
Interleukin-6 Signaling in Triple Negative Breast Cancer Cells Elicits the Annexin A1/Formyl Peptide Receptor 1 Axis and Affects the Tumor Microenvironment

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. Cell Lysis and Immunoblotting
2.3. Flow Cytometry
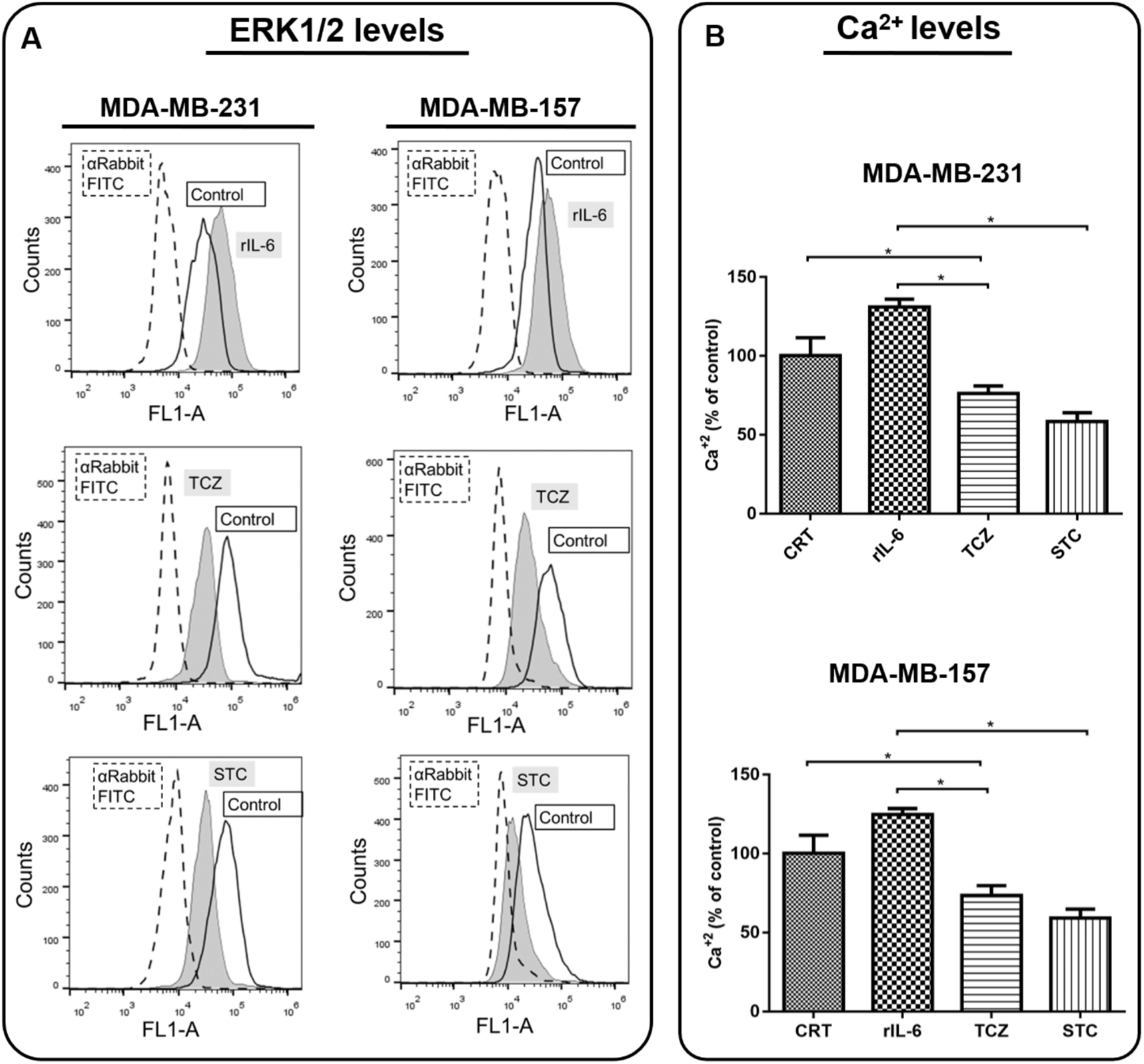
2.4. Calcium Flux
2.5. Immunohistochemistry
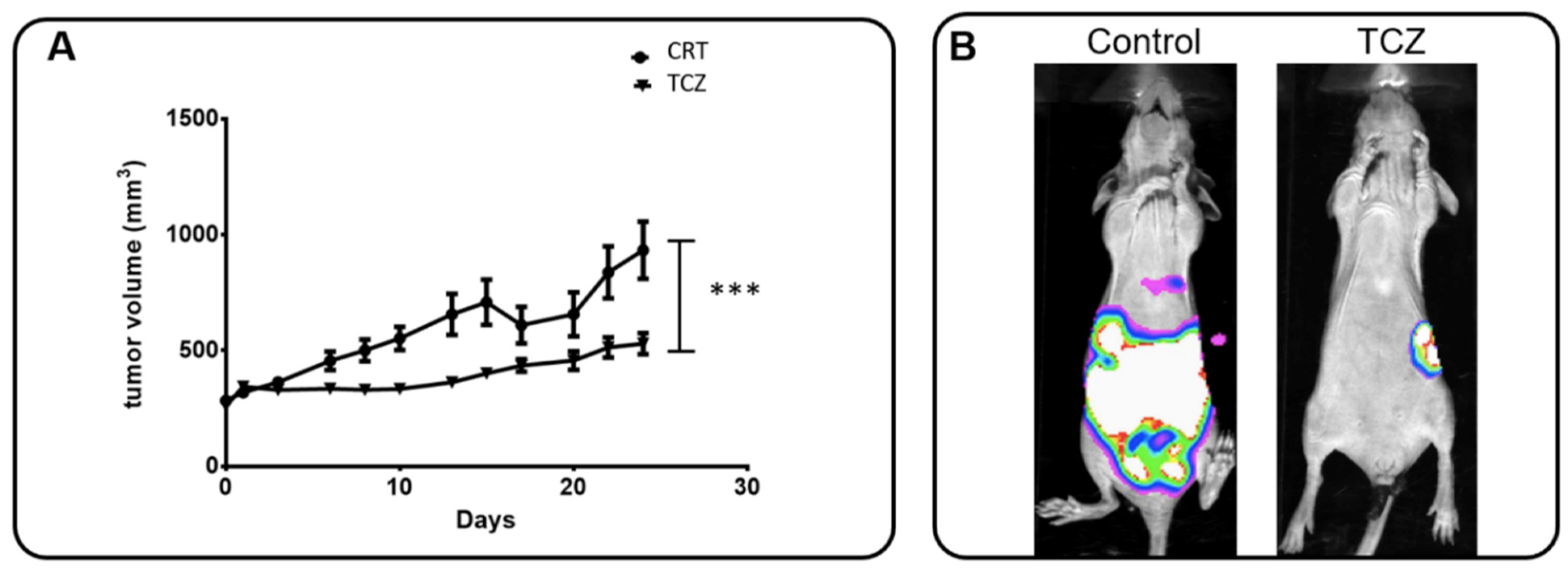
2.6. In Vivo Assay
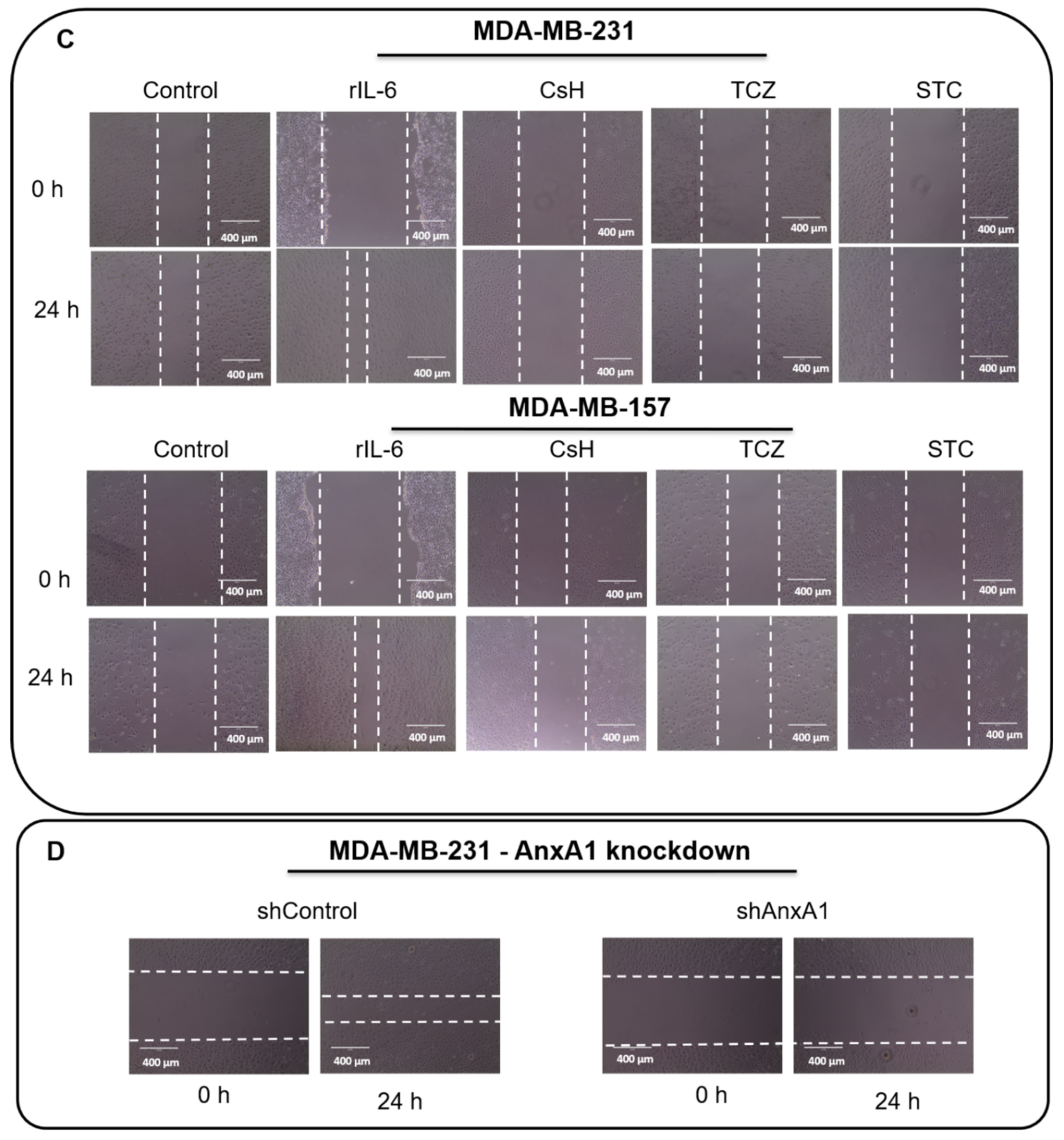
2.7. Wound-Healing Assay
2.8. ELISA
2.9. Quantitative Real-Time PCR
2.10. AnxA1 Knockdown
2.11. Statistical Analysis
3. Results
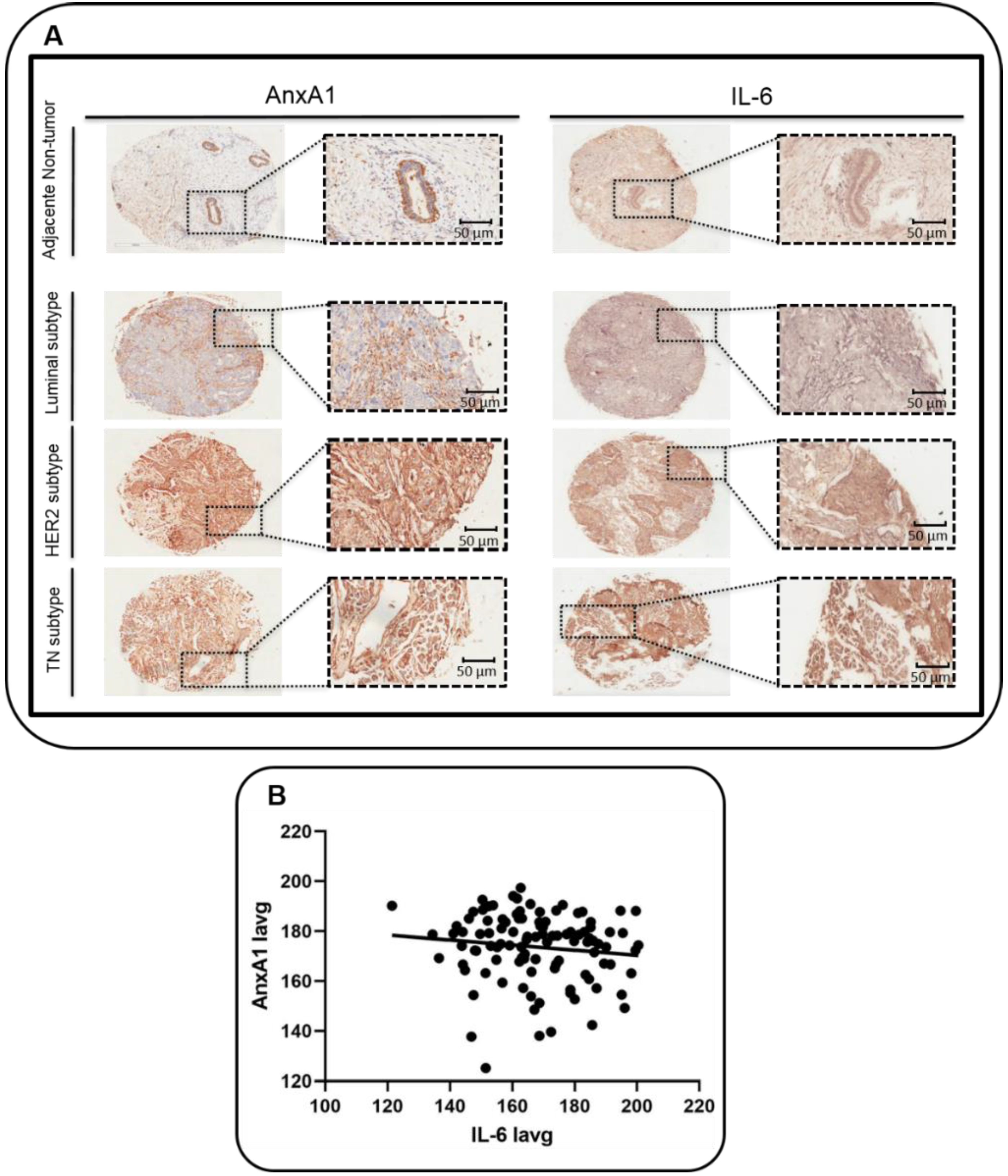
3.1. Correlation between AnxA1 and IL-6 Expression in BC Samples
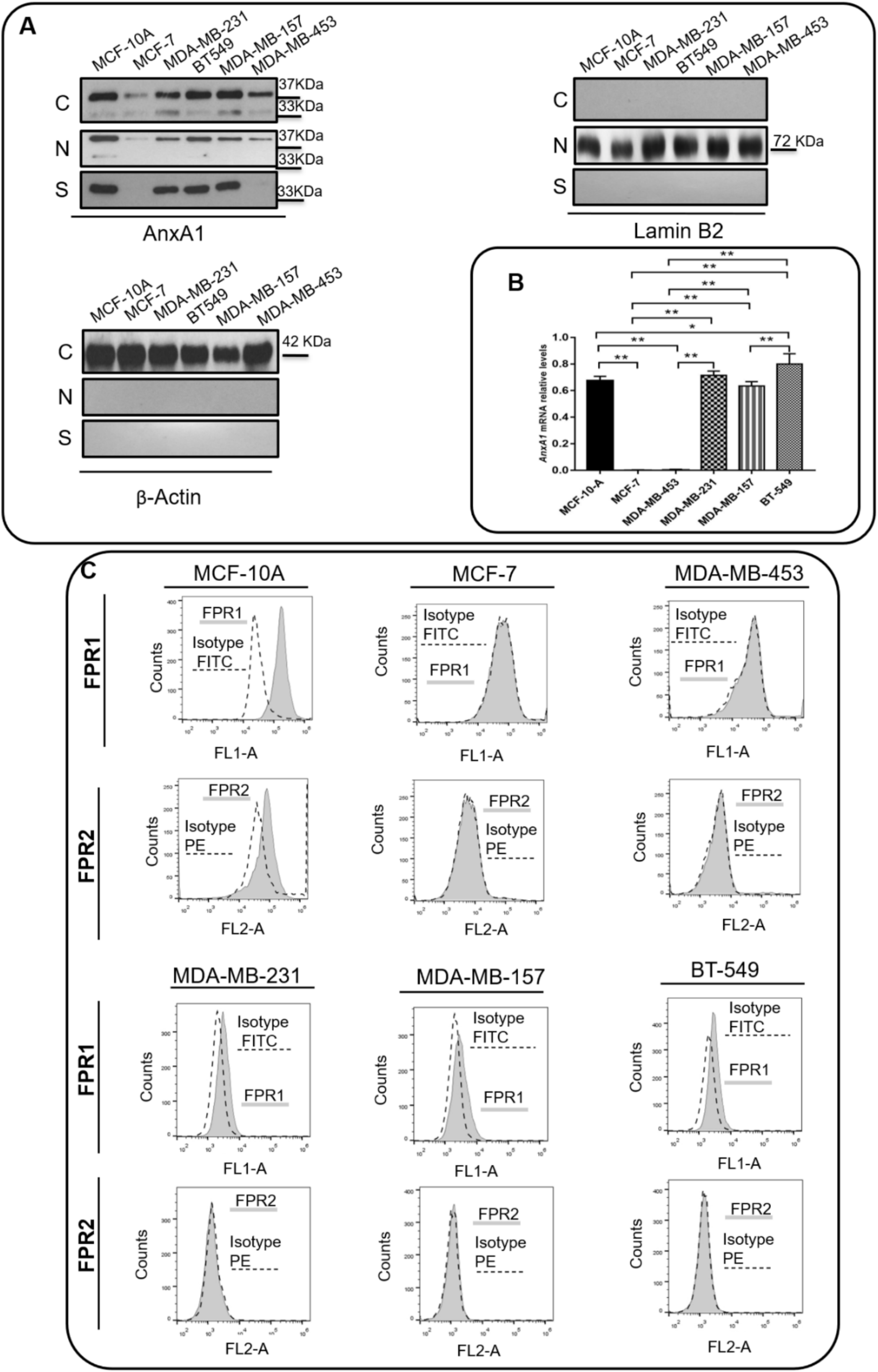
3.2. TNBC Cell Lines Express High Levels of AnxA1 and IL-6
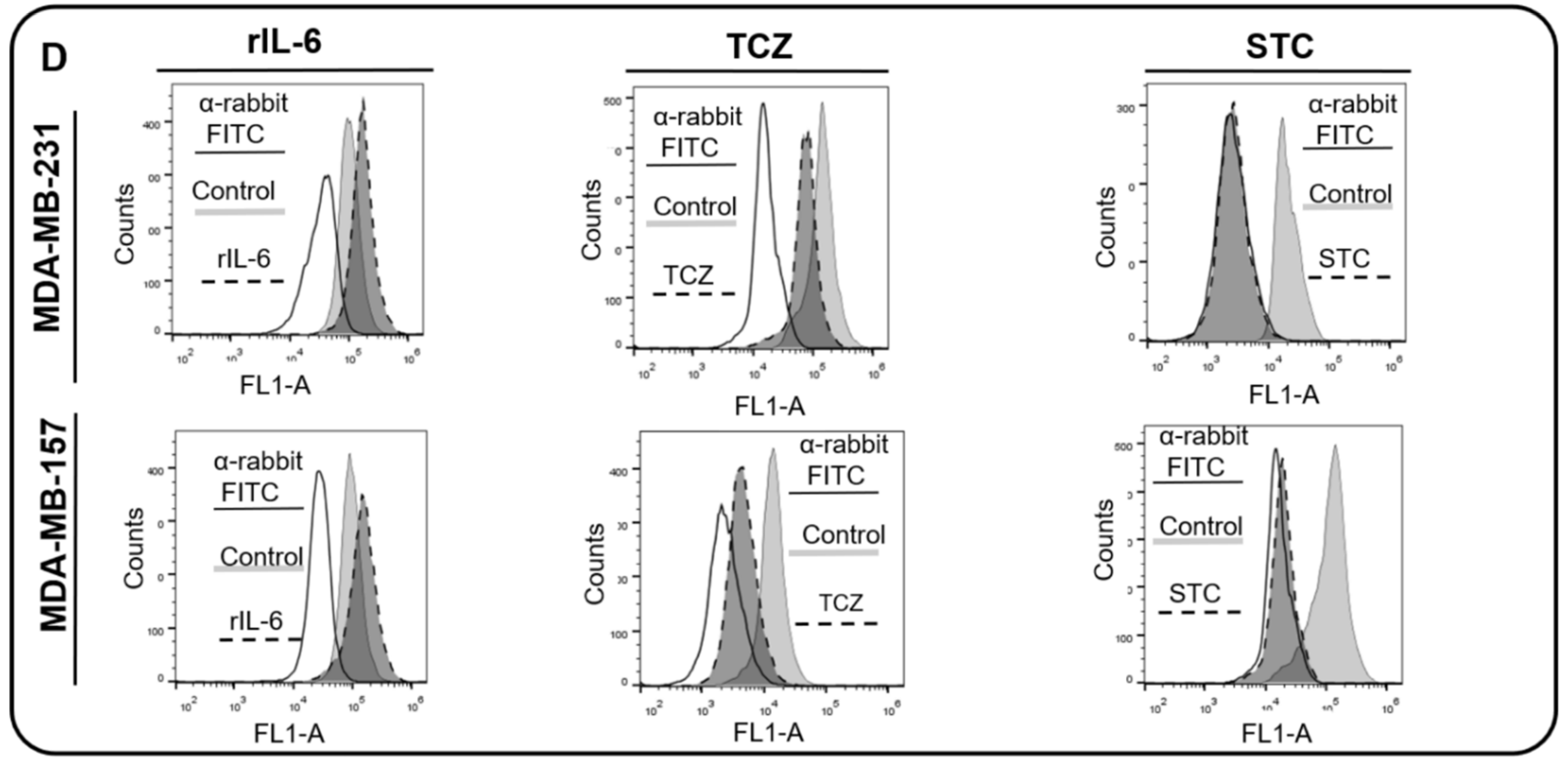
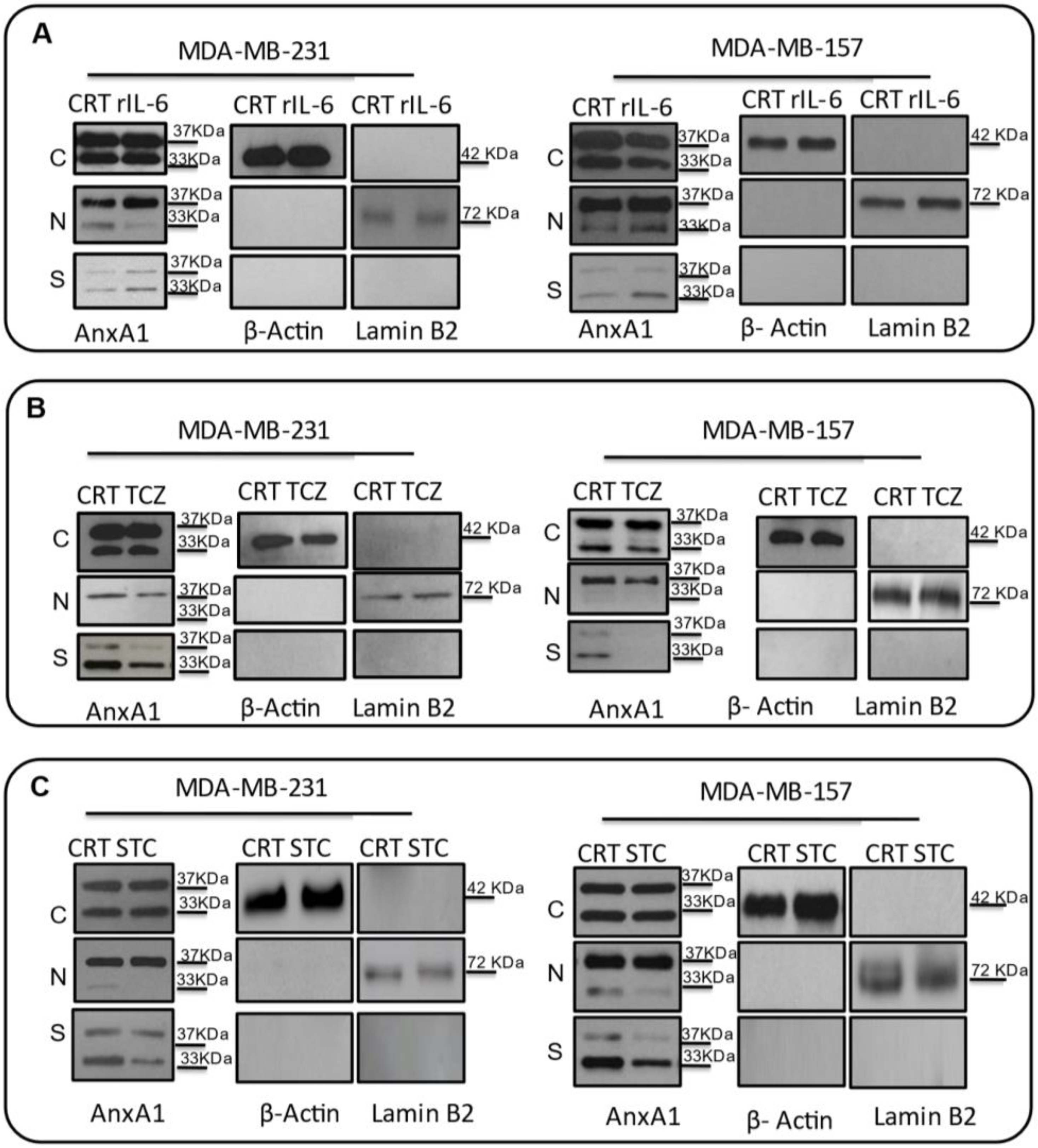
3.3. AnxA1 Autocrine Signaling Leads to a Decrease in IL-6 Expression
3.4. IL-6 Inhibition Is Involved in Tumor Growth
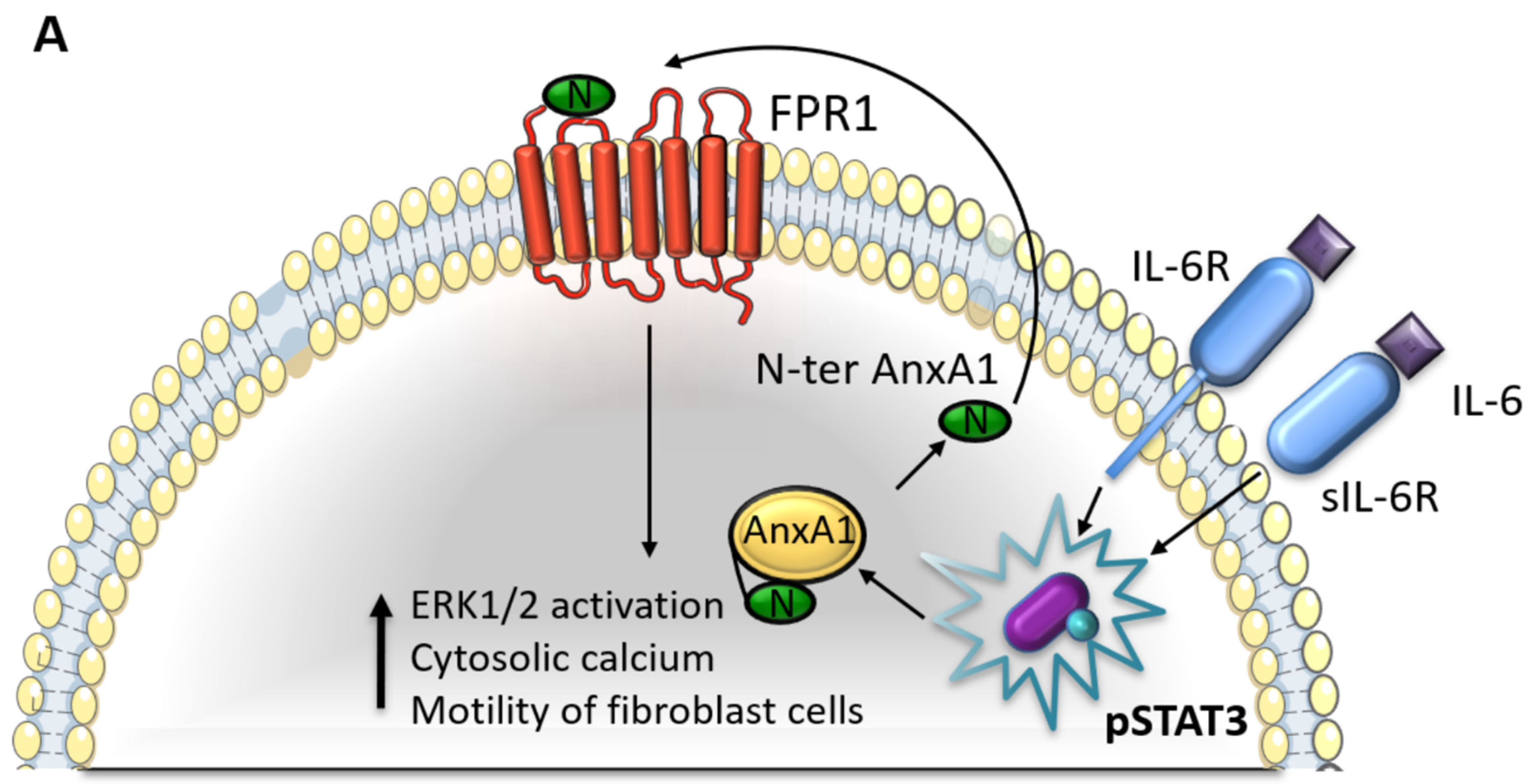
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harborg, S.; Zachariae, R.; Olsen, J.; Johannsen, M.; Cronin-Fenton, D.; Bøggild, H.; Borgquist, S. Overweight and prognosis in triple-negative breast cancer patients: A systematic review and meta-analysis. Npj Breast Cancer 2021, 7, 119. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Chan, P.S.; Lok, V.; Chen, X.; Ding, H.; Jin, Y.; Yuan, J.; Lao, X.-Q.; Zheng, Z.-J.; Wong, M.C. Global incidence and mortality of breast cancer: A trend analysis. Aging 2021, 13, 5748–5803. [Google Scholar] [CrossRef] [PubMed]
- Burguin, A.; Diorio, C.; Durocher, F. Breast Cancer Treatments: Updates and New Challenges. J. Pers. Med. 2021, 11, 808. [Google Scholar] [CrossRef] [PubMed]
- Rossing, M.; Pedersen, C.B.; Tvedskov, T.; Vejborg, I.; Talman, M.-L.; Olsen, L.R.; Kroman, N.; Nielsen, F.C.; Jensen, M.-B.; Ejlertsen, B. Clinical implications of intrinsic molecular subtypes of breast cancer for sentinel node status. Sci. Rep. 2021, 11, 2259. [Google Scholar] [CrossRef]
- Kanwal, B. Untangling Triple-Negative Breast Cancer Molecular Peculiarity and Chemo-Resistance: Trailing Towards Marker-Based Targeted Therapies. Cureus 2021, 13, e16636. [Google Scholar] [CrossRef]
- Berger, E.; Park, T.; Saridakis, A.; Golshan, M.; Greenup, R.; Ahuja, N. Immunotherapy Treatment for Triple Negative Breast Cancer. Pharmaceuticals 2021, 14, 763. [Google Scholar] [CrossRef]
- Hussain, S.P.; Harris, C.C. Inflammation and cancer: An ancient link with novel potentials. Int. J. Cancer 2007, 121, 2373–2380. [Google Scholar] [CrossRef]
- Schetter, A.J.; Heegaard, N.H.H.; Harris, C.C. Inflammation and cancer: Interweaving microRNA, free radical, cytokine and p53 pathways. Carcinogenesis 2009, 31, 37–49. [Google Scholar] [CrossRef] [Green Version]
- Gerke, V.; Moss, S.E. Annexins: From Structure to Function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef]
- Petrella, A.; Festa, M.; Ercolino, S.F.; Zerilli, M.; Stassi, G.; Solito, E.; Parente, L. Induction of annexin-1 during TRAIL-induced apoptosis in thyroid carcinoma cells. Cell Death Differ. 2005, 12, 1358–1360. [Google Scholar] [CrossRef] [Green Version]
- Scannell, M.; Maderna, P. Lipoxins and Annexin-1: Resolution of Inflammation and Regulation of Phagocytosis of Apoptotic Cells. Sci. World J. 2006, 6, 1555–1573. [Google Scholar] [CrossRef] [PubMed]
- Rescher, U.; Gerke, V. Annexins—Unique membrane binding proteins with diverse functions. J. Cell Sci. 2004, 117, 2631–2639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaguchi, M.; Murata, H.; Sonegawa, H.; Sakaguchi, Y.; Futami, J.-I.; Kitazoe, M.; Yamada, H.; Huh, N.-H. Truncation of Annexin A1 Is a Regulatory Lever for Linking Epidermal Growth Factor Signaling with Cytosolic Phospholipase A2 in Normal and Malignant Squamous Epithelial Cells. J. Biol. Chem. 2007, 282, 35679–35686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parente, L.; Solito, E. Annexin 1: More than an anti-phospholipase protein. Agents Actions 2004, 53, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Gavins, F.N.E.; Yona, S.; Kamal, A.M.; Flower, R.J.; Perretti, M. Leukocyte antiadhesive actions of annexin 1: ALXR- and FPR-related anti-inflammatory mechanisms. Blood 2003, 101, 4140–4147. [Google Scholar] [CrossRef] [Green Version]
- Araújo, T.G.; Mota, S.T.S.; Ferreira, H.S.V.; Ribeiro, M.A.; Goulart, L.R.; Vecchi, L. Annexin A1 as a Regulator of Immune Response in Cancer. Cells 2021, 10, 2245. [Google Scholar] [CrossRef]
- Sheikh, M.H.; Solito, E. Annexin A1: Uncovering the Many Talents of an Old Protein. Int. J. Mol. Sci. 2018, 19, 1045. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Teijeiro, S.; Menéndez, S.T.; Villaronga, M.; Pena-Alonso, E.; Rodrigo, J.P.; Morgan, R.O.; Granda-Díaz, R.; Salom, C.; Fernandez, M.P.; García-Pedrero, J.M. Annexin A1 down-regulation in head and neck squamous cell carcinoma is mediated via transcriptional control with direct involvement of miR-196a/b. Sci. Rep. 2017, 7, 6790. [Google Scholar] [CrossRef] [Green Version]
- Mota, S.T.S.; Vecchi, L.; Alves, D.A.; Cordeiro, A.O.; Guimarães, G.S.; Campos-Fernández, E.; Maia, Y.C.P.; Dornelas, B.D.C.; Bezerra, S.M.; de Andrade, V.P.; et al. Annexin A1 promotes the nuclear localization of the epidermal growth factor receptor in castration-resistant prostate cancer. Int. J. Biochem. Cell Biol. 2020, 127, 105838. [Google Scholar] [CrossRef]
- Liu, Y.-F.; Zhang, P.-F.; Li, M.-Y.; Li, Q.-Q.; Chen, Z.-C. Identification of annexin A1 as a proinvasive and prognostic factor for lung adenocarcinoma. Clin. Exp. Metastasis 2011, 28, 413–425. [Google Scholar] [CrossRef]
- Lin, C.-Y.; Jeng, Y.-M.; Chou, H.-Y.; Hsu, H.-C.; Yuan, R.-H.; Chiang, C.-P.; Kuo, M.Y.-P. Nuclear localization of annexin A1 is a prognostic factor in oral squamous cell carcinoma. J. Surg. Oncol. 2008, 97, 544–550. [Google Scholar] [CrossRef] [PubMed]
- Sobral-Leite, M.; Wesseling, J.; Smit, V.T.H.B.M.; Nevanlinna, H.; van Miltenburg, M.H.; Sanders, J.; Hofland, I.; Blows, F.M.; Coulson, P.; Gazinska, P.; et al. Annexin A1 expression in a pooled breast cancer series: Association with tumor subtypes and prognosis. BMC Med. 2015, 13, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vecchi, L.; Zóia, M.A.P.; Santos, T.G.; Beserra, A.D.O.; Ramos, C.; Colombo, B.F.M.; Maia, Y.C.P.; de Andrade, V.P.; Mota, S.T.S.; Araújo, T.; et al. Inhibition of the AnxA1/FPR1 autocrine axis reduces MDA-MB-231 breast cancer cell growth and aggressiveness in vitro and in vivo. Biochim. Biophys. Acta 2018, 1865, 1368–1382. [Google Scholar] [CrossRef] [PubMed]
- de Graauw, M.; van Miltenburg, M.H.; Schmidt, M.K.; Pont, C.; Lalai, R.; Kartopawiro, J.; Pardali, E.; Le Dévédec, S.E.; Smit, V.T.; van der Wal, A.; et al. Annexin A1 regulates TGF-β signaling and promotes metastasis formation of basal-like breast cancer cells. Proc. Natl. Acad. Sci. USA 2010, 107, 6340–6345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, S.L.; Milne, I.R.; Bagley, C.J.; Gamble, J.R.; Vadas, M.A.; Pitson, S.M.; Khew-Goodall, Y. A Proinflammatory Role for Proteolytically Cleaved Annexin A1 in Neutrophil Transendothelial Migration. J. Immunol. 2010, 185, 3057–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solito, E.; De Coupade, C.; Parente, L.; Flower, R.J.; Russo-Marie, F. IL-6 stimulates annexin 1 expression and translocation and suggests a new biological role as class ii acute phase protein. Cytokine 1998, 10, 514–521. [Google Scholar] [CrossRef]
- Ni, C.; Gao, S.; Zheng, Y.; Liu, P.; Zhai, Y.; Huang, W.; Jiang, H.; Lv, Q.; Kong, D.; Jiang, Y. Annexin A1 Attenuates Neutrophil Migration and IL-6 Expression through Fpr2 in a Mouse Model of Streptococcus suis-Induced Meningitis. Infect. Immun. 2021, 89, e00680-20. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in Inflammation, Immunity, and Disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- Lee, S.O.; Yang, X.; Duan, S.; Tsai, Y.; Strojny, L.R.; Keng, P.; Chen, Y. IL-6 promotes growth and epithelial-mesenchymal transition of CD133+ cells of non-small cell lung cancer. Oncotarget 2015, 7, 6626–6638. [Google Scholar] [CrossRef] [Green Version]
- Scheller, J.; Ohnesorge, N.; Rose-John, S. Interleukin-6 Trans-Signalling in Chronic Inflammation and Cancer. Scand. J. Immunol. 2006, 63, 321–329. [Google Scholar] [CrossRef]
- Ma, J.-H.; Qin, L.; Li, X. Role of STAT3 signaling pathway in breast cancer. Cell Commun. Signal. 2020, 18, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshio-Hoshino, N.; Nishimoto, N. The Blockade of IL-6 Signaling in Rational Drug Design. Curr. Pharm. Des. 2008, 14, 1217–1224. [Google Scholar] [CrossRef]
- Culig, Z.; Puhr, M. Interleukin-6: A multifunctional targetable cytokine in human prostate cancer. Mol. Cell. Endocrinol. 2012, 360, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Masjedi, A.; Hashemi, V.; Hojjat-Farsangi, M.; Ghalamfarsa, G.; Azizi, G.; Yousefi, M.; Jadidi-Niaragh, F. The significant role of interleukin-6 and its signaling pathway in the immunopathogenesis and treatment of breast cancer. Biomed. Pharmacother. 2018, 108, 1415–1424. [Google Scholar] [CrossRef]
- Garcia-Tunon, I.; Ricote, M.; Ruiz, A.; Fraile, B.; Paniagua, R.; Royuela, M. IL-6, its receptors and its relationship with bcl-2 and bax proteins in infiltrating and in situ human breast carcinoma. Histopathology 2005, 47, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, A.; Santini, D.; Bonafè, M.; Taffurelli, M.; Avenia, N. Interleukin-6 and pro inflammatory status in the breast tumor microenvironment. World J. Surg. Oncol. 2015, 13, 129. [Google Scholar] [CrossRef] [Green Version]
- Jorcyk, C.L.; Tawara, K. Clinical significance of interleukin (IL)-6 in cancer metastasis to bone: Potential of anti-IL-6 therapies. Cancer Manag. Res. 2011, 3, 177–189. [Google Scholar] [CrossRef]
- Pu, Y.-S.; Hour, T.-C.; Chuang, S.-E.; Cheng, A.-L.; Lai, M.-K.; Kuo, M.-L. Interleukin-6 is responsible for drug resistance and anti-apoptotic effects in prostatic cancer cells. Prostate 2004, 60, 120–129. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, X.L.; Qu, Y.; Wu, J.; Zhu, Y.Q.; Sun, W.J.; Li, L.Z. Autocrine production of interleukin-6 confers cisplatin and paclitaxel resistance in ovarian cancer cells. Cancer Lett. 2010, 295, 110–123. [Google Scholar] [CrossRef]
- Wu, P.; Wu, D.; Zhao, L.; Huang, L.; Shen, G.; Chai, Y. Prognostic role of STAT3 in solid tumors: A systematic review and meta-analysis. Oncotarget 2016, 7, 19863–19883. [Google Scholar] [CrossRef] [Green Version]
- Spitzner, M.; Ebner, R.; Wolff, H.A.; Ghadimi, B.M.; Wienands, J.; Grade, M. STAT3: A Novel Molecular Mediator of Resistance to Chemoradiotherapy. Cancers 2014, 6, 1986–2011. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, K.; Resat, H. Constitutive activation of STAT3 in breast cancer cells: A review. Int. J. Cancer 2015, 138, 2570–2578. [Google Scholar] [CrossRef] [PubMed]
- Yeo, S.K.; Guan, J.-L. Breast Cancer: Multiple Subtypes within a Tumor? Trends Cancer 2017, 3, 753–760. [Google Scholar] [CrossRef] [PubMed]
- Althoubaity, F.K. Molecular classification of breast cancer: A retrospective cohort study. Ann. Med. Surg. 2019, 49, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahmood, S.; Sapiezynski, J.; Garbuzenko, O.B.; Minko, T. Metastatic and triple-negative breast cancer: Challenges and treatment options. Drug Deliv. Transl. Res. 2018, 8, 1483–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S.-C. Triple-negative breast cancer molecular subtyping and treatment progress. Breast Cancer Res. 2020, 22, 129. [Google Scholar] [CrossRef] [PubMed]
- Okano, M.; Oshi, M.; Butash, A.L.; Katsuta, E.; Tachibana, K.; Saito, K.; Okayama, H.; Peng, X.; Yan, L.; Kono, K.; et al. Triple-Negative Breast Cancer with High Levels of Annexin A1 Expression Is Associated with Mast Cell Infiltration, Inflammation, and Angiogenesis. Int. J. Mol. Sci. 2019, 20, 4197. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.H.; Aeberli, D.; Dacumos, A.; Xue, J.R.; Morand, E. Annexin-1 Regulates Macrophage IL-6 and TNF via Glucocorticoid-Induced Leucine Zipper. J. Immunol. 2009, 183, 1435–1445. [Google Scholar] [CrossRef] [Green Version]
- Moraes, L.A.; Ampomah, P.B.; Lim, L.H.K. Annexin A1 in inflammation and breast cancer: A new axis in the tumor microenvironment. Cell Adhes. Migr. 2018, 12, 417–423. [Google Scholar] [CrossRef] [Green Version]
- Mauer, J.; Denson, J.L.; Brüning, J.C. Versatile functions for IL-6 in metabolism and cancer. Trends Immunol. 2015, 36, 92–101. [Google Scholar] [CrossRef]
- Johnson, D.E.; O’Keefe, R.A.; Grandis, J.R. Targeting the IL-6/JAK/STAT3 signalling axis in cancer. Nat. Rev. Clin. Oncol. 2018, 15, 234–248. [Google Scholar] [CrossRef] [PubMed]
- Weng, Y.-S.; Tseng, H.-Y.; Chen, Y.-A.; Shen, P.-C.; Al Haq, A.T.; Chen, L.-M.; Tung, Y.-C.; Hsu, H.-L. MCT-1/miR-34a/IL-6/IL-6R signaling axis promotes EMT progression, cancer stemness and M2 macrophage polarization in triple-negative breast cancer. Mol. Cancer 2019, 18, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.-J.; Yan, L.; Zhang, J.; Zhang, W.-D. STAT3 as a potential therapeutic target in triple negative breast cancer: A systematic review. J. Exp. Clin. Cancer Res. 2019, 38, 195. [Google Scholar] [CrossRef] [PubMed]
- Jin, K.; Pandey, N.B.; Popel, A.S. Simultaneous blockade of IL-6 and CCL5 signaling for synergistic inhibition of triple-negative breast cancer growth and metastasis. Breast Cancer Res. 2018, 20, 54. [Google Scholar] [CrossRef] [Green Version]
- Fasoulakis, Z.; Kolios, G.; Papamanolis, V.; Kontomanolis, E.N. Interleukins Associated with Breast Cancer. Cureus 2018, 10, e3549. [Google Scholar] [CrossRef] [Green Version]
- Studebaker, A.W.; Storci, G.; Werbeck, J.L.; Sansone, P.; Sasser, A.K.; Tavolari, S.; Huang, T.; Chan, M.; Marini, F.C.; Rosol, T.; et al. Fibroblasts Isolated from Common Sites of Breast Cancer Metastasis Enhance Cancer Cell Growth Rates and Invasiveness in an Interleukin-6–Dependent Manner. Cancer Res. 2008, 68, 9087–9095. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Kieffer, Y.; Scholer-Dahirel, A.; Pelon, F.; Bourachot, B.; Cardon, M.; Sirven, P.; Magagna, I.; Fuhrmann, L.; Bernard, C.; et al. Fibroblast heterogeneity and immunosuppressive environment in human breast cancer. Cancer Cell 2018, 33, 463–479.e10. [Google Scholar] [CrossRef] [Green Version]
- Karakasheva, T.A.; Lin, E.W.; Tang, Q.; Qiao, E.; Waldron, T.J.; Soni, M.; Klein-Szanto, A.J.; Sahu, V.; Basu, D.; Ohashi, S.; et al. IL-6 Mediates Cross-Talk between Tumor Cells and Activated Fibroblasts in the Tumor Microenvironment. Cancer Res. 2018, 78, 4957–4970. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, Y.; Kitamura, H.; Xiang, H.; Ohno, Y.; Homma, S.; Kawamura, H.; Takahashi, N.; Kamiyama, T.; Tanino, M.; Taketomi, A. IL6 Modulates the Immune Status of the Tumor Microenvironment to Facilitate Metastatic Colonization of Colorectal Cancer Cells. Cancer Immunol. Res. 2019, 7, 1944–1957. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.A.; Jenkins, B.J. Recent insights into targeting the IL-6 cytokine family in inflammatory diseases and cancer. Nat. Rev. Immunol. 2018, 18, 773–789. [Google Scholar] [CrossRef]
- Hirano, T. IL-6 in inflammation, autoimmunity and cancer. Int. Immunol. 2020, 33, 127–148. [Google Scholar] [CrossRef] [PubMed]
- Alraouji, N.N.; Aboussekhra, A. Tocilizumab inhibits IL-8 and the proangiogenic potential of triple negative breast cancer cells. Mol. Carcinog. 2020, 60, 51–59. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Patients | |||
|---|---|---|---|---|
| AnxA1 N (%) | IL-6 N (%) | |||
| High | Low | High | Low | |
| Age (year) | ||||
| ≥50 | 30 (28.3) | 26 (24.5) | 27 (25.5) | 29 (27.4) |
| <50 | 18 (17) | 32 (30.2) | 26 (24.5) | 24 (22.6) |
| p-value | 0.07 | 0.7 | ||
| Odds Ratio | 2.1 95% CI (0.9–4.5) | 0.8 95% CI (0.4–1.8) | ||
| ER | ||||
| Positive | 14 (13.2) | 27 (25.5) | 21 (19.8) | 20 (18.9) |
| Negative | 34 (32.1) | 31 (29.2) | 32 (30.2) | 33 (31.1) |
| p-value | 0.07 | 0.8 | ||
| Odds Ratio | 0.5 95% CI (0.2–1.1) | 1.1 95% CI (0.5–2.4) | ||
| PR | ||||
| Positive | 10 (9.4) | 18 (17) | 14 (13.2) | 14 (13.2) |
| Negative | 38 (35.9) | 40 (37.7) | 39 (36.8) | 39 (36.8) |
| p-value | 0.2 | 1.0 | ||
| Odds Ratio | 0.6 95% CI (0.3–1.4) | 1.0 95% CI (0.4–2.4) | ||
| HER2-enriched | ||||
| Positive | 18 (17) | 21 (19.8) | 15 (14.1) | 24 (22.6) |
| Negative | 30 (28.3) | 37 (34.9) | 38 (35.9) | 29 (27.4) |
| p-value | 0.9 | 0.07 | ||
| Odds Ratio | 1.1 95% CI (0.5–2.3) | 0.5 95% CI (0.2–1.1) | ||
| Histological grading | ||||
| GIII | 5 (4.6) | 8 (14) | 6 (5.6) | 7 (6.7) |
| GI-GII | 43 (16.3) | 50 (65.1) | 47 (44.3) | 46 (43.4) |
| p-value | 0.6 | 0.8 | ||
| Odds Ratio | 0.7 95% CI (0.2–2.4) | 0.8 95% CI (0.4–3.8) | ||
| Stage | ||||
| IIIA-IIIB | 11 (16.3) | 19 (37.2) | 16 (15.1) | 14 (37.2) |
| I-II (A-B) | 37 (4.6) | 39 (41.9) | 37 (34.9) | 39 (36.8) |
| p-value | 0.3 | 0.7 | ||
| Odds Ratio | 0.6 95% CI (0.3–1.4) | 1.2 95% CI (0.3–2.7) | ||
| Tumor size (cT) | ||||
| T3-T4 | 10 (9.3) | 17 (20.9) | 14 (13.2) | 13 (12.2) |
| T1-T2 | 38 (11.7) | 41 (58.1) | 39 (39.6) | 40 (37.7) |
| p-value | 0.3 | 0.8 | ||
| Odds Ratio | 0.6 95% CI (0.3–1.6) | 1.1 95% CI (0.5–2.6) | ||
| Lymph node (cN) | ||||
| N1-2 | 33 (35.4) | 36 (32.3) | 27 (25.5) | 42 (39.6) |
| N0 | 15 (16.7) | 22 (15.6) | 26 (24.5) | 11 (10.4) |
| p-value | 0.5 | 0.003* | ||
| Odds Ratio | 1.3 95% CI (0.6 -3.0) | 0.3 (95% CI (0.1–0.6) | ||
| Triple Negative | ||||
| Yes | 21 (19.8) | 11 (10.4) | 15 (14.1) | 17 (16) |
| No | 27 (25.5) | 47 (44.3) | 38 (35.9) | 36 (34) |
| p-value | * 0.007 | 0.7 | ||
| Odds Ratio | 3.3 95% CI (1.4–7.9) | 0.8 95% CI (0.4–2.0) | ||
| Luminal A | ||||
| Yes | 9 | 28 | 24 (22.6) | 13 (12.2) |
| No | 39 (36.8) | 30 (28.3) | 29 (27.4) | 40 (37.7) |
| p-value | * 0.002 | * 0.03 | ||
| Odds Ratio | 0.3 95% CI (0.1–0.6) | 2.6 95% CI (1.1–5.8) | ||
| Luminal B | ||||
| Yes | 5 (4.6) | 6 (5.7) | 1 (0.9) | 10 (9.4) |
| No | 43 (40.6) | 52 (49.1) | 52 (49.1) | 43 (40.6) |
| p-value | 0.9 | * 0.02 | ||
| Odds Ratio | 1.0 95% CI (0.3–3.5) | 0.1 95% CI (0.01–0.7) | ||
| HER2 enriched | ||||
| Yes | 13 (12.2) | 13 (12.2) | 13 (12.2) | 13 (12.2) |
| No | 35 (33.1) | 45 (42.5) | 40 (37.7) | 40 (37.7) |
| p-value | 0.6 | 1.0 | ||
| Odds Ratio | 1.2 95% CI (0.5–3.1) | 1.0 95% CI (0.4–2.4) | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vecchi, L.; Mota, S.T.S.; Zóia, M.A.P.; Martins, I.C.; de Souza, J.B.; Santos, T.G.; Beserra, A.d.O.; de Andrade, V.P.; Goulart, L.R.; Araújo, T.G. Interleukin-6 Signaling in Triple Negative Breast Cancer Cells Elicits the Annexin A1/Formyl Peptide Receptor 1 Axis and Affects the Tumor Microenvironment. Cells 2022, 11, 1705. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101705
Vecchi L, Mota STS, Zóia MAP, Martins IC, de Souza JB, Santos TG, Beserra AdO, de Andrade VP, Goulart LR, Araújo TG. Interleukin-6 Signaling in Triple Negative Breast Cancer Cells Elicits the Annexin A1/Formyl Peptide Receptor 1 Axis and Affects the Tumor Microenvironment. Cells. 2022; 11(10):1705. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101705
Chicago/Turabian StyleVecchi, Lara, Sara Teixeira Soares Mota, Mariana Alves Pereira Zóia, Isabella Castro Martins, Jessica Brito de Souza, Tiago Góss Santos, Adriano de Oliveira Beserra, Victor Piana de Andrade, Luiz Ricardo Goulart, and Thaise Gonçalves Araújo. 2022. "Interleukin-6 Signaling in Triple Negative Breast Cancer Cells Elicits the Annexin A1/Formyl Peptide Receptor 1 Axis and Affects the Tumor Microenvironment" Cells 11, no. 10: 1705. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101705