.; Kopsahelis, N.; Machado De Castro, A.; Papanikolaou, S.; Koutinas, A.A. Biotechnological production of fumaric acid: The effect of morphology of Rhizopus arrhizus NRRL 2582. Fermentation 2017, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Yu, C.; Yang, S.T. Effects of soybean meal hydrolysate as the nitrogen source on seed culture morphology and fumaric acid production by Rhizopus oryzae. Process Biochem. 2015, 50, 173–179. [Google Scholar] [CrossRef]
- Zhou, Y.; Du, J.; Tsao, G.T. Mycelial pellet formation by Rhizopus oryzae ATCC 20344. Appl. Biochem. Biotechnol. 2000, 84, 779–789. [Google Scholar] [CrossRef]
- Roa Engel, C.A.; Van Gulik, W.M.; Marang, L.; Van der Wielen, L.A.M.; Straathof, A.J.J. Development of a low pH fermentation strategy for fumaric acid production by Rhizopus oryzae. Enzyme Microb. Technol. 2011, 48, 39–47. [Google Scholar] [CrossRef]
- Das, R.K.; Brar, S.K.; Verma, M. effects of different metallic nanoparticles on germination and morphology of the fungus Rhizopus oryzae 1526 and changes in the production of fumaric acid. Bionanoscience 2015, 5, 217–226. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, S.; Jin, Y.; Yue, X.; Deng, L.; Wang, F.; Tan, T. Production of fumaric acid by immobilized Rhizopus arrhizus RH 7-13-9# on loofah fiber in a stirred-tank reactor. Bioresour. Technol. 2017, 244, 929–933. [Google Scholar] [CrossRef]
- Naude, A.; Nicol, W. Improved continuous fumaric acid production with immobilised Rhizopus oryzae by implementation of a revised nitrogen control strategy. N. Biotechnol. 2018, 44, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, H.; Liu, X.; Zhu, H.; Fan, T.; Deng, L.; Wang, F. A high efficient method for simultaneous fermentation and separation of fumaric acid with a fixed bed ion exchange column. Biochem. Eng. J. 2020, 160, 107610. [Google Scholar] [CrossRef]
- Zhou, Y.; Du, J.; Tsao, G. Comparison of fumaric acid production by Rhizopus oryzae using different neutralizing agents. Bioprocess Biosyst. Eng. 2002, 25, 179–181. [Google Scholar] [CrossRef]
- Rhodes, R.A.; Lagoda, A.A.; Misenheimer, T.J.; Smith, M.L.; Anderson, R.F.; Jackson, R.W. production of fumaric acid in 20-liter fermentors. Appl. Microbiol. 1962, 10, 9–15. [Google Scholar] [CrossRef]
- Guettler, M.V.; Hanchar, R.J.; Rumler, D.S.; Kleff, S. Fermentation Method using a Magnesium Compound Containing Oxygen. U.S. Patent 8,431,372, 30 April 2010. [Google Scholar]
- Yocum, R.R.; Hermann, T.; Yu, X. Fermentation of Glycerol to Organic Acids 2013. WO2013/015770Al, 31 January 2013. [Google Scholar]
- Yocum, R.R.; Dole, S.; Pero, J.G. Production of Organic Acids by Fermentation at Low pH. U.S. Patent US20150240270A1, 28 September 2015. [Google Scholar]
- Cordes, T.; Michelucci, A.; Hiller, K. Itaconic Acid: The surprising role of an industrial compound as a mammalian antimicrobial metabolite. Annu. Rev. Nutr. 2015, 35, 451–473. [Google Scholar] [CrossRef]
- Gopaliya, D.; Kumar, V.; Khare, S.K. Recent advances in itaconic acid production from microbial cell factories. Biocatal. Agric. Biotechnol. 2021, 36, 102130. [Google Scholar] [CrossRef]
- Teleky, B.E.; Vodnar, D.C. Biomass-derived production of itaconic acid as a building block in specialty polymers. Polymers 2019, 11, 1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha da Cruz, J.; Machado de Castro, A.; Camporese Sérvulo, E.F. World market and biotechnological production of itaconic acid. 3 Biotech. 2018, 8. [Google Scholar] [CrossRef]
- Bafana, R.; Pandey, R.A. New approaches for itaconic acid production: Bottlenecks and possible remedies. Crit. Rev. Biotechnol. 2018, 38, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Weastra Determination of market potential for selected platform chemicals: Itaconic acid, Succinic acid, 2,5-Furandicarboxylic acid. BioConSepT 2011, 1–173.
- Okabe, M.; Lies, D.; Kanamasa, S.; Park, E.Y. Biotechnological production of itaconic acid and its biosynthesis in Aspergillus terreus. Appl. Microbiol. Biotechnol. 2009, 84, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Krull, S.; Hevekerl, A.; Kuenz, A.; Prüße, U. Process development of itaconic acid production by a natural wild type strain of Aspergillus terreus to reach industrially relevant final titers. Appl. Microbiol. Biotechnol. 2017, 101, 4063–4072. [Google Scholar] [CrossRef]
- Saha, B.C. Emerging biotechnologies for production of itaconic acid and its applications as a platform chemical. J. Ind. Microbiol. Biotechnol. 2017, 44, 303–315. [Google Scholar] [CrossRef]
- Maassen, N.; Panakova, M.; Wierckx, N.; Geiser, E.; Zimmermann, M.; Bölker, M.; Klinner, U.; Blank, L.M. Influence of carbon and nitrogen concentration on itaconic acid production by the smut fungus Ustilago maydis. Eng. Life Sci. 2014, 14, 129–134. [Google Scholar] [CrossRef]
- Kuenz, A.; Gallenmüller, Y.; Willke, T.; Vorlop, K.D. Microbial production of itaconic acid: Developing a stable platform for high product concentrations. Appl. Microbiol. Biotechnol. 2012, 96, 1209–1216. [Google Scholar] [CrossRef]
- Wierckx, N.; Agrimi, G.; Lübeck, P.S.; Steiger, M.G.; Mira, N.P.; Punt, P.J. Metabolic specialization in itaconic acid production: A tale of two fungi. Curr. Opin. Biotechnol. 2020, 62, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Tabuchi, T.; Sugisawa, T.; Ishidori, T.; Nakahara, T.; Sugiyama, J. Itaconic acid fermentation by a yeast belonging to the genus Candida. Agric. Biol. Chem. 1981, 45, 475–479. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Production of itaconic acid by Pseudozyma antarctica NRRL Y-7808 under nitrogen-limited growth conditions. Enzyme Microb. Technol. 2006, 39, 824–827. [Google Scholar] [CrossRef]
- Winskillm, N. Tricarboxylic acid cycle activity in relation to itaconic acid biosynthesis by Aspergillus terreus. Microbiology 1983, 129, 2877–2883. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Lu, X.; Zong, H.; Li, J.; Zhuge, B. Itaconic acid production in microorganisms. Biotechnol. Lett. 2018, 40, 455–464. [Google Scholar] [CrossRef]
- Geiser, E.; Przybilla, S.K.; Friedrich, A.; Buckel, W.; Wierckx, N.; Blank, L.M.; Bölker, M. Ustilago maydis produces itaconic acid via the unusual intermediate trans-aconitate. Microb. Biotechnol. 2016, 9, 116–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, J.; Tehrani, H.H.; Ernst, P.; Blank, L.M.; Wierckx, N. An optimized Ustilago maydis for itaconic acid production at maximal theoretical yield. J. Fungi 2021, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Vila-Santa, A.; Liu, N.; Prozorov, T.; Xie, D.; Faria, N.T.; Ferreira, F.C.; Mira, N.P.; Shao, Z. Metabolic engineering of an acid-tolerant yeast strain Pichia kudriavzevii for itaconic acid production. Metab. Eng. Commun. 2020, 10, e00124. [Google Scholar] [CrossRef]
- Kolláth, I.S.; Molnár, Á.P.; Soós, Á.; Fekete, E.; Sándor, E.; Kovács, B.; Kubicek, C.P.; Karaffa, L. Manganese deficiency is required for high itaconic acid production from D-xylose in Aspergillus terreus. Front. Microbiol. 2019, 10, 1589. [Google Scholar] [CrossRef] [Green Version]
- Kerssemakers, A.A.J.; Doménech, P.; Cassano, M.; Yamakawa, C.K.; Dragone, G.; Mussatto, S.I. Production of itaconic acid from cellulose pulp: Feedstock feasibility and process strategies for an efficient microbial performance. Energies 2020, 13, 1654. [Google Scholar] [CrossRef] [Green Version]
- Demir, H.T.; Bezirci, E.; Becker, J.; Tehrani, H.H.; Nikerel, E.; Wierck, N.; Türker, M. High level production of itaconic acid at low pH by Ustilago maydis with fed-batch fermentation. Bioprocess Biosyst. Eng. 2021, 44, 749–758. [Google Scholar] [CrossRef]
- Zambanini, T.; Hosseinpour Tehrani, H.; Geiser, E.; Merker, D.; Schleese, S.; Krabbe, J.; Buescher, J.M.; Meurer, G.; Wierckx, N.; Blank, L.M. Efficient itaconic acid production from glycerol with Ustilago vetiveriae TZ1. Biotechnol. Biofuels 2017, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, Q.; Deng, Y.; Li, J.; Chen, X.; Gu, Y.; Lv, X.; Zheng, Z.; Jiang, S.; Li, X. Production of itaconic acid by biotransformation of wheat bran hydrolysate with Aspergillus terreus CICC40205 mutant. Bioresour. Technol. 2017, 241, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, G.; Zhang, J.; Balan, V.; Bao, J. Itaconic acid fermentation using activated charcoal-treated corn stover hydrolysate and process evaluation based on Aspen plus model. Biomass Convers. Biorefinery 2020, 10, 463–470. [Google Scholar] [CrossRef]
- Xie, H.; Ma, Q.; Wei, D.; Wang, F. Metabolic engineering of an industrial Aspergillus niger strain for itaconic acid production. 3 Biotech 2020, 10, 113. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xu, H.; Jiang, J.; Zhang, N.; Xie, J.; Wei, M.; Zhao, J. Production of itaconic acid through microbiological fermentation of inexpensive materials. J. Bioresour. Bioprod. 2019, 4, 135–142. [Google Scholar] [CrossRef]
- Klement, T.; Büchs, J. Itaconic acid—A biotechnological process in change. Bioresour. Technol. 2013, 135, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Ghaffar, T.; Irshad, M.; Anwar, Z.; Aqil, T.; Zulifqar, Z.; Tariq, A.; Kamran, M.; Ehsan, N.; Mehmood, S. Recent trends in lactic acid biotechnology: A brief review on production to purification. J. Radiat. Res. Appl. Sci. 2014, 7, 222–229. [Google Scholar] [CrossRef]
- Komesu, A.; de Oliveira, J.A.R.; da Silva Martins, L.H.; Maciel, M.R.W.; Maciel Filho, R. Lactic acid production to purification: A review. BioResources 2017, 12, 4364–4383. [Google Scholar] [CrossRef]
- Meng, K.; Zhang, G.; Ding, C.; Zhang, T.; Yan, H.; Zhang, D.; Fang, T.; Liu, M.; You, Z.; Yang, C.; et al. Recent advances on purification of lactic acid. Chem. Rec. 2020, 20, 1236–1256. [Google Scholar] [CrossRef] [PubMed]
- Farooq, U.; Anjum, F.M.; Zahoor, T.; Sajjad-Ur-Rahman; Randhawa, M.A.; Ahmed, A.; Akram, K. Optimization of lactic acid production from cheap raw material: Sugarcane molasses. Pakistan J. Bot. 2012, 44, 333–338. [Google Scholar]
- Ajala, E.O.; Olonade, Y.O.; Ajala, M.A.; Akinpelu, G.S. Lactic acid production from lignocellulose–A review of major challenges and selected solutions. ChemBioEng Rev. 2020, 7, 38–49. [Google Scholar] [CrossRef]
- Rawoof, S.A.A.; Kumar, P.S.; Vo, D.V.N.; Devaraj, K.; Mani, Y.; Devaraj, T.; Subramanian, S. Production of optically pure lactic acid by microbial fermentation: A review. Environ. Chem. Lett. 2021, 19, 539–556. [Google Scholar] [CrossRef]
- Jem, K.J.; Tan, B. The development and challenges of poly (lactic acid) and poly (glycolic acid). Adv. Ind. Eng. Polym. Res. 2020, 3, 60–70. [Google Scholar] [CrossRef]
- Naser, A.Z.; Deiab, I.; Darras, B.M. Poly(lactic acid) (PLA) and polyhydroxyalkanoates (PHAs), green alternatives to petroleum-based plastics: A review. RSC Adv. 2021, 11, 17151–17196. [Google Scholar] [CrossRef]
- Chung, M.R.W.Y.; Tan, I.S.; Foo, H.C.Y.; Lam, M.K.; Lim, S. Potential of macroalgae-based biorefinery for lactic acid production from exergy aspect. Biomass Convers. Biorefinery 2021, 1–31. [Google Scholar] [CrossRef]
- Alexandri, M.; Neu, A.K.; Schneider, R.; López-Gómez, J.P.; Venus, J. Evaluation of various Bacillus coagulans isolates for the production of high purity L-lactic acid using defatted rice bran hydrolysates. Int. J. Food Sci. Technol. 2019, 54, 1321–1329. [Google Scholar] [CrossRef]
- Maas, R.H.W.; Bakker, R.R.; Jansen, M.L.A.; Visser, D.; De Jong, E.; Eggink, G.; Weusthuis, R.A. Lactic acid production from lime-treated wheat straw by Bacillus coagulans: Neutralization of acid by fed-batch addition of alkaline substrate. Appl. Microbiol. Biotechnol. 2008, 78, 751–758. [Google Scholar] [CrossRef] [Green Version]
- Awasthi, D.; Wang, L.; Rhee, M.S.; Wang, Q.; Chauliac, D.; Ingram, L.O.; Shanmugam, K.T. Metabolic engineering of Bacillus subtilis for production of D-lactic acid. Biotechnol. Bioeng. 2018, 115, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, J.; Aravindan, R.; Viruthagiri, T. Recent trends in the production, purification and application of lactic acid. Chem. Biochem. Eng. Q. 2008, 22, 245–264. [Google Scholar]
- Abedi, E.; Hashemi, S.M.B. Lactic acid production–Producing microorganisms and substrates sources-state of art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- Holyoak, C.D.; Stratford, M.; Mcmullin, Z.; Cole, M.B.; Crimmins, K.; Brown, A.J.P.; Coote, P.J. Activity of the plasma membrane H+-ATPase and optimal glycolytic flux are required for rapid adaptation and growth of Saccharomyces cerevisiae in the presence of the weak-acid preservative sorbic acid. Appl. Environ. Microbiol. 1996, 62, 3158–3164. [Google Scholar] [CrossRef] [Green Version]
- Porro, D.; Bianchi, M.M.; Brambilla, L.; Menghini, R.; Bolzani, D.; Carrera, V.; Lievense, J.; Liu, C.; Ranzi, B.M.; Frontali, L.; et al. replacement of a metabolic pathway for large-scale production of lactic acid from engineered yeasts. Appl. Environ. Microbiol. 1999, 65, 4211–4215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves De Oliveira, R.; Alexandri, M.; Komesu, A.; Venus, J.; Vaz Rossell, C.E.; Maciel Filho, R. Current advances in separation and purification of second-generation lactic acid. Sep. Purif. Rev. 2020, 49, 159–175. [Google Scholar] [CrossRef]
- Tian, X.; Chen, H.; Liu, H.; Chen, J. Recent advances in lactic acid production by lactic acid bacteria. Appl. Biochem. Biotechnol. 2021, 193, 4151–4171. [Google Scholar] [CrossRef] [PubMed]
- Pleissner, D.; Demichelis, F.; Mariano, S.; Fiore, S.; Navarro Gutiérrez, I.M.; Schneider, R.; Venus, J. Direct production of lactic acid based on simultaneous saccharification and fermentation of mixed restaurant food waste. J. Clean. Prod. 2017, 143, 615–623. [Google Scholar] [CrossRef]
- Zhou, X.; Ye, L.; Wu, J.C. Efficient production of l-lactic acid by newly isolated thermophilic Bacillus coagulans WCP10-4 with high glucose tolerance. Appl. Microbiol. Biotechnol. 2013, 97, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, P. Efficient production of l-lactic acid using co-feeding strategy based on cane molasses/glucose carbon sources. Bioresour. Technol. 2014, 153, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Porro, D.; Brambilla, L.; Ranzi, B.M.; Martegani, E.; Alberghina, L. Development of metabolically engineered Saccharomyces cerevisiae cells for the production of lactic acid. Biotechnol. Prog. 1995, 11, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Baptista, S.L.; Costa, C.E.; Cunha, J.T.; Soares, P.O.; Domingues, L. Metabolic engineering of Saccharomyces cerevisiae for the production of top value chemicals from biorefinery carbohydrates. Biotechnol. Adv. 2021, 47, 107697. [Google Scholar] [CrossRef]
- Sauer, M.; Porro, D.; Mattanovich, D.; Branduardi, P. 16 Years research on lactic acid production with yeast. Biotechnol. Genet. Eng. Rev. 2010, 21, 229–256. [Google Scholar] [CrossRef] [PubMed]
- Abbott, D.A.; Zelle, R.M.; Pronk, J.T.; van Maris, A.J.A. Metabolic engineering of Saccharomyces cerevisiae for production of carboxylic acids: Current status and challenges. FEMS Yeast Res. 2009, 9, 1123–1136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.J.; Bae, J.H.; Ko, H.J.; Lee, S.H.; Sung, B.H.; Han, J.I.; Sohn, J.H. Low-pH production of D-lactic acid using newly isolated acid tolerant yeast Pichia kudriavzevii NG7. Biotechnol. Bioeng. 2018, 115, 2232–2242. [Google Scholar] [CrossRef]
- Eş, I.; Mousavi Khaneghah, A.; Barba, F.J.; Saraiva, J.A.; Sant’Ana, A.S.; Hashemi, S.M.B. Recent advancements in lactic acid production—A review. Food Res. Int. 2018, 107, 763–770. [Google Scholar] [CrossRef]
- Grand View Research Lactic Acid Market Size Worth $5.02 Billion by 2028. Available online: https://www.grandviewresearch.com/press-release/global-lactic-acid-and-poly-lactic-acid-market (accessed on 14 December 2021).
- Srivastava, D.G. Lactic acid production for pharmaceutical applications by Lactobacillus delbrueckii immobilized on chitosan and polyethylene glycol stabilized calcium alginate beads: Process optimization using response surface methodology. World J. Pharm. Res. 2017, 6, 1737–1762. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Bhagwat, S.S.; Cortés-Penã, Y.R.; Ki, D.; Rao, C.V.; Jin, Y.S.; Guest, J.S. Sustainable lactic acid production from lignocellulosic biomass. ACS Sustain. Chem. Eng. 2021, 9, 1341–1351. [Google Scholar] [CrossRef]
- Wasewar, K.L. Separation of lactic acid: Recent advances. Chem. Biochem. Eng. Q. 2005, 19, 159–172. [Google Scholar]
- Li, C.; Gao, M.; Zhu, W.; Wang, N.; Ma, X.; Wu, C.; Wang, Q. Recent advances in the separation and purification of lactic acid from fermentation broth. Process Biochem. 2021, 104, 142–151. [Google Scholar] [CrossRef]
- Lee, H.D.; Lee, M.Y.; Hwang, Y.S.; Cho, Y.H.; Kim, H.W.; Park, H.B. Separation and purification of lactic acid from fermentation broth using membrane-integrated separation Processes. Ind. Eng. Chem. Res. 2017, 56, 8301–8310. [Google Scholar] [CrossRef]
- Liang, S.; Jiang, W.; Song, Y.; Zhou, S.F. Improvement and metabolomics-based analysis of d -lactic acid production from agro-industrial wastes by Lactobacillus delbrueckii submitted to adaptive laboratory evolution. J. Agric. Food Chem. 2020, 68, 7660–7669. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.H.; Kwon, E.Y.; Bae, S.J.; Cho, B.R.; Kim, S.Y.; Hahn, J.S. Improvement of D-lactic acid production in Saccharomyces cerevisiae under acidic conditions by evolutionary and rational metabolic engineering. Biotechnol. J. 2017, 12, 1–7. [Google Scholar] [CrossRef]
- Watcharawipas, A.; Sae-Tang, K.; Sansatchanon, K.; Sudying, P.; Boonchoo, K.; Tanapongpipat, S.; Kocharin, K.; Runguphan, W. Systematic engineering of Saccharomyces cerevisiae for D-lactic acid production with near theoretical yield. FEMS Yeast Res. 2021, 21, 1–15. [Google Scholar] [CrossRef]
- Wang, Y.D.; Liao, J.Y.; Chiang, C.J.; Chao, Y.P. A simple strategy to effectively produce D-lactate in crude glycerol-utilizing Escherichia coli. Biotechnol. Biofuels 2019, 12, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, D.; Tortajada, M.; Ramón, D.; Rojas, A. Production of D-lactic acid by the fermentation of orange peel waste hydrolysate by lactic acid bacteria. Fermentation 2020, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Beitel, S.M.; Coelho, L.F.; Contiero, J. Efficient conversion of agroindustrial waste into D(-) lactic acid by Lactobacillus delbrueckii using fed-batch fermentation. Biomed Res. Int. 2020, 2020, 4194052. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Vadlani, P.V. Lactic acid production from biomass-derived sugars via co-fermentation of Lactobacillus brevis and Lactobacillus plantarum. J. Biosci. Bioeng. 2015, 119, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.; Panesar, P.S.; Saini, M.S. Optimization of process parameters and estimation of kinetic parameters for lactic acid production by Lactobacillus casei MTCC 1423. Biomass Convers. Biorefinery 2019, 9, 253–266. [Google Scholar] [CrossRef]
- Song, J.Y.; Park, J.S.; Kang, C.D.; Cho, H.Y.; Yang, D.; Lee, S.; Cho, K.M. Introduction of a bacterial acetyl-CoA synthesis pathway improves lactic acid production in Saccharomyces cerevisiae. Metab. Eng. 2016, 35, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Sun, X.; Zhu, H.; Jiang, R.; Luo, X.; Yin, L. An optimized fed-batch culture strategy integrated with a one-step fermentation improves L-lactic acid production by Rhizopus oryzae. World J. Microbiol. Biotechnol. 2018, 34, 74. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Liu, H.; Yang, Y.; Zhou, X.; Xiu, Z. High-efficient l-lactic acid production from inedible starchy biomass by one-step open fermentation using thermotolerant Lactobacillus rhamnosus DUT1908. Bioprocess Biosyst. Eng. 2021, 44, 1935–1941. [Google Scholar] [CrossRef]
- Tian, X.; Liu, X.; Zhang, Y.; Chen, Y.; Hang, H.; Chu, J.; Zhuang, Y. Metabolic engineering coupled with adaptive evolution strategies for the efficient production of high-quality L-lactic acid by Lactobacillus paracasei. Bioresour. Technol. 2021, 323, 124549. [Google Scholar] [CrossRef]
- Chen, H.; Chen, B.; Su, Z.; Wang, K.; Wang, B.; Wang, Y.; Si, Z.; Wu, Y.; Cai, D.; Qin, P. Efficient lactic acid production from cassava bagasse by mixed culture of Bacillus coagulans and Lactobacillus rhamnosus using stepwise pH controlled simultaneous saccharification and co-fermentation. Ind. Crops Prod. 2020, 146, 112175. [Google Scholar] [CrossRef]
- Okano, K.; Uematsu, G.; Hama, S.; Tanaka, T.; Noda, H.; Kondo, A.; Honda, K. Metabolic engineering of Lactobacillus plantarum for direct L-lactic acid production from raw corn starch. Biotechnol. J. 2018, 13, 1–6. [Google Scholar] [CrossRef]
- Gao, M.; Ma, X.; Song, N.; Wang, Q.; Wu, C. A newly isolated strain, Lactobacillus paracasei subsp. paracasei 2, produces L-lactic acid from pilot-scale fermentation of food waste under sterile and nonsterile conditions. J. Chem. Technol. Biotechnol. 2020, 95, 3193–3201. [Google Scholar] [CrossRef]
- Li, C.; Ong, K.L.; Cui, Z.; Sang, Z.; Li, X.; Patria, R.D.; Qi, Q.; Fickers, P.; Yan, J.; Lin, C.S.K. Promising advancement in fermentative succinic acid production by yeast hosts. J. Hazard. Mater. 2021, 401, 123414. [Google Scholar] [CrossRef]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzyme Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Putri, D.N.; Sahlan, M.; Montastruc, L.; Meyer, M.; Negny, S.; Hermansyah, H. Progress of fermentation methods for bio-succinic acid production using agro-industrial waste by Actinobacillus succinogenes. Energy Rep. 2020, 6, 234–239. [Google Scholar] [CrossRef]
- E4tech; RE-CORD; WUR. “From the Sugar Platform to Biofuels and Biochemicals”. Final Report for the European Commission; Contract No. ENER/C2/423-2012/SI2.673791; European Commission: Luxembourg, 2015. [Google Scholar]
- Pateraki, C.; Patsalou, M.; Vlysidis, A.; Kopsahelis, N.; Webb, C.; Koutinas, A.A.; Koutinas, M. Actinobacillus succinogenes: Advances on succinic acid production and prospects for development of integrated biorefineries. Biochem. Eng. J. 2016, 112, 285–303. [Google Scholar] [CrossRef]
- Dessie, W.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Opportunities, challenges, and future perspectives of succinic acid production by Actinobacillus succinogenes. Appl. Microbiol. Biotechnol. 2018, 102, 9893–9910. [Google Scholar] [CrossRef] [PubMed]
- Cok, B.; Tsiropoulos, I.; Roes, A.L.; Patel, M.K. Succinic acid production derived from carbohydrates: An energy and greenhouse gas assessment of a platform chemical toward a bio-based economy. Biofuels Bioprod. Bioref 2014, 8, 16–29. [Google Scholar] [CrossRef]
- Ito, Y.; Hirasawa, T.; Shimizu, H. Metabolic engineering of Saccharomyces cerevisiae to improve succinic acid production based on metabolic profiling. Biosci. Biotechnol. Biochem. 2014, 78, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Cheng, K.K.; Wang, G.Y.; Zeng, J.; Zhang, J.A. Improved succinate production by metabolic engineering. Biomed Res. Int. 2013, 2013, 538790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van De Graaf, M.J.; Valianpoer, F.; Fiey, G.; Delattre, L.; Schulten, E.A.M. Process for the Crystallization of Succinic Acid 2015. US20150057425A1, 26 February 2015. [Google Scholar]
- Ahn, J.H.; Jang, Y.S.; Lee, S.Y. Production of succinic acid by metabolically engineered microorganisms. Curr. Opin. Biotechnol. 2016, 42, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Stylianou, E.; Pateraki, C.; Ladakis, D.; Damala, C.; Vlysidis, A.; Latorre-Sánchez, M.; Coll, C.; Lin, C.S.K.; Koutinas, A. Bioprocess development using organic biowaste and sustainability assessment of succinic acid production with engineered Yarrowia lipolytica strain. Biochem. Eng. J. 2021, 174, 108099. [Google Scholar] [CrossRef]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Succinate production in dual-phase Escherichia coli fermentations depends on the time of transition from aerobic to anaerobic conditions. J. Ind. Microbiol. Biotechnol. 2002, 28, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Salma, A.; Abdallah, R.; Fourcade, F.; Amrane, A.; Djelal, H. A New Approach to produce succinic acid through a co-culture system. Appl. Biochem. Biotechnol. 2021, 193, 2872–2892. [Google Scholar] [CrossRef] [PubMed]
- Kuenz, A.; Hoffmann, L.; Goy, K.; Bromann, S.; Prüße, U. High-level production of succinic acid from crude glycerol by a wild type organism. Catalysts 2020, 10, 470. [Google Scholar] [CrossRef]
- Chen, P.; Tao, S.; Zheng, P. Efficient and repeated production of succinic acid by turning sugarcane bagasse into sugar and support. Bioresour. Technol. 2016, 211, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Schroder, H.; Haefner, S.; Von Abendroth, G.; Hollmann, R.; Raddatz, A.; Ernst, H.; Gurski, H. Microbial Ssccinic Acid Producers and Purification of Succinic Acid. U.S. Patent 8,673,598, 18 March 2014. [Google Scholar]
- Li, C.; Gao, S.; Yang, X.; Lin, C.S.K. Green and sustainable succinic acid production from crude glycerol by engineered Yarrowia lipolytica via agricultural residue based in situ fibrous bed bioreactor. Bioresour. Technol. 2018, 249, 612–619. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Cui, Z.; Zhu, Z.; Liu, Y.; Tang, Y.J.; Hou, J.; Qi, Q. Engineering of Yarrowia lipolytica transporters for high-efficient production of biobased succinic acid from glucose. Biotechnol. Biofuels 2021, 14, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Gao, C.; Li, J.; Hou, J.; Lin, C.S.K.; Qi, Q. Engineering of unconventional yeast Yarrowia lipolytica for efficient succinic acid production from glycerol at low pH. Metab. Eng. 2017, 42, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Ong, K.L.; Yang, X.; Lin, C.S.K. Bio-refinery of waste streams for green and efficient succinic acid production by engineered Yarrowia lipolytica without pH control. Chem. Eng. J. 2019, 371, 804–812. [Google Scholar] [CrossRef]
- Porro, D.; Branduardi, P.; Sauer, M.; Mattanovich, D. Old obstacles and new horizons for microbial chemical production. Curr. Opin. Biotechnol. 2014, 30, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Crater, J.S.; Lievense, J.C. Scale-up of industrial microbial processes. FEMS Microbiol. Lett. 2018, 365, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Soares-Silva, I.; Ribas, D.; Sousa-Silva, M.; Azevedo-Silva, J.; Rendulić, T.; Casal, M. Membrane transporters in the bioproduction of organic acids: State of the art and future perspectives for industrial applications. FEMS Microbiol. Lett. 2020, 367, 1–15. [Google Scholar] [CrossRef]
- Lu, J.; Lv, Y.; Qian, X.; Jiang, Y.; Wu, M.; Zhang, W.; Zhou, J.; Dong, W.; Xin, F.; Jiang, M. Current advances in organic acid production from organic wastes by using microbial co-cultivation systems. Biofuels Bioprod. Biorefining 2020, 14, 481–492. [Google Scholar] [CrossRef]
- Steiger, M.G.; Mattanovich, D.; Sauer, M. Microbial organic acid production as carbon dioxide sink. FEMS Microbiol. Lett. 2017, 364, 1–4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Branduardi, P.; Sauer, M. Microbial carbon dioxide fixation: New tricks for an old game. FEMS Microbiol. Lett. 2018, 365, 1–7. [Google Scholar] [CrossRef] [PubMed]
Scheme 1.
Structures of organic acids produced at pilot scale by microbial processes and discussed in this review.
Scheme 1.
Structures of organic acids produced at pilot scale by microbial processes and discussed in this review.

Scheme 2.
Representation of the main metabolic pathways involved in the production of citric, fumaric, itaconic, lactic and succinic acid.
Scheme 2.
Representation of the main metabolic pathways involved in the production of citric, fumaric, itaconic, lactic and succinic acid.

Table 1.
The 2004 and 2010 “revisited” US Department of Energy’s top Sugar-derived Building Block Chemicals [2,3].
| 2004 | 2010 |
|---|---|
| 1,4-Diacids (Succinic, fumaric and malic acids) | 3-Hydroxypropionic acid/aldehyde |
| 2,5-Furan dicarboxylic | Bio-hydrocarbons |
| 3-Hydroxybutyrolactone | Ethanol |
| 3-Hydroxy propionic acid | Furans |
| Aspartic acid | Glycerol and derivatives |
| Glucaric acid | Lactic acid |
| Aylitol/Arabitol | Levulinic acid |
| Glutamic acid | Succinic acid |
| Itaconic acid | Sorbitol |
| Levulinic acid | Xylitol |
| Glycerol | |
| Sorbitol |
Table 3.
Fumaric acid production with different substrates, microorganisms, and fermentation strategies.
Table 3.
Fumaric acid production with different substrates, microorganisms, and fermentation strategies.
| Microorganism | Substrate | Fumaric Acid (g/L) | Fermentation Mode | Reference |
|---|---|---|---|---|
| R. arrhizus NRRL 1526 | Glucose | 130 | Batch | [74] |
| R. arrhizus NRRL 1526 | Perennial grasses | 8.9 | Shake flask | [75] |
| R. oryzae ATCC20344 | Lignocellulosic Syrup | 34.2 | Shake flask | [82] |
| R. arrhizus RH7-13 | Food Waste | 32.7 | Shake flask | [78] |
| R. oryzae NRRL 1526 | Brewery wastewater | 31.3 | Shake flask | [85] |
| R. arrhizus RH-07-13 | Glucose and glycerol | 22.8 | Shake flask | [84] |
| R. arrhizus RH-7−13-9 | Glucose | 17.5 | Simultaneous fermentation and separation | [94] |
| Mutant E. coli E2 | Glycerol | 41.5 | Fed-batch | [69] |
| Mutant S. cerevisiae | Glucose | 33.1 | Shake flask | [71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Di Lorenzo, R.D.; Serra, I.; Porro, D.; Branduardi, P. State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts 2022, 12, 234. https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020234
AMA Style
Di Lorenzo RD, Serra I, Porro D, Branduardi P. State of the Art on the Microbial Production of Industrially Relevant Organic Acids. Catalysts. 2022; 12(2):234. https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020234
Chicago/Turabian StyleDi Lorenzo, Raffaella Desirè, Immacolata Serra, Danilo Porro, and Paola Branduardi. 2022. "State of the Art on the Microbial Production of Industrially Relevant Organic Acids" Catalysts 12, no. 2: 234. https://0-doi-org.brum.beds.ac.uk/10.3390/catal12020234
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.