1. Introduction
Persistent hyperfiltration is one of the major causes of a loss of glomerular function and the progression of chronic kidney disease (CKD), but the underlying mechanism is not well understood. Hemodynamic parameters, including renal blood flow, glomerular capillary pressure (P
GC), single nephron glomerular filtration rate (SNGFR), filtration fraction, and decreased hydraulic conductivity associated with glomerular hypertrophy, have been used to study hyperfiltration, leading to a detailed analysis of these parameters [
1,
2]. Our laboratory and a few other groups have recently started investigating biomechanical forces that mediate the effects of hyperfiltration in the glomerulus [
3,
4,
5,
6].
Two types of biomechanical forces are believed to mediate the effects of hyperfiltration on glomerular function and structure, namely, tensile stress and fluid flow shear stress (FFSS), which were recently reviewed by Srivastava et al. [
5,
6]. Podocytes localized in Bowman’s space are most vulnerable to increased biomechanical forces [
7,
8,
9]. Primary processes from the large soma of podocytes further branch into foot processes covering the glomerular basement membrane around the capillary and interdigitate to form slit pore junctions. The increased intra-capillary pressure, working outwardly perpendicular to the direction of blood flow in the capillary, stretches foot processes and generates tensile stress on the basolateral aspect of podocytes [
5]. The second biomechanical force, FFSS, is generated on podocyte soma and primary processes by the glomerular ultrafiltrate flowing through Bowman’s space [
5,
7]. Thus, P
GC and SNGFR become the principal determinants of tensile stress and FFSS, respectively [
5,
6]. We have shown a 1.5–2.0-fold increase in the calculated FFSS over podocytes in solitary kidney in mice and rats [
9].
The in vitro application of FFSS to podocytes resulted in an altered actin cytoskeleton, increased the secretion of prostaglandin E
2 (PGE
2), and increased the expression of cyclooxygenase (COX)2 and PGE
2 receptor EP2, but not EP4 [
10,
11,
12]. Additionally, we demonstrated an upregulated COX2-PGE
2-EP2 axis with an unchanged EP4 expression in unilaterally nephrectomized (UNX) mice, as well as an elevated urinary PGE
2 in children born with solitary kidney [
10,
13]. These findings established the significance of the COX2-PGE
2-EP2 axis in mediating the mechanoreception of FFSS by podocytes.
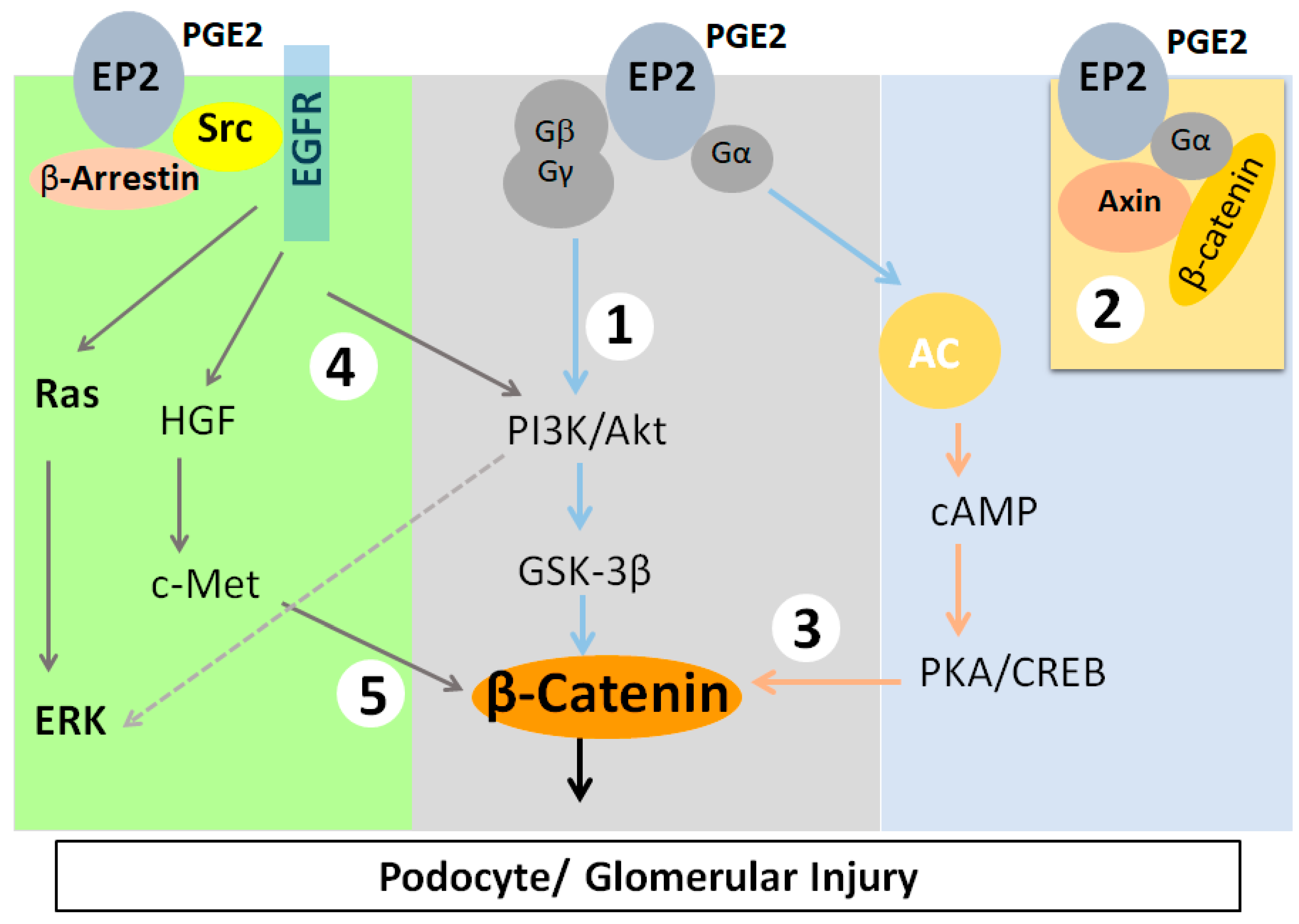
PGE
2 binding with EP2 receptor results in signaling events that induce β-catenin translocation to the nucleus (
Figure 1). Bioinformatics analysis to address the mechanotransduction that converts mechanical stimuli into biochemical changes identified Glycogen Synthase Kinase-3β (GSK3β) using Kinase enrichment analysis and Protein Kinase B (AKT1), GSK3β, and β-catenin through protein–protein interaction network analysis. Validation of these analyses using immortalized mouse podocytes led to the conclusion that AKT-GSK3β-β-catenin signaling plays a key role in podocyte response to FFSS in vitro [
14,
15].
Intracellular levels of β-catenin, a transcription factor, are normally kept low by the association of phosphorylated β-catenin with a degradation complex that targets it to the ubiquitin-proteasome degradation pathway in the absence of Wnt ligand. In the presence of Wnt bound to membrane receptor complex (LRP5/6 and frizzled), it is released from the degradation complex for translocation to the nucleus. Phosphorylation by Casein Kinase at Ser45, followed by GSK3β, at Ser33, Ser37, and Thr41 directs β-catenin for degradation, whereas phosphorylation by AKT at Ser552 and by Protein Kinase A (PKA) at Ser675 results in the nuclear translocation of β-catenin [
16,
17,
18]. Activated β-catenin signaling has been identified in Adriamycin-induced podocyte injury and albuminuria, oxidative stress-induced podocyte dysfunction, diabetic nephropathy, and focal segmental glomerulosclerosis [
19,
20]. The significance of β-catenin in solitary kidney injury and dysfunction is not known.
The present studies were undertaken to assess and confirm the upregulation of AKT-GSK3β-β-catenin signaling using a mouse model of UNX. We performed UNX in TOPGAL mice, a specific reporter mouse line with β-galactosidase activity as a direct in vivo readout for activated β-catenin signaling [
21]. The increased FFSS in solitary kidney following UNX resulted in the activation of β-catenin in TOPGAL mice. Next, we carried out in vitro experiments using cultured podocytes, which demonstrated phosphorylation of β-catenin at Ser552 as the main driver of β-catenin nuclear translocation. Additionally, we analyzed the publicly available datasets in the gene expression omnibus (GEO) database repository, which showed an upregulation of AKT-GSK3β-β-catenin in solitary kidney following UNX in mice [
22,
23]. Thus, studies using in vivo UNX TOPGAL mice, cultured podocytes in vitro, and bioinformatic data analysis on UNX mice provide combined evidence supporting our original in vitro findings and a better understanding of the molecular events associated with hyperfiltration-induced glomerular injury in solitary kidney.
2. Materials and Methods
2.1. Unilateral Nephrectomy of TOPGAL Mice
Animal studies involving TOPGAL mice were carried out using protocols approved by the Institutional Animal Care and Use Committee (IACUC), Institutional Biosafety Committee/Subcommittee on Research Safety (IBC/SRS), and the Research and Development (R&D) Committee at the VA Medical Center, Kansas City, MO. The study was carried out in compliance with the ARRIVE guidelines. All methods were performed in accordance with the relevant guidelines and regulations. The mice were maintained at AAALAC-approved facilities with unrestricted access to food and water under light/dark cycles of 12/12 h. The TOPGAL [Tg(TCF/Lef1-lacZ)34Efu] mouse carries a lacZ transgene on a CD1 background (Stock #4623, The Jackson Laboratory, Bar Harbor, ME, USA). The transgene contains the lacZ gene under the control of a promoter consisting of three consensus lymphoid enhancer binding factor 1/transcription factor 3 (LEF/TCF)-binding motifs upstream of a minimal Fos promoter [
21]. This allele is responsive to canonical Wnt/β-catenin signal transduction. Thus, the TOPGAL mouse is a specific reporter strain, with β–galactosidase activity as a direct in vivo readout for activated β–catenin signaling.
Four-week-old TOPGAL mice (
n = 47) underwent unilateral nephrectomy (UNX) to remove the right kidney (UNX) or sham operation (Sham). The animals recovered in their cages following the surgery, and the left kidney was harvested for analysis at 4 or 12 weeks following UNX. The day prior to the harvesting of the kidneys, urine was collected for urine albumin and creatinine measurements. At the time of the tissue retrieval, an intracardiac perfusion was performed with ice cold 4%PFA in PBS (pH 7.8) for 10 min at 4.4 mL/min, followed by ice cold 18% sucrose in PBS (pH 7.8) for 10 min at 4.4 mL/min. The fixative solutions were prepared and filtered to remove small particles which otherwise may block the capillaries. The resected kidney was then incubated overnight in 30% sucrose in PBS (pH 7.8) on ice [
24]. A part of the kidney was embedded in optimal cutting temperature compound (OCT) and flash frozen in liquid nitrogen. A part of the kidney was fixed in 10% formalin, processed, and embedded in paraffin.
2.2. Urine Albumin and CREATININE
Urine albumin was measured in triplicates using an Albuwell M kit (#1011, Exocell Inc., Philadelphia, PA, USA), following the manufacturer’s instructions. Urine creatinine was measured in duplicates using a companion creatinine kit (#1012, Exocell Inc., Philadelphia, PA, USA), following the manufacturer’s instructions.
2.3. Morphometric Analysis of Glomeruli in the TOPGAL Mouse Kidney
Paraffin-embedded tissue was sectioned at 3–5 μm and stained with Periodic Acid-Schiff (PAS) stain. PAS-stained kidney sections were evaluated for glomerular characteristics, including the size, mesangial matrix, and mesangial cell count. The glomerular area and perimeter included only the glomerular tufts and not the Bowman’s space or the parietal epithelial cells. The mesangial area included the PAS-stained matrix and cells. The mesangial cell counts were derived from the most proliferative mesangial stalk of each glomerulus, but not immediately adjacent to a vascular stalk. The counts were taken from cross sections and not the longitudinal section of the stalk, as described by Roberts et al. [
25]. Images were obtained using an Olympus BX60 (Hamburg, Germany) for light microscopy and analyzed using the Image J software suite (National Institute of Health and the Laboratory for Optical and Computational Instrumentation at the University of Wisconsin, Madison, WI, USA) [
26].
2.4. X-Gal Staining for β-Galactosidase Expression in TOPGAL Mice
The tissue sections (8–10 μm in thickness) were fixed by immersion in 2% glutaraldehyde, 0.01% sodium deoxycholate, 0.02% IGEPAL-CA630, and 1 mM MgCl
2 in PBS [pH = 7.8]) for 10 min and washed in a LacZ wash buffer (2 mM MgCl
2, 0.01% Na-deoxycholate, and 0.02% NP40 in PBS). The sections were immersed and incubated overnight with gentle rocking in a staining solution (1 mg/mL X-gal, 5 mM potassium ferricyanide, 5 mM potassium ferrocyanide, and 2 mM MgCl
2 in PBS, pH = 8.0) at 30 °C in the dark for 24 h in a humidified atmosphere [
24]. The tissue sections were washed with PBS, counterstained using a nuclear fast red staining solution, and washed with water for 1–2 min. Finally, the tissue sections were dehydrated through grades of ethanol and xylene and mounted in a non-aqueous mounting fluid. Using the images obtained after X-gal staining, endothelial cells were localized on the intraluminal aspect of the glomerular capillary, while podocytes were on the extraluminal aspect of the capillary wall. Semi-quantitative analysis was performed using a scale of 0–4 for X-gal staining within the glomerulus. Each glomerulus examined was assigned a score based upon the percentage of total podocytes that showed positive staining: 0 (no staining), 1 (<10% of podocytes), 2 (10–25% of podocytes), 3 (25–50% of podocytes), or 4 (>50% of podocytes staining).
2.5. Immunofluorescence Staining of TOPGAL Mouse Kidneys and Immortalized Mouse Podocytes
The tissue sections (4 μm) were fixed at 4 °C in 1:1 acetone and alcohol for 10–15 min, washed with PBS, and blocked in 5% donkey serum for 3 h at room temperature. The blocked sections were washed with PBS and incubated with primary antibodies for Podocalyxin (Goat Polyclonal Antibody from R&D, Catalog number AF1556, 1:1000 dilution), Total β-catenin (Rabbit Monoclonal Antibody from Cell Signaling, cs-8480S, 1:100 dilution), phospho-β-catenin (Ser675, Rabbit Monoclonal Antibody from Cell Signaling, cs-4176S, 1:100), phospho-β-catenin (Ser552, Rabbit Monoclonal Antibody from Cell Signaling, cs-5651S, 1:200 dilution), or β-galactosidase (Rabbit Polyclonal Antibody from Sigma, Catalog Number A-11132, 1:100 dilution) in a blocking solution overnight at 4 °C. The tissue slices were then washed and incubated with fluorescent-tagged secondary antibody (Donkey anti-rabbit Alexa Fluor 488, Invitrogen Catalog # A-21206 and Donkey anti-goat Alexa Fluor 594, Invitrogen Catalog # A-32758) for 1 h at room temperature and washed. The tissue sections were counterstained with Evan’s Blue for 4 min, washed, and mounted in an aqueous mounting fluid.
The cultured podocytes were fixed in 4% paraformaldehyde in PBS for 15 min at room temperature, washed in PBS, permeabilized in Triton X-100 0.1% in PBS for 10 min, and washed again. The cells were blocked in 1% goat serum/2.5% BSA in PBS for 2–3 h at room temperature, washed, and then incubated with primary antibodies in a blocking solution overnight at 4 °C, as described above for the tissue sections. The cells were washed and incubated in Alexa Fluor 488-tagged goat anti-rabbit secondary antibody (1:200 dilution) for 1 h at room temperature, washed, and mounted in an aqueous mounting fluid.
2.6. Confocal Microscopy and Measurement of Immunofluorescence Intensity
Confocal microscopy with Z-stacking was performed on a Zeiss LSM 510 META microscope with an LSM 510 laser module. We obtained images at 63× magnification (in immersion oil). The z-stack depth of the confocal images is approximately 4 μm (+/− 1.5 μm) with a an optical section (z-slice) depth of 0.317 μm. We converted stacked images into maximum-intensity-projection images for analysis. The cells were treated with FFSS in triplicate, and 3–5 images were obtained per experiment. The photomultiplier tube voltage was kept constant throughout image acquisition to ensure comparable relative fluorescence intensities for all experimental conditions. Quantitative analysis of confocal microscope images was carried using the open-source Image-J/FIJI software to determine the predominant subcellular location of different phospho-β-catenin [
27]. The fluorescent staining for β-catenin protein in the nucleus was quantitated by measuring the relative fluorescence units (RFU) selected in the region of interest. We measured the net fluorescence integrated density in the nucleus using the area of the nucleus and the mean intensity of β-catenin immunofluorescence within the nucleus. We measured a minimum of 50 nuclei at each experimental time point. A similar protocol was followed for quantification in kidney tissue from TOPGAL animals.
2.7. Podocyte Cell Culture
Conditionally immortalized mouse podocytes containing thermosensitive tsA58 mutant T-antigen (kindly provided by Dr. Peter Mundel) were seeded on collagen-I coated 75 cm
2 polystyrene flasks and first propagated in RPMI 1640 containing L-glutamine, 10% fetal bovine serum, 100 units/mL penicillin, and 0.1 mg/mL streptomycin (Invitrogen, Carlsbad, CA, USA) supplemented with 10 units/mL of γ-interferon (Cell Sciences, Norwood, MA, USA) under permissive conditions at 33 °C, with 95% humidity and 5% CO
2 [
28]. The cells were then transferred to non-permissive conditions (37 °C without γ-interferon) to induce differentiation. The cells on standard glass slides (3 slides/dish, 12 mL medium) were maintained for differentiation and studied on day 14.
2.8. Fluid Flow Shear Stress (FFSS) Application
Fluid flow shear stress was applied to the differentiated podocytes using a FlexCell Streamer Gold apparatus (FlexCell International, Hillsborough, NC, USA), as described previously [
10,
14]. FFSS was applied at 2 dynes/cm
2 for 2 h and untreated podocytes (control group) were placed in the same incubator, without exposure to FFSS. The slides were returned to the dishes for recovery up to 24 h at 37 °C under a 5% CO
2-humidified atmosphere. The samples obtained from untreated podocytes at the end of the FFSS treatment, and at 2 h and 24 h following the FFSS treatment, were termed Control, End-FFSS, Post-2 h, and Post-24 h, respectively.
2.9. Gene Expression Omnibus Dataset on Unilaterally Nephrectomized Mice
The Gene Expression Omnibus (GEO, NCBI, (
http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/geo, access on 24 July 2017) public functional genomics data repository was searched for “uninephrectomy” and filtered for “
Mus musculus”, which returned 58 hits (as of 24 July 2017). We excluded results for uninephrectomy associated with ischemia-perfusion injury, diabetic nephropathy, drug treatment, and pooled samples. We identified the GEO dataset, GSE 53996, titled “Effects of high-fat induced obesity on gene expression in mouse kidney”. In the animal protocol used for obtaining this GEO dataset, male C57/BJ mice aged 6 weeks were randomly assigned to left uninephrectomy (UNX) or sham procedures and fed a high-fat diet or control chow diet. The mice were divided into 4 groups: Sham-chow, UNX-chow, Sham-HFD, and UNX-HFD. All mice were sacrificed under anesthesia at 20 weeks after surgery, and their kidneys were harvested. Microarray was performed on Agilent-026655 Mouse GE 4x44K v2 [
29,
30]. To evaluate the transcriptomic changes induced by unilateral nephrectomy alone, avoiding the effect of the high-fat diet, the present bioinformatics analysis included only the groups maintained on normal chow. These included the sham samples (
n = 4): GSM1305179 LFD-sham_1, GSM1305180 LFD-sham_2, GSM1305181 LFD-sham_3, GSM1305182 LFD-sham_4, and UNX samples (
n = 4): GSM1305183 LFD-UNX_1, GSM1305184 LFD-UNX_2, GSM1305185 LFD-UNX_3, and GSM1305186 LFD-UNX_4.
2.10. Functional Annotation, Pathway, and Network Analyses Using Ingenuity Pathway Analysis (IPA)
To identify over-represented canonical pathways, networks, and upstream regulators, differentially expressed genes (p < 0.05) between sham and unilateral nephrectomy were submitted for the Ingenuity Pathways Knowledge Base tool (IPA, Ingenuity Systems, Inc., Redwood City, CA, USA). Core analysis was performed by comparing the significant genes with the IPA knowledge base, comprising curated pathways within the program on 24 July 2017. IPA predicted significant biological functions and pathways (p < 0.05, Fischer’s exact test) affected in the kidney following unilateral nephrectomy. In addition, IPA was used to predict the upstream biological regulators and the downstream effects on cellular and organismal biology. In the analysis of the upstream regulators, such as transcription factors, kinases, receptors, etc., the further the activation z-score is away from zero, the more likely it is that the direction of change of the target genes is consistent with the upstream regulator being in an activated or inhibited state. The settings for the core analyses were as follows: Ingenuity Knowledge Base; Endogenous Chemicals included Direct and Indirect relationships; molecules per pathway: 35; and networks per analysis: 25.
2.11. Statistics
We used the SPSS 23 statistical software for the preparation of the statistics and graphs. ANOVA was used for three-group comparisons of the podocyte experiments, and Students’ t-test was used for two-group comparisons. A p-value < 0.05 was considered significant.
4. Discussion
Hyperfiltration is a major underlying factor in the onset and progression of CKD, which is associated with podocyte injury, resulting in impairment of the glomerular filtration barrier and a decreased renal function over time. The present in vivo results validate previous in vitro studies that described the significance of β-catenin signaling in FFSS-treated podocytes [
14,
15]. The present results show glomerular hypertrophy and an increased mesangial matrix in unilaterally nephrectomized TOPGAL mice. The glomerular hypertrophy was more marked in juxtaglomerular glomeruli in the early period (4 weeks) and in cortical glomeruli in the later period (12 weeks). The results also show an increased albuminuria at 3 months, increased nuclear translocation of β-catenin at 4 weeks, phosphorylation of β-catenin at Ser552 and not Ser675 in UNX mice, which was further complemented by in vitro studies in podocytes, and the bioinformatic analysis of publicly available independent datasets supports our previously reported activation of “COX2-PGE
2-EP2” and “AKT-GSK3β-β-catenin” in podocytes in vitro. These studies advance our understanding of the mechanism of CKD progression to ESRD in adult kidney donors with acquired solitary kidney and in children born with solitary functioning kidney.
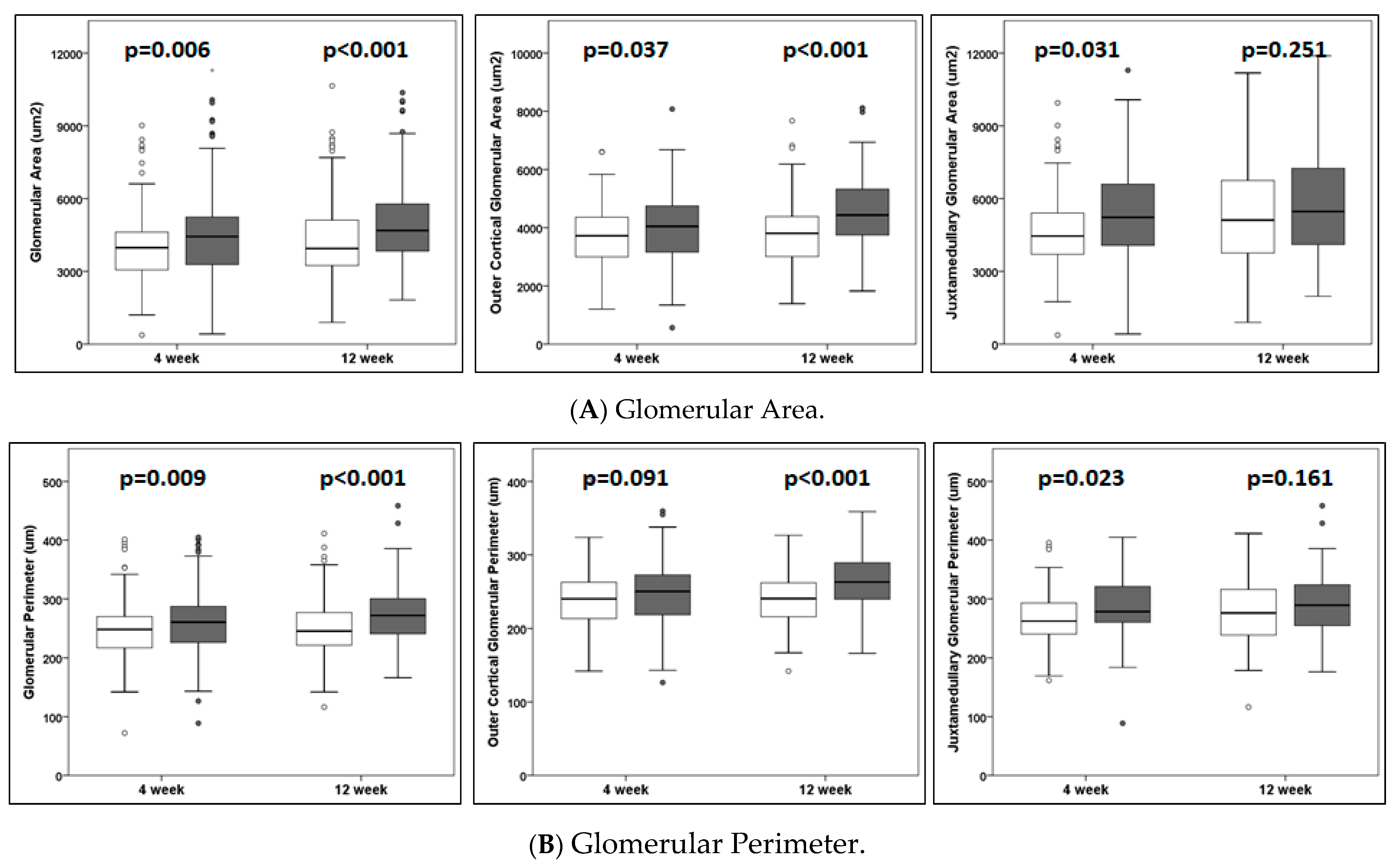
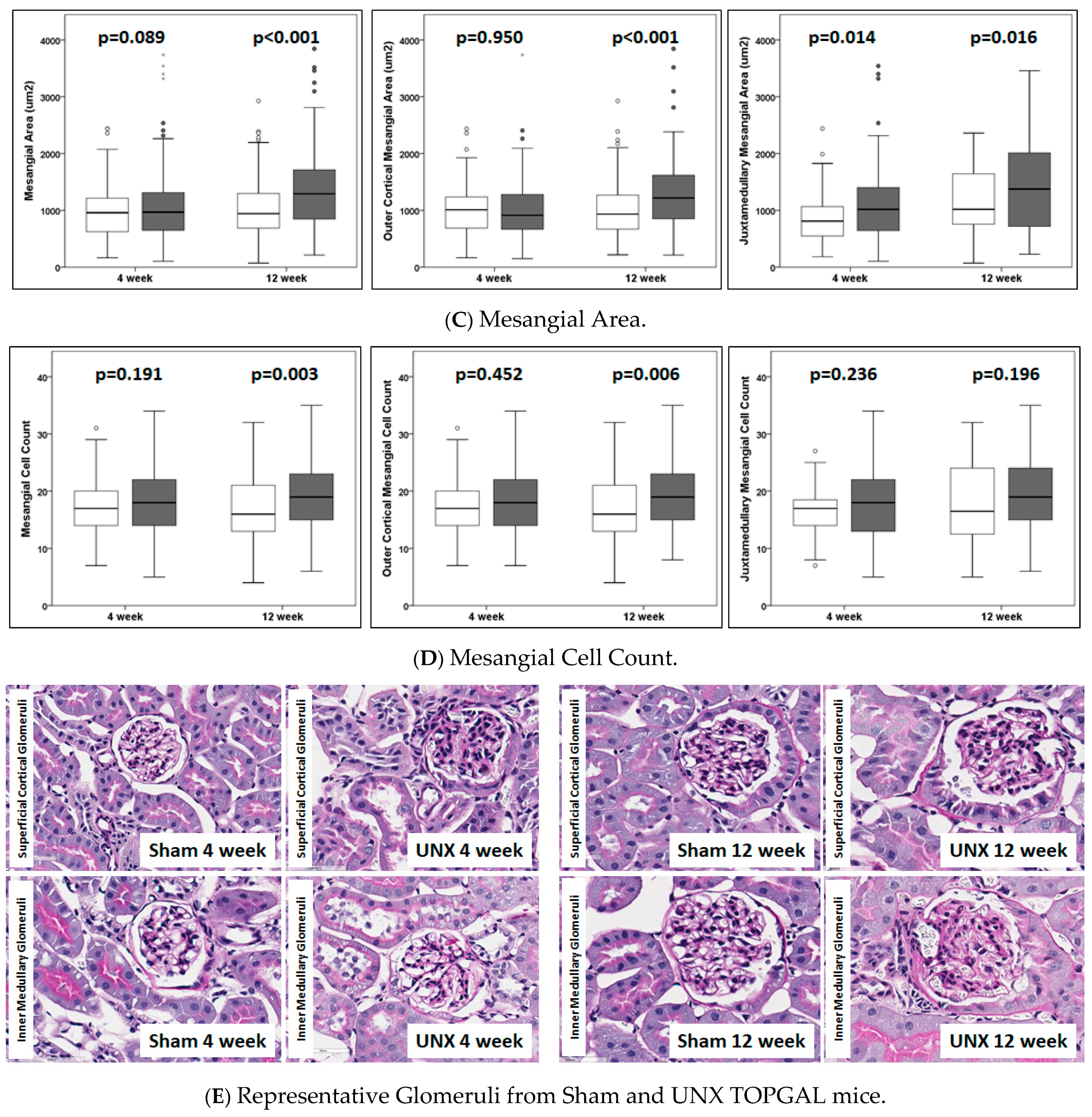
Glomerular hypertrophy is considered a hallmark of decreasing kidney function and proteinuric disease. Key in vivo findings, glomerular hypertrophy, and an increased mesangial matrix in TOPGAL mice were evident by 4 weeks post-UNX and showed further progression at 12 weeks (
Figure 2 and
Table 1). Previously, we reported an increased glomerular diameter and area as early as 2–3 weeks in C57BL/6J mice and by 4 weeks in sv129 mice following UNX [
9,
10]. Celsi et al. [
1,
2] reported an increase in glomerular diameter and area at ~7 weeks and not at ~2 weeks following UNX in Sprague Dawley rats. To further refine and advance our findings, we examined and compared the adaptive changes in cortical and juxtamedullary glomeruli. The glomerular hypertrophy was more marked in the juxtamedullary region at 4 weeks and in the cortical region by 12 weeks. A centrifugal pattern of renal maturation from the juxtamedullary region towards the superficial cortical layers leads to subtle regional differences in the structure and function of the nephrons [
31]. Thus, the larger juxtamedullary glomeruli, compared to the cortical glomeruli, in the present study correspond with normal size differences (
Table 1). Vascular damage from hypertension occurs first and more severely in the juxtamedullary glomeruli [
32]. A pathologic glomerular lesion of focal segmental glomerulosclerosis is most common in or restricted to the juxtamedullary cortex [
33]. Thus, the presently observed early glomerular adaptive changes in the juxtamedullary region are of special significance in the development of secondary focal segmental glomerulosclerosis from hyperfiltration-mediated injury.
The unilaterally nephrectomized TOPGAL mice showed an increased urinary albumin by 12 weeks, which was not evident at 4 weeks (
Figure 3). Recently, we showed sv129 mice with increased urine albumin following unilateral nephrectomy [
10]. The onset of albuminuria is preceded by increased albumin in the glomerular filtrate, as a result of a damaged glomerular filtration barrier. Using an in vitro assay developed in our laboratory, we demonstrated that a variety of agents and conditions increase glomerular albumin permeability (P
alb). We have also demonstrated that an increase in P
alb precedes the onset of proteinuria in animal models of diabetes, hypertension, radiation nephropathy, puromycin-induced nephrosis, and focal segmental glomerulosclerosis [
34,
35,
36,
37]. We have also demonstrated that treatment of isolated rat glomeruli with PGE
2 or FFSS increased P
alb, which could be blocked by indomethacin, indicating a role of PGE
2 [
10,
38,
39]. Thus, previously reported work and present observations show that UNX-induced increases in FFSS damage the glomerular filtration barrier causing increased albumin permeability and, subsequently, proteinuria. Similarly, children born with solitary kidney start to develop hypertension and proteinuria in late adolescence, suggesting that glomerular injury due to hyperfiltration gradually leads to proteinuria over several years in these children [
40,
41]. Thus, morphological changes, glomerular hypertrophy, and an increased mesangial matrix complement the observed functional outcome in TOPGAL mice following UNX.
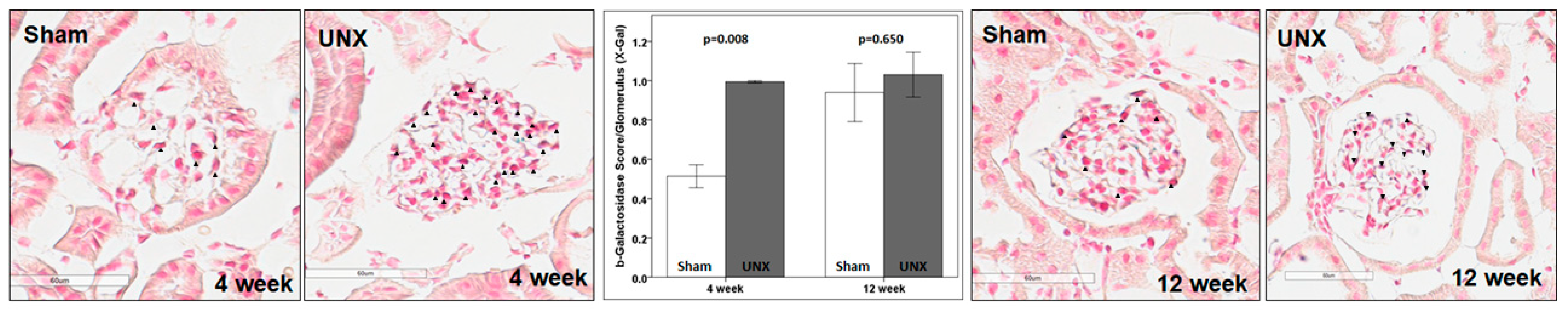
The TOPGAL mouse is a specific reporter strain with β-galactosidase activity, as a direct in vivo readout for activated β-catenin signaling. Using the TOPGAL mice, we demonstrated an increase in the β-galactosidase activity, as direct evidence for the in vivo activation of β-catenin following UNX. We used two different methods (described under Methods) to evaluate the β-galactosidase expression in TOPGAL and the data from each method support this finding. The increased β-galactosidase activity (i.e., β-catenin signaling) was detectable at 4 weeks following UNX and returned to the baseline by 12 weeks. We anticipated a transient activation of β-catenin signaling, since it is a transcription factor, and its activation would cause progressive changes.
A small but significant magnitude of increase in β-galactosidase activity at 4 weeks but not at 12 weeks in β-galactosidase expression is consistent with other reports. For example, the β-galactosidase activity in TOPGAL mice is observed during fetal and neonatal periods in the skeleton and is not detectable in mature animals, unless induced by mechanical loading or fracture in vivo [
42,
43]. In vivo bone loading in 5- and 12-months-old TOPGAL mice for 1 day and 5 day was shown to result in a β-galactosidase expression in 5% to 10% of osteocytes that peaked at 24 h and declined to baseline by 72 h [
44,
45]. Additionally, the expression of β-galactosidase in callus and periosteum following a fracture stimulus was observed in TOPGAL mice. Treatment with GSK-3β inhibitor resulted in an increased β-catenin activation and rapid fracture healing, showing the importance of β-catenin in response to injury [
43]. Thus, stress or injury stimulate β-catenin signaling in a TOPGAL mouse model of UNX, which is similar to that observed in models of bone loading and fracture.
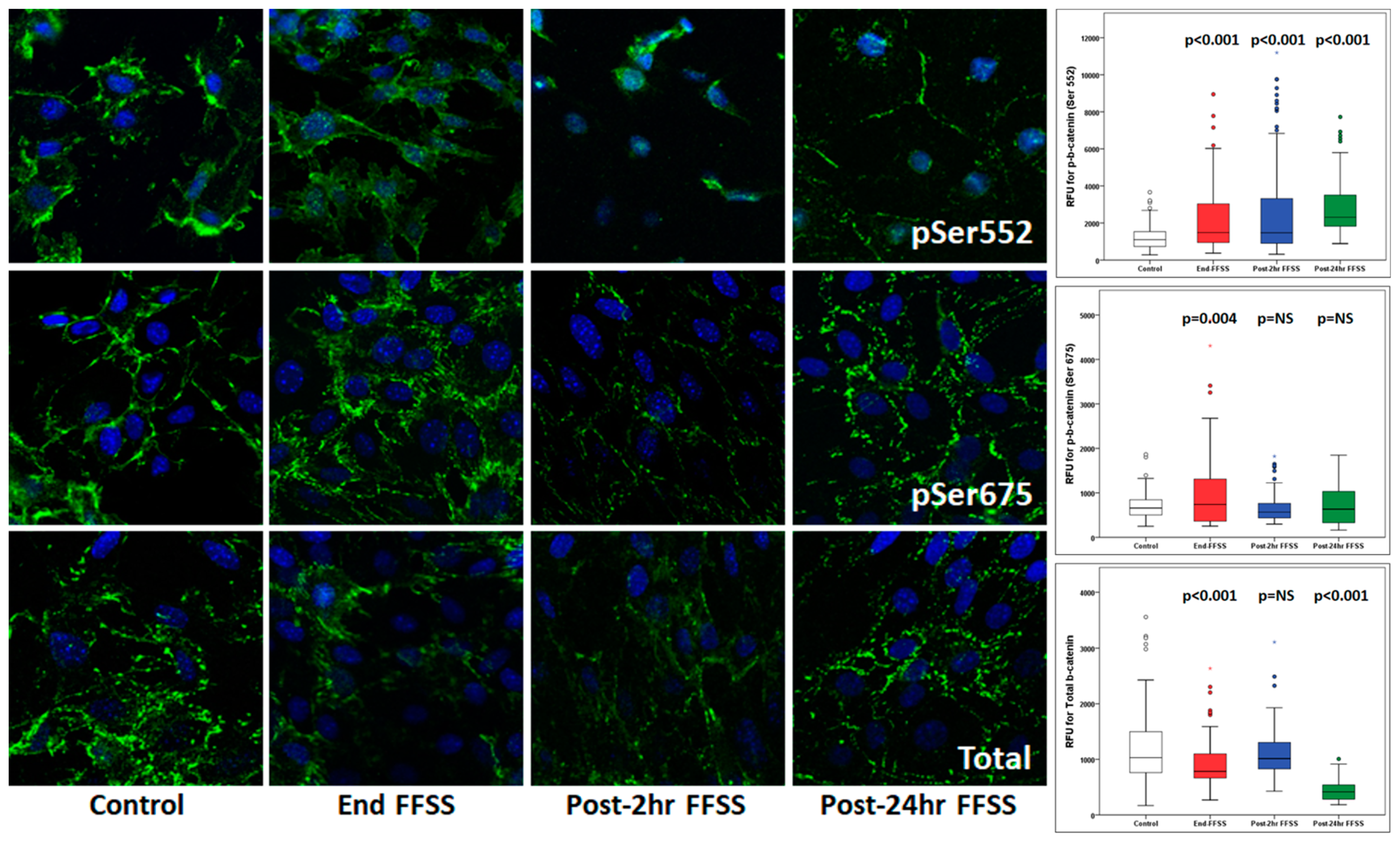
β-catenin phosphorylation by casein kinase (Ser45) followed by GSK3β (Ser33,37 and Thr41) directs it toward degradation through the ubiquitin-proteasome degradation pathway [
18]. In contrast, phosphorylation by AKT (predominantly) at Ser552 and by PKA (predominantly) at Ser675 results in the nuclear translocation of β-catenin for transcriptional activity via LEF/TCF [
16,
17]. We previously identified β-catenin, a transcription factor, in podocytes treated with FFSS through Enrichr and IPA bioinformatic analyses as an upstream regulator [
14]. Western blot analysis of podocytes treated with PGE
2 (1 µM) or FFSS showed an increased phosphorylation of β-catenin (Ser552) but not β-catenin (Ser675) [
14]. Western blotting also showed the upregulation of CD44/ERBB2/β-catenin pathway in podocytes following FFSS [
15].
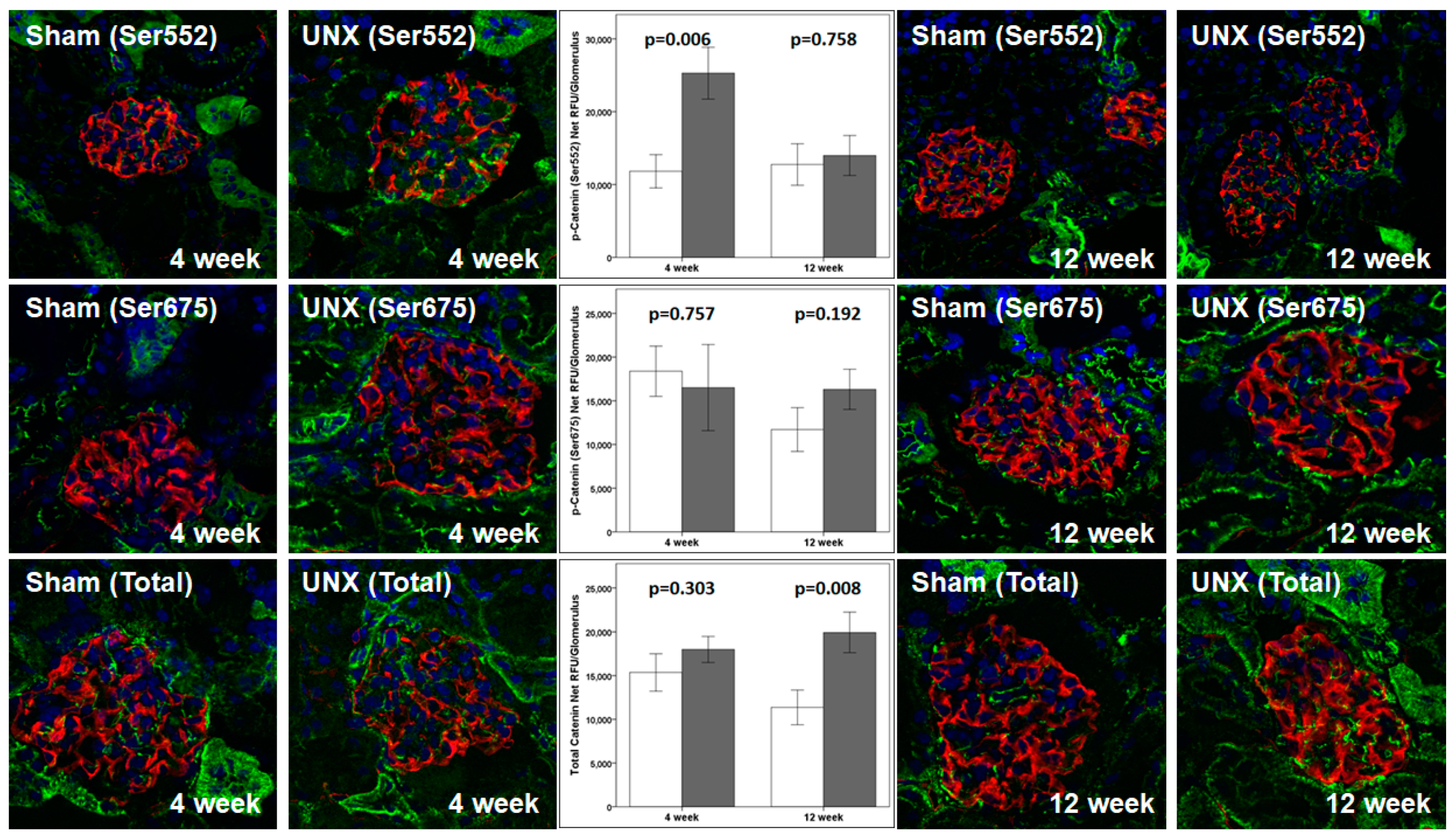
Presently, we evaluated the localization and movement of phospho-β-catenin (Ser552), phospho-β-catenin (Ser675), and total β-catenin using confocal microscopy. The results (
Table 4 and
Figure 7) show the translocation to and accumulation of phospho-β-catenin (Ser552) in the nucleus. A transient increase in phospho-β-catenin (Ser675) suggests a role of AKT-GSK3β-β-catenin, compared to cAMP-PKA-β-catenin, as observed previously [
14]. Additionally, confocal microscopy of the kidney from unilaterally nephrectomized TOPGAL mice showed an upregulated phospho-β-catenin (Ser552) at 4 weeks and Total β-catenin at 12 weeks, while phospho-β-catenin (Ser675) remained unchanged (
Table 3 and
Figure 6). The significance of β-catenin signaling is also highlighted by other reports, as it was found to mediate oxidative stress-induced podocyte dysfunction in vitro and in vivo [
19]. Podocyte injury and albuminuria caused by Adriamycin were ameliorated in β-catenin knock-out mice, and the over-expression of podocyte β-catenin was also detected in human diabetic nephropathy and focal segmental glomerulosclerosis [
20]. Additionally, treatment of osteocytes with PGE
2 results in the EP2-activated phosphorylation of GSK3β and increased nuclear translocation of β-catenin, followed by binding with LEF/TCF and transcriptional regulation of genes, including COX2, suggesting a role of β-catenin in skeletal physiology [
46,
47,
48,
49,
50]. Thus, understanding the importance of β-catenin in FFSS-mediated injury in solitary kidney will be critical for developing strategies to treat the progression of CKD in transplant donors with acquired solitary kidney and children born with solitary functioning kidneys.
Considering its potential clinical significance, we adopted a third pre-clinical approach to corroborate our finding that hyperfiltration-induced kidney injury causes the upregulation of β-catenin signaling. To this end, we performed bioinformatic analysis using publicly available dataset in the Gene Expression Omnibus. As shown in
Table 5,
Table 6,
Table 7 and
Table 8, the IPA analyses showed G-protein coupled receptor signaling as the main pathway, with prostanoid receptor EP2 as the G-protein-coupled protein receptor, AKT1, GSK3β, Erbb, and mTOR as the kinases, and β-catenin as the transcription factor. Previously, we validated the phosphorylation of AKT1 (Ser473), GSK3β (Ser9), and β-catenin (Ser552 and Ser675) [
14]. Additionally, we reported the altered phosphorylation of Erbb2 (Tyr1221/1222), mTOR (Ser2448), ERK1/2 (Thr202/Tyr204), and p38MAPK (Thr180/Tyr182), but not PKA (Thr197) [
14,
15]. Thus, the mechanoperception and mechanotransduction pathways that we have previously identified were validated from a dataset generated by other investigators [
29,
30]. Therefore, based on our previous work, the present in vitro and in vivo results, and an analysis of a publicly available dataset, we propose β-catenin signaling to be relevant for understanding hyperfiltration-induced injury in solitary kidney.
Since PGE
2 receptors are known to activate diverse signaling pathways, the activity of several proteins is likely affected due to FFSS. Of the four PGE
2 receptors, only EP2 activation results in the nuclear translocation of activated β-catenin through five known intermediate mechanisms (
Figure 1). EP2 is a G-protein-coupled receptor that activates the heterotrimeric (αβγ) Gs protein, leading to (i) the release of the Gβγ complex, activating PI3K/AKT, which phosphorylates and inactivates GSK3β [
51,
52], (ii) binding of the Gα complex to Axin and the release of β-catenin from the Axin-β-catenin-GSK3β complex, (iii) generation of cAMP and activation of PKA, which activate β-catenin [
53,
54,
55], (iv) recruitment of β-arrestin-1 and phosphorylation of Src-kinase, which transactivates the EGFR signaling network of the PI3K/AKT, HGF/c-Met, and Ras/ERK pathways [
56,
57,
58], and (v) activation of PI3K/AKT leading to the activation of ERK [
59]. Studies suggest that the phosphorylation of β-catenin at Ser552 or Ser675 induces β-catenin translocation to the nucleus, with AKT preferentially phosphorylating β-catenin at Ser552, while PKA preferentially phosphorylating it at Ser675 [
16,
17,
55,
60]. Thus, there is a strong link between G-protein-coupled receptor signaling via the EP2 receptor and β-catenin signaling, which could be targeted to mitigate injury.
We believe that there are at least two clinical scenarios where susceptibility to CKD needs special attention, as the reason(s) for the progression to ESRD is still unclear, and specific treatment(s) are not available: (a) in adults following kidney donation and (b) in children born with solitary functioning kidney. We recently reviewed the accumulating evidence that living kidney donation increases the risk of ESRD in donors, despite stringent screening [
61]. The risk of ESRD in donors is 8.3-fold higher, compared to eligible healthy individuals, as reported in a meta-analysis of 52 studies [
62,
63,
64,
65]. There is a cumulative increase in the incidence of ESRD over time following kidney donation, with a reported mean interval of 27.1 ± 9.8 years [
66,
67,
68]. Secondly, a considerable number of children born with solitary kidney develop albuminuria during adolescence and progress to end-stage renal disease as young adults [
69,
70,
71,
72]. Children with SFK manifest renal injury at a median age of ~15 years with hypertension, proteinuria, and/or an eGFR of less than 60 mL/min/1.73 m
2 [
69,
70,
71,
72]. The significance of FFSS in hyperfiltration-mediated glomerular injury in the progression of CKD is being better understood and appreciated. With growing evidence, we propose EP2 and/or β-catenin as suitable targets for mitigating glomerular injury.


 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}