
Dose and Dose Rate-Dependent Effects of Low-Dose Irradiation on Inflammatory Parameters in ApoE-Deficient and Wild Type Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice, Irradiation and Analysis of Plasma Samples
2.2. Expression of Immune Parameters in Spleen
2.3. In Vitro Experiments
2.4. Statistical Analysis
3. Results
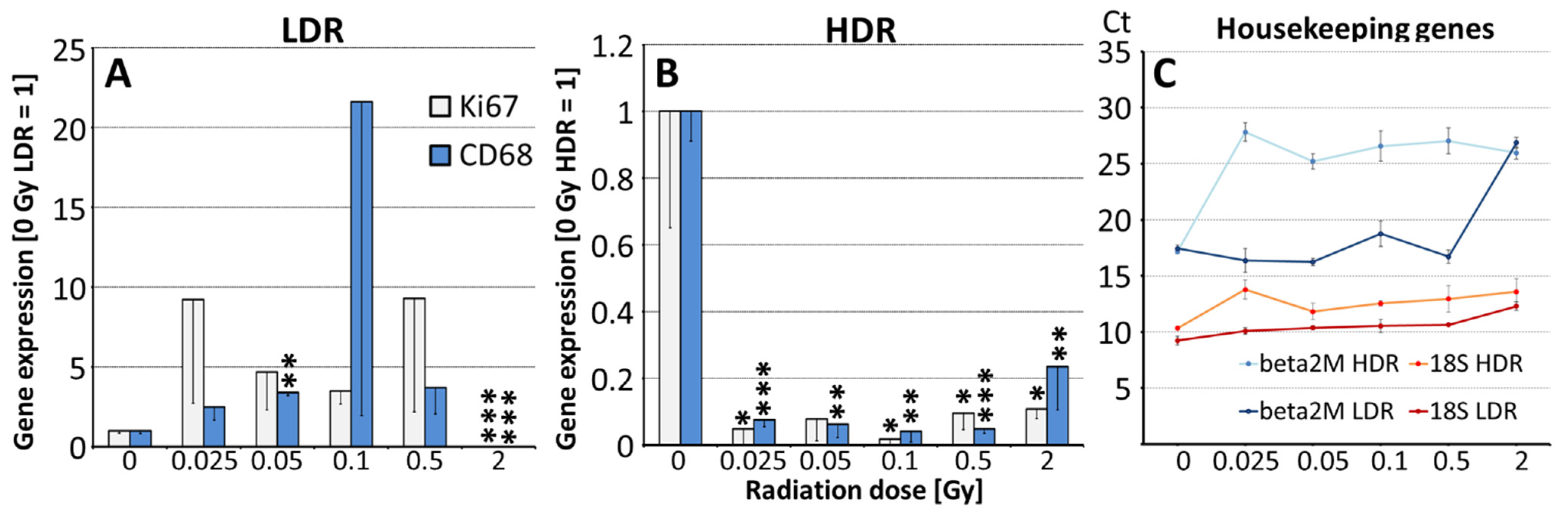
3.1. In Vivo Effects of Irradiation on the Spleen
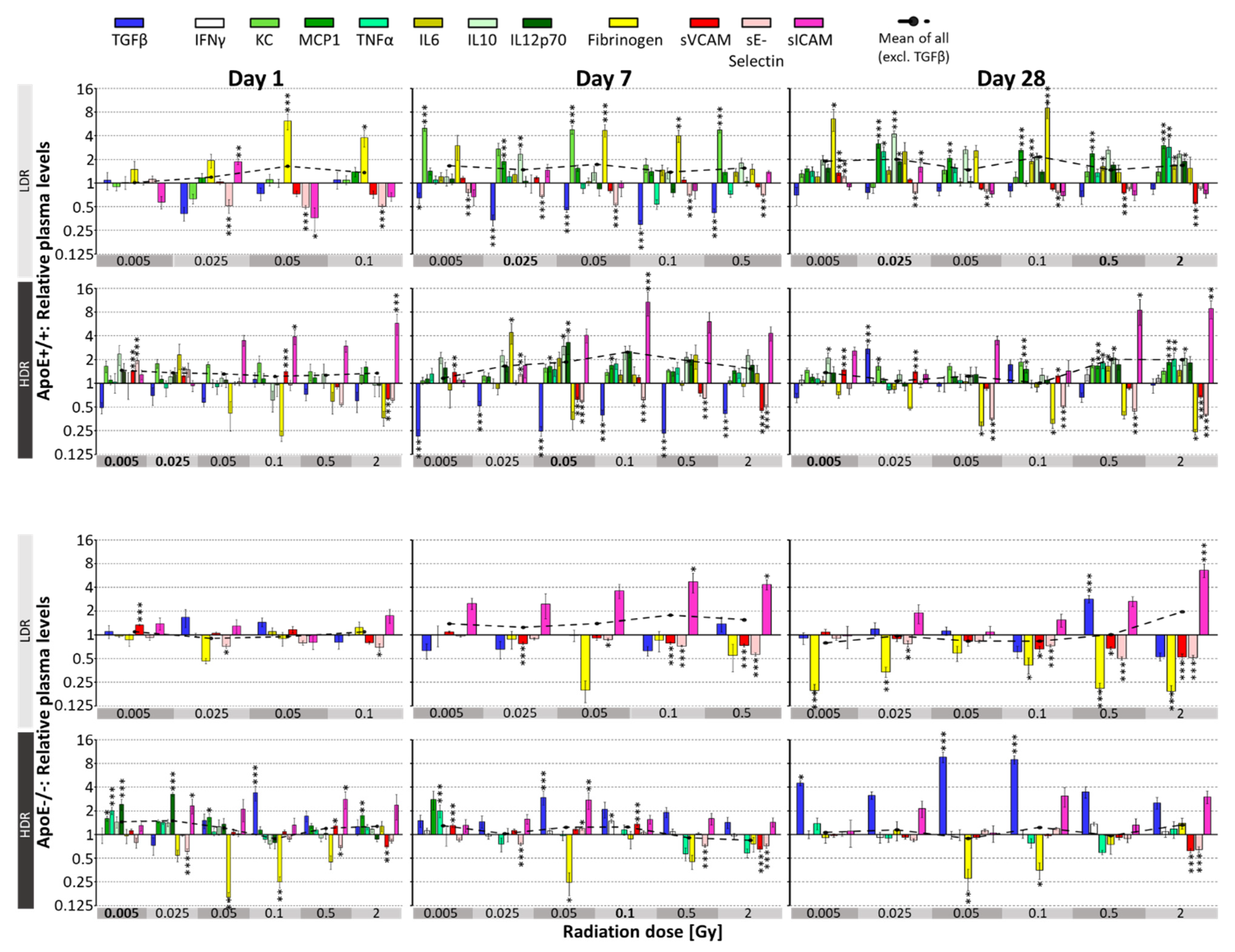
3.2. Irradiation-Induced Effects on Plasma Parameters in Mice
3.3. Age-Related Effects on Plasma Parameters
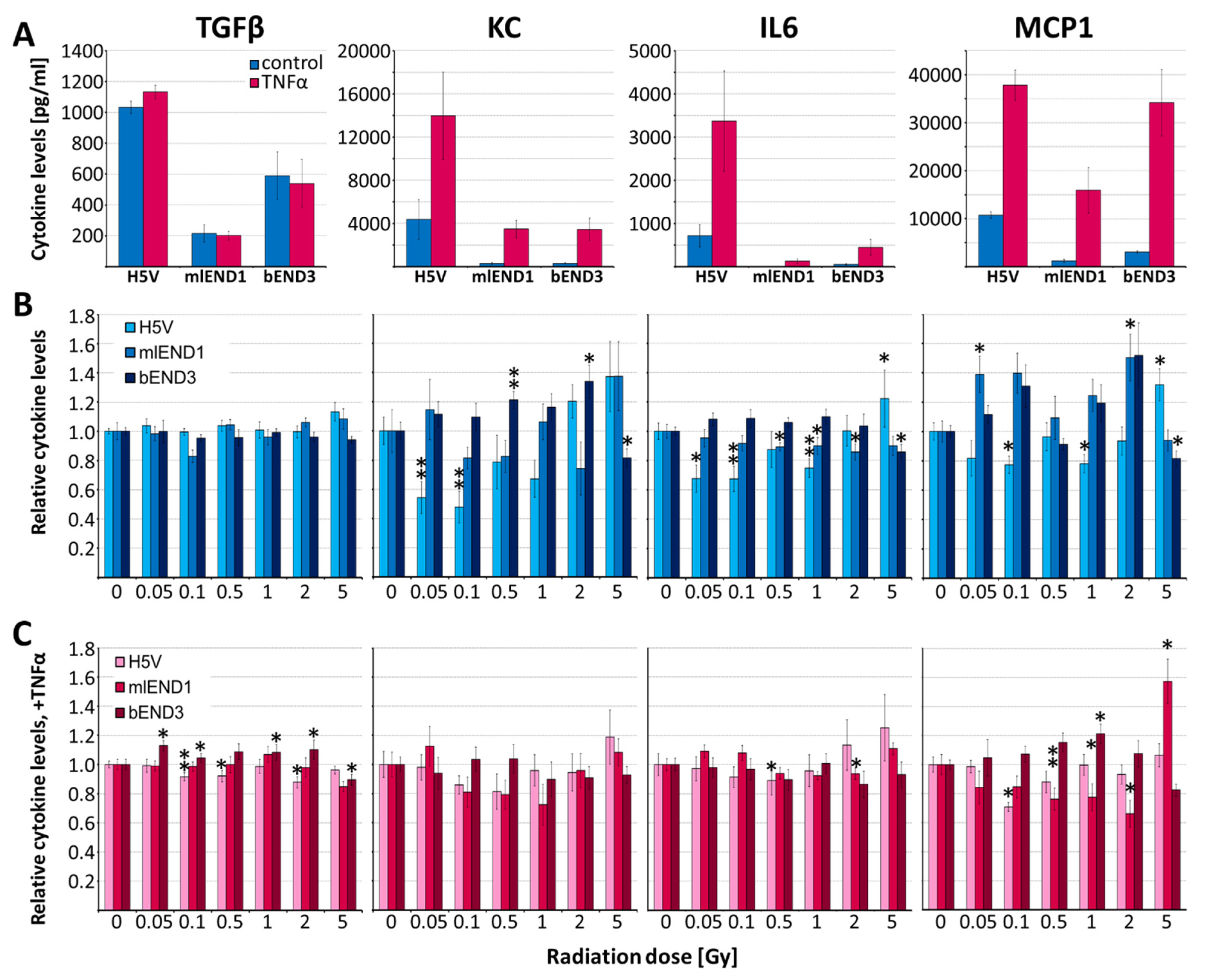
3.4. Irradiation of EC Lines—Effects on Cytokine Release and Monocyte Adhesion
4. Discussion
4.1. Systemic In Vivo Responses after Whole Body Irradiation (WBI)
4.2. Age-Related Plasma Effects
4.3. In Vitro Effects of Irradiation on Endothelial Cells
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ApoE | Apolipoprotein E |
| CD68 | Cluster of Differentiation 68 |
| EC | endothelial cell |
| G-CSF | Granulocyte-Colony Stimulating Factor |
| INF | Interferon |
| IL | Interleukin |
| IR | irradiation |
| KC | Keratinocyte-derived chemokine |
| Ki67 | Marker of proliferation Ki-67 |
| LDR | low dose rate, HDR: high dose rate |
| MCP1 | Monocyte Chemoattractant Protein 1 |
| NO | nitric oxide |
| RANTES | Regulated on Activation Normal T cell Expressed and Secreted |
| ROS | reactive oxygen species |
| sE-selectin | soluble Endothelial Leukocyte Adhesion Molecule-1 |
| sICAM | soluble Intercellular Adhesion Molecule 1 |
| sVCAM | soluble Vascular Adhesion Molecule 1 |
| TGFβ | Transforming Growth Factor beta 1 |
References
- Formenti, S.C. Immunological aspects of local radiotherapy: Clinical relevance. Discov. Med. 2010, 9, 119–124. [Google Scholar]
- Hildebrandt, G.; Jahns, J.; Hindemith, M.; Spranger, S.; Sack, U.; Kinne, R.W.; Madaj-Sterba, P.; Wolf, U.; Kamprad, F. Effects of low dose radiation therapy on adjuvant induced arthritis in rats. Int. J. Radiat. Biol. 2000, 76, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Rödel, F.; Frey, B.; Gaipl, U.; Keilholz, L.; Fournier, C.; Manda, K.; Schöllnberger, H.; Hildebrandt, G.; Rödel, C. Modulation of inflammatory immune reactions by low-dose ionizing radiation: Molecular mechanisms and clinical application. Curr. Med. Chem. 2012, 19, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Large, M.; Hehlgans, S.; Reichert, S.; Gaipl, U.S.; Fournier, C.; Rödel, C.; Weiss, C.; Rödel, F. Study of the anti-inflammatory effects of low-dose radiation: The contribution of biphasic regulation of the antioxidative system in endothelial cells. Strahlenther. Und Onkol. 2015, 191, 742–749. [Google Scholar] [CrossRef]
- Seegenschmiedt, M.H.; Micke, O.; Willich, N. Radiation therapy for nonmalignant diseases in Germany. Current concepts and future perspectives. Strahlenther. Onkol. 2004, 180, 718–730. [Google Scholar] [CrossRef]
- Juniku, N.; Micke, O.; Seegenschmiedt, M.H.; Muecke, R. Radiotherapie bei schmerzhaften benignen muskuloskeletalen Erkrankungen: Ergebnisse einer retrospektiven klinischen Qualitätskontrolle. Strahlenther. Und Onkol. 2019, 195, 1068–1073. [Google Scholar] [CrossRef] [Green Version]
- Heyd, R.; Tselis, N.; Ackermann, H.; Röddiger, S.J.; Zamboglou, N. Radiation therapy for painful heel spurs: Results of a prospective randomized study. Strahlenther. Onkol. 2007, 183, 3–9. [Google Scholar] [CrossRef]
- Montero, A.; Sabater, S.; Rödel, F.; Gaipl, U.S.; Ott, O.J.; Seegenschmiedt, M.H.; Arenas, M. Is it time to redefine the role of low-dose radiotherapy for benign disease? Ann. Rheum. Dis. 2020, 79, e34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sautter-Bihl, M.L.; Liebermeister, E.; Scheurig, H.; Heinze, H.G. Analgetische bestrahlung degenerativentzündlicher skeletterkrankungen: Nutzen und risiko. Dtsch. Med. Wochenschr. 1993, 118, 493–498. [Google Scholar] [CrossRef]
- Lumniczky, K.; Impens, N.; Armengol, G.; Candéias, S.; Georgakilas, A.G.; Hornhardt, S.; Martin, O.A.; Rödel, F.; Schaue, D. Low dose ionizing radiation effects on the immune system. Environ. Int. 2021, 149, 106212. [Google Scholar] [CrossRef] [PubMed]
- Torres Royo, L.; Antelo Redondo, G.; Árquez Pianetta, M.; Arenas Prat, M. Low-Dose radiation therapy for benign pathologies. Rep. Pract. Oncol. Radiother. 2020, 25, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, G.; Maggiorella, L.; Rödel, F.; Rödel, V.; Willis, D.; Trott, K.-R. Mononuclear cell adhesion and cell adhesion molecule liberation after X-irradiation of activated endothelial cells in vitro. Int. J. Radiat. Biol. 2002, 78, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, G.; Seed, M.P.; Freemantle, C.N.; Alam, C.A.; Colville-Nash, P.R.; Trott, K.R. Mechanisms of the anti-inflammatory activity of low-dose radiation therapy. Int. J. Radiat. Biol. 1998, 74, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Schaue, D.; Marples, B.; Trott, K.R. The effects of low-dose X-irradiation on the oxidative burst in stimulated macrophages. Int. J. Radiat. Biol. 2002, 78, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Kern, P.M.; Keilholz, L.; Forster, C.; Hallmann, R.; Herrmann, M.; Seegenschmiedt, M.-H. Low-dose radiotherapy selectively reduces adhesion of peripheral blood mononuclear cells to endothelium in vitro. Radiother. Oncol. 2000, 54, 273–282. [Google Scholar] [CrossRef]
- Rödel, F.; Schaller, U.; Schultze-Mosgau, S.; Beuscher, H.-U.; Keilholz, L.; Herrmann, M.; Voll, R.; Sauer, R.; Hildebrandt, G. The induction of TGF-beta(1) and NF-kappaB parallels a biphasic time course of leukocyte/endothelial cell adhesion following low-dose X-irradiation. Strahlenther. Onkol. 2004, 180, 194–200. [Google Scholar] [CrossRef] [PubMed]
- Roedel, F.; Kley, N.; Beuscher, H.U.; Hildebrandt, G.; Keilholz, L.; Kern, P.; Voll, R.; Herrmann, M.; Sauer, R. Anti-inflammatory effect of low-dose X-irradiation and the involvement of a TGF-beta1-induced down-regulation of leukocyte/endothelial cell adhesion. Int. J. Radiat. Biol. 2002, 78, 711–719. [Google Scholar] [CrossRef]
- Conti, P.; DiGioacchino, M. MCP-1 and RANTES are mediators of acute and chronic inflammation. Allergy Asthma Proc. 2001, 22, 133–137. [Google Scholar] [CrossRef]
- Raines, E.W.; Ferri, N. Thematic review series: The immune system and atherogenesis. Cytokines affecting endothelial and smooth muscle cells in vascular disease. J. Lipid Res. 2005, 46, 1081–1092. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Yin, L.; Zhang, K.; Sun, W.; Yang, S.; Zhang, B.; Salzman, P.; Wang, W.; Liu, C.; Vidyasagar, S.; et al. Response patterns of cytokines/chemokines in two murine strains after irradiation. Cytokine 2012, 58, 169–177. [Google Scholar] [CrossRef]
- Schröder, S.; Juerß, D.; Kriesen, S.; Manda, K.; Hildebrandt, G. Immunomodulatory properties of low-dose ionizing radiation on human endothelial cells. Int. J. Radiat. Biol. 2019, 95, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Schröder, S.; Kriesen, S.; Paape, D.; Hildebrandt, G.; Manda, K. Modulation of inflammatory reactions by low-dose ionizing radiation: Cytokine release of murine endothelial cells is dependent on culture conditions. J. Immunol. Res. 2018, 2018, 2856518. [Google Scholar] [CrossRef] [PubMed]
- Cool, D.A. Review of the ICRP system of protection: The approach to existing exposure situations. Ann. ICRP 2015, 44, 179–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Association of Nuclear Medicine; European Federation of Organizations for Medical Physics; European Federation of Radiographer Societies; European Society of Radiology; European Society for Radiotherapy and Oncology. Common strategic research agenda for radiation protection in medicine. Insights Imaging 2017, 8, 183–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, F.A.; Akleyev, A.V.; Hauer-Jensen, M.; Hendry, J.H.; Kleiman, N.J.; MacVittie, T.J.; Aleman, B.M.; Edgar, A.B.; Mabuchi, K.; Muirhead, C.R.; et al. ICRP publication 118: ICRP statement on tissue reactions and early and late effects of radiation in normal tissues and organs—threshold doses for tissue reactions in a radiation protection context. Ann. ICRP 2012, 41, 1–322. [Google Scholar] [CrossRef]
- Harrison, J.D.; Balonov, M.; Bochud, F.; Martin, C.J.; Menzel, H.-G.; Smith-Bindman, R.; Ortiz-López, P.; Simmonds, J.R.; Wakeford, R. The use of dose quantities in radiological protection: ICRP publication 147 Ann ICRP 50(1) 2021. J. Radiol. Prot. 2021, 41, 410–422. [Google Scholar] [CrossRef] [PubMed]
- Grace, M.B.; McLeland, C.B.; Gagliardi, S.J.; Smith, J.M.; Jackson, W.E.; Blakely, W.F. Development and assessment of a quantitative reverse transcription-PCR assay for simultaneous measurement of four amplicons. Clin. Chem. 2003, 49, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Mathias, D.; Mitchel, R.E.J.; Barclay, M.; Wyatt, H.; Bugden, M.; Priest, N.D.; Whitman, S.C.; Scholz, M.; Hildebrandt, G.; Kamprad, M.; et al. Low-dose irradiation affects expression of inflammatory markers in the heart of ApoE −/− mice. PLoS ONE 2015, 10, e0119661. [Google Scholar] [CrossRef] [Green Version]
- Mahley, R.W.; Rall, S.C. Apolipoprotein E: Far more than a lipid transport protein. Annu. Rev. Genom. Hum. Genet. 2000, 1, 507–537. [Google Scholar] [CrossRef]
- Zhang, H.; Wu, J.; Zhu, J. The role of apolipoprotein E in Guillain-Barré syndrome and experimental autoimmune neuritis. J. Biomed. Biotechnol. 2010, 2010, 357412. [Google Scholar] [CrossRef] [Green Version]
- Libby, P. Inflammation in atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2045–2051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banda, M.; Bommineni, A.; Thomas, R.A.; Luckinbill, L.S.; Tucker, J.D. Evaluation and validation of housekeeping genes in response to ionizing radiation and chemical exposure for normalizing RNA expression in real-time PCR. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2008, 649, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Bijwaard, K.E.; Aguilera, N.S.I.; Monczak, Y.; Trudel, M.; Taubenberger, J.K.; Lichy, J.H. Quantitative real-time reverse transcription-PCR assay for cyclin D1 expression: Utility in the diagnosis of mantle cell lymphoma. Clin. Chem. 2001, 47, 195–201. [Google Scholar] [CrossRef]
- Colotta, F.; Allavena, P.; Sica, A.; Garlanda, C.; Mantovani, A. Cancer-related inflammation, the seventh hallmark of cancer: Links to genetic instability. Carcinogenesis 2009, 30, 1073–1081. [Google Scholar] [CrossRef] [Green Version]
- Garlanda, C.; Parravicini, C.; Sironi, M.; de Rossi, M.; Wainstok de Calmanovici, R.; Carozzi, F.; Bussolino, F.; Colotta, F.; Mantovani, A.; Vecchi, A. Progressive growth in immunodeficient mice and host cell recruitment by mouse endothelial cells transformed by polyoma middle-sized T antigen: Implications for the pathogenesis of opportunistic vascular tumors. Proc. Natl. Acad. Sci. USA 1994, 91, 7291–7295. [Google Scholar] [CrossRef] [Green Version]
- Sorokin, L.; Girg, W.; Göpfert, T.; Hallmann, R.; Deutzmann, R. Expression of novel 400-kDa laminin chains by mouse and bovine endothelial cells. Eur. J. Biochem. 1994, 223, 603–610. [Google Scholar] [CrossRef]
- Patties, I.; Haagen, J.; Dörr, W.; Hildebrandt, G.; Glasow, A. Late inflammatory and thrombotic changes in irradiated hearts of C57BL/6 wild-type and atherosclerosis-prone ApoE-deficient mice. Strahlenther. Onkol. 2015, 191, 172–179. [Google Scholar] [CrossRef]
- Patties, I.; Habelt, B.; Rosin, B.; Dörr, W.; Hildebrandt, G.; Glasow, A. Late effects of local irradiation on the expression of inflammatory markers in the Arteria saphena of C57BL/6 wild-type and ApoE-knockout mice. Radiat. Environ. Biophys. 2014, 53, 117–124. [Google Scholar] [CrossRef]
- Baselet, B.; Sonveaux, P.; Baatout, S.; Aerts, A. Pathological effects of ionizing radiation: Endothelial activation and dysfunction. Cell. Mol. Life Sci. 2019, 76, 699–728. [Google Scholar] [CrossRef] [Green Version]
- Boerma, M.; Sridharan, V.; Mao, X.-W.; Nelson, G.A.; Cheema, A.K.; Koturbash, I.; Singh, S.P.; Tackett, A.J.; Hauer-Jensen, M. Effects of ionizing radiation on the heart. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2016, 770, 319–327. [Google Scholar] [CrossRef] [Green Version]
- Sievert, W.; Trott, K.-R.; Azimzadeh, O.; Tapio, S.; Zitzelsberger, H.; Multhoff, G. Late proliferating and inflammatory effects on murine microvascular heart and lung endothelial cells after irradiation. Radiother. Oncol. 2015, 117, 376–381. [Google Scholar] [CrossRef]
- Monceau, V.; Meziani, L.; Strup-Perrot, C.; Morel, E.; Schmidt, M.; Haagen, J.; Escoubet, B.; Dörr, W.; Vozenin, M.-C. Enhanced sensitivity to low dose irradiation of ApoE−/− mice mediated by early pro-inflammatory profile and delayed activation of the TGFβ1 cascade involved in fibrogenesis. PLoS ONE 2013, 8, e57052. [Google Scholar] [CrossRef] [Green Version]
- Koturbash, I.; Merrifield, M.; Kovalchuk, O. Fractionated exposure to low doses of ionizing radiation results in accumulation of DNA damage in mouse spleen tissue and activation of apoptosis in a p53/Atm-independent manner. Int. J. Radiat. Biol. 2017, 93, 148–155. [Google Scholar] [CrossRef]
- Takahashi, A.; Ohnishi, K.; Asakawa, I.; Kondo, N.; Nakagawa, H.; Yonezawa, M.; Tachibana, A.; Matsumoto, H.; Ohnishi, T. Radiation response of apoptosis in C57BL/6N mouse spleen after whole-body irradiation. Int. J. Radiat. Biol. 2001, 77, 939–945. [Google Scholar] [CrossRef]
- Paul, A.M.; Overbey, E.G.; da Silveira, W.A.; Szewczyk, N.; Nishiyama, N.C.; Pecaut, M.J.; Anand, S.; Galazka, J.M.; Mao, X.W. Immunological and hematological outcomes following protracted low dose/low dose rate ionizing radiation and simulated microgravity. Sci. Rep. 2021, 11, 11452. [Google Scholar] [CrossRef]
- Lin, I.H.; Hau, D.M.; Chen, W.C.; Chen, K.T. Effects of low dose gamma-ray irradiation on peripheral leukocyte counts and spleen of mice. Chin. Med. J. (Engl.) 1996, 109, 210–214. [Google Scholar]
- de la Pinta, C.; Fernández Lizarbe, E.; Montero Luis, Á.; Domínguez Rullán, J.A.; Sancho García, S. Treatment of symptomatic splenomegaly with low doses of radiotherapy: Retrospective analysis and review of the literature. Tech. Innov. Patient Support Radiat. Oncol. 2017, 3-4, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Alexandru, M.; Rodica, A.; Dragos-Eugen, G.; Mihai-Teodor, G. Assessing the spleen as an organ at risk in radiation therapy and its relationship with radiation-induced lymphopenia: A retrospective study and literature review. Adv. Radiat. Oncol. 2021, 100761. [Google Scholar] [CrossRef]
- Bogdándi, E.N.; Balogh, A.; Felgyinszki, N.; Szatmári, T.; Persa, E.; Hildebrandt, G.; Sáfrány, G.; Lumniczky, K. Effects of low-dose radiation on the immune system of mice after total-body irradiation. Radiat. Res. 2010, 174, 480–489. [Google Scholar] [CrossRef]
- DeBo, R.J.; Register, T.C.; Caudell, D.L.; Sempowski, G.D.; Dugan, G.; Gray, S.; Owzar, K.; Jiang, C.; Bourland, J.D.; Chao, N.J.; et al. Molecular and cellular profiling of acute responses to total body radiation exposure in ovariectomized female cynomolgus macaques. Int. J. Radiat. Biol. 2015, 91, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Giddings, J.C. Soluble adhesion molecules in inflammatory and vascular diseases. Biochem. Soc. Trans. 2005, 33, 406–408. [Google Scholar] [CrossRef] [PubMed]
- Ishii, Y.; Kitamura, S. Soluble intercellular adhesion molecule-1 as an early detection marker for radiation pneumonitis. Eur. Respir. J. 1999, 13, 733–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, F.A.; Hoving, S.; Russell, N.S. Vascular damage as an underlying mechanism of cardiac and cerebral toxicity in irradiated cancer patients. Radiat. Res. 2010, 174, 865–869. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, R.; Claessens, M.; Cocquyt, E.; Mysara, M.; Decrock, E.; Baatout, S.; Aerts, A.; Leybaert, L. X-irradiation induces acute and early term inflammatory responses in atherosclerosis-prone ApoE-/- mice and in endothelial cells. Mol. Med. Rep. 2021, 23, 399. [Google Scholar] [CrossRef]
- Mitchel, R.E.J.; Hasu, M.; Bugden, M.; Wyatt, H.; Little, M.P.; Gola, A.; Hildebrandt, G.; Priest, N.D.; Whitman, S.C. Low-dose radiation exposure and atherosclerosis in ApoE⁻/⁻ mice. Radiat. Res. 2011, 175, 665–676. [Google Scholar] [CrossRef] [Green Version]
- Gaugler, M.H.; Squiban, C.; van der Meeren, A.; Bertho, J.M.; Vandamme, M.; Mouthon, M.A. Late and persistent up-regulation of intercellular adhesion molecule-1 (ICAM-1) expression by ionizing radiation in human endothelial cells in vitro. Int. J. Radiat. Biol. 1997, 72, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Jensen, T.; Kierulf, P.; Sandset, P.M.; Klingenberg, O.; Joø, G.B.; Godal, H.C.; Skjønsberg, O.H. Fibrinogen and fibrin induce synthesis of proinflammatory cytokines from isolated peripheral blood mononuclear cells. Thromb. Haemost. 2007, 97, 822–829. [Google Scholar] [CrossRef]
- Koenig, W. Fibrin(ogen) in cardiovascular disease: An update. Thromb. Haemost. 2003, 89, 601–609. [Google Scholar] [CrossRef]
- Mackiewicz, A.; Ganapathi, M.K.; Schultz, D.; Brabenec, A.; Weinstein, J.; Kelley, M.F.; Kushner, I. Transforming growth factor beta 1 regulates production of acute-phase proteins. Proc. Natl. Acad. Sci. USA 1990, 87, 1491–1495. [Google Scholar] [CrossRef] [Green Version]
- Richter, V.; Rassoul, F.; Purschwitz, K.; Hentschel, B.; Reuter, W.; Kuntze, T. Circulating vascular cell adhesion molecules VCAM-1, ICAM-1, and E-selectin in dependence on aging. Gerontology 2003, 49, 293–300. [Google Scholar] [CrossRef]
- Ren, H.-Y.; Khera, A.; de Lemos, J.A.; Ayers, C.R.; Rohatgi, A. Soluble endothelial cell-selective adhesion molecule and incident cardiovascular events in a multiethnic population. Am. Heart J. 2017, 191, 55–61. [Google Scholar] [CrossRef]
- Miles, E.A.; Rees, D.; Banerjee, T.; Cazzola, R.; Lewis, S.; Wood, R.; Oates, R.; Tallant, A.; Cestaro, B.; Yaqoob, P.; et al. Age-related increases in circulating inflammatory markers in men are independent of BMI, blood pressure and blood lipid concentrations. Atherosclerosis 2008, 196, 298–305. [Google Scholar] [CrossRef]
- Cominacini, L.; Garbin, U.; Pasini, A.F.; Davoli, A.; Campagnola, M.; Contessi, G.B.; Pastorino, A.M.; Lo Cascio, V. Antioxidants inhibit the expression of intercellular cell adhesion molecule-1 and vascular cell adhesion molecule-1 induced by oxidized LDL on human umbilical vein endothelial cells. Free Radic. Biol. Med. 1997, 22, 117–127. [Google Scholar] [CrossRef]
- Schröder, S.; Broese, S.; Baake, J.; Juerß, D.; Kriesen, S.; Hildebrandt, G.; Manda, K. Effect of ionizing radiation on human EA.hy926 endothelial cells under inflammatory conditions and their interactions with A549 tumour cells. J. Immunol. Res. 2019, 2019, 9645481. [Google Scholar] [CrossRef] [Green Version]
- Meeren, A.V.; Bertho, J.M.; Vandamme, M.; Gaugler, M.H. Ionizing radiation enhances IL-6 and IL-8 production by human endothelial cells. Mediat. Inflamm. 1997, 6, 185–193. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wu, L.-M.; Wu, J. Cross-talk between apolipoprotein E and cytokines. Mediat. Inflamm. 2011, 2011, 949072. [Google Scholar] [CrossRef] [Green Version]
- Gerhardt, S.; König, V.; Doll, M.; Hailemariam-Jahn, T.; Hrgovic, I.; Zöller, N.; Kaufmann, R.; Kippenberger, S.; Meissner, M. Dimethylfumarate protects against TNF-α-induced secretion of inflammatory cytokines in human endothelial cells. J. Inflamm. 2015, 12, 49. [Google Scholar] [CrossRef] [Green Version]
- Brown, Z.; Gerritsen, M.E.; Carley, W.W.; Strieter, R.M.; Kunkel, S.L.; Westwick, J. Chemokine gene expression and secretion by cytokine-activated human microvascular endothelial cells. Differential regulation of monocyte chemoattractant protein-1 and interleukin-8 in response to interferon-gamma. Am. J. Pathol. 1994, 145, 913–921. [Google Scholar]
- Cervelli, T.; Panetta, D.; Navarra, T.; Andreassi, M.G.; Basta, G.; Galli, A.; Salvadori, P.A.; Picano, E.; Del Turco, S. Effects of single and fractionated low-dose irradiation on vascular endothelial cells. Atherosclerosis 2014, 235, 510–518. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene/Product Size | Forward Primer 5′-3′ | Probe 5′-3′ | Reversed Primer 5′-3′ |
|---|---|---|---|
| mCD68/73 bp NM_001291058 | CTCAGCTGCCTGACAAGGGA | FAM-TCGGGCCATGTTTCTCTTGCAACCGT-BHQ1 | AGAGGCAGCAAGAGGGACTG |
| mKi67/99 bp NM_001081117 | AGCAGACGAGCAAGAGACAA | FAM.-CCCAGCACTCCAAAGAAACCCAC-BHQ1 | TACAGGGAGAGTTTGCATGG |
| mB2m/91 bp [33] NM_009735 | TGAGACTGATACATACGCCTGCA | HEX-ATGGCCGAGCCCAAGACCGTC-BHQ1 | GATGCTTGATCACATGTCTCGATC |
| m18S/102 bp NR_003278 | AGGAATTCCCAGTAAGTGCG | HEX-TCCCTGCCCTTTGTACACACCGCC-BHQ1 | GCCTCACTAAACCATCCAA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glasow, A.; Patties, I.; Priest, N.D.; Mitchel, R.E.J.; Hildebrandt, G.; Manda, K. Dose and Dose Rate-Dependent Effects of Low-Dose Irradiation on Inflammatory Parameters in ApoE-Deficient and Wild Type Mice. Cells 2021, 10, 3251. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113251
Glasow A, Patties I, Priest ND, Mitchel REJ, Hildebrandt G, Manda K. Dose and Dose Rate-Dependent Effects of Low-Dose Irradiation on Inflammatory Parameters in ApoE-Deficient and Wild Type Mice. Cells. 2021; 10(11):3251. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113251
Chicago/Turabian StyleGlasow, Annegret, Ina Patties, Nicholas D. Priest, Ronald E. J. Mitchel, Guido Hildebrandt, and Katrin Manda. 2021. "Dose and Dose Rate-Dependent Effects of Low-Dose Irradiation on Inflammatory Parameters in ApoE-Deficient and Wild Type Mice" Cells 10, no. 11: 3251. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113251