
Microglia-Based Sex-Biased Neuropathology in Early-Stage Alzheimer’s Disease Model Mice and the Potential Pharmacologic Efficacy of Dioscin

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Drug Administration
2.2. Tissue Preparation
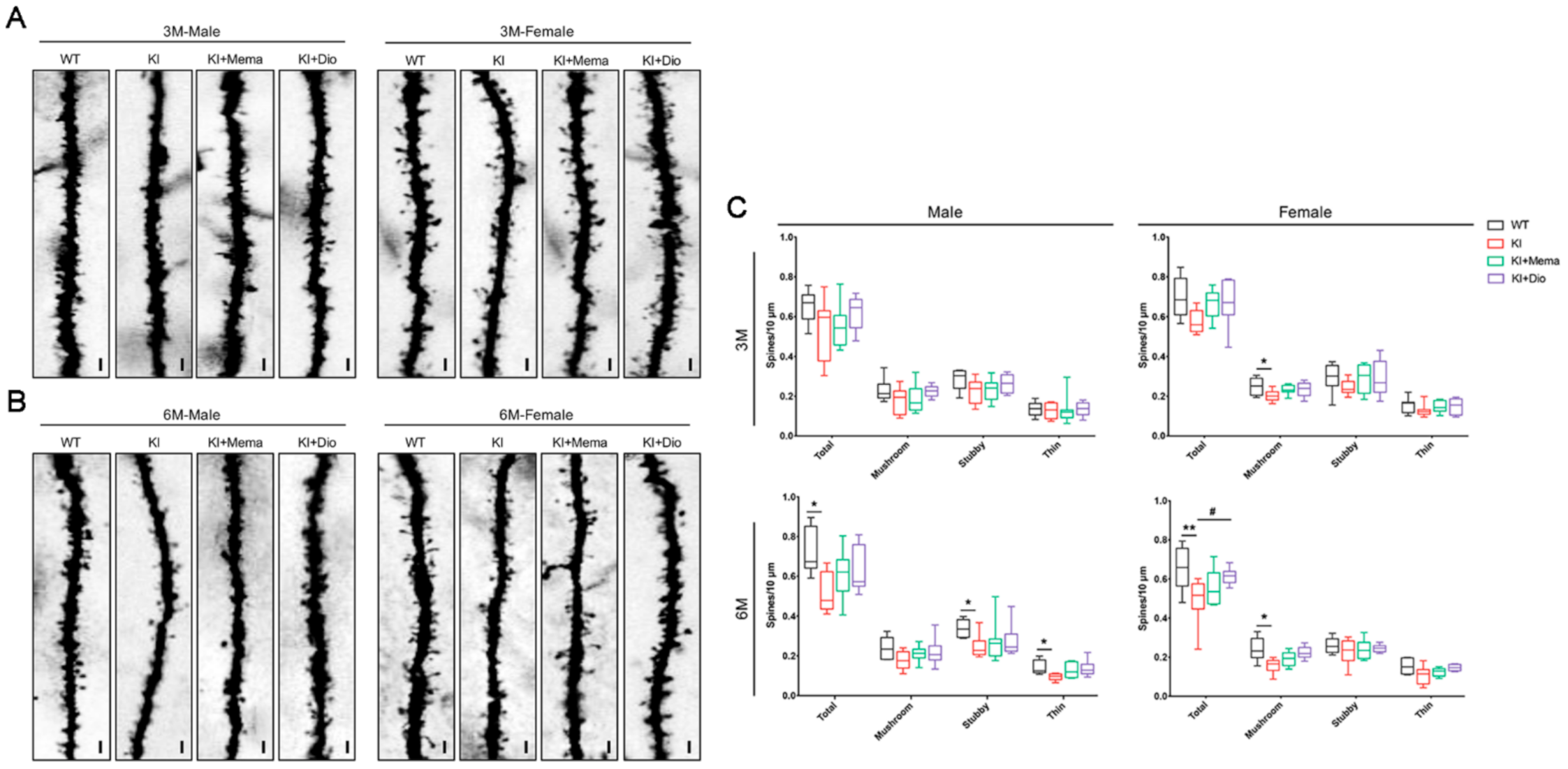
2.3. Golgi Staining and Dendritic Spine Analysis
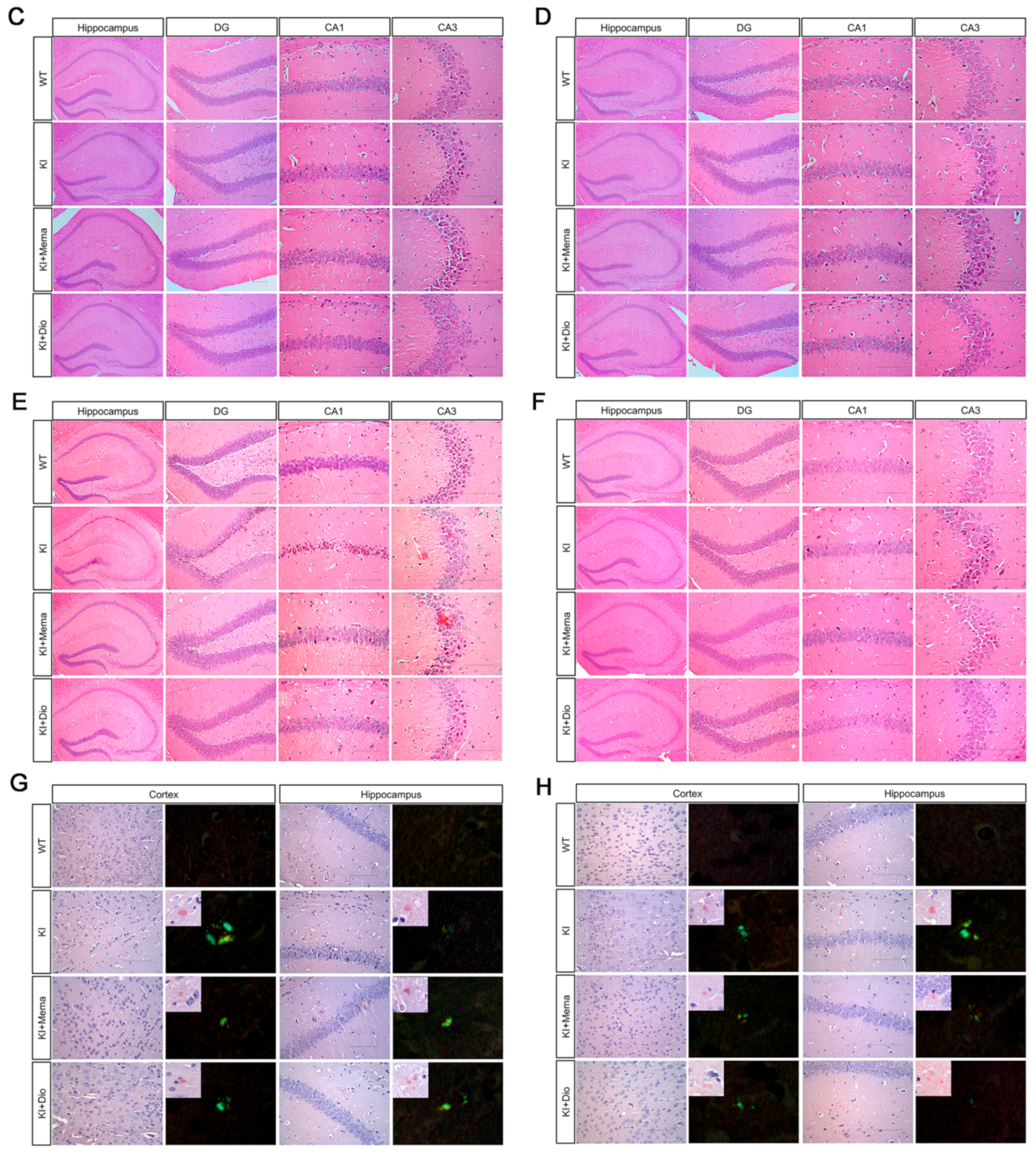
2.4. Histologic Observations
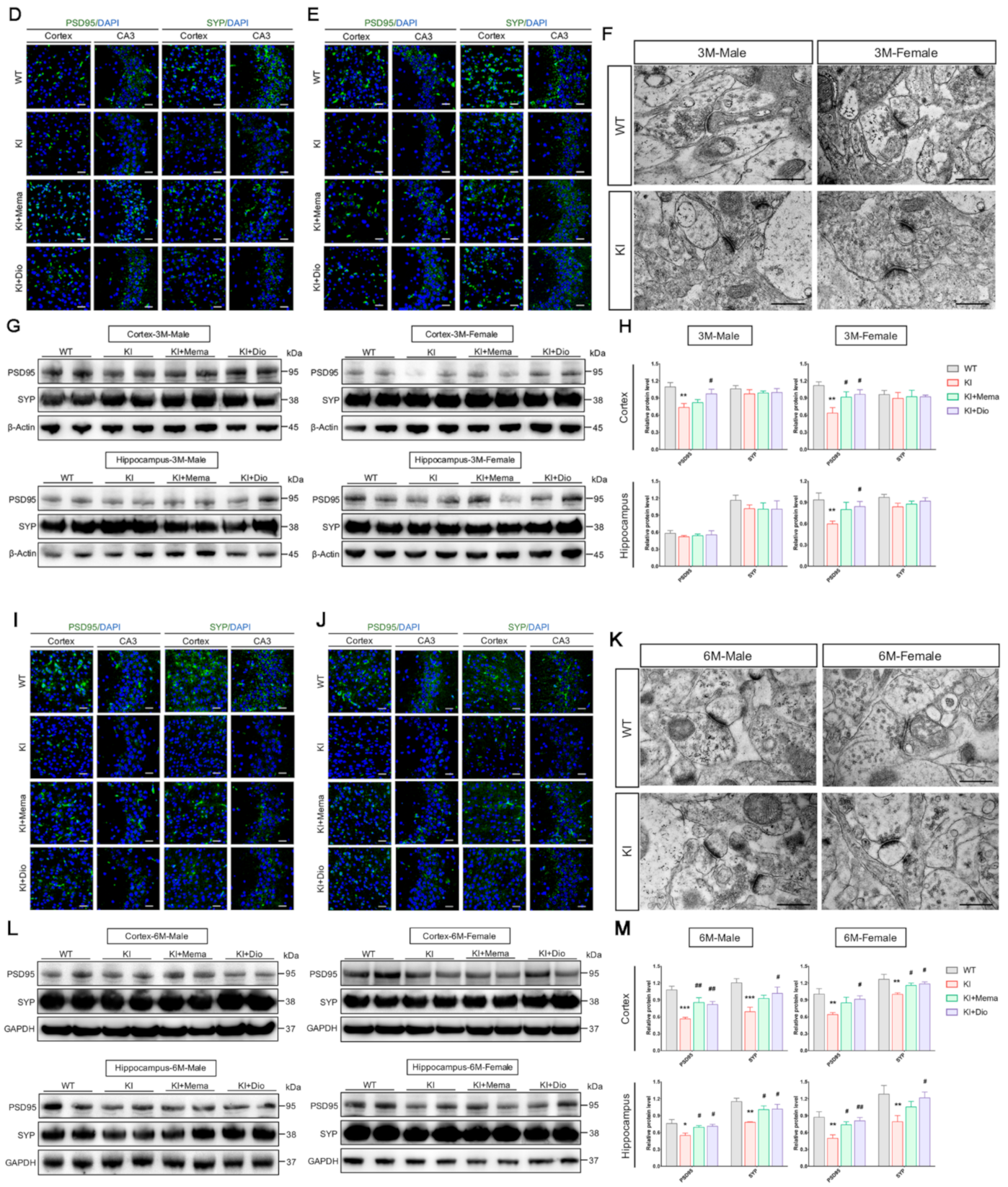
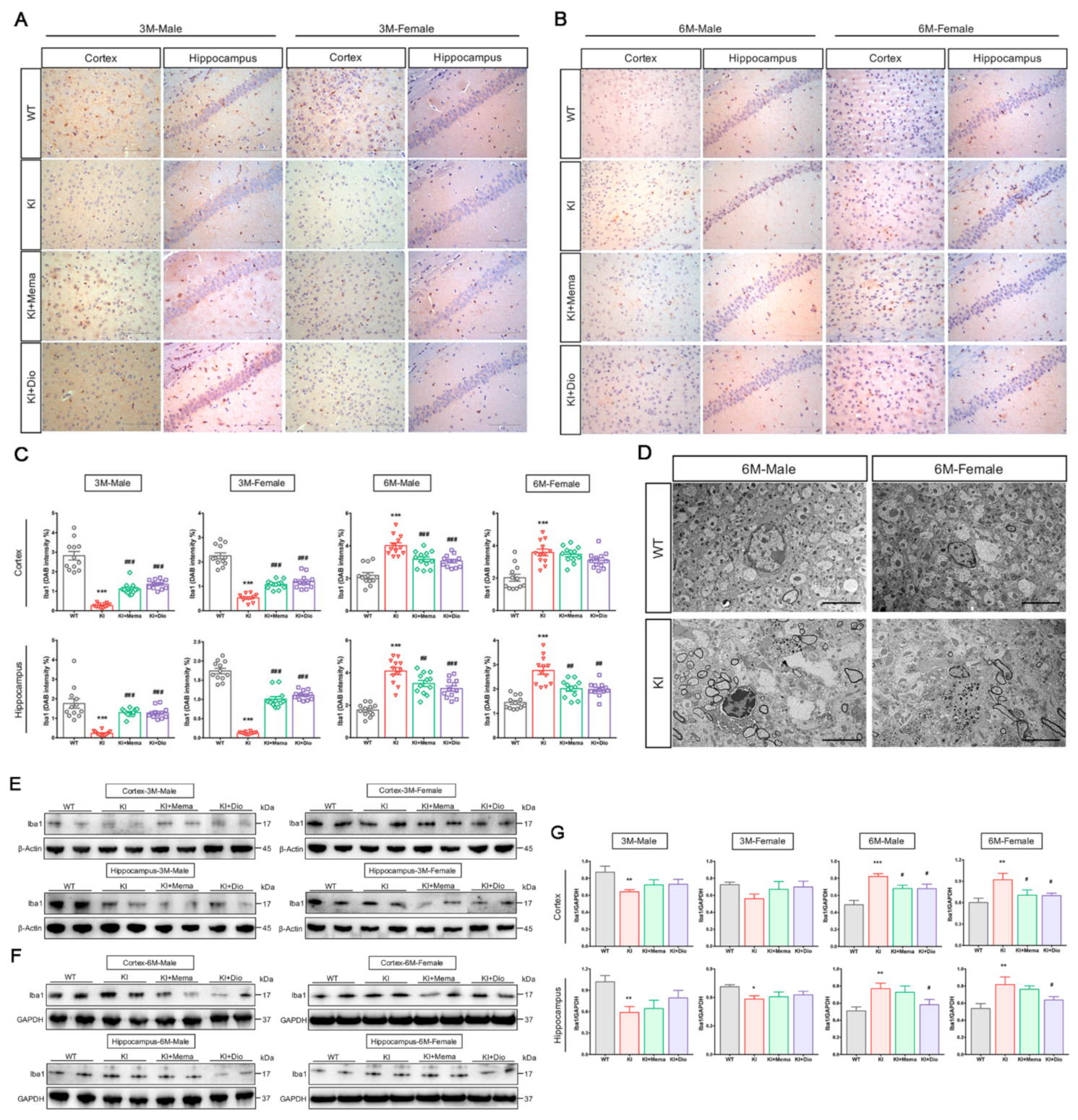
2.5. Transmission Electron Microscopic Observations
2.6. Immunohistochemistry
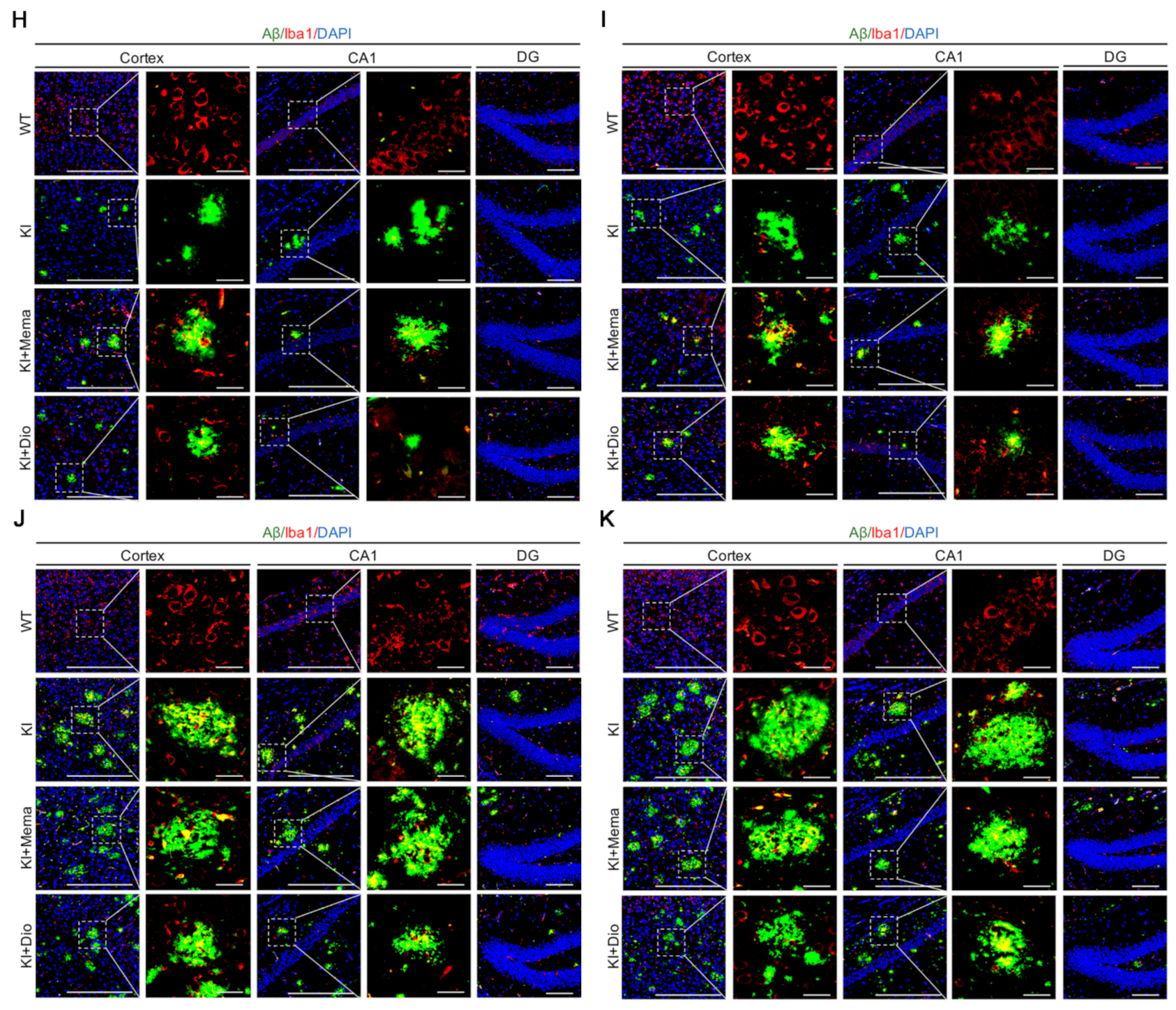
2.7. Immunofluorescence
2.8. Western Blot Analyses
2.9. ELISA Assay
2.10. Statistical Analyses
3. Results
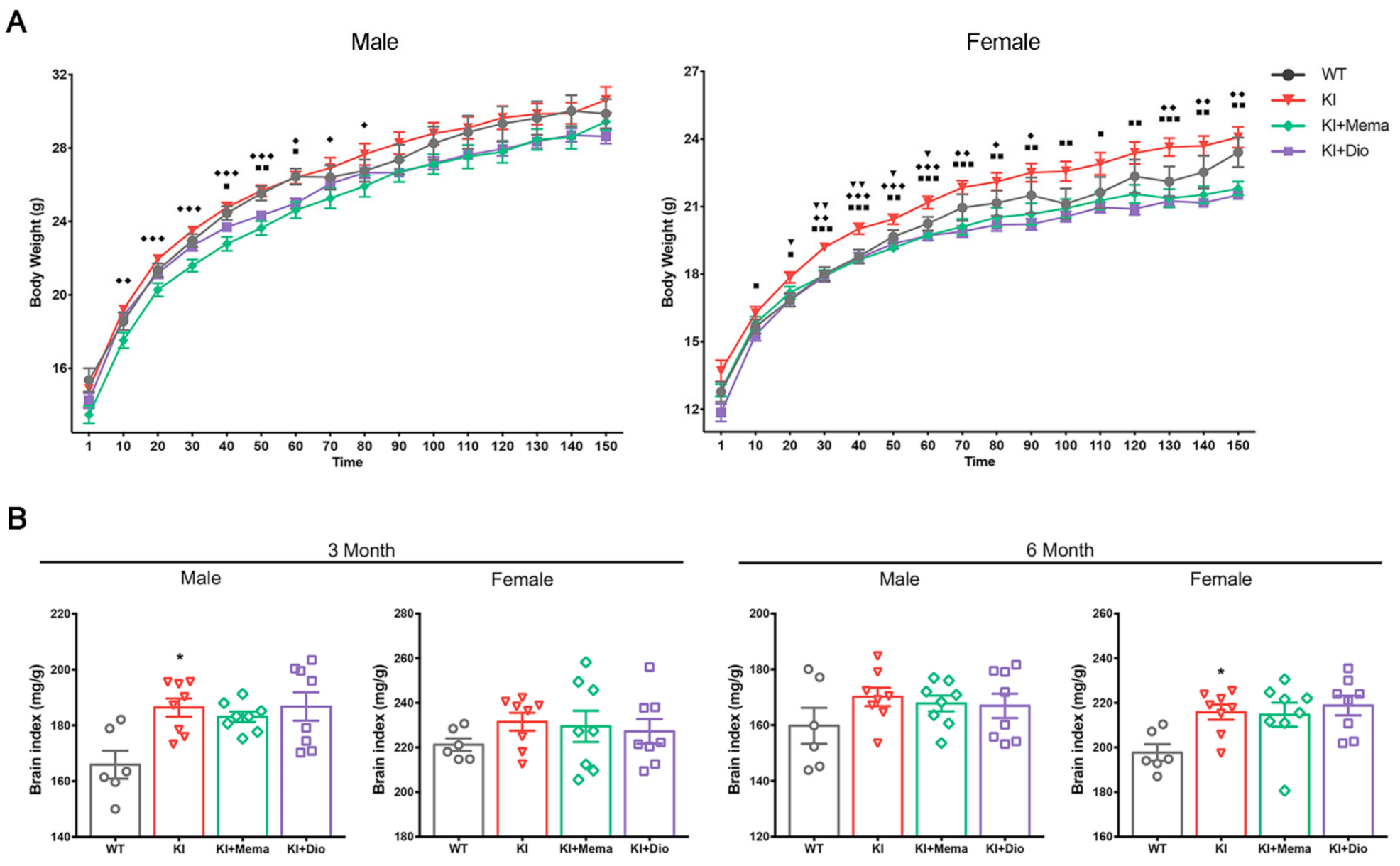
3.1. Pathologic Changes in AppNL-G-F Mice and Dio Treatment
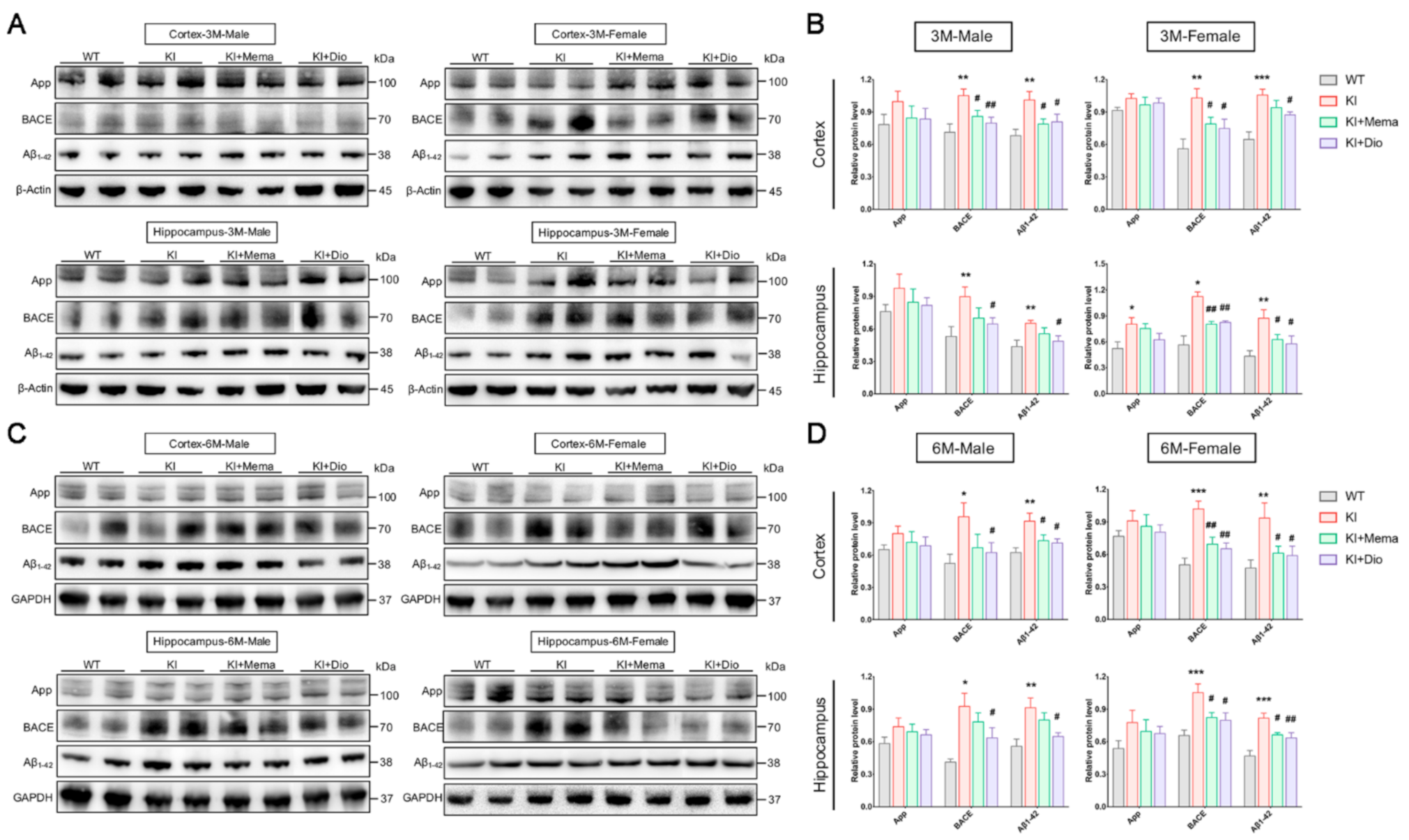
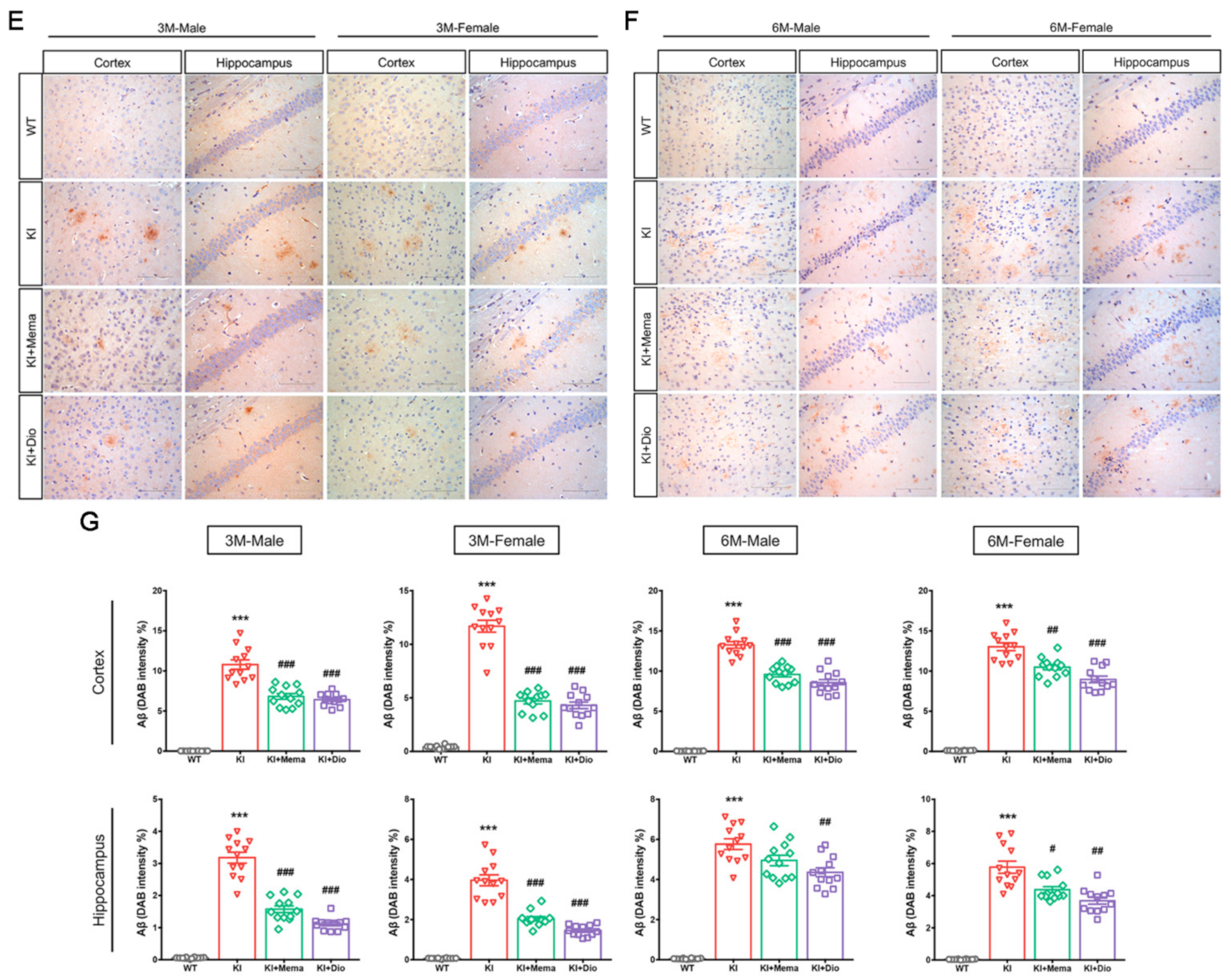
3.2. Dio Inhibits the Abnormal Accumulation of Aβ Precursors and Alleviates Aβ Burden in Early-Stage AD Model Mice
3.3. Dio Alters Synaptic Dysfunction in Young Adult AppNL-G-F Mice
3.4. Dio Affects Microglial Activation in Young Adult Appnl-G-F Mice
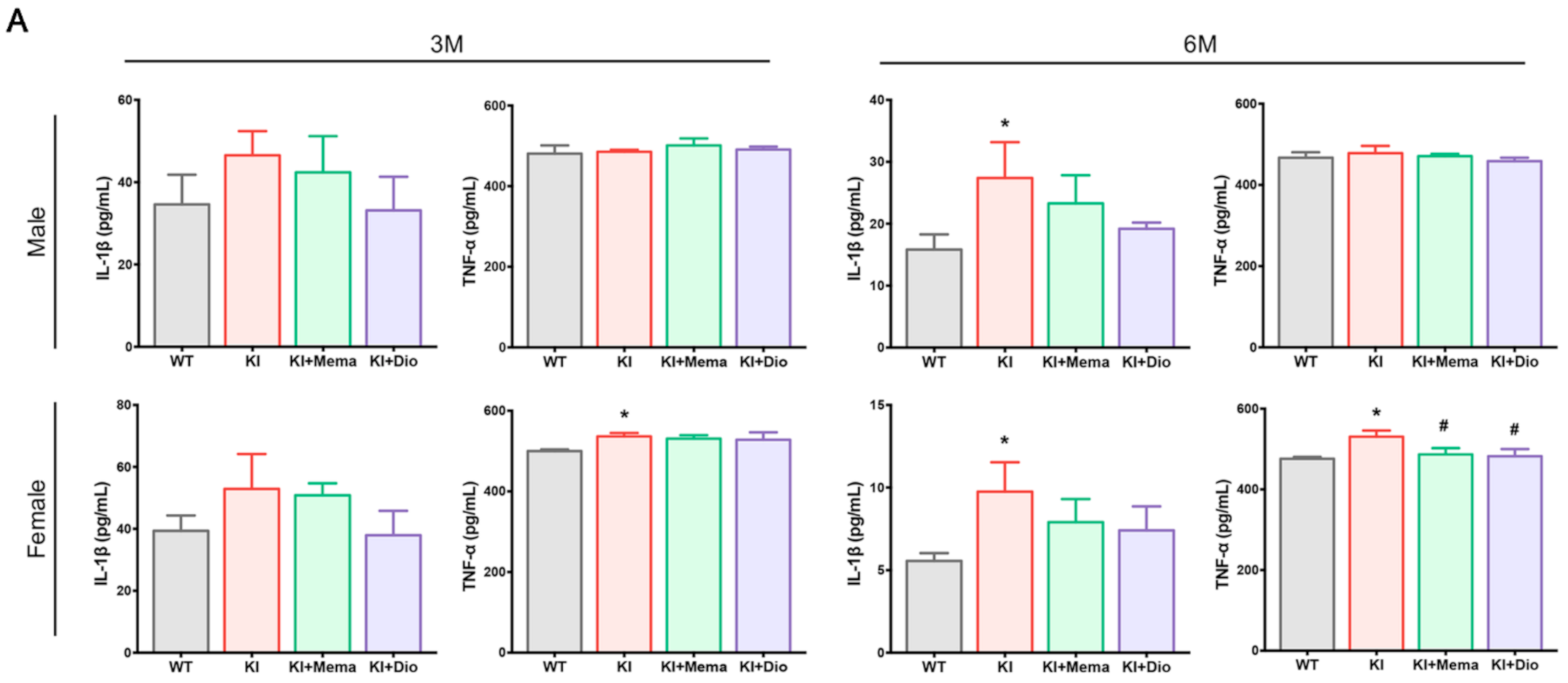
3.5. Dio Regulates Neuroinflammatory Responses in Young Adult Appnl-G-F Mouse Brain by Affecting NF-Κb/Β-Catenin
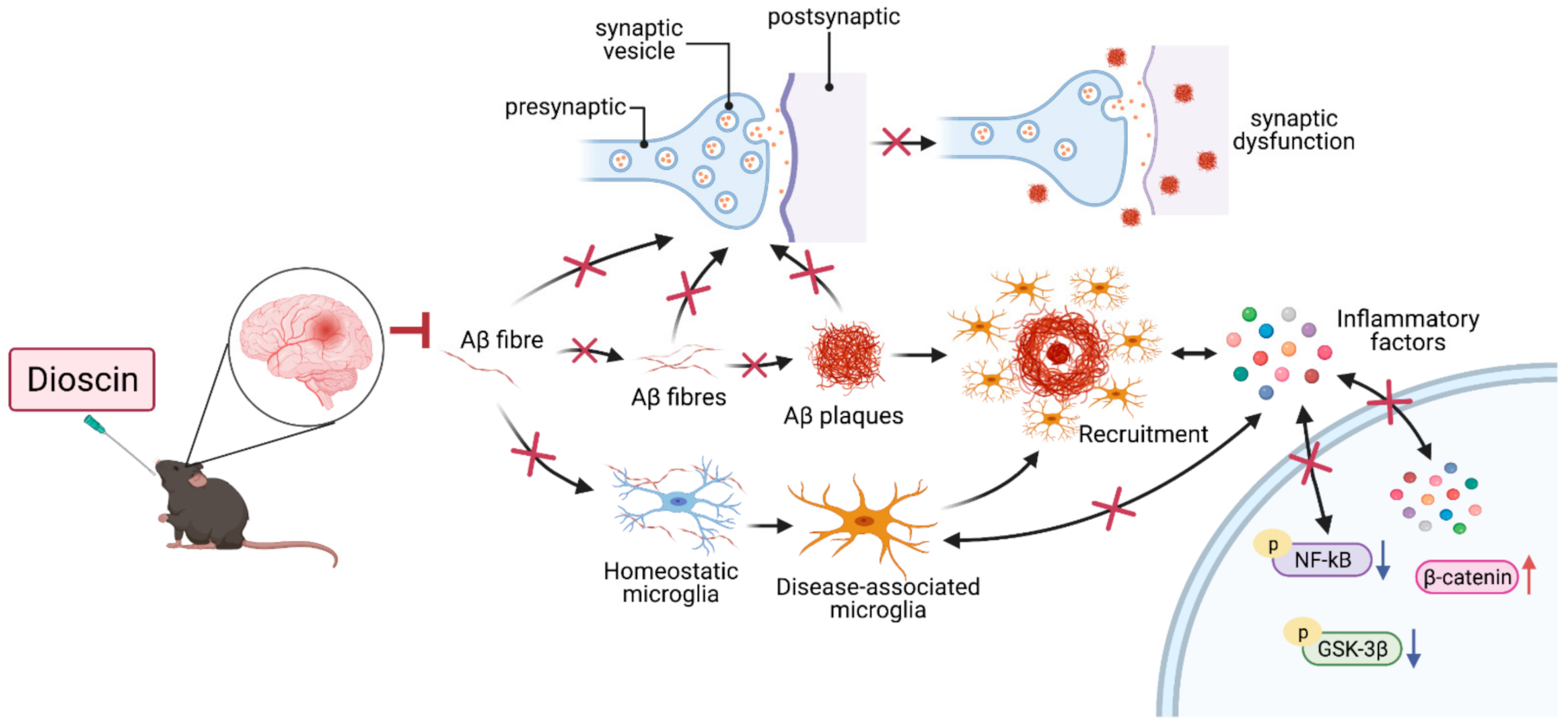
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hodson, R. Alzheimer’s disease. Nat. Cell Biol. 2018, 559, S1. [Google Scholar] [CrossRef] [Green Version]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. Nia-Aa Research Framework: Toward a Biological Definition of Alzheimer’s Disease. Alzheimers Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Calsolaro, V.; Edison, P. Neuroinflammation in Alzheimer’s Disease: Current Evidence and Future directions. Alzheimers Dement. 2016, 12, 719–732. [Google Scholar] [CrossRef]
- Sasaguri, H.; Nilsson, P.; Hashimoto, S.; Nagata, K.; Saito, T.; De Strooper, B.; Hardy, J.; Vassar, R.; Winblad, B.; Saido, T.C. App Mouse Models for Alzheimer’s Disease Preclinical Studies. Embo J. 2017, 36, 2473–2487. [Google Scholar] [CrossRef] [PubMed]
- Sevigny, J.; Chiao, P.; Bussière, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef]
- Hardy, J.; Selkoe, D.J. The Amyloid Hypothesis of Alzheimer’s Disease: Progress and Problems on the Road to Therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.C.; Zhao, N.; Yamaguchi, Y.; Cirrito, J.R.; Kanekiyo, T.; Holtzman, D.M.; Bu, G. Neuronal Heparan Sulfates Promote Amyloid Pathology by Modulating Brain Amyloid-β Clearance and Aggregation in Alzheimer’s Disease. Sci. Transl. Med. 2016, 8, 332ra44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mucke, L.; Selkoe, D.J. Neurotoxicity of Amyloid -Protein: Synaptic and Network Dysfunction. Cold Spring Harb. Perspect. Med. 2012, 2, a006338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mawuenyega, K.G.; Sigurdson, W.; Ovod, V.; Munsell, L.; Kasten, T.; Morris, J.C.; Yarasheski, K.E.; Bateman, R.J. Decreased Clearance of CNS β-Amyloid in Alzheimer’s Disease. Science 2010, 330, 1774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busche, M.A.; Hyman, B.T. Synergy between amyloid-β and tau in Alzheimer’s disease. Nat. Neurosci. 2020, 23, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Jonsson, T.; Atwal, J.K.; Steinberg, S.; Snaedal, J.; Jonsson, P.V.; Bjornsson, S.; Stefansson, H.; Sulem, P.; Gudbjartsson, D.; Maloney, J.; et al. A Mutation in App Protects against Alzheimer’s Disease and Age-Related Cognitive Decline. Nature 2012, 488, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, K.; Chapman, P.; Nilsen, S.; Eckman, C.; Harigaya, Y.; Younkin, S.; Yang, F.; Cole, G. Correlative Memory Deficits, Abeta Elevation, and Amyloid Plaques in Transgenic Mice. Science 1996, 274, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Sacher, C.; Blume, T.; Beyer, L.; Peters, F.; Eckenweber, F.; Sgobio, C.; Deussing, M.; Albert, N.L.; Unterrainer, M.; Lindner, S.; et al. Longitudinal Pet Monitoring of Amyloidosis and Microglial Activation in a Second-Generation Amyloid-Β Mouse Model. J. Nucl. Med. 2019, 60, 1787–1793. [Google Scholar] [CrossRef] [PubMed]
- Saito, T.; Matsuba, Y.; Mihira, N.; Takano, J.; Nilsson, P.; Itohara, S.; Iwata, N.; Saido, T.C. Single App Knock-in Mouse Models of Alzheimer’s Disease. Nat. Neurosci. 2014, 17, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Masuda, A.; Kobayashi, Y.; Kogo, N.; Saito, T.; Saido, T.C.; Itohara, S. Cognitive Deficits in Single App Knock-in Mouse Models. Neurobiol. Learn Mem. 2016, 135, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, Y.; Sekiya, M.; Saito, T.; Saido, T.C.; Iijima, K.M. Cognitive and Emotional Alterations in App Knock-in Mouse Models of Aβ Amyloidosis. BMC Neurosci. 2018, 19, 46. [Google Scholar] [CrossRef] [PubMed]
- Castillo, E.; Leon, J.; Mazzei, G.; Abolhassani, N.; Haruyama, N.; Saito, T.; Saido, T.; Hokama, M.; Iwaki, T.; Ohara, T.; et al. Comparative Profiling of Cortical Gene Expression in Alzheimer’s Disease Patients and Mouse Models Demonstrates a Link between Amyloidosis and Neuroinflammation. Sci. Rep. 2017, 7, 17762. [Google Scholar] [CrossRef] [Green Version]
- Sala Frigerio, C.; Wolfs, L.; Fattorelli, N.; Thrupp, N.; Voytyuk, I.; Schmidt, I.; Mancuso, R.; Chen, W.T.; Woodbury, M.E.; Srivastava, G.; et al. The Major Risk Factors for Alzheimer’s Disease: Age, Sex, and Genes Modulate the Microglia Response to Aβ Plaques. Cell Rep. 2019, 27, 1293–1306. [Google Scholar] [CrossRef] [Green Version]
- Latif-Hernandez, A.; Shah, D.; Craessaerts, K.; Saido, T.; Saito, T.; De Strooper, B.; Van der Linden, A.; D’Hooge, R. Subtle behavioral changes and increased prefrontal-hippocampal network synchronicity in APPNL−G−F mice before prominent plaque deposition. Behav. Brain Res. 2019, 364, 431–441. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wu, L.; Pchitskaya, E.; Zakharova, O.; Saito, T.; Saido, T.; Bezprozvanny, I. Neuronal Store-Operated Calcium Entry and Mushroom Spine Loss in Amyloid Precursor Protein Knock-in Mouse Model of Alzheimer’s Disease. J. Neurosci. 2015, 35, 13275–13286. [Google Scholar] [CrossRef] [Green Version]
- Cai, B.; Zhang, Y.; Wang, Z.; Xu, D.; Jia, Y.; Guan, Y.; Liao, A.; Liu, G.; Chun, C.; Li, J. Therapeutic Potential of Diosgenin and Its Major Derivatives against Neurological Diseases: Recent Advances. Oxid. Med. Cell Longev. 2020, 2020, 3153082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sethi, G.; Shanmugam, M.K.; Warrier, S.; Merarchi, M.; Arfuso, F.; Kumar, A.P.; Bishayee, A. Pro-Apoptotic and Anti-Cancer Properties of Diosgenin: A Comprehensive and Critical Review. Nutrients 2018, 10, 645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, F.C.; Jiang, J.G. Effects of Diosgenin and Its Derivatives on Atherosclerosis. Food Funct. 2019, 10, 7022–7036. [Google Scholar] [CrossRef] [PubMed]
- Gan, Q.; Wang, J.; Hu, J.; Lou, G.; Xiong, H.; Peng, C.; Zheng, S.; Huang, Q. The role of diosgenin in diabetes and diabetic complications. J. Steroid Biochem. Mol. Biol. 2020, 198, 105575. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.-X.; Huang, Y.; Zheng, L.-L.; Zhang, L.; Su, L.; Wu, Y.-H.; Li, J.; Zhou, L.-C.; Huang, J.; Tang, Y.; et al. Design, synthesis and evaluation of diosgenin carbamate derivatives as multitarget anti-Alzheimer’s disease agents. Eur. J. Med. Chem. 2020, 187, 111913. [Google Scholar] [CrossRef]
- Tohda, C.; Lee, Y.A.; Goto, Y.; Nemere, I. Diosgenin-Induced Cognitive Enhancement in Normal Mice Is Mediated by 1,25d₃-Marrs. Sci. Rep. 2013, 3, 3395. [Google Scholar] [CrossRef] [Green Version]
- Mehla, J.; Lacoursiere, S.G.; Lapointe, V.; McNaughton, B.L.; Sutherland, R.J.; McDonald, R.J.; Mohajerani, M.H. Age-dependent behavioral and biochemical characterization of single APP knock-in mouse (APPNL-G-F/NL-G-F) model of Alzheimer’s disease. Neurobiol. Aging 2019, 75, 25–37. [Google Scholar] [CrossRef]
- Danysz, W.; Parsons, C.G. Alzheimer’s Disease, Β-Amyloid, Glutamate, Nmda Receptors and Memantine--Searching for the Connections. Br. J. Pharmacol. 2012, 167, 324–352. [Google Scholar] [CrossRef] [Green Version]
- Whyte, L.S.; Hemsley, K.M.; Lau, A.A.; Hassiotis, S.; Saito, T.; Saido, T.C.; Hopwood, J.J.; Sargeant, T.J. Reduction in Open Field Activity in the Absence of Memory Deficits in the App Nl-G-F Knock-in Mouse Model of Alzheimer’s Disease. Behav. Brain Res. 2018, 336, 177–181. [Google Scholar] [CrossRef]
- Tu, S.; Okamoto, S.-I.; Lipton, S.A.; Xu, H. Oligomeric Aβ-induced synaptic dysfunction in Alzheimer’s disease. Mol. Neurodegener. 2014, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Selkoe, D.J. Alzheimer’s Disease Is a Synaptic Failure. Science 2002, 298, 789–791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koffie, R.M.; Hyman, B.T.; Spires-Jones, T.L. Alzheimer’s Disease: Synapses Gone Cold. Mol. Neurodegener. 2011, 6, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Beja-Glasser, V.F.; Nfonoyim, B.M.; Frouin, A.; Li, S.; Ramakrishnan, S.; Merry, K.M.; Shi, Q.; Rosenthal, A.; Barres, B.A.; et al. Complement and Microglia Mediate Early Synapse Loss in Alzheimer Mouse Models. Science 2016, 352, 712–716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monasor, L.S.; Müller, S.A.; Colombo, A.V.; Tanrioever, G.; König, J.; Roth, S.; Liesz, A.; Berghofer, A.; Piechotta, A.; Prestel, M.; et al. Fibrillar Aβ triggers microglial proteome alterations and dysfunction in Alzheimer mouse models. eLife 2020, 9. [Google Scholar] [CrossRef]
- Heneka, M.T.; Carson, M.J.; El Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef] [Green Version]
- Tackenberg, C.; Ghori, A.; Brandt, R. Thin, Stubby or Mushroom: Spine Pathology in Alzheimer’s Disease. Curr. Alzheimer Res. 2009, 6, 261–268. [Google Scholar] [CrossRef]
- Simpson, D.S.A.; Oliver, P.L. ROS Generation in Microglia: Understanding Oxidative Stress and Inflammation in Neurodegenerative Disease. Antioxidants 2020, 9, 743. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Zhou, W.; Liu, S.; Deng, Y.; Cai, F.; Tone, M.; Tone, Y.; Tong, Y.; Song, W. Increased NF-κB Signalling up-Regulates Bace1 Expression and Its Therapeutic Potential in Alzheimer’s Disease. Int. J. Neuropsychopharmacol. 2012, 15, 77–90. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Rojas, C.; Burgos, P.V.; Inestrosa, N.C. Inhibition of Wnt Signaling Induces Amyloidogenic Processing of Amyloid Precursor Protein and the Production and Aggregation of Amyloid- β (Aβ) 42 Peptides. J. Neurochem. 2016, 139, 1175–1191. [Google Scholar] [CrossRef]
- Jin, N.; Zhu, H.; Liang, X.; Huang, W.; Xie, Q.; Xiao, P.; Ni, J.; Liu, Q. Sodium Selenate Activated Wnt/β-catenin Signaling and Repressed Amyloid-β Formation in a Triple Transgenic Mouse Model of Alzheimer’s Disease. Exp. Neurol. 2017, 297, 36–49. [Google Scholar] [CrossRef]
- Nejak-Bowen, K.; Kikuchi, A.; Monga, S.P. Beta-catenin-NF-κB Interactions in Murine Hepatocytes: A Complex to Die For. Hepatology 2013, 57, 763–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wang, K.; Wei, X.; Xie, T.; Lv, B.; Zhou, Q.; Wang, X. Interaction of NF-κB and Wnt/β-catenin Signaling Pathways in Alzheimer’s Disease and Potential Active Drug Treatments. Neurochem. Res. 2021, 46, 711–731. [Google Scholar] [CrossRef] [PubMed]
- Bayod, S.; Felice, P.; Andrés, P.; Rosa, P.; Camins, A.; Pallàs, M.; Canudas, A.-M. Downregulation of canonical Wnt signaling in hippocampus of SAMP8 mice. Neurobiol. Aging 2015, 36, 720–729. [Google Scholar] [CrossRef] [PubMed]
- Rosi, M.C.; Luccarini, I.; Grossi, C.; Fiorentini, A.; Spillantini, M.G.; Prisco, A.; Scali, C.; Gianfriddo, M.; Caricasole, A.; Ters-tappen, G.C.; et al. Increased Dickkopf-1 Expression in Transgenic Mouse Models of Neurodegenerative Disease. J. Neurochem. 2010, 112, 1539–1551. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Hottiger, M.O. Crosstalk between Wnt/β-Catenin and NF-κB Signaling Pathway during Inflammation. Front. Immunol. 2016, 7, 378. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | CAT# | Source | MW (kDa) | Dilution | Application |
| Anti-Aβ1-42 | ab201060 | Abcam | 37–50 | 1:500/1:1000/1:200 | WB/IHC/IF |
| Anti-Iba1 | ab48004 | Abcam | 17 | 1:1000/1:250/1:200 | WB/IHC/IF |
| Anti-PSD95 | #2507 | CST | 95 | 1:1000/1:400 | WB/IF |
| Anti-Synaptophysin | #36406 | CST | 38 | 1:1000/1:100 | WB/IF |
| Anti-BACE | #5606 | CST | 70 | 1:1000 | WB |
| Anti-App | #2452 | CST | 100–140 | 1:1000 | WB |
| Anti-NF-κB p65 | #8242 | CST | 65 | 1:1000 | WB |
| Anti-pNF-κB p65 | sc166748 | SCB | 65 | 1:1000 | WB |
| Anti-GSK-3β | #9832 | CST | 46 | 1:1000 | WB |
| Anti-pGSK-3β(Ser9) | #9323 | CST | 46 | 1:1000 | WB |
| Anti-β-catenin | sc-7963 | SCB | 92 | 1:1000 | WB |
| Anti-GAPDH | #2118 | CST | 37 | 1:1000 | WB |
| Anti-β-Actin | #4970 | CST | 45 | 1:1000 | WB |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Zhou, Q.; Zhang, J.-H.; Wang, K.-Y.; Saito, T.; Saido, T.C.; Wang, X.; Gao, X.; Azuma, K. Microglia-Based Sex-Biased Neuropathology in Early-Stage Alzheimer’s Disease Model Mice and the Potential Pharmacologic Efficacy of Dioscin. Cells 2021, 10, 3261. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113261
Liu X, Zhou Q, Zhang J-H, Wang K-Y, Saito T, Saido TC, Wang X, Gao X, Azuma K. Microglia-Based Sex-Biased Neuropathology in Early-Stage Alzheimer’s Disease Model Mice and the Potential Pharmacologic Efficacy of Dioscin. Cells. 2021; 10(11):3261. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113261
Chicago/Turabian StyleLiu, Xiao, Qian Zhou, Jia-He Zhang, Ke-Yong Wang, Takashi Saito, Takaomi C. Saido, Xiaoying Wang, Xiumei Gao, and Kagaku Azuma. 2021. "Microglia-Based Sex-Biased Neuropathology in Early-Stage Alzheimer’s Disease Model Mice and the Potential Pharmacologic Efficacy of Dioscin" Cells 10, no. 11: 3261. https://0-doi-org.brum.beds.ac.uk/10.3390/cells10113261