Sex Differences in High Fat Diet-Induced Metabolic Alterations Correlate with Changes in the Modulation of GRK2 Levels

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Metabolic Assays
2.3. Tissue Collection and Processing
2.4. Western Blotting
2.5. Statistical Analysis
3. Results
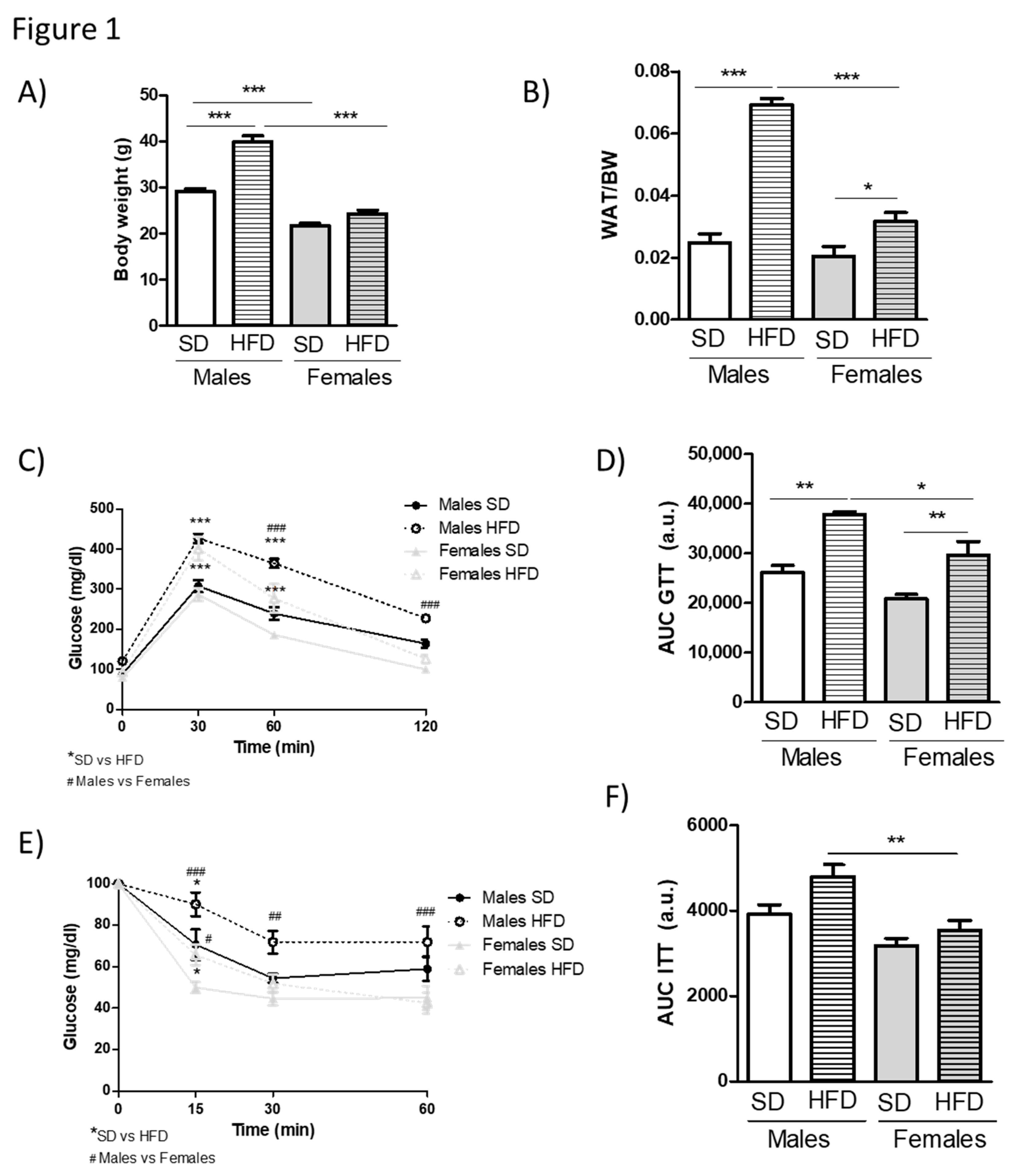
3.1. HFD Feeding Causes More Pronounced Obesity and Metabolic Alterations in Young Male Than in Young Female Mice
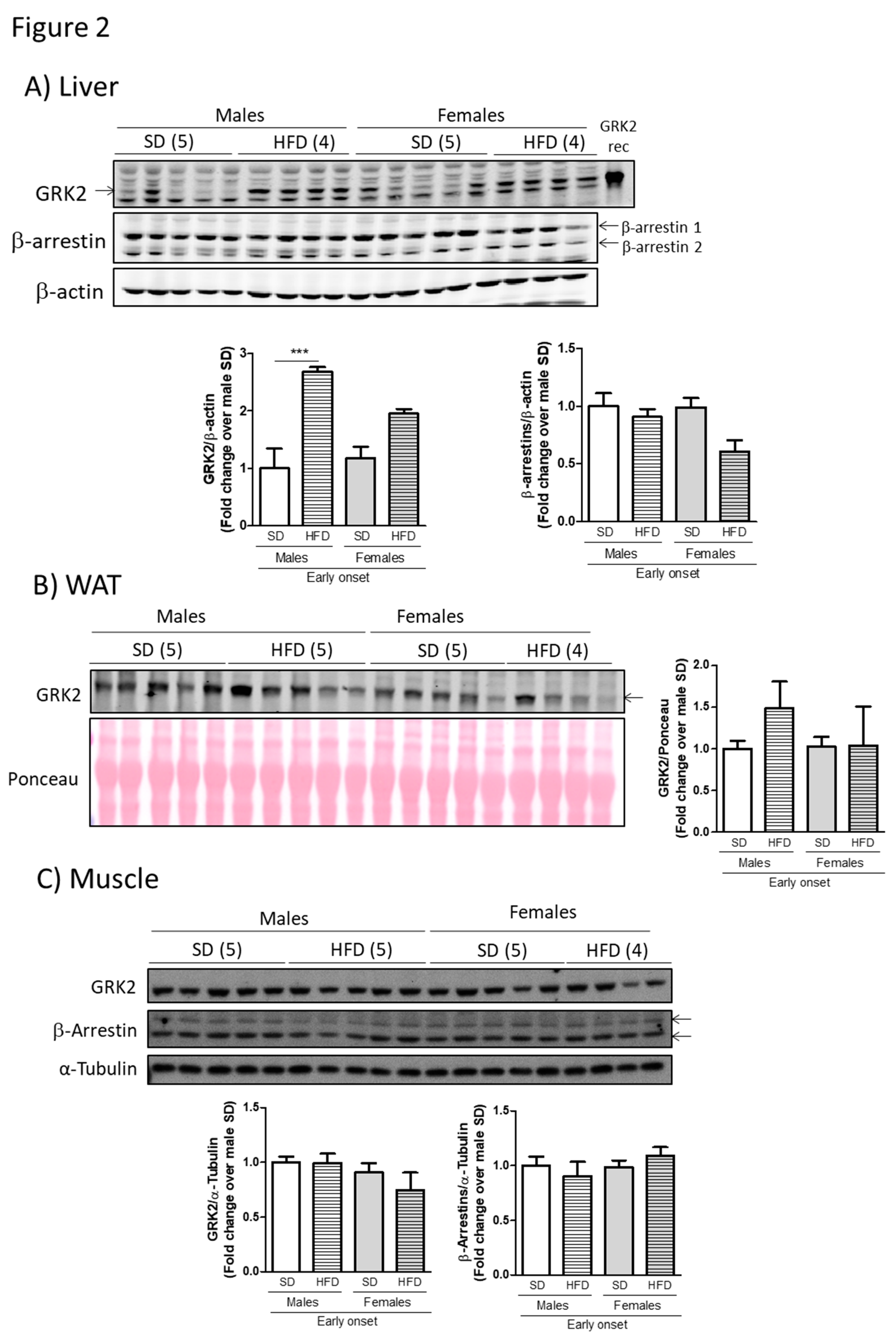
3.2. The Milder Phenotype Observed in Young Female Mice Fed a Hfd Associates with Lower Levels of GRK2 in Metabolically Relevant Tissues Compared to Males
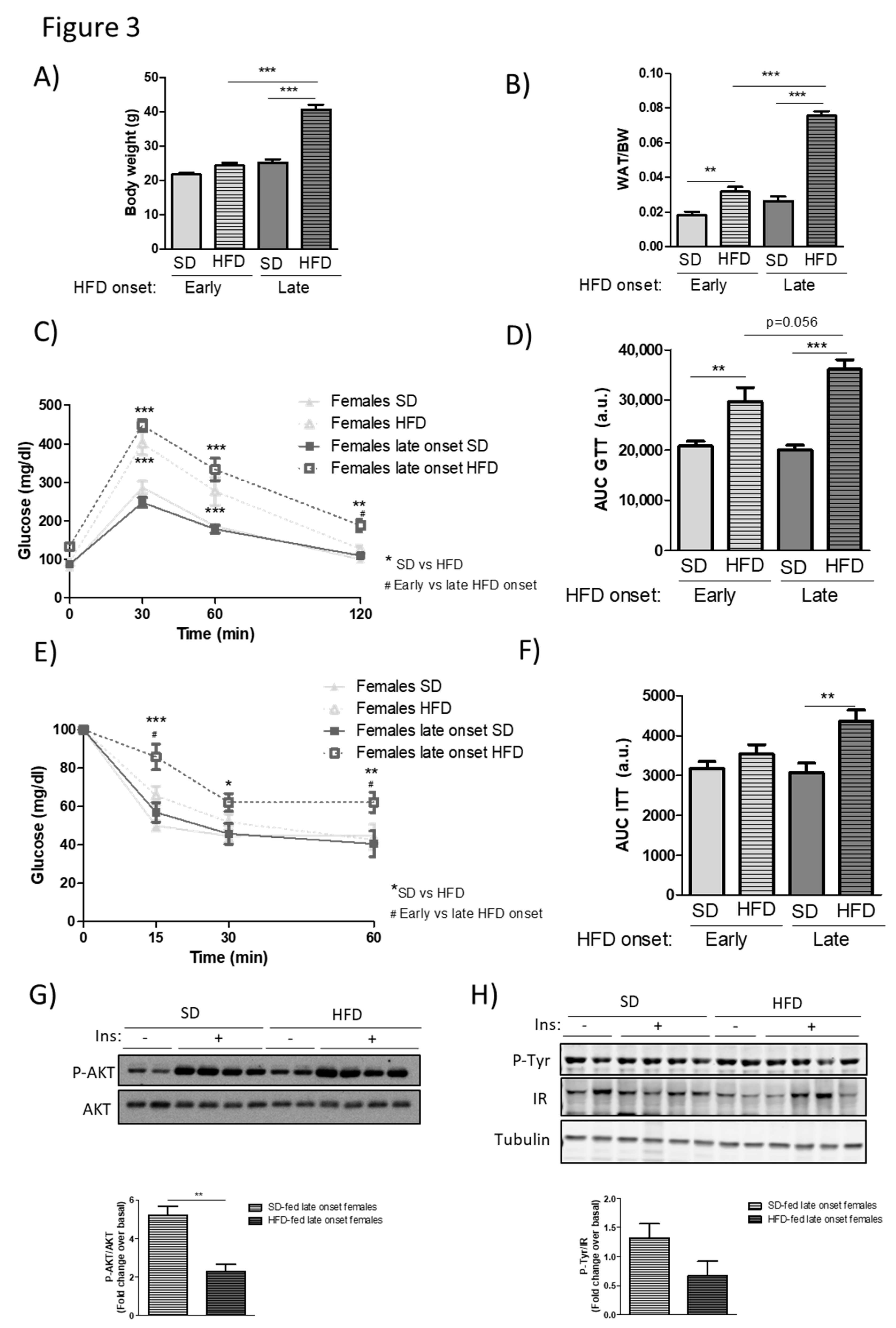
3.3. Middle-Aged Female Mice are No Longer Protected from the Metabolic Alterations Caused by HFD Feeding
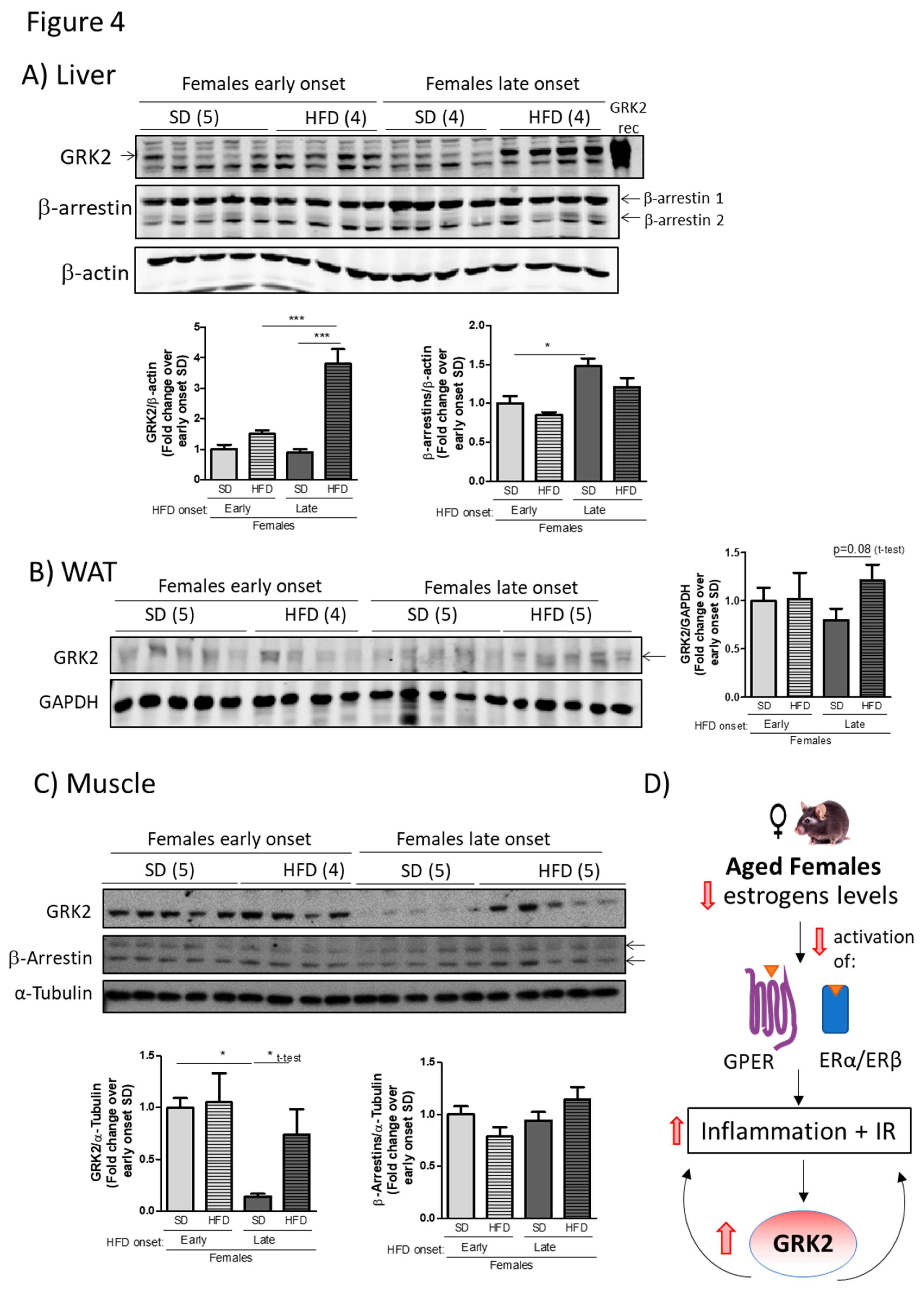
3.4. Age-Associated Loss of Protection Against a HFD in Female Mice Correlates with an Altered Pattern of GRK2 Protein Levels in Metabolically Relevant Tissues in Such Dietary Conditions
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huebschmann, A.G.; Huxley, R.R.; Kohrt, W.M.; Zeitler, P.; Regensteiner, J.G.; Reusch, J.E.B. Sex differences in the burden of type 2 diabetes and cardiovascular risk across the life course. Diabetologia 2019, 62, 1761–1772. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Arnold, A.P.; Reue, K. A Guide for the Design of Pre-clinical Studies on Sex Differences in Metabolism. Cell Metab. 2017, 25, 1216–1230. [Google Scholar] [CrossRef] [PubMed]
- Hevener, A.L.; Zhou, Z.; Moore, T.M.; Drew, B.G.; Ribas, V. The impact of ERalpha action on muscle metabolism and insulin sensitivity-Strong enough for a man, made for a woman. Mol. Metab. 2018, 15, 20–34. [Google Scholar] [CrossRef] [PubMed]
- Salinero, A.E.; Anderson, B.M.; Zuloaga, K.L. Sex differences in the metabolic effects of diet-induced obesity vary by age of onset. Int. J. Obes. (Lond.) 2018, 42, 1088–1091. [Google Scholar] [CrossRef]
- Clegg, D.; Hevener, A.L.; Moreau, K.L.; Morselli, E.; Criollo, A.; Van Pelt, R.E.; Vieira-Potter, V.J. Sex hormones and cardiometabolic health: Role of estrogen and estrogen receptors. Endocrinology 2017, 158, 1095–1105. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F.; Manson, J.E.; Stevenson, J.C.; Fonseca, V.A. Menopausal hormone therapy and type 2 diabetes prevention: Evidence, mechanisms, and clinical implications. Endocr. Rev. 2017, 38, 173–188. [Google Scholar] [CrossRef]
- Sharma, G.; Prossnitz, E.R. G-protein-coupled estrogen receptor (GPER) and sex-specific metabolic homeostasis. Adv. Exp. Med. Biol. 2017, 1043, 427–453. [Google Scholar]
- Penela, P.; Murga, C.; Ribas, C.; Lafarga, V.; Mayor, F., Jr. The complex G protein-coupled receptor kinase 2 (GRK2) interactome unveils new physiopathological targets. Br. J. Pharm. 2010, 160, 821–832. [Google Scholar] [CrossRef]
- Penela, P.; Ribas, C.; Sanchez-Madrid, F.; Mayor, F., Jr. G protein-coupled receptor kinase 2 (GRK2) as a multifunctional signaling hub. Cell Mol. Life Sci. 2019, 1–24. [Google Scholar] [CrossRef]
- Crudden, C.; Shibano, T.; Song, D.; Suleymanova, N.; Girnita, A.; Girnita, L. Blurring boundaries: Receptor tyrosine kinases as functional G protein-coupled receptors. Int. Rev. Cell Mol. Biol. 2018, 339, 1–40. [Google Scholar]
- Mayor, F., Jr.; Cruces-Sande, M.; Arcones, A.C.; Vila-Bedmar, R.; Briones, A.M.; Salaices, M.; Murga, C. G protein-coupled receptor kinase 2 (GRK2) as an integrative signalling node in the regulation of cardiovascular function and metabolic homeostasis. Cell Signal. 2018, 41, 7. [Google Scholar] [CrossRef] [PubMed]
- Sato, P.Y.; Chuprun, J.K.; Schwartz, M.; Koch, W.J. The evolving impact of g protein-coupled receptor kinases in cardiac health and disease. Physiol. Rev. 2015, 95, 377–404. [Google Scholar] [CrossRef] [PubMed]
- Murga, C.; Arcones, A.C.; Cruces-Sande, M.; Briones, A.M.; Salaices, M.; Mayor, F., Jr. g protein-coupled receptor kinase 2 (GRK2) as a potential therapeutic target in cardiovascular and metabolic diseases. Front. Pharm. 2019, 10, 112. [Google Scholar] [CrossRef] [PubMed]
- Anis, Y.; Leshem, O.; Reuveni, H.; Wexler, I.; Ben Sasson, R.; Yahalom, B.; Laster, M.; Raz, I.; Ben Sasson, S.; Shafrir, E.; et al. Antidiabetic effect of novel modulating peptides of G-protein-coupled kinase in experimental models of diabetes. Diabetologia 2004, 47, 1232–1244. [Google Scholar] [CrossRef]
- Vila-Bedmar, R.; Cruces-Sande, M.; Lucas, E.; Willemen, H.L.; Heijnen, C.J.; Kavelaars, A.; Mayor, F., Jr.; Murga, C. Reversal of diet-induced obesity and insulin resistance by inducible genetic ablation of GRK2. Sci. Signal. 2015, 8, ra73. [Google Scholar] [CrossRef]
- Vila-Bedmar, R.; Garcia-Guerra, L.; Nieto-Vazquez, I.; Mayor, F., Jr.; Lorenzo, M.; Murga, C. Fernandez-Veledo, S. GRK2 contribution to the regulation of energy expenditure and brown fat function. Faseb. J. 2012, 26, 3503–3514. [Google Scholar] [CrossRef]
- Usui, I.; Imamura, T.; Babendure, J.L.; Satoh, H.; Lu, J.C.; Hupfeld, C.J.; Olefsky, J.M. G protein-coupled receptor kinase 2 mediates endothelin-1-induced insulin resistance via the inhibition of both Galphaq/11 and insulin receptor substrate-1 pathways in 3T3-L1 adipocytes. Mol. Endocrinol. 2005, 19, 2760–2768. [Google Scholar] [CrossRef]
- Ciccarelli, M.; Chuprun, J.K.; Rengo, G.; Gao, E.; Wei, Z.; Peroutka, R.J.; Gold, J.I.; Gumpert, A.; Chen, M.; Otis, N.J.; et al. G protein-coupled receptor kinase 2 activity impairs cardiac glucose uptake and promotes insulin resistance after myocardial ischemia. Circulation 2011, 123, 1953–1962. [Google Scholar] [CrossRef]
- Lucas, E.; Jurado-Pueyo, M.; Fortuno, M.A.; Fernandez-Veledo, S.; Vila-Bedmar, R.; Jimenez-Borreguero, L.J.; Lazcano, J.J.; Gao, E.; Gomez-Ambrosi, J.; Fruhbeck, G.; et al. Downregulation of G protein-coupled receptor kinase 2 levels enhances cardiac insulin sensitivity and switches on cardioprotective gene expression patterns. Biochim. Biophys. Acta 1842, 2448–2456. [Google Scholar] [CrossRef]
- Penela, P.; Barradas, M.; Alvarez-Dolado, M.; Munoz, A.; Mayor, F., Jr. Effect of hypothyroidism on G protein-coupled receptor kinase 2 expression levels in rat liver, lung, and heart. Endocrinology 2001, 142, 987–991. [Google Scholar] [CrossRef]
- Pettersson, U.S.; Walden, T.B.; Carlsson, P.O.; Jansson, L.; Phillipson, M. Female mice are protected against high-fat diet induced metabolic syndrome and increase the regulatory T cell population in adipose tissue. PLoS ONE 2012, 7, e46057. [Google Scholar] [CrossRef] [PubMed]
- Medrikova, D.; Jilkova, Z.M.; Bardova, K.; Janovska, P.; Rossmeisl, M.; Kopecky, J. Sex differences during the course of diet-induced obesity in mice: Adipose tissue expandability and glycemic control. Int, J. Obes. (Lond.) 2012, 36, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Cruces-Sande, M.; Vila-Bedmar, R.; Arcones, A.C.; Gonzalez-Rodriguez, A.; Rada, P.; Gutierrez-de-Juan, V.; Vargas-Castrillon, J.; Iruzubieta, P.; Sanchez-Gonzalez, C.; Formentini, L.; et al. Involvement of G protein-coupled receptor kinase 2 (GRK2) in the development of non-alcoholic steatosis and steatohepatitis in mice and humans. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 3655–3667. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Rossi, M.; Cui, Y.; Lee, R.J.; Sakamoto, W.; Perry, N.A.; Urs, N.M.; Caron, M.G.; Gurevich, V.V.; Godlewski, G.; et al. Hepatic beta-arrestin 2 is essential for maintaining euglycemia. J. Clin. Investig. 2017, 127, 2941–2945. [Google Scholar] [CrossRef]
- Mauvais-Jarvis, F. Gender differences in glucose homeostasis and diabetes. Physiol. Behav. 2018, 187, 20–23. [Google Scholar] [CrossRef]
- Frias, J.P.; Macaraeg, G.B.; Ofrecio, J.; Yu, J.G.; Olefsky, J.M.; Kruszynska, Y.T. Decreased susceptibility to fatty acid-induced peripheral tissue insulin resistance in women. Diabetes 2001, 50, 1344–1350. [Google Scholar] [CrossRef]
- Hevener, A.; Reichart, D.; Janez, A.; Olefsky, J. Female rats do not exhibit free fatty acid-induced insulin resistance. Diabetes 2002, 51, 1907–1912. [Google Scholar] [CrossRef]
- Nishikawa, S.; Yasoshima, A.; Doi, K.; Nakayama, H.; Uetsuka, K. Involvement of sex, strain and age factors in high fat diet-induced obesity in C57BL/6J and BALB/cA mice. Exp. Anim. 2007, 56, 263–272. [Google Scholar] [CrossRef]
- Bellino, F.L. Nonprimate animal models of menopause: Workshop report. Menopause 2000, 7, 14–24. [Google Scholar] [CrossRef]
- Diaz Brinton, R. Minireview: Translational animal models of human menopause: Challenges and emerging opportunities. Endocrinology 2012, 153, 3571–3578. [Google Scholar] [CrossRef]
- Lucas, E.; Cruces-Sande, M.; Briones, A.M.; Salaices, M.; Mayor, F., Jr.; Murga, C.; Vila-Bedmar, R. Molecular physiopathology of obesity-related diseases: Multi-organ integration by GRK2. Arch. Physiol. Biochem. 2015, 121, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Charbonneau, A.; Unson, C.G.; Lavoie, J.M. High-fat diet-induced hepatic steatosis reduces glucagon receptor content in rat hepatocytes: Potential interaction with acute exercise. J. Physiol. 2007, 579, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Usui, I.; Imamura, T.; Satoh, H.; Huang, J.; Babendure, J.L.; Hupfeld, C.J.; Olefsky, J.M. GRK2 is an endogenous protein inhibitor of the insulin signaling pathway for glucose transport stimulation. Embo. J. 2004, 23, 2821–2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, H.; Worrall, C.; Shen, H.; Issad, T.; Seregard, S.; Girnita, A.; Girnita, L. Selective recruitment of G protein-coupled receptor kinases (GRKs) controls signaling of the insulin-like growth factor 1 receptor. Proc. Natl. Acad. Sci. USA 2012, 109, 7055–7060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crudden, C.; Song, D.; Cismas, S.; Trocme, E.; Pasca, S.; Calin, G.A.; Girnita, A.; Girnita, L. Below the surface: IGF-1R therapeutic targeting and its endocytic journey. Cells 2019, 8, 1223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauvais-Jarvis, F. Menopause, estrogens, and glucose homeostasis in women. Adv. Exp. Med. Biol. 2017, 1043, 217–225. [Google Scholar] [PubMed]
- Mauvais-Jarvis, F. Estrogen and androgen receptors: Regulators of fuel homeostasis and emerging targets for diabetes and obesity. Trends Endocrinol. Metab. 2011, 22, 24–33. [Google Scholar] [CrossRef] [Green Version]
- Ribas, V.; Drew, B.G.; Le, J.A.; Soleymani, T.; Daraei, P.; Sitz, D.; Mohammad, L.; Henstridge, D.C.; Febbraio, M.A.; Hewitt, S.C.; et al. Myeloid-specific estrogen receptor alpha deficiency impairs metabolic homeostasis and accelerates atherosclerotic lesion development. Proc. Natl. Acad. Sci. USA 2011, 108, 16457–16462. [Google Scholar] [CrossRef] [Green Version]
- Morford, J.J.; Wu, S.; Mauvais-Jarvis, F. The impact of androgen actions in neurons on metabolic health and disease. Mol. Cell Endocrinol. 2018, 465, 92–102. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arcones, A.C.; Cruces-Sande, M.; Ramos, P.; Mayor, F.; Murga, C. Sex Differences in High Fat Diet-Induced Metabolic Alterations Correlate with Changes in the Modulation of GRK2 Levels. Cells 2019, 8, 1464. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8111464
Arcones AC, Cruces-Sande M, Ramos P, Mayor F, Murga C. Sex Differences in High Fat Diet-Induced Metabolic Alterations Correlate with Changes in the Modulation of GRK2 Levels. Cells. 2019; 8(11):1464. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8111464
Chicago/Turabian StyleArcones, Alba C., Marta Cruces-Sande, Paula Ramos, Federico Mayor, and Cristina Murga. 2019. "Sex Differences in High Fat Diet-Induced Metabolic Alterations Correlate with Changes in the Modulation of GRK2 Levels" Cells 8, no. 11: 1464. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8111464