
Exosome-Associated circRNAs as Key Regulators of EMT in Cancer


Department of Molecular Medicine, Sapienza University of Rome, Viale Regina Elena, 324, 00161 Rome, Italy
*
Author to whom correspondence should be addressed.
Cells 2022, 11(10), 1716; https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101716
Submission received: 10 March 2022
/
Revised: 17 May 2022
/
Accepted: 19 May 2022
/
Published: 23 May 2022
(This article belongs to the Special Issue Extracellular Vesicle-Associated Non-Coding RNAs)
Abstract
:Epithelial-to-mesenchymal transition (EMT) is a dynamic program of cell plasticity aberrantly reactivated in cancer. The crosstalk between tumor cells and the tumoral microenvironment (TME) has a pivotal importance for the induction of the EMT and the progression toward a malignant phenotype. Notably, exosomes are key mediators of this crosstalk as vehicles of specific molecular signals that include the class of circular RNAs (circRNAs). This review specifically focuses on the role of exosome-associated circRNAs as key regulators of EMT in cancer. The relevance of these molecules in regulating the intercellular communication in TME and tumor progression is highlighted. Moreover, the here-presented evidence indicates that exosome-associated circRNA modulation should be taken in account for cancer diagnostic and therapeutic approaches.
1. Introduction
Epithelial-to-mesenchymal transition (EMT) is a complex process by which epithelial polarized cells lose cell–cell interactions and acquire a mesenchymal migratory phenotype. Different EMT subtypes can be categorized according to when and where the process occurs: (i) type 1 concerns developmental and organogenesis events; (ii) type 2 plays a major role in wound healing, tissue regeneration, and fibrosis; and (iii) type 3 characterizes epithelial cancer. The reactivation of the transdifferentiation process, indeed, renders tumor cells able to metastasize, thanks to the acquisition of migration ability and invasiveness properties. The relevance of the EMT in cancer biology is further underlined by the evidence that EMT traits are often coupled to stemness properties, increased tumorigenicity, metabolic reprogramming, and a pro-survival phenotype in stress conditions. Therefore, therapeutic treatments targeting EMT could interfere with multiple aspects that foster tumor progression.
Several observations unveiled the dynamics of the EMT process that occurs and progresses throughout a series of gradual molecular and morphological changes of epithelial cell phenotype and that, in some cases, can revert through an inverse transdifferentiation process, named mesenchymal-to-epithelial transition (MET) [1,2,3,4]. Many of the intermediate states that the epithelial/mesenchymal cells acquire during these transitions can be more or less stably maintained thanks to a balance between several pro- and anti-EMT elements, so that many situations of partial EMT can be encountered. This is particularly relevant in cancer, where each intermediate state could show distinct properties of invasiveness and play specific roles in the malignancy.
At the base of the dynamism of these processes and of the variety of intermediate cellular phenotypes that can be observed, there are complex molecular mechanisms resulting from the cooperation of and the interplay between numerous regulatory and effector molecules, belonging to different pathways, many of which still have to be characterized. In this context, epigenetic mechanisms controlling the expression of the EMT/MET genetic programs are becoming increasingly relevant, especially those involving non-coding RNAs.
With the rapid development of sequencing technology, numerous circular RNAs (circRNAs), structurally distinct from other coding and non-coding RNAs, have been identified in human transcriptomes. Notably, the expression of circRNAs can be cell-type and cell-state specific, indicating their complex regulation and functional role in the cell [5]. Recent studies have revealed that circRNAs contribute to the growth and progression of several human cancers. Moreover, several studies proposed some circRNAs as prognostic biomarkers because their presence and level in body fluids can predict the final output of many pathological conditions [6,7,8]. Furthermore, the finding of circRNAs in biological fluids indicates that they may also be important mediators of communication between cells. These RNAs are turning out to be important biological regulators, acting as sponges for miRNAs, as scaffolds for proteins, and also as coding molecules.
In cancer, the crosstalk between tumor cells and other elements of the tumoral microenvironment (TME), as well as between tumor cells and distant tissues targets of metastasis, has a pivotal importance for the induction of the EMT and the transition toward a malignant phenotype [9]. This intercellular communication significantly impacts on the capacity of tumoral cells to acquire invasiveness and regulates EMT plasticity. In turn, it makes the tumoral and distant tissue microenvironments favorable to the tumor growth, the colonization of secondary sites and the metastasis formation.
In EMT and cancer, cell–cell communication is importantly mediated by extracellular vesicles (EVs), an heterogeneous class of lipid-bilayer structures, including exosomes and microvesicles, that differ for size and biogenesis and are virtually produced by all types of cells. EVs’ specific cargo (including proteins, lipids, DNA and RNAs) has a relevant functional impact on receiving cells and profoundly differs between untransformed and transformed cells, as well as between epithelial cells and their counterpart that has undergone EMT [10,11,12,13]. Notably, besides other regulatory non-coding RNAs, such as microRNAs and lncRNAs, circRNAs have been recently unveiled as important elements of the exosome and microvesicle cargo, where they can also be enriched with respect to the level present in producing cells. Moreover, recent studies specifically demonstrated the key role in cancer progression and in the EMT of circRNAs embedded in the exosomes derived from tumoral cells, as well as from cells of the tumor niche.
Therefore, this review focuses on the current knowledge about exosomal circRNAs with a role in the EMT, to provide readers with a state-of-the-art understanding of this specific research topic, still largely unexplored. The relevance of circRNAs in regulating tumor progression as well as the tumor cell-microenvironment network is highlighted. Moreover, the perspective section illustrates the potential therapeutic applications that the advances in the molecular functions of exosomal circRNAs promise.
2. Cell-to-Cell Communication in Tumor Microenvironment
A tumor is a complex and dynamic collection of transformed cells and different other cellular and non-cellular components that reciprocally interact. The microenvironment that surrounds tumor cells comprehends, indeed, both different cell types and several non-cellular elements. In particular, TME include fibroblasts, endothelial cells, adipocytes, immune cells, stem cells, and acellular components such as the extracellular matrix (ECM), EVs and a variable repertoire of cytokines, chemokines and growth factors. Moreover, physical and chemical elements, such as low pH and hypoxia, are critical microenvironmental elements. Tumor cells gradually build their own TME, conditioning its components to their advantage. Therefore TME, which at first can exert an antitumor effect, is progressively modified to become a fundamental element in promoting tumor growth and progression. The complex and dynamic interplay between cancer cells and TME cellular components, mediated by secreted factors and EVs, results in local inflammation, immunosuppression and angiogenesis and, eventually, in cancer cell survival, proliferation, local invasion, and dissemination, together with the acquisition of drug resistance. The EMT of tumor cells, a crucial element of malignancy, is a relevant outcome of the crosstalk between cancer cells and TME components [14].
Primary tumors and TME cells, moreover, are also able to actively conditionate distant tissues and to establish the so-called pre-metastatic niches, where a complex interplay between cellular components and tumor-secreted elements generate a fertile soil for the following secondary tumor colonization and metastasis [15].
Notably, the informational content of tumor-derived vesicles, in particular of exosomes released by the primary tumor (tumor-derived exosomes, TDEs), for paracrine and endocrine signaling, educate receiving cells in the niche and exert a key role in the establishment of a microenvironment favorable to metastasis. Therefore, the exosomes, derived both from cancer cells and from TME cellular elements, play a crucial role in all phases of the tumor process, from onset to secondary site colonization.
Exosomes as Mediators of Intercellular Communication in Cancer
Exosomes are small extracellular vesicles with a diameter of 30–150 nm that can be produced in the endolysosomal and multivesicular body compartments by most cells and released into the extracellular environment [16]. EV complex cargo, including a wide range of biomolecules (lipids, DNA, proteins, lncRNAs, miRNAs, mRNAs, circRNAs, metabolites, and even organelles), protected from enzyme degradation by a lipid bilayer membrane, can be delivered to target cells, thus influencing their phenotype and cellular processes. The most attractive feature of exosomes is that their content reflects the pathophysiological status of the producing cells because the loading of specific molecules is finely regulated [17,18,19,20,21,22].
Several studies demonstrated the pivotal role of exosomes in cancer biology. Notably, TDEs deliver autocrine and paracrine signals that reprogram and educate the cellular components of the tumor niche, such as normal parenchymal and stromal cells, cancer-associated fibroblasts (CAFs), endothelial cells, and local immune cells, thus creating the microenvironmental conditions suitable for tumor progression [23]. Moreover, the systemic release of TDEs contributes to the establishment of the pre-metastatic niche and metastasis by reprogramming cells of secondary sites and, eventually, determining the organotropism of circulating metastatic cells and their engraftment [24].
The functional effects induced by the exosome-released cargo in recipient cells include the modulation of immunosuppression (recruitment of suppressive immune cells or inhibition of T cell activation and proliferation, impairment of the adaptive immune response), angiogenesis (activation of endothelial cells, expression/secretion of matrix metalloproteinase), local inflammation (induction of the expression/secretion of inflammatory factors from stromal and immune cells), extracellular matrix remodeling (ECM deposition by stromal cells, and exosome adhesion to ECM molecules to facilitate colonization of circulating tumor cells) [24,25].
Part of these effects can be ascribed to the EMT phenotype of the EV-producing cells in the tumoral environment. Previous studies have found that the EMT not only can occur in a subpopulation of cells within the tumor mass, triggering the generation of circulating cancer cells, but that it can also be significantly induced in other neighboring epithelial normal and transformed cells by tumor-derived exosomes, thus triggering the acquisition of a mesenchymal phenotype and metastatic properties [26], respectively. Therefore, exosomes released by EMT-induced cells can in turn confer EMT features to neighboring cells, thus producing a synergistic effect [27,28].
These overall pro-EMT signals can also be supplied by cells of the microenvironment. Among stromal cells, cancer-associated fibroblasts (CAFs) are the main cellular components of TME in most solid cancers. Several studies have described the role of CAFs in inducing EMT in tumor cells through the exosome-mediated delivery of cytokines such as TGFβ and of miRNAs, promoting cell migration, invasion, and metastasis [29,30,31,32]. In addition to stromal cells, tumor-associated macrophages (TAMs) were shown to contribute to gastric cancer cell migration through the exosome-mediated induction of EMT-related changes such as the upregulation of matrix metalloproteinases and EMT transcription factors and cytoskeleton remodeling [33]. Exosomal miRNAs from hypoxic bone marrow-derived mesenchymal stem cells (MSCs) were found to promote the invasion and metastasis of lung cancer cells by STAT3-driven EMT [34]. Cancer stem cell-derived (CSCs) exosomes were shown to accelerate the EMT-mediated tumorigenesis and metastasis in vivo of human renal clear cell carcinoma cells [35].
Therefore, cancer-related exosomes released by cancer cells, CSCs, or tumor microenvironment associated cells can contribute to a dynamic inter-cellular crosstalk, inducing and reinforcing tumor progression through the delivery of active biomolecules able to activate signaling pathways drivers of EMT.
Some of the molecules delivered by tumor-derived exosomes have been identified and include RNAs with regulative roles, such as miRNAs, lncRNAs, and circRNAs [36]. While the role of miRNAs and lncRNAs in tumor onset and progression has already been documented and largely described, the role of circRNAs in cancer-related EMT is still emerging, and its study is in full development. Recent reports revealed that cirRNAs are highly enriched and stable in exosomes compared to the producer cells [37] and closely related to cancer where they mediate multiple processes during tumor progression [38].
3. Circular RNAs (circRNAs)
Circular RNAs are single stranded, low copy, covalently closed-loop molecules. The lack of 5′ and 3′ ends renders them resistant to exonuclease-mediated degradation and they are therefore more stable than linear transcripts [39]. CircRNAs were discovered in eukaryotic cells about forty years ago [40] and found from archaea to mammals [41,42,43]; nevertheless, they were considered for a long time as simple splicing noise to be ignored [44]. However, recently a growing amount of literature pointed to the relevance of these RNAs in cell differentiation and physio-pathology. The evidence of different functional roles, coupled to the developmental stage/tissue specific expression and to evolutionarily conserved sequences, highlighted that the circRNA class does not include byproducts of the splicing process but molecules whose biogenesis is finely regulated.
3.1. Biogenesis of circRNAs
Recent advances in high-throughput RNA-seq approaches have allowed the identification of a large number of circRNAs in non-polyadenylated transcriptomes. These molecules mainly derive from exons (exonic circRNAs or ecircRNAs); however, circRNAs that consist of both exons and introns (exonic-intronic ciRNAs or EIciRNAs) or only introns (circular intronic RNAs or ciRNAs), as well as intergenic circRNAs, have been found [45].
Exonic circRNAs can derive from the pre-mRNA back-splicing mechanism (Figure 1), following the pairing of a downstream splice donor site with an upstream splice acceptor [46]. The back-splicing mechanism can be facilitated by intron looping mediated by internal base pairing, as may be in the presence of reverse complementary Alu repeats [42,47,48,49,50,51]. The interaction between distant sequences can also be favored by RNA Binding Proteins (RBPs). For instance, DHX9 acts as a negative regulator of the production of circRNAs by means of its specific binding to transcribed inverted Alu repeats [52]. Instead, RNA-binding proteins FUS and Quaking (QKI) induce the production of several circRNAs by binding introns bordering the back-splicing junctions [53,54]. The splicing factor musclebind (MBL/MBNL1) strongly and specifically binds to circMbl, a circular form that is generated from the second exon of its own mRNA; this interaction increases circRNA levels in dependance on the binding of MBL on intronic consensus sites flanking the exon [55]. Other RBPs are suggested to impact on back-splicing mechanisms, including the heterogeneous nuclear ribonucleoprotein L (HNRNPL) [56]. Interestingly, Dudekula et al. used an innovative web tool (CircInteractome, for circRNA interactome) to map the RBP-binding sites in the flanking sequences upstream and downstream of mature circRNAs. By specifically using circ_0000020, they provided evidence of multiple binding sites for HuR, FMRP, and EIF4A3 [57]. Notably, a recent work by Minzhe et al. proposed a pipeline, called Clirc, to profile circRNAs in CLIP-Seq datasets and to analyze circRNA-RBPs interactions [58].
A further mechanism of circRNA biogenesis, discovered in S. pombe, implies the formation of a lariat exon-containing precursor, whose internal splicing causes the removal of the intronic sequence and the circularization of the exon (Figure 1) [59]. This mechanism can explain circRNA biogenesis when sequences facilitating a direct back-splicing are lacking.
In ciRNA biogenesis, exons can also be circularized, retaining introns between them, thus originating EIciRNAs [60]. Furthermore, ciRNA biosynthesis may depend on a failure in debranching and requires a consensus motif containing a 7nt GU-rich element near the 5′ splice site and an 11nt C-rich element near the branchpoint site [61].
Interestingly, by applying a new chiastic clipping signal-based algorithm (CIRI) to ENCODE RNA-seq data, Gao et al. also identified intergenic circRNAs in human transcriptome [62].
From all that has been said so far, it is clear that the formation of circRNAs is a finely regulated process; therefore, how much and which circRNAs are produced by the same precursor RNA may depend on the cell type and on the particular conditions in which cells and tissue are found.
Furthermore, different circRNAs produced starting from the processing of the same precursor RNA can exert different and even opposite effects on the same cellular process. This has to be taken into consideration when mentioning, with the same name, circRNAs that may only share their origin.
3.2. Mechanisms of Action of circRNAs
CircRNAs have been described as regulating gene expression through different molecular mechanisms.
CircRNAs can exert both transcriptional and translational control of the expression of their parental genes. EIciRNAs (such as circEIF3J and circPAIP2) localize in the nucleus, where they can form a complex with U1 snRNP and Pol II at promoter regions of parental genes and regulate their transcription in cis. Notably, this function requires the interaction between the circRNA and U1 snRNA [60]. Similarly, ciRNAs can positively modulate Pol II elongation activity at their own genes [61].
CircRNAs in some cases function as negative regulators of their own linear mRNAs. Some of them, in fact, contain the translation start codon thus precluding that the protein coding transcripts from which they derived can be translated [42,63]. Furthermore, circPABPN1 (has_circ_0031288) arises from the PABPN1 mRNA and can bind to HuR, thus interfering with the capacity of this RBP to interact with PABPN1 mRNA. This suppresses its translation, modulated positively by HuR [64]. Notably, the circular transcript of the YAP gene, circYAP, forms a complex with YAP mRNA and the translation initiation site-associated proteins eIF4G and PABP. In this manner, it blocks the interaction of PABP on the 3′ tail with eIF4G on the 5′-cap, thus suppressing YAP mRNA translation initiation [65]. Another remarkable mechanism of post-transcriptional downregulation is used by circMbl. The MBL protein is a general splicing factor, playing a crucial role both in canonical splicing and in the circularization of its own mRNA, depending on its level. When the protein level is high, circMbl production increases, while the production of its own mRNA decreases. Moreover, MBL protein is massively recruited by intronic binding sites retained in the circRNA. This adds a further level of negative control of circMbl on its own MBL protein coding linear transcript [55].
Notably, circRNAs can act as decoys for proteins involved in gene regulation. CircVPS13C functions as a key regulator of pituitary adenoma growth by competitively interacting with a ribosome-binding protein of the endoplasmic reticulum membrane, thus decreasing the stability of the mRNA coding for the antiproliferative protein [66]. A further example of how circRNAs can impact gene regulation by modulating the stability of the transcript comes from circRasGEF1B. It acts as a regulator of lipopolysaccharide (LPS) response in macrophage cells by regulating the stability of ICAM-1 mRNA [67]. Otherwise, circPan3 controls self-renewal capacity of multipotent intestinal stem cells (ISCs) by protecting the mRNA of an IL-13 receptor subunit from KSRP-mediated degradation [68].
A large body of evidence has proven that circRNAs can act as miRNA sponges, thus competing with target mRNAs for their binding and interfering with the miRNA-mediated post-transcriptional regulation of gene expression. Hansen et al. showed that ciRS-7 (circular RNA sponge for miR-7, also known as CDR1as, i.e., cerebellar degeneration related protein 1 antisense circRNA), identified in human and mouse brains, contains more than 70 target sites for miR-7. CiRS-7 overexpression strongly suppresses miR-7 activity in neocortical and hippocampal neurons, resulting in an increased level of its targets [69,70]. The same model of action was demonstrated for several other circRNAs, for example: circ-SRY (for sex-determining region Y) sponges miR-138 in mice testes [69]; circ-ITCH, derived from several exons of itchy E3 ubiquitin protein ligase (ITCH), sponges miR-7, miR-17 and miR-214, in esophageal squamous cell carcinoma [71], and miR-214 also in osteogenic differentiation [72].
Moreover, other circRNA functions include the capacity of forming complexes with specific proteins by acting as scaffolds. Protein binding may result in their sequestration or in the modulation of their compartmentalization. For example, Du et al. reported that the circ-Foxo3 negatively controls the cell cycle progression by binding CDK2 and p21 and impairing their function [73]. The same circRNA interacts in the cytoplasm with the anti-senescent protein ID-1 and the transcription factor E2F1, as well as the anti-stress proteins FAK and HIF1α, impairing their anti-senescent and anti-stress roles [74]. CircACC1, instead, exerts a critical role in controlling cellular response to metabolic stress by binding to regulatory subunits of the AMPK complex and regulating its assembly [75].
Furthermore, recent evidence indicates that circRNAs can be translatable, even if they lack the 5′ cap and the 3′ poly(A) tail. Specifically, Zhang et al. demonstrated that the circular RNA transcribed by the SNF2 histone linker PHD RING helicase (SHPRH) gene can be translated into a 17 kDa SHPRH-146aa protein that protects the full-length SHPRH from degradation by the ubiquitin proteasome, leading to inhibition of cell proliferation and tumorigenicity in glioblastoma [76]. Moreover, Legnini et al. reported that circ-ZNF609 can be translated in a cap-independent manner and functions in myogenesis by controlling myoblasts proliferation [77].
Mechanisms of function of circRNAs are depicted in Figure 2.
4. Functional Role of circRNAs in EMT and Tumor Progression
Increasing evidence supports a regulatory role of circRNAs in cancer-associated EMT and are mainly referred to studies on human tissues or cellular models, even if circRNAs show a high degree of conservation across species and their mechanisms of action have been studied in different organisms, including the mouse [43].
Firstly, the expression of circRNAs is modulated during the transition, and EMT transcription factors (EMT-TFs) can be directly involved in controlling their expression. For example, in hepatocellular carcinoma, Twist positively controls circCul2 (circ10720) expression to increase vimentin levels. This circRNA, indeed, sponges miR-1246, miR-578, and miR-490-5p, targeting this mesenchymal marker [78].
Secondly, a growing amount of evidence points to the functional role of circRNAs as competing endogenous RNAs in controlling the levels of both EMT-TFs and signaling molecules involved in the induction of the transition.
In melanoma, circRNA_0084043 promotes cell proliferation, migration, and invasion by sponging miR-153-3p, then determining Snail upregulation [79]. In cervical cancer, circRNA-000284 induces tumor cell proliferation and invasion by sponging miR-506, which targets Snail-2 [80]. Similarly, circPRMT5 binds to miR-30c in cells of urothelial carcinoma of the bladder, thus modulating the levels of its target Snail [6]. Another circRNA involved in the metastatic EMT of bladder cancer is circ-HIPK3, produced from exon 2 of the homeodomain-interacting protein kinase 3 (HIPK3) gene, also known as bladder cancer-related circular RNA-2 (BCRC-2). It sponges miR-558 in bladder cancer cells [81]; interestingly, miR-558 suppresses heparinase, whose expression has been closely related to EMT [82]. Moreover, circHIPK3 serves as a sponge for multiple other miRNAs in human cells, exhibiting a pleiotropic activity [83]. In papillary thyroid cancer, circNUP214 sponges miR-145, causing the upregulation of ZEB2 and promoting cell proliferation, migration, and invasion (PTC) [84]. In non-small cell (NSC) lung cancer cells, circAGFG1 enhances proliferation, invasion, and migration by acting as competing endogenous RNA of miR-203 and positively regulating its target, ZNF281 [85]. Moreover, in breast cancer, circANKS1B (hsa_circ_0007294) sponges miR-148a-3p and miR-152-3p, leading to the increase of the expression of the transcription factor USF1, in turn activating TGF-β1/Smad signaling [86]. In breast cancer progression is also involved circ-ROBO1 (has_circ_0124696), which promotes breast cancer-derived liver metastasis by sponging miR-217-5p [87], known to target Zeb1 [88]. Furthermore, in oral squamous cell carcinoma cells, circUHRF1 (hsa_circ_0002185) exhibits a sponge activity for miR-526b-5p, thereby regulating c-Myc and promoting TGF-β1 transcription [89]. Myc is also a target for miR-449c-5p and circNOTCH1, upregulated in gastric cancer and promotes tumor growth and metastasis by interfering with miR-449c-5p/MYC/NOTCH1 axis [90].
However, circRNAs can also act as sponges to negatively regulate the EMT process. This is the case with circAKT3 (hsa_circ_0017252), repressed in clear cell renal cell carcinoma metastasis, which binds to miR-296-3p and interferes with the post-transcriptional down-regulation of E-cadherin mediated by this miRNA [91]. Moreover, the circAMOTL1L (hsa_circRNA_000350) down-regulation is required in EMT of prostate cancer cells, in the light of the sponge activity of this circRNA on miR-193a-5p, known to target a subset of cadherin superfamily members [92]. Similarly, in lung cancer cells, hsa_circ_0007059 overexpression causes up-regulation of E-cadherin and inhibition of Vimentin, Twist, and Zeb1. This circRNA impairs miR-378 activity, thus negatively regulating the EMT process via inactivation of the Wnt/β-catenin and ERK1/2 pathways [93]. Wnt/βcatenin signaling can also be interfered in triple-negative breast cancer by circ-ITCH. It was found significantly downregulated in tumor cells, and its restoration was shown to inhibit proliferation, invasion, and metastasis by sponging miR-214 and miR-17 [94]. Circ-ITCH also inhibits bladder cancer progression by sponging miRNAs targeting PTEN [95], known to suppress EMT in cancer stem cells [96]. Circ0026344 restrains EMT by suppressing miR-183-dependent Wnt/β-catenin signaling in colon cancer [97]. In small cell lung cancer cells, cESRP1 binds to miR-93-5p, upregulating its target Smad7, a negative feedback regulator of the TGF-β-mediated signaling [98]. Another oncosuppressor is circPTPRA, which, in non-small lung cancer cells, suppresses EMT in vitro and in vivo by sequestering miR-96-5p and upregulating RASSF8 [99]. An inhibitory effect on proliferation and metastasis of human cancers has been attributed to circPTK2. In non-small cell lung cancer tissues, TGF-β down-regulates circPTK2 (hsa_circ_0008305) that functions as a sponge of the pro-EMT regulators miR-429 and miR-200b-3p. Conversely, circPTK2 overexpression significantly decreases the expression of Snail and limits the invasiveness of tumor cells [100]. An anti-oncogenic effect of circPTK2 has also been observed in glioblastoma [101] and in gastric cancer where it sponges miR-196a-3p, a down-regulator of the oncosuppressor gene AATK [102]. Interestingly, from the same PTK2 pre-mRNA are derived another two circRNAs, circPTK2 (hsa_circ_0005273) and circPTK2 (hsa_circ_0003221), containing different spliced sequences and exerting oncogenic functions in colorectal [103] and bladder cancer [104] as well as laryngeal squamous cell carcinoma [105]. CircST6GALNAC6 (hsa_circ_0088708) is downregulated in bladder cancer tissues and cells. This circRNA inhibits the EMT in vitro and the metastasis process in vivo. Mechanistically, Tan et al. demonstrated that circST6GALNAC6 serves as a sponge that directly binds miR-200a-3p [106], the main regulator of EMT [107]. CircEHMT1, whose levels are strongly decreased in breast cancer tissues, inhibits migration and invasion of tumor cells targeting miR-1233-3p that, in turn, control MMP2 by means of the transcription factor KLF4 [108].
Beside sponge activity, the functional role of circRNAs can include the binding with specific proteins to modulate the activation of molecular pathways involved in EMT induction. For example, circPTK2 (hsa_circ_0005273) can promote EMT of colon cancer cells by binding to vimentin on sites Ser38, Ser55, and Ser82 [103]. Moreover, Bi et al. proposed that circRNA_102171 promotes proliferation, migration and invasion of papillary thyroid cancer cells by interacting with CTNNBIP1. This binding interferes with the association of this protein with the β-catenin/TCF complex and causes the activation of the Wnt/β-catenin pathway [109].
Furthermore, Pan et al. demonstrated that circFNDC3B negatively controls the EMT in colon cancer progression via encoding a protein, named circFNDC3B-218aa, able to inhibit Snail expression [110].
Finally, in breast cancer, circYAP is negatively regulated; this circRNA, indeed, suppresses translation initiation of YAP protein [65], a key regulator of the EMT [111].
CircRNAs involved in EMT are listed in Table 1.
5. Exosomal circRNAs in Cell-Tumor Microenvironment Crosstalk
EV-associated circRNAs exert a key role in the modulation of EMT features as mediators of communication between primary tumor cells and TME. Accordingly, circRNAs in biological fluids are valuable as non-invasive biomarkers for diagnosis of different cancer stages, independently of which EV species (often not investigated) delivers them [7]. However, growing evidence points to the exosomes as vehicles of circRNAs in the cell-tumor microenvironment crosstalk.
At first, tumor cells with high metastatic potential can donate to recipient epithelial cells exosomes whose specific cargo of circRNAs confers them EMT features and pro-metastatic properties. This is accomplished through the induction of signaling pathways triggering the transdifferentiation process or the positive modulation of mesenchymal markers. In vitro studies demonstrated that exosomes from high metastatic LM3 hepatoma cells increase the migration and invasion potential of non-metastatic (HepG2) and low metastatic (97L) cells, and these effects are blocked by circPTGR1 silencing. Mechanistically, the sponge activity of circPTGR1 on the tumor suppressor miR-449a results in the positive regulation of its target MET, in turn modulating the phenotypic change. Notably, in HCC the prognosis of patients with low exosomal circPTGR1 levels in serum is better than that of patients with high expression of this circRNA [113,127]. Low overall survival of HCC patients also correlates with high levels in the tumor of circMMP2 and low levels of miR-136-5p. In vitro circMMP2, delivered by exosomes from highly invasive cells, confers EMT properties to normal hepatocytes and nonmetastatic HepG2 cells; conversely, circMMP2 silencing in HCC cells weakens their ability to metastasize. Molecularly, it has been shown that circMMP2 causes the upregulation of MMP2 expression via sponging miR-136-5p [114]. CircRNA100338 is a further exosomal circRNA produced by highly invasive HCC cells that positively regulates the metastatic capacity of recipient hepatocytes and hepatoma cells, upregulating MMP9 expression. Accordingly, high levels of this circRNA in serum of HCC patients have been proposed as a risk indicator of metastasis and poor survival [115]. Interestingly, the exosomal circRNA-mediated upregulation of MMP2 is also involved in the process of ovarian cancer cell metastasization. CircPUM1, indeed able to upregulate MMP2 by sponging of miRNA-6753-5p and miR-615-5p, is expressed by cancer cells and transferred to peritoneum. Here, overexpressed MMP2 participates in the process of the mesothelium-to-mesenchymal transition, favoring cancer dissemination [116].
In gastric cancer, exosome-embedded circ133 is delivered from hypoxic cells to normoxic cells and promotes cell migration by acting on the miR-133a/GEF-H1/RhoA axis. Furthermore, this circRNA is enriched in the plasma exosomes of patients, and its levels increase with tumor progression [117]. Metastasis and poor prognosis in gastric cancer also correlate with circNHSL1 upregulation. Furthermore, in vitro studies have demonstrated that this circRNA has an oncogenic role by sponging miR-1306-3p and relieving the repression of its target SIX1. This transcriptional factor, in turn, induces the expression of the mesenchymal marker vimentin [118]. Interestingly, SIX1 promotes EMT of mammary carcinoma cells by increasing TGFβ signaling [128] and regulates the transdifferentiation of colorectal cancer cells by post-transcriptional activation of the EMT-TF ZEB1 [129]. Moreover, circ-0000284 expression levels are high in cholangiocarcinoma cell lines and tumor tissues. This circRNA is secreted in exosomes and delivered to surrounding normal cells to promote their migration and proliferation by acting as competitive endogenous RNA [119].
In laryngeal squamous cell carcinoma progression, the levels of circRASSF2 are significantly elevated in exosomes from plasma and tumor tissues. As reported by Tian et al., this circRNA modulates the miR-302b-3p/insulin-like growth factor 1 receptor (IGF-1R) axis, thus promoting proliferation, migration, and invasion of tumor cells [120]. Notably, miR-302 is known to induce mesenchymal-epithelial transition during reprogramming of fibroblasts and to block TGFβ-induced EMT of epithelial cells [130].
Additionally, untransformed epithelial cells can communicate via EV-associated circRNAs to in situ tumor cells to counteract their malignant behavior and promote cell apoptosis or the arrest of the cell cycle. For example, exosome-embedded circ0051443 is produced by normal hepatocytes and delivered to hepatocarcinoma cells in which it regulates BAK1 expression through competitive binding to miR-331-3p, thus blocking tumor progression. Accordingly, circ-0051443 levels are significantly lower in the plasma exosomes and tissues from HCC patients than healthy controls [121]. Of note, miR-331-3p acts as a negative regulator of EMT, migration and metastasis also in non-small-cell lung cancer, by targeting ErbB2 and VAV2 (that control Rac1, PAK1, and β-catenin) [131].
CircRNA-containing exosomes are also key regulators of the crosstalk between tumor cells and non-transformed stromal host cells, including endothelial cells, fibroblasts, and immune cells. EV-delivered circRNAs can regulate angiogenesis induction, invasion, and metastasis activation as well as immune surveillance evasion.
For example, HCC cells secrete circUHRF1 via exosomes, and high levels of this circRNA in HCC patient plasma not only correlates to poor clinical prognosis but also to NK cell dysfunction. CircUHRF1 acts as a sponge for miR-449c-5p, determining in NK cells the upregulation of its target TIM-3 that, in turn, inhibits IFN-γ and TNF-α secretion and the antitumoral immunity [112]. It can be supposed that miR-449c-5p regulation in primary tumor cells can also affect their ability to metastasize, in analogy to that reported in gastric cancer, where miR-449c-5p function is inhibited by circNOTCH1 to modulate Myc and Notch1 [90]. Interestingly, Notch signaling positively regulates the EMT, invasion, and growth of breast cancer cells by inducing Slug expression [132]. Moreover, circRNA100338, secreted in exosomes by HCC highly invasive cells, is internalized by HUVEC cells and binds to several RBPs, affecting cell proliferation, angiogenesis, and permeability [115]. A role for circASAP1 (hsa_circ_0085616) has been recently reported in HCC where it upregulates Mitogen-Activated Protein Kinase 1 expression by sponging miR-326 and miR-532-5p, thus inducing high metastatic properties in cancer cells and regulating TAM infiltration [122].
Furthermore, Xu et al. reported that circ-CCAC1 from cholangiocarcinoma-derived EVs transferred to endothelial monolayer cells induces the disruption of endothelial barrier integrity and promotes angiogenesis. Circ-CCAC1 acts by sequestering EZH2 in the cytoplasm, thereby preventing its nuclear translocation. This determines the positive regulation of SH3GL2, a negative regulator of ZO-1 and occludin. Furthermore, circ-CCAC1 regulates the miR-514a-5p/YY1 axis by means of a competitive endogenous RNA mechanism, thus controlling the proliferation and invasiveness of producing cholangiocarcinoma cells [123].
Moreover, EVs can be produced by cells in TME and internalized by tumor cells impacting their phenotype. For example, in vitro and in vivo experiments have shown that circ-DB secreted in EVs from adipocytes promotes the growth of HCC cells by inhibiting miR-34a and activating deubiquitination-related USP7 [124]. Notably, miR-34a is a key EMT regulator, inhibiting several EMT-TFs [133].
Tumor-associated macrophages (TAM) are further regulators of tumor progression through EV-associated circRNAs. In cholangiocarcinoma, proliferation, migration, and invasion are modulated by circ_0020256 conveyed by TAM-secreted exosomes. This circRNA acts via miR-432-5p sponging to determine the rescue of its-mediated repression of the transcription factor E2F3. In fact, crc_0020256 overexpression causes the downregulation of E-cadherin expression and the upregulation of N-Cadherin, and these effects can be reversed by the miR-432-5p mimics [125].
In the molecular interplay between cancer cells and the surrounding microenvironment, a role is played by CAFs, able to secrete several pro-tumoral molecules. Little is known at present about CAF-derived exosomal circRNAs. The CAF-derived exosomal circSL7A6 has been proposed to play a role in EMT by sequestering miRNA21 and the anti-EMT miRNA-200 members in colon cancer [126]. Interestingly, a role for circHIF1A, found in exosomal cargo of CAFs in breast cancer, has been recently reported to modulate the expression of CD44 and ultimately induce stemness properties to cancer cells [134].
6. Conclusions and Perspectives
EMT has a key role in tumor progression and metastasis, and this makes the molecules and signaling pathways involved in this process attractive targets for therapeutic approaches. However, this process is not of the “all or nothing” type, and the complex interplay between cells in EMT and tumor niche demands a great attention in developing possible strategies.
It is a matter of fact that the recent identification of distinct circRNAs, enriched in exosomes from cells undergoing EMT, as key mediators of the crosstalk between cells of the TME, strongly encourages the use of these molecules as novel biomarkers of invasive tumors. Moreover, while more efforts are needed to clarify the role of single molecules in invasive tumor networks, recent findings, as described above, already suggest that specific circRNAs could be conceivably addressed to achieve targeted therapeutic intervention, as previously proposed for other RNAs (i.e., miRNAs and lncRNAs) with a better documented role in cancer development and progression. A therapeutic approach targeting specific circRNAs enriched in exosomes could be able, indeed, to limit the cell-to cell communication that fosters tumor outcome. Further studies are required to clarify the contribution in EMT of circRNAs delivered by microvesicles or if they can also be delivered by exomeres in TME [135].
As discussed above, circRNAs can exert an oncogenic or a suppressive role in tumor progression. In the first case, siRNAs targeting the back-splice junction sequences may provide a simple strategy to achieve potential beneficial outcomes. Conversely, circRNAs with anti-EMT roles could be overexpressed or delivered by exosomes. Being circular molecules, they exhibit a great stability that represents an advantage with respect to linear RNAs and would allow a long-term expression at lower dosage. Interestingly, overexpression vectors, including lentivirals and adeno-associated viral vectors, have been successfully used in vitro and in vivo for expression and translation of circRNAs [136,137,138,139]. However, the more promising strategies of silencing or overexpression are based on possible EV-based therapeutic approaches, that should imply the engineering of EV-producing cells where the vesicles are enriched with the nucleic acids of interest.
This could be a limiting step, taking in account that the cellular mechanisms controlling the sorting of specific molecules are still poorly known. However, the use of nanoparticles or synthetic exosomes as carriers can be conceived for better results [140]. Interestingly, Kojima et al. reported a set of EXOsomal transfer into cell (EXOtic) devices for the efficient and customizable production of exosomes by mammalian cells and demonstrated the delivery of mRNA cargo by engineered producing cells implanted in mice [139]. Furthermore, nanoparticles have been successfully used for the delivery in mice of constructs overexpressing circEHMT1, an anti-EMT circRNA, as a therapeutic approach for breast cancer [108].
Author Contributions
Writing—original draft preparation, writing—review and editing, L.A., A.M. and C.C.; conceptualization, supervision, funding acquisition, C.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lu, W.; Kang, Y. Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef]
- Bakir, B.; Chiarella, A.M.; Pitarresi, J.R.; Rustgi, A.K. EMT, MET, Plasticity, and Tumor Metastasis. Trends Cell Biol. 2020, 30, 764–776. [Google Scholar] [CrossRef]
- Pastushenko, I.; Brisebarre, A.; Sifrim, A.; Fioramonti, M.; Revenco, T.; Boumahdi, S.; Van Keymeulen, A.; Brown, D.; Moers, V.; Lemaire, S.; et al. Identification of the tumour transition states occurring during EMT. Nature 2018, 556, 463–468. [Google Scholar] [CrossRef]
- Brabletz, S.; Schuhwerk, H.; Brabletz, T.; Stemmler, M.P. Dynamic EMT: A multi-tool for tumor progression. EMBO J. 2021, 40, e108647. [Google Scholar] [CrossRef]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-Type Specific Features of Circular RNA Expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Chen, X.; Chen, R.X.; Wei, W.S.; Li, Y.H.; Feng, Z.H.; Tan, L.; Chen, J.W.; Yuan, G.J.; Chen, S.L.; Guo, S.J.; et al. PRMT5 circular RNA promotes metastasis of urothelial carcinoma of the bladder through sponging miR-30c to induce epithelial mesenchymal transition. Clin. Cancer Res.. 2018, 24, 6319–6330. [Google Scholar] [CrossRef] [Green Version]
- Memczak, S.; Papavasileiou, P.; Peters, O.; Rajewsky, N. Identification and Characterization of Circular RNAs As a New Class of Putative Biomarkers in Human Blood. PLoS ONE 2015, 10, e0141214. [Google Scholar] [CrossRef]
- Bahn, J.H.; Zhang, Q.; Li, F.; Chan, T.M.; Lin, X.; Kim, Y.; Wong, D.T.; Xiao, X. The landscape of microRNA, Piwi-interacting RNA, and circular RNA in human saliva. Clin. Chem. 2015, 61, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Amicone, L.; Marchetti, A. Microenvironment and tumor cells: Two targets for new molecular therapies of hepatocellular carcinoma. Transl. Gastroenterol. Hepatol. 2018, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Conigliaro, A.; Cicchini, C. Exosome-Mediated Signaling in Epithelial to Mesenchymal Transition and Tumor Progression. J. Clin. Med. 2018, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Tao, S.C.; Guo, S.C. Role of extracellular vesicles in tumour microenvironment. Cell Commun. Signal. 2020, 18, 163. [Google Scholar] [CrossRef]
- Zhu, S.; Li, S.; Yi, M.; Li, N.; Wu, K. Roles of Microvesicles in Tumor Progression and Clinical Applications. Int. J. Nanomed. 2021, 16, 7071–7090. [Google Scholar] [CrossRef]
- Bian, X.; Xiao, Y.T.; Wu, T.; Yao, M.; Du, L.; Ren, S.; Wang, J. Microvesicles and chemokines in tumor microenvironment: Mediators of intercellular communications in tumor progression. Mol. Cancer 2019, 18, 50. [Google Scholar] [CrossRef]
- Anderson, N.M.; Simon, M.C. The tumor microenvironment. Curr. Biol. 2020, 30, R921–R925. [Google Scholar] [CrossRef]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Villarroya-Beltri, C.; Gutierrez-Vazquez, C.; Sanchez-Cabo, F.; Perez-Hernandez, D.; Vazquez, J.; Martin-Cofreces, N.; Martinez-Herrera, D.J.; Pascual-Montano, A.; Mittelbrunn, M.; Sanchez-Madrid, F. Sumoylated hnRNPA2B1 controls the sorting of miRNAs into exosomes through binding to specific motifs. Nat. Commun. 2013, 4, 2980. [Google Scholar] [CrossRef] [Green Version]
- Santangelo, L.; Giurato, G.; Cicchini, C.; Montaldo, C.; Mancone, C.; Tarallo, R.; Battistelli, C.; Alonzi, T.; Weisz, A.; Tripodi, M. The RNA-Binding Protein SYNCRIP Is a Component of the Hepatocyte Exosomal Machinery Controlling MicroRNA Sorting. Cell Rep. 2016, 17, 799–808. [Google Scholar] [CrossRef] [Green Version]
- Temoche-Diaz, M.M.; Shurtleff, M.J.; Nottingham, R.M.; Yao, J.; Fadadu, R.P.; Lambowitz, A.M.; Schekman, R. Distinct mechanisms of microRNA sorting into cancer cell-derived extracellular vesicle subtypes. Elife 2019, 8, e47544. [Google Scholar] [CrossRef]
- Lee, H.; Li, C.; Zhang, Y.; Zhang, D.; Otterbein, L.E.; Jin, Y. Caveolin-1 selectively regulates microRNA sorting into microvesicles after noxious stimuli. J. Exp. Med. 2019, 216, 2202–2220. [Google Scholar] [CrossRef]
- Leidal, A.M.; Huang, H.H.; Marsh, T.; Solvik, T.; Zhang, D.; Ye, J.; Kai, F.; Goldsmith, J.; Liu, J.Y.; Huang, Y.H.; et al. The LC3-conjugation machinery specifies the loading of RNA-binding proteins into extracellular vesicles. Nat. Cell Biol. 2020, 22, 187–199. [Google Scholar] [CrossRef]
- McKenzie, A.J.; Hoshino, D.; Hong, N.H.; Cha, D.J.; Franklin, J.L.; Coffey, R.J.; Patton, J.G.; Weaver, A.M. KRAS-MEK SignalingControls Ago2 Sorting into Exosomes. Cell Rep. 2016, 15, 978–987. [Google Scholar] [CrossRef] [Green Version]
- Maia, J.; Caja, S.; Strano Moraes, M.C.; Couto, N.; Costa-Silva, B. Exosome-Based Cell-Cell Communication in the Tumor Microenvironment. Front. Cell Dev. Biol. 2018, 6, 18. [Google Scholar] [CrossRef]
- Guo, Y.; Ji, X.; Liu, J.; Fan, D.; Zhou, Q.; Chen, C.; Wang, W.; Wang, G.; Wang, H.; Yuan, W.; et al. Effects of exosomes on pre-metastatic niche formation in tumors. Mol. Cancer 2019, 18, 39. [Google Scholar] [CrossRef] [Green Version]
- Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer: Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [Google Scholar] [CrossRef] [Green Version]
- Blackwell, R.H.; Foreman, K.E.; Gupta, G.N. The Role of Cancer-Derived Exosomes in Tumorigenicity & Epithelial-to-Mesenchymal Transition. Cancers 2017, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, M.; Silva, J.; Herrera, A.; Herrera, M.; Peña, C.; Martin, P.; Gil-Calderón, B.; Larriba, M.J.; Coronado, M.J.; Soldevilla, B.; et al. Exosomes enriched in stemness/metastatic-related mRNAS promote oncogenic potential in breast cancer. Oncotarget 2015, 6, 40575–40587. [Google Scholar] [CrossRef] [Green Version]
- Whiteside, T.L. The role of tumor-derived exosomes in epithelial mesenchymal transition (EMT). Transl. Cancer Res.. 2017, 6 (Suppl. 1), S90–S92. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Wang, J.; Li, M.; Cao, C.; Tan, J.; Ma, D.; Gao, Q. Tgfβ1 in Fibroblasts-Derived Exosomes Promotes Epithelial-Mesenchymal Transition of Ovarian Cancer Cells. Oncotarget 2017, 8, 96035–96047. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.L.; Wang, W.; Lan, X.L.; Zeng, Z.C.; Liang, Y.S.; Yan, Y.R.; Song, F.Y.; Wang, F.F.; Zhu, X.H.; Liao, W.J.; et al. CAFs Secreted Exosomes Promote Metastasis and Chemotherapy Resistance by Enhancing Cell Stemness and Epithelial-Mesenchymal Transition in Colorectal Cancer. Mol. Cancer 2019, 18, 91. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wei, H.; Wang, J.; Li, L.; Chen, A.; Li, Z. MicroRNA-181d-5p-Containing Exosomes Derived from CAFs Promote EMT by Regulating CDX2/HOXA5 in Breast Cancer. Mol. Ther. Nucleic Acids 2020, 19, 654–667. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Yan, Y.; Yang, Y.; Hong, X.; Wang, M.; Yang, Z.; Liu, B.; Ye, L. MiR-210 in Exosomes Derived from CAFs Promotes non-Small Cell Lung Cancer Migration and Invasion Through PTEN/PI3K/AKT Pathway. Cell. Signal. 2020, 73, 109675. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.; Luo, Q.; Wang, W.; Li, J.; Wang, T.; Wang, P.; Chen, L.; Zhang, P.; Chen, H.; Liu, Y.; et al. Tumor-associated macrophages-derived exosomes promote the migration of gastric cancer cells by transfer of functional apolipoprotein E. Cell Death Dis. 2018, 9, 434. [Google Scholar] [CrossRef]
- Zhang, X.; Sai, B.; Wang, F.; Wang, L.; Wang, Y.; Zheng, L.; Li, G.; Tang, J.; Xiang, J. Hypoxic BMSC-derived exosomal miRNAs promote metastasis of lung cancer cells via STAT3-induced EMT. Mol. Cancer 2019, 18, 40. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Yang, G.; Zhao, D.; Wang, J.; Bai, Y.; Peng, Q.; Wang, H.; Fang, R.; Chen, J.; Wang, Z.; et al. CD103-positive CSC exosome promotes EMT of clear cell renal cell carcinoma: Role of remote MiR-19b-3p. Mol. Cancer 2019, 18, 86. [Google Scholar] [CrossRef] [PubMed]
- Shrivastava, S.; Morris, K.V. The Multifunctionality of Exosomes; from the Garbage Bin of the Cell to a Next Generation Gene and Cellular Therapy. Genes 2021, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Research 2015, 25, 981–984. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Feng, W.; Kong, M.; Liu, R.; Wu, A.; Shen, L.; Tang, Z.; Wang, F. Exosomal circRNAs: A new star in cancer. Life Sci. 2021, 269, 119039. [Google Scholar] [CrossRef]
- Suzuki, H.; Tsukahara, T. A view of pre-mRNA splicing from RNase R resistant RNAs. Int. J. Mol. Sci. 2014, 15, 9331–9342. [Google Scholar] [CrossRef] [Green Version]
- Hsu, M.T.; Coca-Prados, M. Electron microscopic evidence for the circular form of RNA in the cytoplasm of eukaryotic cells. Nature 1979, 280, 339–340. [Google Scholar] [CrossRef]
- Danan, M.; Schwartz, S.; Edelheit, S.; Sorek, R. Transcriptome-wide discovery of circular RNAs in Archaea. Nucleic Acids Res. 2012, 40, 3131–3142. [Google Scholar] [CrossRef] [PubMed]
- Jeck, W.R.; Sorrentino, J.A.; Wang, K.; Slevin, M.K.; Burd, C.E.; Liu, J.; Marzluff, W.F.; Sharpless, N.E. Circular RNAs are abundant, conserved, and associated with ALU repeats. RNA 2013, 19, 141–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeck, W.R.; Sharpless, N.E. Detecting and characterizing circular RNAs. Nat. Biotechnol. 2014, 32, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Cocquerelle, C.; Mascrez, B.; Hetuin, D.; Bailleul, B. Mis-splicing yields circular RNA molecules. FASEB J. 1993, 7, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Kristensen, L.S.; Andersen, M.S.; Stagsted, L.V.W.; Ebbesen, K.K.; Hansen, T.B.; Kjems, J. The biogenesis, biology and characterization of circular RNAs. Nat. Rev. Genet 2019, 20, 675–691. [Google Scholar] [CrossRef]
- Braun, S.; Domdey, H.; Wiebauer, K. Inverse Splicing of a Discontinuous Pre-mRNA Intron Generates a Circular Exon in a HeLa Cell Nuclear Extract. Nucleic Acids Res. 1996, 24, 4152–4157. [Google Scholar] [CrossRef]
- Ivanov, A.; Memczak, S.; Wyler, E.; Torti, F.; Porath, H.T.; Orejuela, M.R.; Piechotta, M.; Levanon, E.Y.; Landthaler, M.; Dieterich, C.; et al. Analysis of intron sequences reveals hallmarks of circular RNA biogenesis in animals. Cell Rep. 2015, 10, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Liang, D.; Wilusz, J.E. Short intronic repeat sequences facilitate circular RNA production. Genes Dev 2014, 28, 2233–2247. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.O.; Wang, H.B.; Zhang, Y.; Lu, X.; Chen, L.L.; Yang, L. Complementary sequence mediated exon circularization. Cell 2014, 159, 134–147. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, Z. Efficient back splicing produces translatable circular mRNAs. RNA 2015, 21, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Starke, S.; Jost, I.; Rossbach, O.; Schneider, T.; Schreiner, S.; Hung, L.H.; Bindeiref, A. Exon circularization requires canonical splice signals. Cell Rep. 2015, 10, 103–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, T.; Ilik, I.A.; Maticzka, D.; Bhardwaj, V.; Pessoa Rodrigues, C.; Mittler, G.; Manke, T.; Backofen, R.; Akhtar, A. DHX9 suppresses RNA processing defects originating from the Alu invasion of the human genome. Nature 2017, 544, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Errichelli, L.; Dini Modigliani, S.; Laneve, P.; Colantoni, A.; Legnini, I.; Capauto, D.; Rosa, A.; De Santis, R.; Scarfò, R.; Peruzzi, G.; et al. FUS affects circular RNA expression in murine embryonic stem cell-derived motor neurons. Nat. Commun. 2017, 8, 14741. [Google Scholar] [CrossRef] [PubMed]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Fei, T.; Chen, Y.; Xiao, T.; Li, W.; Cato, L.; Zhang, P.; Cotter, M.B.; Bowden, M.; Lis, R.T.; Zhao, S.G.; et al. Genome-wide CRISPR screen identifies HNRNPL as a prostate cancer dependency regulating RNA splicing. Proc. Natl. Acad. Sci. USA 2017, 114, E5207–E5215. [Google Scholar] [CrossRef] [Green Version]
- Dudekula, D.B.; Panda, A.C.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Gorospe, M. CircInteractome: A web tool for exploring circular RNAs and their interacting proteins and microRNAs. RNA Biol. 2016, 13, 34–42. [Google Scholar] [CrossRef] [Green Version]
- Minzhe, Z.; Tao, W.; Guanghua, X.; Yang, X. Large-Scale Profiling of RBP-circRNA Interactions from Public CLIP-Seq Datasets. Genes 2020, 11, 54. [Google Scholar] [CrossRef] [Green Version]
- Barrett, S.P.; Wang, P.L.; Salzman, J. Circular RNA biogenesis can proceed through an exon-containing lariat precursor. ELife 2015, 4, e07540. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, D.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, C.W.; Chan, D.C.; Kuo, A.; Leder, P. The mouse formin (Fmn) gene: Abundant circular RNA transcripts and gene-targeted deletion analysis. Molecular Medicine 1998, 4, 614–628. [Google Scholar] [CrossRef] [Green Version]
- Abdelmohsen, K.; Panda, A.C.; Munk, R.; Grammatikakis, I.; Dudekula, D.B.; De, S.; Kim, J.; Noh, J.H.; Kim, K.M.; Martindale, J.L.; et al. Identification of HuR target circular RNAs uncovers suppression of PABPN1 translation by CircPABPN1. RNA Biol. 2017, 14, 361–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Yuan, Z.; Du, K.Y.; Fang, L.; Lyu, J.; Zhang, C.; He, A.; Eshaghi, E.; Zeng, K.; Ma, J.; et al. Translation of yes-associated protein (YAP) was antagonized by its circular RNA via suppressing the assembly of the translation initiation machinery. Cell Death Differ. 2019, 26, 2758–2773. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Chen, S.; Du, Q.; Bian, P.; Chen, Y.; Liu, Z.; Zheng, J.; Sai, K.; Mou, Y.; Chen, Z.; et al. CircVPS13C promotes pituitary adenoma growth by decreasing the stability of IFITM1 mRNA via interacting with RRBP1. Oncogene 2022, 41, 1550–1562. [Google Scholar] [CrossRef]
- Ng, W.L.; Marinov, G.K.; Liau, F.S.; Lam, Y.L.; Lim, Y.-Y.; Eaa, C.-K. Inducible RasGEF1B circular RNA is a positive regulator of ICAM-1 in the TLR4/LPS pathway. RNA Biol. 2016, 13, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Zhu, P.; Zhu, X.; Wu, J.; He, L.; Lu, T.; Wang, Y.; Liu, B.; Ye, B.; Sun, L.; Fan, D.; et al. IL-13cret seed by ILC2s promotes the self-renewal of intestinal stem cells through circular RNA circPan3. Nat. Immunol. 2019, 20, 183. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef]
- Li, F.; Zhang, L.; Li, W.; Deng, J.; Zheng, J.; An, M.; Lu, J.; Zhou, Y. Circular RNA ITCH has inhibitory effect on ESCC by suppressing the Wnt/β-catenin pathway. Oncotarget 2015, 6, 6001–6013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, D.; Xu, G.-Z.; Wu, J.-Z.; Liu, H.; Tang, J.-Y.; Wang, C.-G. Circ-ITCH sponges miR-214 to promote the osteogenic differentiation in osteoporosis via upregulating YAP1 2021, 12, 340. Cell Death Dis. 2021, 12, 340. [Google Scholar] [CrossRef] [PubMed]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, W.W.; Yang, W.; Chen, Y.; Wu, Z.K.; Foster, F.S.; Yang, Z.; Li, X.; Yang, B.B. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur. Heart J. 2017, 38, 1402–1412. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, Y.; Wu, S.; Zhou, Z.; Ding, X.; Shi, R.; Thorne, R.F.; Zhang, X.D.; Hu, W.; Wu, M. CircACC1 Regulates Assembly and Activation of AMPK Complex under Metabolic Stress. Cell Metab. 2019, 30, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Huang, N.; Yang, X.; Luo, J.; Yan, S.; Xiao, F.; Chen, W.; Gao, X.; Zhao, K.; Zhou, H.; et al. A novel protein encoded by the circular form of the SHPRH gene suppresses glioma tumorigenesis. Oncogene 2018, 37, 1805–1814. [Google Scholar] [CrossRef]
- Legnini, I.; Di Timoteo, G.; Rossi, F.; Morlando, M.; Briganti, F.; Sthandier, O.; Fatica, A.; Santini, T.; Andronache, A.; Wade, M.; et al. Circ-ZNF609 Is a Circular RNA that Can Be Translated and Functions in Myogenesis. Mol. Cell 2017, 66, 22–37.e9. [Google Scholar] [CrossRef] [Green Version]
- Meng, J.; Chen, S.; Han, J.X.; Qian, B.; Wang, X.R.; Zhong, W.L.; Qin, Y.; Zhang, H.; Ga, W.F.; Lei, Y.Y.; et al. Twist1 regulates vimentin through Cul2 circular RNA to promote EMT in hepatocellular carcinoma. Cancer Res. 2018, 78, 4150–4162. [Google Scholar] [CrossRef] [Green Version]
- Luan, W.; Shi, Y.; Zhou, Z.; Xia, Y.; Wang, J. circRNA_0084043 promote malignant melanoma progression via miR-153-3p/Snail axis. Biochem. Biophys. Res. Commun. 2018, 502, 22–29. [Google Scholar] [CrossRef]
- Ma, H.-B.; Yao, Y.-N.; Yu, J.-J.; Chen, X.-X.; Li, H.-F. Extensive profiling of circular RNAs and the potential regulatory role of circRNA-000284 in cell proliferation and invasion of cervical cancer via sponging miR-506. Am. J. Transl. Res. 2018, 10, 592–604. [Google Scholar]
- Li, Y.; Zheng, F.; Xiao, X.; Xie, F.; Tao, D.; Huang, C.; Liu, D.; Wang, M.; Wang, L.; Zeng, F.; et al. CircHIPK3 sponges miR-558 to suppress heparanase expression in bladder cancer cells. EMBO Rep. 2017, 18, 1646–1659. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.; Fourgeaud, C.; Derieux, S.; Mirshahi, S.; Contant, G.; Pimpie, C.; Lo Dico, R.; Soria, J.; Pocard, M.; Mirshahi, M. The close relationship between heparanase and epithelial mesenchymal transition in gastric signet-ring cell adenocarcinoma. Oncotarget 2018, 9, 33778–33787. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Bao, C.; Guo, W.; Li, S.; Chen, J.; Chen, B.; Luo, Y.; Lyu, D.; Li, Y.; Shi, G.; et al. Circular RNA profiling reveals an abundant circHIPK3 that regulates cell growth by sponging multiple miRNAs. Nat. Commun. 2016, 7, 11215. [Google Scholar] [CrossRef]
- Li, X.; Tian, Y.; Hu, Y.; Yang, Z.; Zhang, L.; Luo, J. CircNUP214 sponges miR-145 to promote the expression of ZEB2 in thyroid cancer cells. Biochem. Biophys. Res. Commun. 2018, 507, 168–172. [Google Scholar] [CrossRef]
- Xue, Y.B.; Ding, M.Q.; Xue, L.; Luo, J.H. CircAGFG1 sponges miR-203 to promote EMT and metastasis of non-small-cell lung cancer by upregulating ZNF281 expression. Thorac. Cancer 2019, 10, 1692–1701. [Google Scholar] [CrossRef]
- Zeng, K.; He, B.; Yang, B.B.; Xu, T.; Chen, X.; Xu, M.; Liu, X.; Sun, H.; Pan, Y.; Wang, S. The prometastasis effect of circANKS1B in breast cancer. Mol. Cancer 2018, 17, 160. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, L.; Wu, P.; Li, X.; Tang, Y.; Ou, X.; Zhang, Y.; Xiao, X.; Wang, J.; Tang, H. The circROBO1/KLF5/FUS feedback loop regulates the liver metastasis of breast cancer by inhibiting the selective autophagy of afadin. Mol. Cancer 2022, 21, 29. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wang, B.; Luo, H.; Zhang, Q.; Xu, M. miR-217 represses TGF-β1-induced airway smooth muscle cell proliferation and migration through targeting ZEB1. Biomed. Pharmacother. 2018, 108, 27–35. [Google Scholar] [CrossRef]
- Zhao, W.; Cui, Y.; Liu, L.; Qi, X.; Liu, J.; Ma, S.; Hu, X.; Zhang, Z.; Wang, Y.; Li, H.; et al. Splicing factor derived circular RNA circUHRF1 accelerates oral squamous cell carcinoma tumorigenesis via feedback loop. Cell Death Differ. 2019, 27, 919–933. [Google Scholar] [CrossRef]
- Zhao, X.; Zhong, Q.; Cheng, X.; Wang, S.; Wu, R.; Leng, X.; Shao, L. miR-449c-5p availability is antagonized by circ-NOTCH1 for MYC-induced NOTCH1 upregulation as well as tumor metastasis and stemness in gastric cancer. J. Cell Biochem. 2020, 121, 4052–4063. [Google Scholar] [CrossRef]
- Xue, D.; Wang, H.; Chen, Y.; Shen, D.; Lu, J.; Wang, M.; Zebibula, A.; Xu, L.; Wu, H.; Li, G.; et al. Circ-AKT3 inhibits clear cell renal cell carcinoma metastasis via altering miR-296-3p/E-cadherin signals. Mol. Cancer 2019, 18, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z.; Qu, C.-B.; Zhang, Y.; Zhang, W.-F.; Wang, D.-D.; Gao, C.-C.; Ma, L.; Chen, J.-S.; Liu, K.-L.; Zheng, B.; et al. Dysregulation of p53-RBM25-mediated circAMOTL1L biogenesis contributes to prostate cancer progression through the circAMOTL1L-miR-193a-5p-Pcdha pathway. Oncogene 2019, 38, 2516–2532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, S.; Yu, Y.; Liu, L.; Meng, J.; Li, G. Circular RNA hsa_circ_0007059 restrains proliferation and epithelial-mesenchymal transition in lung cancer cells via inhibiting microRNA-378. Life Sci. 2019, 233, 116692. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.T.; Liu, L.B.; Li, X.M.; Wang, Y.F.; Xie, P.J.; Li, Q.; Wang, R.; Wei, Q.; Kang, Y.H.; Meng, R.; et al. Circ-ITCH regulates triple-negative breast cancer progression through the Wnt/beta-catenin pathway. Neoplasma 2019, 66, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yuan, W.; Yang, X.; Li, P.; Wang, J.; Han, J.; Tao, J.; Li, P.; Yang, H.; Lv, Q.; et al. Circular RNA circ-ITCH inhibits bladder cancer progression by sponging miR-17/miR-224 and regulating p21, PTEN expression. Mol. Cancer 2018, 17, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Liu, J.; Chao, J.; Scheuerman, M.P.; Rahimi, S.A.; Lee, L.Y.; Li, S. PTEN suppresses epithelial-mesenchymal transition and cancer stem cell activity by downregulating Abi1. Sci. Rep. 2020, 10, 12685. [Google Scholar] [CrossRef]
- Shen, T.; Cheng, X.; Liu, X.; Xia, C.; Zhang, H.; Pan, D.; Zhang, X.; Li, Y. Circ_0026344 restrains metastasis of human colorectal cancer cells via miR-183. Artif. Cells Nanomed. Biotechnol. 2019, 47, 4038–4045. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Yang, Y.; Wu, J.; Niu, Y.; Yao, Y.; Zhang, J.; Huang, X.; Liang, S.; Chen, R.; Chen, S.; et al. Circular RNA cESRP1 sensitises small cell lung cancer cells to chemotherapy by sponging miR-93-5p to inhibit TGF-β signalling. Cell Death Differ. 2020, 27, 1709–1727. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Zheng, Y.; Jiang, Y.; Li, X.; Geng, J.; Shen, Y.; Li, Q.; Wang, X.; Zhao, C.; Chen, Y.; et al. The circRNA circPTPRA suppresses epithelial-mesenchymal transitioning and metastasis of NSCLC cells by sponging miR-96-5p. EBioMedicine 2019, 44, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Tong, X.; Zhou, Z.; Wang, S.; Lei, Z.; Zhang, T.; Liu, Z.; Zeng, Y.; Li, C.; Zhao, J.; et al. Circular RNA hsa_circ_0008305 (circPTK2) inhibits TGF-β-induced epithelial-mesenchymal transition and metastasis by controlling TIF1γ in non-small cell lung cancer. Mol. Cancer 2018, 17, 140. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Wang, N.; Lian, M. CircRNA circPTK2 Might Suppress Cancer Cell Invasion and Migration of Glioblastoma by Inhibiting miR-23a Maturation. Neuropsychiatr. Dis. Treat 2021, 17, 2767–2774. [Google Scholar] [CrossRef] [PubMed]
- Ling, G.; Tingting, X.; Mingde, Q.; Xiaofeng, X.; Linhua, J.; Xinguo, Z. CircPTK2 Suppresses the Progression of Gastric Cancer by Targeting the MiR-196a-3p/AATK Axis. Front. Oncol. 2021, 11, 706415. [Google Scholar] [CrossRef]
- Yang, H.; Li, X.; Meng, Q.; Sun, H.; Wu, S.; Hu, W.; Liu, G.; Li, X.; Yang, Y.; Chen, R. CircPTK2 (hsa_circ_0005273) as a novel therapeutic target for metastatic colorectal cancer. Mol. Cancer 2020, 19, 13. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.-Q.; Yang, M.-G.; Liu, H.-J.; Su, C.-Q. Circular RNA hsa_circ_0003221 (circPTK2) promotes the proliferation and migration of bladder cancer cells. J. Cell Biochem. 2018, 119, 3317–3325. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jin, J.; Chang, T. CircPTK2 (hsa_circ_0003221) Contributes to Laryngeal Squamous Cell Carcinoma by the miR-1278/YAP1 Axis. J. Oncol. 2021, 13, 2408384. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Kang, Y.; Li, H.; He, H.-Q.; Zheng, L.; Wu, S.-Q.; Ai, K.; Zhang, L.; Xu, R.; Zhang, X.-Z.; et al. circST6GALNAC6 suppresses bladder cancer metastasis by sponging miR-200a-3p to modulate the STMN1/EMT axis. Cell Death Dis. 2021, 12, 168. [Google Scholar] [CrossRef] [PubMed]
- Gregory, P.A.; Bert, A.G.; Paterson, E.L.; Barry, S.C.; Tsykin, A.; Farshid, G.; Vadas, M.A.; Khew-Goodall, Y.; Goodall, G.J. The miR-200 family and miR-205 regulate epithelial to mesenchymal transition by targeting ZEB1 and SIP1. Nat. Cell Biol. 2008, 10, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Wu, Y.; Zeng, B.; Sun, J.; Li, Y.; Luo, J.; Wang, L.; Yi, Z.; Li, H.; Ren, G. CircEHMT1 inhibits metastatic potential of breast cancer cells by modulating miR-1233-3p/KLF4/MMP2 axis. Biochem. Biophys. Res. Commun. 2020, 526, 306–313. [Google Scholar] [CrossRef]
- Bi, W.; Huang, J.; Nie, C.; Liu, B.; He, G.; Han, J.; Pang, R.; Ding, Z.; Xu, J.; Zhang, J. CircRNA circRNA_102171 promotes papillary thyroid cancer progression through modulating CTNNBIP1-dependent activation of beta-catenin pathway. J. Exp. Clin. Cancer Res. 2018, 37, 275. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Cai, J.; Lin, J.; Zhou, H.; Peng, J.; Liang, J.; Xia, L.; Yin, Q.; Zou, B.; Zheng, J.; et al. A novel protein encoded by circFNDC3B inhibits tumor progression and EMT through regulating Snail in colon cancer. Mol. Cancer 2020, 19, 71. [Google Scholar] [CrossRef] [Green Version]
- Noce, V.; Battistelli, C.; Cozzolino, A.M.; Consalvi, V.; Cicchini, C.; Strippoli, R.; Tripodi, M.; Marchetti, A.; Amicone, L. YAP integrates the regulatory Snail/HNF4α circuitry controlling epithelial/hepatocyte differentiation. Cell Death Dis. 2019, 10, 768. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.-F.; Gao, C.; Huang, X.-Y.; Lu, J.-C.; Guo, X.-J.; Shi, G.-M.; Cai, J.-B.; Ke, A.-W. Cancer cell-derived exosomal circUHRF1 induces natural killer cell exhaustion and may cause resistance to anti-PD1 therapy in hepatocellular carcinoma. Mol. Cancer 2020, 19, 110. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Liu, W.; Zou, Y.; Wang, G.; Deng, Y.; Luo, J.; Zhang, Y.; Li, H.; Zhang, Q.; Yang, Y.; et al. Three isoforms of exosomal circPTGR1 promote hepatocellular carcinoma metastasis via the miR449a-MET pathway. EBioMedicine 2019, 40, 432–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Kang, H.; Gao, M.; Jin, L.; Zhang, F.; Chen, D.; Li, M.; Xiao, L. Exosome-transmitted circ_MMP2 promotes hepatocellular carcinoma metastasis by upregulating MMP2. Mol. Oncol. 2020, 14, 1365–1380. [Google Scholar] [CrossRef]
- Huang, X.Y.; Huang, Z.L.; Huang, J.; Xu, B.; Huang, X.Y.; Xu, Y.H.; Zhou, J.; Tang, Z.Y. Exosomal circRNA-100338 promotes hepatocellular carcinoma metastasis via enhancing invasiveness and angiogenesis. J. Exp. Clin. Cancer Res. 2020, 39, 20. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Zong, Z.-H.; Liu, Y.; Chen, S.; Wang, L.-L.; Zhao, Y. circPUM1 Promotes Tumorigenesis and Progression of Ovarian Cancer by Sponging miR-615-5p and miR-6753-5p. Mol. Ther.—Nucleic Acids 2019, 18, 882–892. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Zhang, H.; Yang, Y.; Wang, X.; Deng, T.; Liu, R.; Ning, T.; Bai, M.; Li, H.; Zhu, K.; et al. Hypoxia induced exosomal circRNA promotes metastasis of Colorectal Cancer via targeting GEF-H1/RhoA axis. Theranostics 2020, 10, 8211–8226. [Google Scholar] [CrossRef]
- Zhu, Z.; Rong, Z.; Luo, Z.; Yu, Z.; Zhang, J.; Qiu, Z.; Huan, C. Circular RNA circNHSL1 promotes gastric cancer progression through the miR-1306- 3p/SIX1/vimentin axis. Mol. Cancer 2019, 18, 126. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Hu, Y.; Lv, X.; Li, B.; Gu, D.; Li, Y.; Sun, Y.; Su, Y. Circ-0000284 arouses malignant phenotype of cholangiocarcinoma cells and regulates the biological functions of peripheral cells through cellular communication. Clin. Sci. 2019, 133, 1935–1953. [Google Scholar] [CrossRef]
- Tian, L.; Cao, J.; Jiao, H.; Zhang, J.; Ren, X.; Liu, X.; Liu, M.; Sun, Y. CircRASSF2 promotes laryngeal squamous cell carcinoma progression by regulating the miR-302b-3p/IGF-1R axis. Clin. Sci. 2019, 133, 1053–1066. [Google Scholar] [CrossRef]
- Chen, W.; Quan, Y.; Fan, S.; Wang, H.; Liang, J.; Huang, L.; Chen, L.; Liu, Q.; He, P.; Ye, Y. Exosome-transmitted circular RNA hsa_circ_0051443 suppresses hepatocellular carcinoma progression. Cancer Lett. 2020, 475, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.-Q.; Zhou, S.-L.; Li, J.; Zhou, Z.-J.; Wang, P.-C.; Xin, H.-Y.; Mao, L.; Luo, C.-B.; Yu, S.-Y.; Huang, X.-W.; et al. Circular RNA Sequencing Identifies CircASAP1 as a Key Regulator in Hepatocellular Carcinoma Metastasis. Hepatology 2020, 72, 906–922. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Leng, K.; Yao, Y.; Kang, P.; Liao, G.; Han, Y.; Shi, G.; Ji, D.; Huang, P.; Zheng, W.; et al. A Circular RNA, Cholangiocarcinoma-Associated Circular RNA 1, Contributes to Cholangiocarcinoma Progression, Induces Angiogenesis, and Disrupts Vascular Endothelial Barriers. Hepatology 2021, 73, 1419–1435. [Google Scholar] [CrossRef]
- Zhang, H.; Deng, T.; Ge, S.; Liu, Y.; Bai, M.; Zhu, K.; Fan, Q.; Li, J.; Ning, T.; Tian, F.; et al. Exosome circRNA secreted from adipocytes promotes the growth of hepatocellular carcinoma by targeting deubiquitination-related USP7. Oncogene 2019, 38, 2844–2859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Chen, Z.; Li, Z.; Li, S.; Wen, Z.; Cao, L.; Chen, Y.; Xue, P.; Li, H.; Zhang, D. Tumor-associated macrophages promote cholangiocarcinoma progression via exosomal Circ_0020256. Cell Death Dis. 2022, 13, 94. [Google Scholar] [CrossRef]
- Chen, G.; Huigang, L.; Zhengha, Q. Matrine reduces the secretion of exosomal circSLC7A6 from cancer-associated fibroblast to inhibit tumorigenesis of colorectal cancer by regulating CXCR5. Biochem. Biophys. Res. Commun. 2020, 527, 638–645. [Google Scholar] [CrossRef]
- Chen, S.-P.; Liu, B.-X.; Xu, J.; Pei, X.-F.; Liao, Y.-J.; Yuan, F.; Zheng, F. MiR-449a suppresses the epithelial-mesenchymal transition and metastasis of hepatocellular carcinoma by multiple targets. BMC Cancer 2015, 15, 706. [Google Scholar] [CrossRef] [Green Version]
- Micalizzi, D.S.; Christensen, K.L.; Jedlicka, P.; Coletta, R.D.; Barón, A.E.; Harrell, J.C.; Horwitz, K.B.; Billheimer, D.; Heichman, K.A.; Welm, A.L.; et al. The SIX1 homeoprotein induces human mammary carcinoma cells to undergo epithelial-mesenchymal transition and metastasis in mice through increasing TGF-beta signaling. J. Clin. Investig. 2009, 119, 2678–2690. [Google Scholar] [CrossRef]
- Ono, H.; Imoto, I.; Kozaki, K.; Tsuda, H.; Matsui, T.; Kurasawa, Y.; Muramatsu, T.; Sugihara, K.; Inazawa, J. SIX1 promotes epithelial-mesenchymal transition in colorectal cancer through ZEB1 activation. Oncogene 2012, 31, 4923–4934. [Google Scholar] [CrossRef] [Green Version]
- Subramanyam, D.; Lamouille, S.; Judson, R.L.; Liu, J.Y.; Bucay, N.; Derynck, R.; Blelloch, R. Multiple targets of miR-302 and miR-372 promote reprogramming of human fibroblasts to induced pluripotent stem cells. Nat. Biotechnol. 2011, 29, 443–448. [Google Scholar] [CrossRef]
- Li, X.; Zhu, J.; Liu, Y.; Duan, C.; Chang, R.; Zhang, C. MicroRNA-331-3p inhibits epithelial-mesenchymal transition by targeting ErbB2 and VAV2 through the Rac1/PAK1/β-catenin axis in non-small-cell lung cancer. Cancer Sci. 2019, 110, 1883–1896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, S.; Zhao, X.; Zhang, X.; Luo, M.; Zuo, X.; Huang, S.; Wang, Y.; Gu, S.; Zhao, X. Notch1 signaling regulates the epithelial-mesenchymal transition and invasion of breast cancer in a Slug-dependent manner. Mol. Cancer 2015, 14, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, D.; Fu, J.; Chen, H.; Cheng, J.; Fu, J. Roles of MicroRNA-34a in Epithelial to Mesenchymal Transition, Competing Endogenous RNA Sponging and Its Therapeutic Potential. Int. J. Mol. Sci. 2019, 20, 861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, Y.; Du, J.; Min, Z.; Ma, L.; Zhang, W.; Zhu, W.; Liu, Y. Carcinoma-associated fibroblasts derived exosomes modulate breast cancer cell stemness through exonic circHIF1A by miR-580-5p in hypoxic stress. Cell Death Discov. 2021, 7, 141. [Google Scholar] [CrossRef]
- Zhang, Q.; Higginbotham, J.N.; Jeppesen, D.K.; Yang, Y.P.; Li, W.; McKinley, E.T.; Graves-Deal, R.; Ping, J.; Britain, C.M.; Dorsett, K.A.; et al. Transfer of Functional Cargo in Exomeres. Cell Rep. 2019, 27, 940–954.e6. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Du, W.W.; Li, X.; Yee, A.J.; Yang, B.B. Foxo3 activity promoted by non-coding effects of circular RNA and Foxo3 pseudogene in the inhibition of tumor growth and angiogenesis. Oncogene 2016, 35, 3919–3931. [Google Scholar] [CrossRef]
- Meganck, R.M.; Borchardt, E.K.; Castellanos Rivera, R.M.; Scalabrino, M.L.; Wilusz, J.E.; Marzluff, W.F.; Asokan, A. Tissue-Dependent Expression and Translation of Circular RNAs with Recombinant AAV Vectors In Vivo. Mol. Ther. Nucleic Acids 2018, 13, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Lavenniah, A.; Luu, T.D.A.; Li, Y.P.; Lim, T.B.; Jiang, J.; Ackers-Johnson, M.; Foo, R.S. Engineered Circular RNA Sponges Act as miRNA Inhibitors to Attenuate Pressure Overload-Induced Cardiac Hypertrophy. Mol. Ther. 2020, 28, 1506–1517. [Google Scholar] [CrossRef]
- Kojima, R.; Bojar, D.; Rizzi, G.; Hamri, G.C.; El-Baba, M.D.; Saxena, P.; Auslander, S.; Tan, K.R.; Fussenegger, M. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson‘s disease treatment. Nat. Commun. 2018, 9, 1305. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.U.; Park, I.K.; Lee, Y.K.; Hwang, S.R. The Biological Function and Therapeutic Potential of Exosomes in Cancer: Exosomes as Efficient Nanocommunicators for Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 7363. [Google Scholar] [CrossRef]
Figure 1.
Biogenesis of circular RNAs. Pre-mRNA can be processed via canonical splicing; however, a lariat-driven RNA circularization can result in the formation of ecircRNAs, intron lariat, and linear mRNAs (A). Circularization in EIciRNAs or ecircRNA can depend on the bridge formed by RBP pairing (B) or be triggered by internal base pairing (e.g., for inverted homologous Alu repeats) (C). Lariat formation of ciRNAs can be favored by GU and C-rich elements (D).
Figure 1.
Biogenesis of circular RNAs. Pre-mRNA can be processed via canonical splicing; however, a lariat-driven RNA circularization can result in the formation of ecircRNAs, intron lariat, and linear mRNAs (A). Circularization in EIciRNAs or ecircRNA can depend on the bridge formed by RBP pairing (B) or be triggered by internal base pairing (e.g., for inverted homologous Alu repeats) (C). Lariat formation of ciRNAs can be favored by GU and C-rich elements (D).

Figure 2.
Functional mechanisms of circRNAs. EIciNAs can form a complex with U1 small nuclear ribonucleoprotein (U1 snRNP) and bind RNA PolII to enhance transcription (A). CiRNAs can interact with RNA PolII to positively modulate transcription (B). CircRNAs can act as sponges of RBPs, thus impacting their pleiotropic functions (C); they can control translation initiation (D), protein sequestering, localization or function (E). Moreover, circRNAs can affect mRNA stability (F); sponge miRNAs, thus interfering with the post-transcriptional regulation of their target mRNAs (G), or act as scaffold for proteins (H). Some circRNAs can also be translated (I).
Figure 2.
Functional mechanisms of circRNAs. EIciNAs can form a complex with U1 small nuclear ribonucleoprotein (U1 snRNP) and bind RNA PolII to enhance transcription (A). CiRNAs can interact with RNA PolII to positively modulate transcription (B). CircRNAs can act as sponges of RBPs, thus impacting their pleiotropic functions (C); they can control translation initiation (D), protein sequestering, localization or function (E). Moreover, circRNAs can affect mRNA stability (F); sponge miRNAs, thus interfering with the post-transcriptional regulation of their target mRNAs (G), or act as scaffold for proteins (H). Some circRNAs can also be translated (I).

Figure 3.
Exosome-mediated cell-to-cell communication in TME. Exosomes-associated pro- and anti-EMT circRNAs in the communication between tumor cells and surrounding microenvironmental elements, including normal epithelial cells, tumor-associated macrophages, endothelial cells, cancer-associated fibroblasts, and adipocytes.
Figure 3.
Exosome-mediated cell-to-cell communication in TME. Exosomes-associated pro- and anti-EMT circRNAs in the communication between tumor cells and surrounding microenvironmental elements, including normal epithelial cells, tumor-associated macrophages, endothelial cells, cancer-associated fibroblasts, and adipocytes.


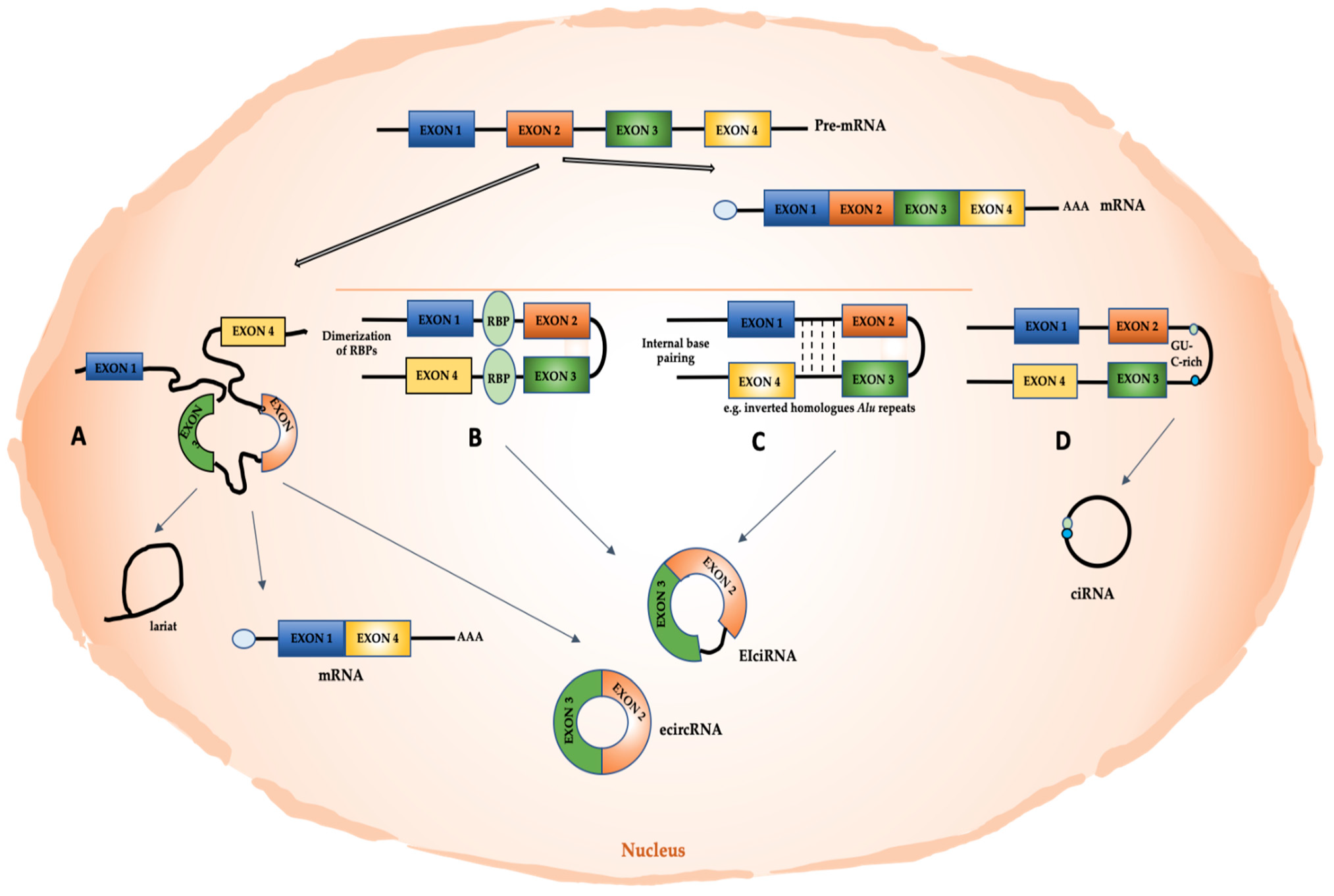{kind=link}
{kind=link}
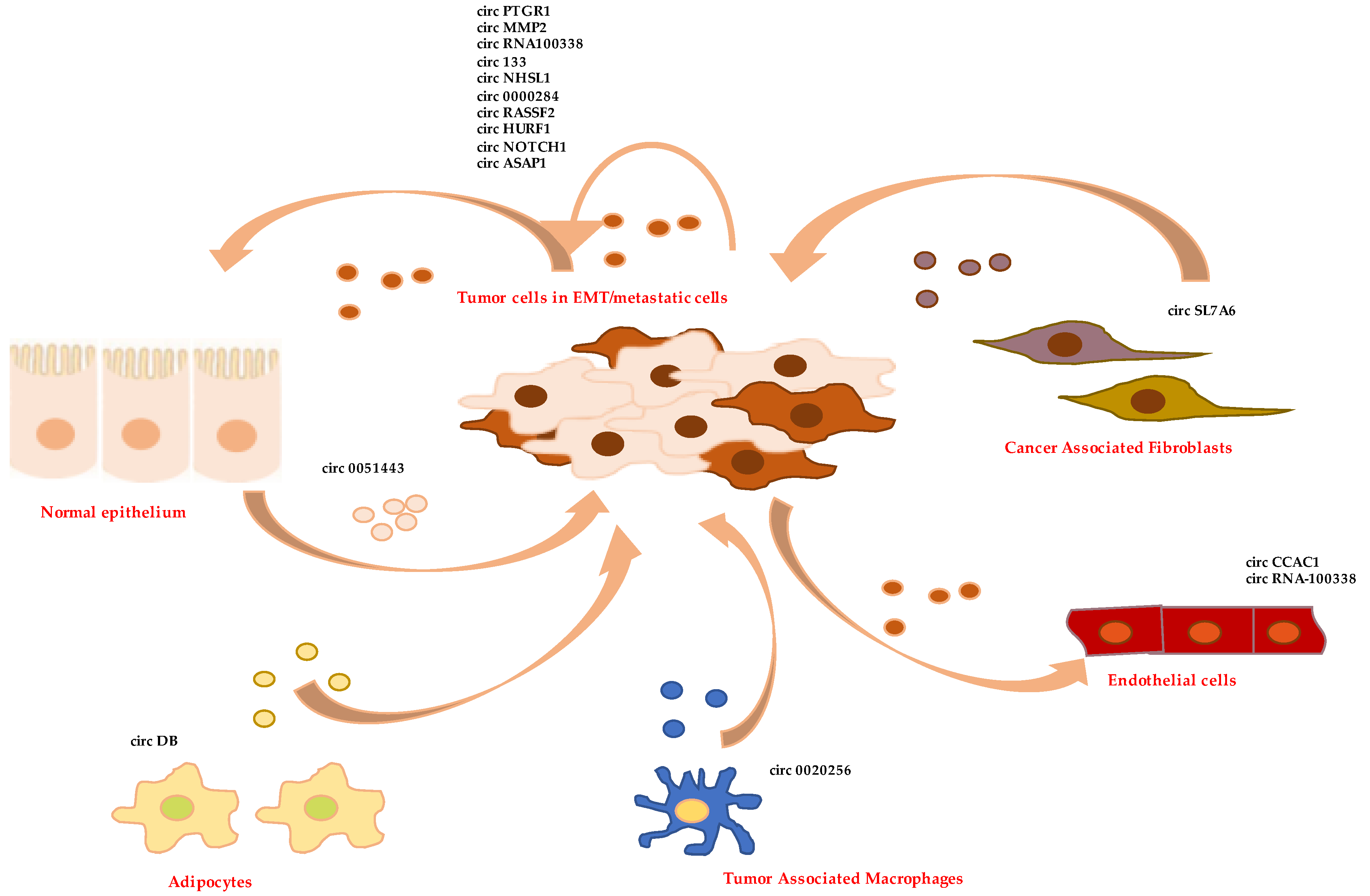{kind=link}
Table 1.
CircRNAs with a role in EMT and tumor progression.
| circRNA | Putative Function | Type of Cancer/EMT Cells | Effect on EMT/Metastasis | Localization | Reference |
|---|---|---|---|---|---|
| circPRMT5 | miRNA sponge | bladder carcinoma | pro | not investigated | [6] |
| circYAP | translation initiation | breast cancer | anti | not investigated | [65] |
| circCul2 | miRNA sponge | HCC | pro | not investigated | [78] |
| circ_0084043 | miRNA sponge | melanoma | pro | not investigated | [79] |
| circRNA-000284 | miRNA sponge | cervical cancer | pro | not investigated | [80] |
| circHIPK3 | miRNA sponge | different cancers | pro | not investigated | [81,83] |
| circNUP214 | miRNA sponge | thyroid cancer | pro | not investigated | [84] |
| circAGFG1 | miRNA sponge | NSC lung cancer | pro | not investigated | [85] |
| circANKS1B | miRNA sponge | breast cancer | pro | not investigated | [86] |
| circROBO1 | miRNA sponge | breast cancer | pro | not investigated | [87] |
| circUHRF1 | miRNA sponge | oral squamous carcinoma/HCC | pro | not investigated/ exosome | [89,112] |
| circNOTCH1 | miRNA sponge | gastric cancer | pro | exosome | [90] |
| circAKT3 | miRNA sponge | clear renal cancer | anti | not investigated | [91] |
| circAMOTL1L | miRNA sponge | prostate cancer | anti | not investigated | [92] |
| hsa_circ_0007059 | miRNA sponge | lung cancer | anti | not investigated | [93] |
| circITCH | miRNA sponge | TNBC/bladder carcinoma | anti | not investigated | [94,95] |
| circ_0026344 | miRNA sponge | colon cancer | anti | not investigated | [97] |
| circESRP1 | miRNA sponge | small cell lung cancer | anti | not investigated | [98] |
| circPTPRA | miRNA sponge | NSC lung cancer | anti | not investigated | [99] |
| circPTK2 (hsa_circ_0008305) | miRNA sponge | different cancers | anti | not investigated | [100,101,102] |
| circPTK2 (hsa_circ_0005273) | protein binding | colon cancer | pro | not investigated | [103] |
| circPTK2 (hsa_circ_0003221) | miRNA sponge | different cancers | pro | not investigated | [104,105] |
| circST6GALNAC6 (hsa_circ_0088708) | miRNA sponge | bladder cancer | anti | not investigated | [106] |
| circEHMT1 | miRNA sponge | breast cancer | anti | not investigated | [108] |
| circRNA_102171 | decoy for protein | papillary thyroid cancer | pro | not investigated | [109] |
| circFNDC3B | translated in protein | colon cancer | anti | not investigated | [110] |
| circPTGR1 | miRNA sponge | HCC | pro | exosome | [113] |
| circMMP2 | miRNA sponge | HCC | pro | exosome | [114] |
| circRNA_100338 | not investigated | HCC | pro | exosome | [115] |
| circPUM1 | miRNA sponge | ovarian cancer | pro | exosome | [116] |
| circ133 | miRNA sponge | gastric cancer | pro | exosome | [117] |
| circNHSL1 | miRNA sponge | gastric cancer | pro | exosome | [118] |
| circ-0000284 | not investigated | cholangiocarcinoma | pro | exosome | [119] |
| circRASSF2 | miRNA sponge | laryngeal squamous cell carcinoma | pro | exosome | [120] |
| hsa_circ_0051443 | miRNA sponge | HCC | anti | hepatocyte-secreted exosome | [121] |
| circASAP1 | miRNA sponge | HCC | pro | exosome | [122] |
| circCCAC1 | protein decoy and miRNA sponge | cholangiocarcinoma | pro | exosome | [123] |
| circ-DB | miRNA sponge | HCC | pro | adipocyte-secreted exosomes | [124] |
| circ_0020256 | miRNA sponge | cholangiocarcinoma | pro | TAM-secreted exosomes | [125] |
| circSL7A6 | miRNA sponge | colon cancer | pro | CAF-secreted exosomes | [126] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Amicone, L.; Marchetti, A.; Cicchini, C. Exosome-Associated circRNAs as Key Regulators of EMT in Cancer. Cells 2022, 11, 1716. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101716
AMA Style
Amicone L, Marchetti A, Cicchini C. Exosome-Associated circRNAs as Key Regulators of EMT in Cancer. Cells. 2022; 11(10):1716. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101716
Chicago/Turabian StyleAmicone, Laura, Alessandra Marchetti, and Carla Cicchini. 2022. "Exosome-Associated circRNAs as Key Regulators of EMT in Cancer" Cells 11, no. 10: 1716. https://0-doi-org.brum.beds.ac.uk/10.3390/cells11101716
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.