Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma
by
 ,
,
Ko-Hsiu Lu
1,2 ,
,
Renn-Chia Lin
1,2,3,
Jia-Sin Yang
4,5,
Wei-En Yang
4,5,
Russel J. Reiter
6,* and 
Shun-Fa Yang
4,5,*
1
Department of Orthopedics, Chung Shan Medical University Hospital, Taichung 402, Taiwan
2
School of Medicine, Chung Shan Medical University, Taichung 402, Taiwan
3
Division of Hyperbaric Oxygen Therapy and Wound Medicine, Chung Shan Medical University Hospital, Taichung 402, Taiwan
4
Institute of Medicine, Chung Shan Medical University, Taichung 402, Taiwan
5
Department of Medical Research, Chung Shan Medical University Hospital, Taichung 402, Taiwan
6
Department of Cell Systems and Anatomy, UT Health, San Antonio, TX 78229, USA
*
Authors to whom correspondence should be addressed.
Cells 2019, 8(12), 1618; https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121618
Submission received: 26 November 2019
/
Revised: 6 December 2019
/
Accepted: 9 December 2019
/
Published: 12 December 2019
(This article belongs to the Special Issue Melatonin in Human Health and Diseases)
Abstract
:Osteosarcoma, the most common primary bone malignancy, occurs most frequently in adolescents with a peak of incidence at 11–15 years. Melatonin, an indole amine hormone, shows a wide range of anticancer activities. The decrease in melatonin levels simultaneously concurs with the increase in bone growth and the peak age distribution of osteosarcoma during puberty, so melatonin has been utilized as an adjunct to chemotherapy to improve the quality of life and clinical outcomes. While a large amount of research has been conducted to understand the complex pleiotropic functions and the molecular and cellular actions elicited by melatonin in various types of cancers, a few review reports have focused on osteosarcoma. Herein, we summarized the anti-osteosarcoma effects of melatonin and its underlying molecular mechanisms to illustrate the known significance of melatonin in osteosarcoma and to address cellular signaling pathways of melatonin in vitro and in animal models. Even in the same kind of osteosarcoma, melatonin has been sparingly investigated to counteract tumor growth, apoptosis, and metastasis through different mechanisms, depending on different cell lines. We highlighted the underlying mechanism of anti-osteosarcoma properties evoked by melatonin, including antioxidant activity, anti-proliferation, induction of apoptosis, and the inhibition of invasion and metastasis. Moreover, we discussed the drug synergy effects of the role of melatonin involved and the method to fortify the anti-cancer effects on osteosarcoma. As a potential therapeutic agent, melatonin is safe for children and adolescents and is a promising candidate for an adjuvant by reinforcing the therapeutic effects and abolishing the unwanted consequences of chemotherapies.
1. Introduction
Cancer is a well-known public health problem associated with high mortality and disability rates worldwide [1]. According to the data reported by the World Health Organization (WHO), new cancer-related cases have increased to 18.1 million and cancer-induced deaths to 9.6 million in 2018 [2]. Cancer is a leading cause of death for children, with 300,000 new cases diagnosed each year. Osteosarcoma is the most prevalent primary bone cancer with a peak of incidence at 10–15 years and the second incidence peak in older adulthood [3,4,5]. This malignant bone tumor often first develops in the metaphysis of long bones (distal femur and proximal tibia) in about six in every million children and two in every million adults [6]. It is therefore feasible that there may be a relationship between the incidence of osteosarcoma and the rate of bone growth [4].
Even though surgical en bloc resection of the cancer or amputation of the extensive diseased extremity to achieve a complete radical excision has been the treatment of choice for osteosarcoma, the main cause of most treatment failures and the high mortality rate is still its highly metastatic potential [7]. Reassuringly, the combination of surgery and chemotherapy for osteosarcoma has increased the long-term survival chances of approximately 68% through limb-sparing surgeries based on radiological staging, surgical techniques, and new chemotherapy protocols [8]. Nonetheless, potent metastatic transfers to the lungs are still responsible for most treatment failures and are accountable for one of the most lethal pediatric malignancies. Thus, novel agents that target particular intracellular pathways related to the distinctive properties of osteosarcoma cells need to be developed.
Melatonin levels, which are high in children, with the highest peak between ages four–seven [9], begin to decrease prior to the onset of puberty and continue to decline during puberty [10]. Initially, it is rather bewildering how such findings, compared with the incidence of osteosarcoma, have generally received little attention. After establishing a hypothesis of a correlation between the declination in melatonin levels (with provides oncostatic protection) and the increase in the chances of developing the fatal osteosarcoma in young patients, melatonin has been studied for its anti-osteosarcoma action and as an adjunct to the conventional chemotherapy for osteosarcoma to improve the prognosis of the fatal disease [11]. Moreover, melatonin is non-toxic and can augment anticancer action while ameliorating the side effects of many other chemotherapeutic drugs. Therefore, it is not surprising that increasing attention has been devoted to melatonin as it may be integrated into adjuvant therapies to amplify the therapeutic effects and to diminish the side effects of chemotherapies [12].
2. Biosynthesis, Metabolism, Actions, and Signaling Pathways of Melatonin
2.1. Biosynthesis of Melatonin
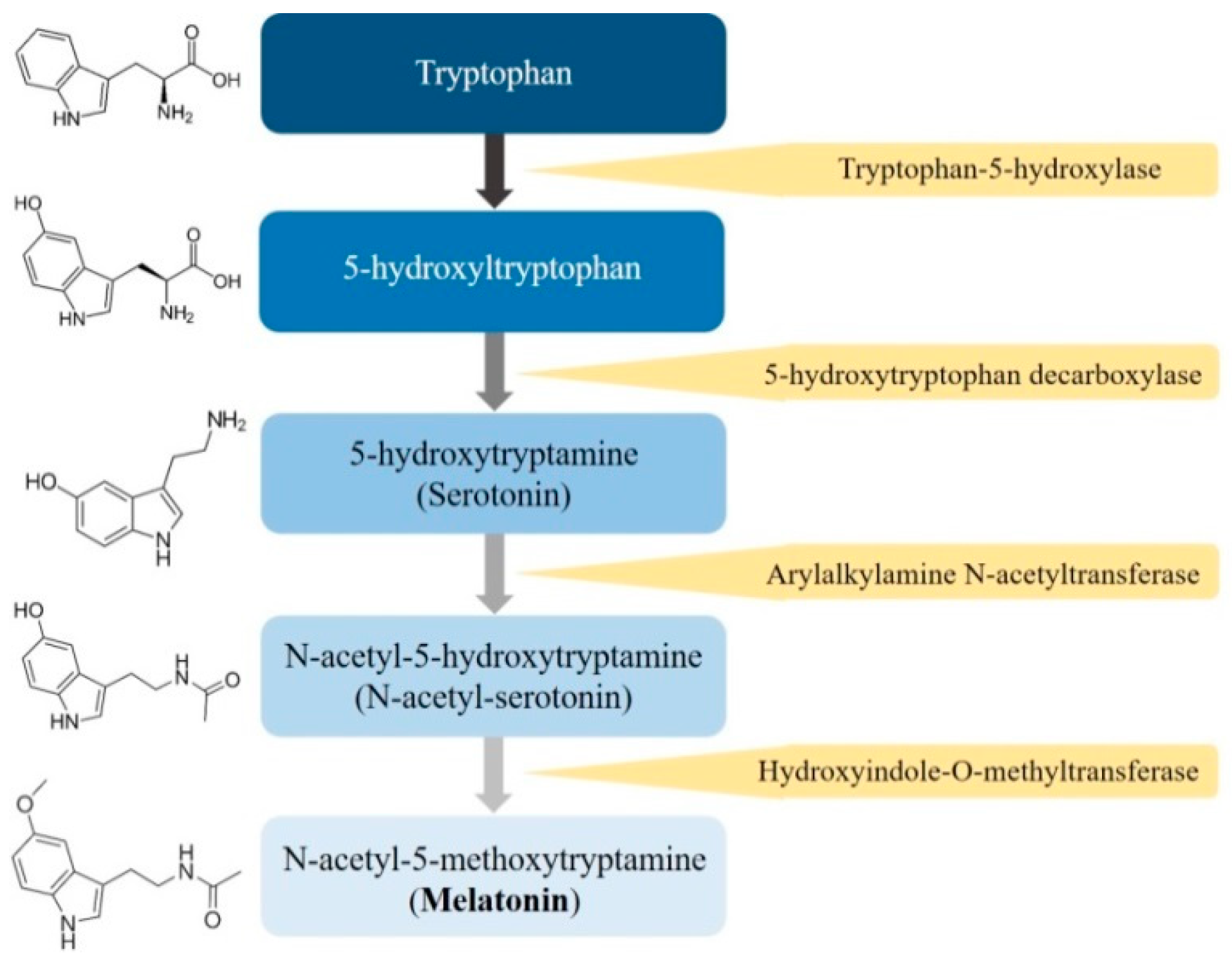
Melatonin (N-acetyl-5-methoxytryptamine), a ubiquitously amphiphilic indole amine, is synthesized from its precursor, tryptophan, and is secreted primarily by the pineal gland and several other organs, including the retina, gastrointestinal tract, bile, skin, bone marrow, and lymphocytes, of humans and mammals [13,14]. In mammals, melatonin synthesis is dependent on the day-night cycle and is part of an intricate enzymatic pathway in which tryptophan is hydroxylated to 5-hydroxytryptophan by tryptophan-5-hydroxylase [15] (Figure 1). Then, 5-hydroxytryptophan is decarboxylated into serotonin by 5-hydroxy-l-tryptophan decarboxylase; afterwards, serotonin is acetylated into N-acetylserotonin, and N-acetylserotonin is subsequently methylated to melatonin, which is quickly released into the bloodstream.
2.2. Metabolism of Melatonin
As a circadian pattern, the synthesis and secretion of melatonin is regulated by the central circadian clock, which is located in the suprachiasmatic nucleus of the hypothalamus [16], and its synthesis is stimulated by the night and inhibited by light [17]. As in instances of reduced light exposure, norepinephrine is released from sympathetic nerve endings to strengthen the intracellular levels of c-AMP and activates the protein kinase A, which in term regulates the function of arylalkylamine N-acetyltransferase to limit the rates of melatonin production [15]. The levels of melatonin in cerebrospinal fluid and blood reach maximal concentration near the middle of the darkest period (between 02:00 and 04:00) and decrease throughout the daytime [18]. Nearly 80% of the melatonin is synthesized at night with nocturnal serum concentrations varying between 80 and 120 pg/mL and low serum concentrations (10–20 pg/mL) during daylight hours [9]. After an oral intake of 500 μg melatonin, the mean (± SD) half-life of the elimination is 47 ± 3 min [19]. The clearance of melatonin is rapid through the liver by cytochrome P450 enzymes CYP1A1 and 1A2 enzymes undergoing 6-hydroxylation to 6-hydroxymelatonin [20]. Large amounts of the metabolites are secreted into the bile [21] with minor amounts being further conjugated with sulfuric acid (90%) or glucuronic acid (10%) and being excreted in the urine as 6-sulfatoxymelatonin [22]. Additionally, about 5% of serum melatonin is excreted as the unmetabolized drug [23]. In case of intravenous administration, the hepatic bio-degradation is less important due to the lack of hepatic first pass.
2.3. Actions of Melatonin
While melatonin is regulated by the central circadian clock, it transmits during the nighttime and helps organize target organs and organ systems into appropriate homeostatic metabolic rhythms [24] and also reverses to modulate the central circadian clock and peripheral oscillators in tissues and organs, which makes melatonin a circadian pacemaker [25]. Meanwhile, circadian secretion of melatonin also synchronizes the immune system via a reciprocal association. As the relevance of melatonin upholds the circadian rhythm and orchestrates many signaling pathways, melatonin is not only a hormone, but also a cell protector [26] involved in immunomodulation, anti-oxidative processes, and hematopoiesis [20]. Consequently, a disruption of the circadian rhythm can interfere with the nocturnal melatonin signal to disturb normal homeostatic metabolic rhythms and many physiologic cell functions, which, if deregulated, may cause an accelerated pace towards cancer [27]. In addition to sleep induction, biological rhythms resynchronization, anti-oxidation, and anti-inflammation, melatonin possesses a wide spectrum of biological effects such as immunomodulation and apoptosis induction [28]. Furthermore, melatonin is involved in the regulation of immune functions and the tumor microenvironment and acts as an anticancer agent. Hence, it is not surprising that melatonin has been popularly used as a healthcare product in the global market.
2.4. Signaling Pathways of Melatonin
The amphiphilic indole amine is capable of easily penetrating into cells and exerting various biological effects that either attributes to its interactions with the cell surface and intracellular receptors or directs effects as a radical scavenger [29]. By affecting its receptors or acting as a direct antioxidant agent, melatonin modulates many signaling pathways essential for cellular functions [26], such as cellular metabolism, DNA damage response, cell-to-cell communication, and more. As a result, melatonin behaves via receptor-dependent and receptor-independent mechanisms for its important oncostatic properties [30]. The receptors affected by melatonin belong to the G protein coupled receptors superfamily, including three subtypes of MT1, MT2, and MT3. In mammals, MT1 (encoded by MTNR1A) and MT2 (encoded by MTNR1B) are mainly responsible for regulating the melatonin’s downstream effects [31].
The MT1 receptor, mainly located in the pars-tuberalis of the pituitary gland, the suprachiasmatic nuclei of the hypothalamus [16], the skin, and the retina, is believed to be involved in the inhibitory effect of melatonin in mammalian brains to act as a master regulator of brain function. The MT2 receptor, which is widely distributed in osteoblasts, vessels of extremities, and the retina, is active in the phase shifting of the circadian clock [32]. However, the receptor-independent mechanisms are related to the prevention of circadian disruption, antioxidant activity, tumor metabolism and cancer immunity, regulation of apoptosis, and inhibition on angiogenesis and migration [30,33]. Melatonin’s anticancer properties in many cancers have recently attracted attention; thereby, the role of melatonin in the treatment of cancer cells has been extensively studied and the number of articles has been increasing rapidly, given that amplification of the anticancer properties constitutes a possible treatment feature in osteosarcoma. Recently, a compelling body of evidence has documented significant anti-osteosarcoma effects of melatonin in vitro and in vivo.
3. Function of Melatonin Relating to the Bone
3.1. Effects of Melatonin on Mesenchymal Stem Cells
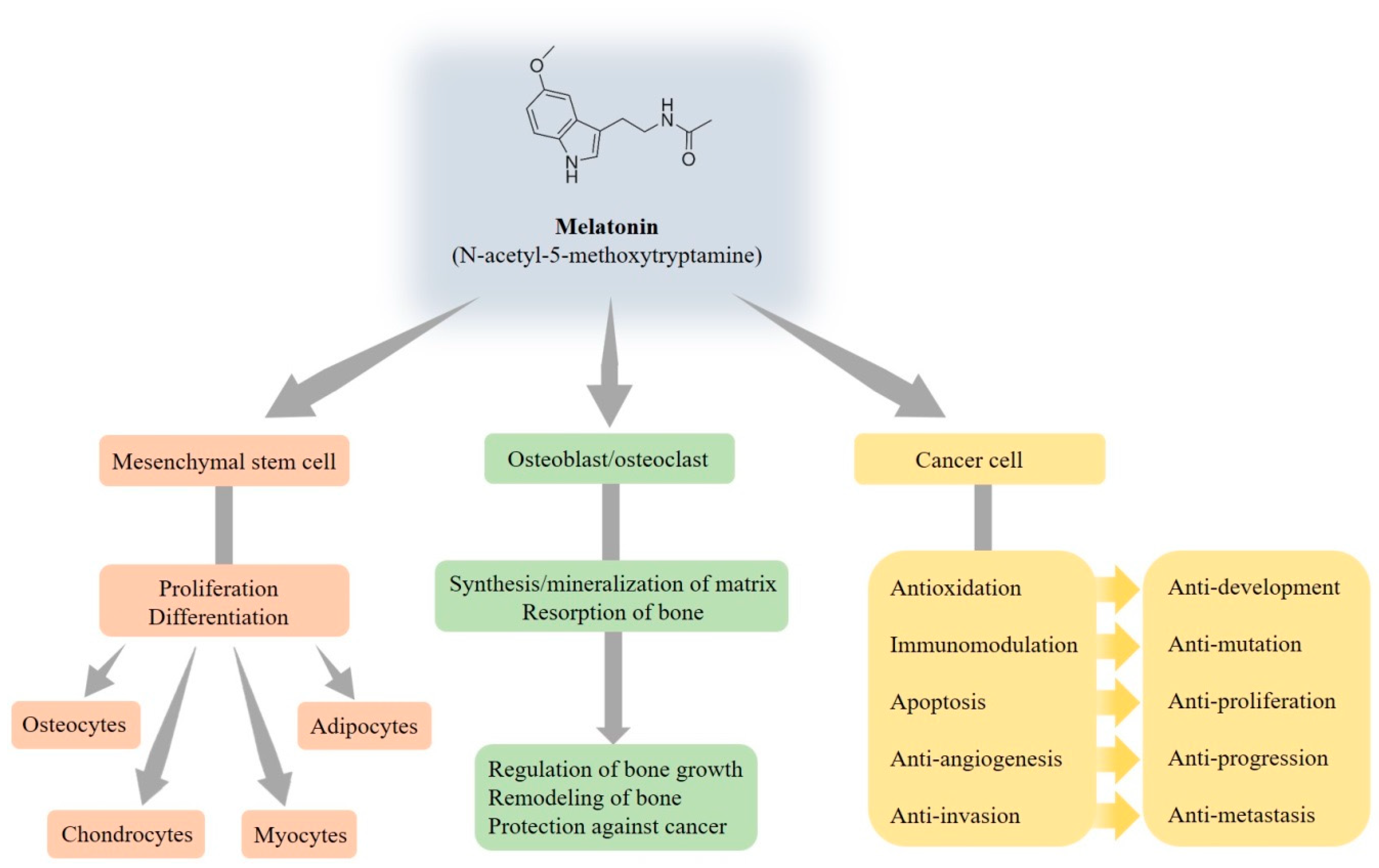
It is widely demonstrated that melatonin is able to regulate signaling pathways that drive commitment and the differentiation of mesenchymal stem cells, which are multipotent progenitor cells, into several cell types such as osteocytes, chondrocytes, myocytes, and adipocytes [34] (Figure 2). Mesenchymal stem cells’ differentiation is finely regulated by the action of mechanical and molecular signals from the extracellular environment, but melatonin may also be an important regulator of precursor cell commitment and differentiation [35]. It is universally approved that adipogenesis and osteogenesis is a reciprocal relationship in bone marrow. Despite no apparent effect on the proliferation of human mesenchymal stem cells, melatonin directly inhibits adipogenic differentiation toward the adipocyte lineage and simultaneously fosters osteogenic differentiation by suppressing peroxisome proliferator-activated receptor γ (PPARγ) expression and enhancing Runt-related transcription factor 2 (Runx2) expression [36]. Furthermore, melatonin enhances the differentiation of human mesenchymal stem cells to osteoblasts via MT2 receptors and the mitogen/extracellular signal-regulated kinase (MEK)/extracellular signal-regulated kinase (ERK) 1/2 signaling cascade [37].
3.2. Effects of Melatonin on Osteoblasts and Osteoclasts
In addition to the regulation of the sleep cycle, endocrine system, reproductive cycle, bone metabolism, cell cycle, and mitochondrial function, melatonin influences bone growth and metabolism to promote the differentiation and mineralization of osteoblast cells grown [38]. Additionally, melatonin participates in the regulation of bone homeostasis via the modulation of osteoblast and osteoclast activities by receptor-independent and dependent pathways, and then bone remodeling is established by the synthesis of bone matrix by osteoblasts and the resorption of bone by osteoclasts [39]. Meanwhile, melatonin induces osteoblastic differentiation and mineralization for osteogenesis and bone formation through the bone morphogenetic protein (BMP)/ERK/Wnt signaling pathways [40].
Disrupted circadian rhythms alter the expression of clock genes and deregulate oncogenes, which ultimately promote tumor development and progression. Melatonin indeed represses in vitro osteoblast differentiation and the mineralization of the matrix to regulate bone growth [41,42], so the fall of melatonin has been shown to be an indicator of skeletal maturation [43]. Recent studies have reported that melatonin possesses multiple tumor-suppressing properties for a myriad of tumors [30,33,44,45]. It is worthy to mention that individuals with distinct defects in osteoblast functions are at an increased risk of contracting cancer, indicating that osteosarcoma may be partly related to abnormal osteoblast functions. However, melatonin, on the contrary, possesses the ability to enhance normal osteoblast functions and thus plays a protective role against cancer.
3.3. Effects of Melatonin on Cancer Cells
Melatonin has been sparingly investigated to counteract tumor growth and metastasis through various mechanisms, depending not only on different cancer types, but also the different cell lines in the same kind cancer. Even the actions of melatonin between cancer cells and normal cells are obviously different [30,44,45,46,47,48]. For example, melatonin is antiapoptotic in normal cells [49], but proapoptotic in cancer cells [50]. Due to the widespread subcellular distribution and free radical scavenging actions, numerous studies have suggested that melatonin abrogates oxidative damage to inhibit cancer development [51], and this relates, in part, to its pro-oxidative actions in cancer cells [50]. To enhance the immunomodulatory potential of the destined cell mutation, melatonin augments the immune responses and alleviates immunodeficiency states [52], along with enhancing immunosurveillance by stimulating the activities of immune cells, including T and B lymphocytes, monocytes, macrophages, and natural killer cells, and stimulating the several cytokine productions, for example, interferon-γ, interleukin (IL)-1, IL-2, IL-6, and tumor necrosis factor-α [53]. As a result of accumulating evidence, melatonin possesses pleiotropic bioactivities and exhibits a very wide antitumor repertoire.
As mentioned earlier, the role of melatonin in inhibiting tumor proliferation has been well documented [48]. Melatonin has directed proapoptotic on cancer cells, limits the cellular uptake of key factors for tumor growth and their signaling molecules [54], and inhibits cell cycle kinetics and telomerase activity to restrain tumor cell growth [55]. To suppress cancer progression and metastasis, melatonin reduces vascular endothelial growth factor secretion and the formation and release of endothelin-1 to inhibit angiogenesis [56]. Through the remodeling of the extracellular matrix (ECM), the reorganization of the cytoskeleton, and epithelial–mesenchymal transition (EMT) [47,48], melatonin counteracts the angiogenic responses and thus abrogates cancer cell invasion and metastasis.
4. Molecular Actions of Melatonin in Osteosarcoma
4.1. Cytotoxic Activity of Melatonin in Human Osteosarcoma
Apoptosis, or programmed cell death, a key regulator of physiological growth control and regulation of tissue homeostasis, is characterized by typical morphological and biochemical hallmarks, including cell shrinkage, nuclear DNA fragmentation, and membrane blebbing [57]. To undergo apoptosis, the activation of an important initiator and effector caspases would be initiated through the activation of the extrinsic (receptor) pathway or the stimulation of the intrinsic (mitochondria) pathway [58,59]. Multiple stress-inducible molecules, such as mitogen-activated protein kinase (MAPK)/ERK, c-Jun N-terminal kinase (JNK), and nuclear factor-κB, have been implied in transmitting the apoptotic pathway [60,61]. Currently, most anticancer strategies in clinical oncology focus on triggering apoptosis in cancer cells. On the contrary, failure to undergo apoptosis may result in treatment resistance. Thereby, understanding the molecular events that regulate apoptosis in response to chemotherapy provides novel opportunities to develop the molecular targeted therapy through the intrinsic and/or extrinsic pathways for the intractable osteosarcoma.
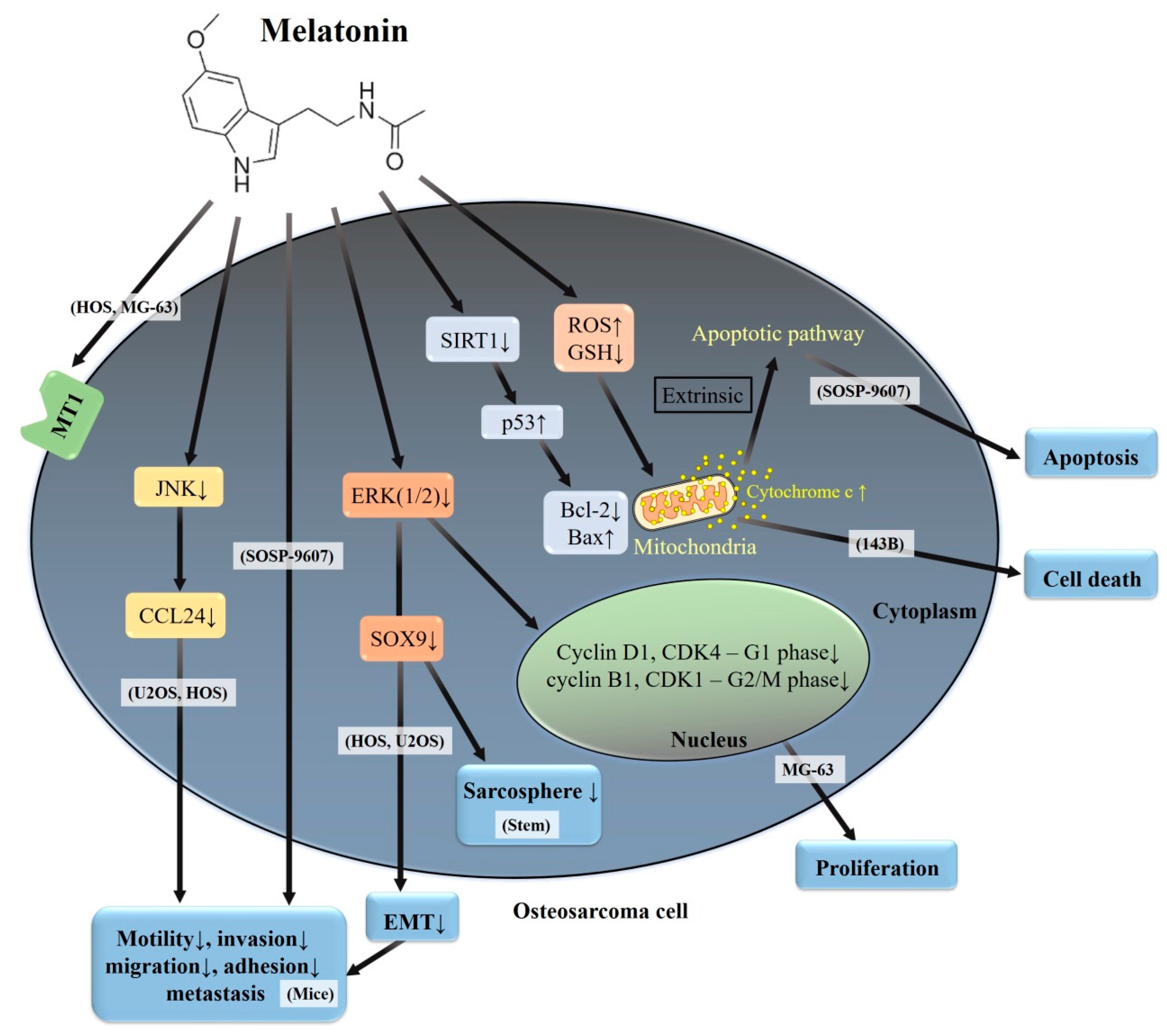
Compelling evidence has highlighted that melatonin demonstrates both cytotoxic and anti-metastatic activities in various cancer cells and they appear to be cell-type specific—even in human osteosarcoma cell lines (Table 1). Abundant expression of MT1-mRNA in human osteosarcoma HOS and MG-63 cells and other malignant and non-malignant bone tumors has been demonstrated [62], and these findings suggest an irreplaceable role for MT1 in bone pathology (Figure 3). In human osteosarcoma 143B cells, 100 μM of melatonin reduces mitochondrial reactive oxygen species (ROS) generation, cell death, and mitochondrial ROS-induced depletion of cardiolipin in order to improve the retardation of mitochondrial movement and dynamics [63]; however, lower concentrations (10−5–10−13 M) of melatonin have no effect on the growth, morphology or cell cycle [64]. It seems that the adequate concentration of melatonin exhibits a considerable potential for rescuing cardiolipin-dependent mitochondrial dynamics-associated mitochondrial pathologies in 143B cells.
Through the downregulation of the expressions of cyclin D1 and CDK4 (related to G1 phase), as well as cyclin B1 and CDK1 (related to G2/M phase), melatonin inhibits the proliferation of human osteosarcoma MG-63 cells [65]; meanwhile, the inhibition of the ERK1/2 signaling pathway is involved in the melatonin’s anti-proliferative effect, induction of G1 and G2/M phase cell arrest, and downregulation of expressions of cyclin D1, CDK4, cyclin B1, and CDK1 in MG-63 cells [66]. Furthermore, melatonin affects the cellular redox status to create a substantial correlation between apoptosis and ROS generation in human osteosarcoma SOSP-9607 cells [67]. In addition to the alleviation of the adhesive and migratory abilities, melatonin induces apoptosis in SOSP-9607 cells via the down-regulation of sirtuin 1 (SIRT1, a conserved nicotinamide adenine dinucleotide-dependent deacetylase), which has an association with age and cancer [68], thus leading to increased p53 acetylation. Then, acetylated p53 triggers the intrinsic apoptotic pathway by decreasing Bcl-2 levels and increasing Bax and cytochrome c expressions.
4.2. Anti-Metastatic Effects of Melatonin in Human Osteosarcoma
As the tumor develops, it becomes increasingly important for the cancer cells to sustain their growth, to increase the tumor size through angiogenesis process, and to metastasize throughout the body. Rampant metastasis of cancerous cells is a serious feature of all malignancies and most cancer morbidities and mortalities are associated with metastatic spread. Accordingly, it is a significant goal to bring insight in order to understand the signaling pathways underlying the complicated process of metastasis that occurs through a series of steps termed the invasion–metastasis cascade [69]. Briefly, the invasion–metastatic cascade pathways include the detachment of cancer cells and degradation of ECM, induction of cellular motility, invasion and migration, adhesion themselves to endothelial cells, and re-establishment of cancer cell growth at a distant site [70].
It is generally accepted that EMT in the developmental program plays a critical role in promoting metastasis in epithelium-derived carcinoma throughout the metastasis process. In contrast, sarcoma cells of mesenchymal origin embed themselves inside the ECM and rarely establish tight contact with neighboring cells. During the past decade, the mechanism of EMT involvement in invasion and migration allows carcinoma cells to dissociate from each other and to degrade ECM, but the investigators further applied the mechanism to sarcoma cells, e.g., osteosarcoma, with the ability to initiate the invasion–metastasis cascade [47,48,71,72,73]. In this regard, sarcoma cells make ECM easier and it may be one of the reasons why osteosarcoma, arising from the aberrant transformation of mesenchymal cells, exhibits a highly metastatic potential. Regrettably, lung metastasis is the main cause of death rather than the primary tumor itself [74].
Of the sirtuin family members, SIRT1 is the most well-known to participate in a number of processes, including cell growth, variability, p53 acetylation, DNA damage, and apoptosis [75,76]. As mentioned before, higher concentrations (up to 1000 μM) of melatonin induce apoptosis in human osteosarcoma SOSP-9607 cells via the down-regulation of sirtuin 1, leading to p53 hyperacetylation, whereas lower concentrations (12.5–50 μM) of melatonin inhibit SOSP-9607 cellular adhesion and migration [67] (Table 2). Another in vitro study shows that melatonin potently suppresses migration and invasion in human osteosarcoma HOS and U2OS cells, and inhibits the sarcosphere formation of osteosarcoma stem cells via the down-regulation of SOX9-mediated signaling pathway [77]. Additionally, melatonin suppresses EMT via the downregulation of SOX9-mediated signaling to inhibit migration and invasion of HOS and U2 cells and decreases tumor initiating cells and lung metastasis of osteosarcoma in the nude mice model. Likewise, melatonin significantly downregulates the expression of mesenchymal marker vimentin, as well as decreases the expression of β-catenin and N-cadherin, indicating that melatonin can partially block the EMT progress of U2 cells. Furthermore, p-ERK is prominently downregulated and the transcription factor SOX9 expression level is significantly repressed under the melatonin treatment, indicating that melatonin might suppress the EMT program and stem cells in U2 cells through the ERK pathway and downregulation of SOX9 expression. Collectively, in vitro and in vivo evidence provides that SOX9 has a crucial role in mediating melatonin-dependent inhibition of the initiation and metastasis of HOS and U2 cells.
While melatonin, up to 2.0 mM, has no cytotoxic effects, it intriguingly suppresses cellular motility, migration, and invasion in human osteosarcoma U2OS and HOS cells and represses the gene expression of C-C motif chemokine ligand 24 (CCL24) in U2OS cells [45]. Manipulation of CCL24 levels influenced the motility of U2OS and HOS cells as cell migration and invasion can be enhanced by the addition of recombinant human CCL24 and can be attenuated by the silencing of CCL24. While both inhibitors of JNK1/2 (SP600125 and DN-JNK) exaggerate melatonin’s attenuation of the expression of CCL24 mRNA and the migratory potential in U2OS cells, the inhibitor of ERK1/2 (U0126) cannot affect melatonin’s actions. Altogether, U2OS and HOS cell-derived CCL24 contributes to cellular invasion and migration through the upstream JNK signaling pathway; this finding implies a promotional role of CCL24 in osteosarcoma metastasis; the action is inhibited by melatonin.
4.3. Synergistic Effects and Utilization of Micro/Nanoparticles Delivery and Inclusion Complex Systems in Human Osteosarcoma
In managing patients diagnosed with any form of osteosarcoma, powerful therapeutic agents with less to no harmful side effects are the mainstay. Numerous approaches have been undertaken to improve survival rates and to diminish adverse side effects of drugs therapy, including the concomitant use of chemotherapy with melatonin due to its efficacy and safety [78,79]. Even in in vitro studies unresponsive to melatonin alone, the pineal hormone may advance the cytostatic and the cytotoxic effects triggered by other compounds or conventional drugs. Accumulating evidence suggests beneficial effects of the co-administration of melatonin with conventional drugs, which leads to synergistic effects, thus increasing cancer cell inhibition [48,50], indicating that melatonin is preeminently suitable as a drug in combination chemotherapy. For osteosarcoma, it is well perceived that melatonin inhibits activity, blocks the cell cycle at the G1-stage, and induces apoptosis in human osteosarcoma SaOS-2 cells [80] (Table 3). While melatonin has an antagonistic effect with a lower concentration of cis-platinum, it has a synergistic effect with methotrexate or a higher concentration of cis-platinum.
With current chemotherapeutic schedules for osteosarcoma, lung metastasis is almost impossible to cure [81]. Melatonin modulates anti-apoptotic processes in normal cells and activates bone cell proliferation and differentiation at 1 nM–1 μM concentrations [82], whereas it inhibits the proliferation of bone cancer cells to trigger pro-apoptotic signals at 4–10 mM concentrations [65]. The idea of encapsulating the drugs inside a micro/nanoparticle, fabricated from biodegradable polymers for drug delivery systems, to control drug release at desired sites, has become very intriguing [83]. Owing to the relatively short half-life (30–57 min) of melatonin in the blood [84], melatonin loaded poly(d,l-lactide-co-glycolide) (PLGA) nanoparticles and microparticles have been employed to fortify the inhibitory effect of human osteosarcoma MG-63 cell proliferation, providing an expectation about the usage of melatonin as an adjunct to the routine chemotherapy of osteosarcoma [83]. Moreover, 9 mM of melatonin concentration released from the melatonin/2-hydroxypropyl-β-cyclodextrin (HPβCD) inclusion complex loaded chitosan scaffolds causes time dependent cell death by reducing the proportion of the cells in the G2/M phase rather than the S phase, so the inclusion complex can be considered an alternative system for human osteosarcoma therapy [85].
5. Conclusions
Cellular and molecular signaling pathways, including oxidative stress, proliferation, apoptosis, and metastasis, involved in osteosarcoma, have been partially clarified in the past decade. Several in vitro and animal studies have evaluated the effect of melatonin on osteosarcoma, including synergistic effects with conventional chemotherapies and utilization of micro/nanoparticles delivery and inclusion complex systems for fortifying the effect. Accumulating evidence from experimental studies has supported the anti-osteosarcoma properties of melatonin, which may be a promising candidate as an adjuvant agent in osteosarcoma treatment. However, more studies are needed to completely elucidate the synergistic effect of melatonin with other compounds in osteosarcoma, both to raise success rates of therapy and also to abolish side effects. As melatonin is a potential therapeutic agent, we herein address several key concerns regarding its potential for treating patients with osteosarcoma. Finally, we need robust evidence from carefully designed and conducted clinical trials to underpin the treatment of osteosarcoma patients, particularly in young people.
Author Contributions
Conceptualization, K.-H.L., R.-C.L., S.-F.Y. and R.J.R.; writing—original draft preparation, K.-H.L., J.-S.Y., W.-E.Y. and S.-F.Y.; writing—review and editing, K.-H.L., S.-F.Y. and R.J.R. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research received no external funding.
Acknowledgments
The authors would like to express sincere thanks to Eric Wun-Hao Lu of American School in Taichung for his assistance in the preparation of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| BAX | Bcl-2-associated X protein |
| Bcl-2 | B-cell lymphoma/leukemia 2 |
| CCL24 | C-C motif chemokine ligand 24 |
| EMT | epithelial–mesenchymal transition |
| ERK | extracellular signal-regulated protein kinase |
| GSH | glutathione |
| JNK | c-Jun N-terminal kinase |
| MT1 | melatonin receptor 1 |
| ROS | reactive oxygen species |
| SIRT1 | sirtuin 1 |
References
- Li, T.; Jiang, S.; Yang, Y. Database selection and heterogeneity-more details, more credibility. JAMA Oncol. 2018, 4, 1295. [Google Scholar] [CrossRef] [PubMed]
- The World Health Organization (WHO). Cancer. Available online: https://www.who.int/cancer/en/ (accessed on 10 December 2019).
- Mirabello, L.; Troisi, R.J.; Savage, S.A. Osteosarcoma incidence and survival rates from 1973 to 2004: Data from the surveillance, epidemiology, and end results program. Cancer 2009, 115, 1531–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picci, P.; Mercuri, M.; Ferrari, S.; Alberghini, M.; Briccoli, A.; Ferrari, C.; Pignotti, E.; Bacci, G. Survival in high-grade osteosarcoma: Improvement over 21 years at a single institution. Ann. Oncol. 2010, 21, 1366–1373. [Google Scholar] [CrossRef] [PubMed]
- Mertens, W.C.; Bramwell, V. Osteosarcoma and other tumors of bone. Curr. Opin. Oncol. 1994, 6, 384–390. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Briccoli, A.; Mercuri, M.; Bertoni, F.; Picci, P.; Tienghi, A.; Del Prever, A.B.; Fagioli, F.; Comandone, A.; Bacci, G. Postrelapse survival in osteosarcoma of the extremities: Prognostic factors for long-term survival. J. Clin. Oncol. 2003, 21, 710–715. [Google Scholar] [CrossRef]
- Ciernik, I.F.; Niemierko, A.; Harmon, D.C.; Kobayashi, W.; Chen, Y.L.; Yock, T.I.; Ebb, D.H.; Choy, E.; Raskin, K.A.; Liebsch, N.; et al. Proton-based radiotherapy for unresectable or incompletely resected osteosarcoma. Cancer 2011, 117, 4522–4530. [Google Scholar] [CrossRef] [Green Version]
- Oertel, S.; Blattmann, C.; Rieken, S.; Jensen, A.; Combs, S.E.; Huber, P.E.; Bischof, M.; Kulozik, A.; Debus, J.; Schulz-Ertner, D. Radiotherapy in the treatment of primary osteosarcoma--a single center experience. Tumori 2010, 96, 582–588. [Google Scholar] [CrossRef]
- Karasek, M.; Winczyk, K. Melatonin in humans. J. Physiol. Pharm. 2006, 57, 19–39. [Google Scholar]
- Waldhauser, F.; Weiszenbacher, G.; Frisch, H.; Zeitlhuber, U.; Waldhauser, M.; Wurtman, R.J. Fall in nocturnal serum melatonin during prepuberty and pubescence. Lancet 1984, 1, 362–365. [Google Scholar] [CrossRef]
- Panzer, A. Melatonin in osteosarcoma: An effective drug? Med. Hypotheses 1997, 48, 523–525. [Google Scholar] [CrossRef]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Reiter, R.J. Melatonin uses in oncology: Breast cancer prevention and reduction of the side effects of chemotherapy and radiation. Expert. Opin. Investig. Drugs 2012, 21, 819–831. [Google Scholar] [CrossRef] [PubMed]
- Acuna-Castroviejo, D.; Escames, G.; Venegas, C.; Diaz-Casado, M.E.; Lima-Cabello, E.; Lopez, L.C.; Rosales-Corral, S.; Tan, D.X.; Reiter, R.J. Extrapineal melatonin: Sources, regulation, and potential functions. Cell Mol. Life Sci. 2014, 71, 2997–3025. [Google Scholar] [CrossRef] [PubMed]
- Remy, P.; Doder, M.; Lees, A.; Turjanski, N.; Brooks, D. Depression in parkinson’s disease: Loss of dopamine and noradrenaline innervation in the limbic system. Brain 2005, 128, 1314–1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schomerus, C.; Korf, H.W. Mechanisms regulating melatonin synthesis in the mammalian pineal organ. Ann. N. Y. Acad. Sci. 2005, 1057, 372–383. [Google Scholar] [CrossRef] [PubMed]
- Reiter, R.J. Pineal melatonin: Cell biology of its synthesis and of its physiological interactions. Endocr. Rev. 1991, 12, 151–180. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Jiang, S.; Lu, C.; Yang, W.; Yang, Z.; Hu, W.; Xin, Z.; Yang, Y. Melatonin: Another avenue for treating osteoporosis? J. Pineal Res. 2019, 66, e12548. [Google Scholar] [CrossRef] [Green Version]
- Brzezinski, A. Melatonin in humans. N. Engl. J. Med. 1997, 336, 186–195. [Google Scholar] [CrossRef]
- Di, W.L.; Kadva, A.; Johnston, A.; Silman, R. Variable bioavailability of oral melatonin. N. Engl. J. Med. 1997, 336, 1028–1029. [Google Scholar] [CrossRef]
- Vijayalaxmi; Thomas, C.R., Jr.; Reiter, R.J.; Herman, T.S. Melatonin: From basic research to cancer treatment clinics. J. Clin. Oncol. 2002, 20, 2575–2601. [Google Scholar] [CrossRef]
- Tan, D.; Manchester, L.C.; Reiter, R.J.; Qi, W.; Hanes, M.A.; Farley, N.J. High physiological levels of melatonin in the bile of mammals. Life Sci. 1999, 65, 2523–2529. [Google Scholar] [CrossRef]
- Wetterberg, L. Melatonin and clinical application. Reprod. Nutr. Dev. 1999, 39, 367–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, H.J.; Wurtman, R.J.; Moskowitz, M.A.; Archer, M.C.; Ho, M.H. Daily rhythm in human urinary melatonin. Science 1975, 187, 169–171. [Google Scholar] [CrossRef] [PubMed]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stehle, J.H.; von Gall, C.; Korf, H.W. Melatonin: A clock-output, a clock-input. J. Neuroendocr. 2003, 15, 383–389. [Google Scholar] [CrossRef]
- Luchetti, F.; Canonico, B.; Betti, M.; Arcangeletti, M.; Pilolli, F.; Piroddi, M.; Canesi, L.; Papa, S.; Galli, F. Melatonin signaling and cell protection function. FASEB J. 2010, 24, 3603–3624. [Google Scholar] [CrossRef]
- Stevens, R.G.; Brainard, G.C.; Blask, D.E.; Lockley, S.W.; Motta, M.E. Breast cancer and circadian disruption from electric lighting in the modern world. CA Cancer J. Clin. 2014, 64, 207–218. [Google Scholar] [CrossRef]
- Carlberg, C. Gene regulation by melatonin. Ann. N. Y. Acad. Sci. 2000, 917, 387–396. [Google Scholar] [CrossRef]
- Reiter, R.J. Oxidative damage in the central nervous system: Protection by melatonin. Prog. Neurobiol. 1998, 56, 359–384. [Google Scholar] [CrossRef]
- Srinivasan, V.; Spence, D.W.; Pandi-Perumal, S.R.; Trakht, I.; Cardinali, D.P. Therapeutic actions of melatonin in cancer: Possible mechanisms. Integr. Cancer 2008, 7, 189–203. [Google Scholar] [CrossRef]
- Deming, S.L.; Lu, W.; Beeghly-Fadiel, A.; Zheng, Y.; Cai, Q.; Long, J.; Shu, X.O.; Gao, Y.T.; Zheng, W. Melatonin pathway genes and breast cancer risk among chinese women. Breast Cancer Res. Treat. 2012, 132, 693–699. [Google Scholar] [CrossRef] [Green Version]
- Maria, S.; Samsonraj, R.M.; Munmun, F.; Glas, J.; Silvestros, M.; Kotlarczyk, M.P.; Rylands, R.; Dudakovic, A.; van Wijnen, A.J.; Enderby, L.T.; et al. Biological effects of melatonin on osteoblast/osteoclast cocultures, bone, and quality of life: Implications of a role for mt2 melatonin receptors, mek1/2, and mek5 in melatonin-mediated osteoblastogenesis. J. Pineal Res. 2018, 64. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.M.; Belancio, V.P.; Dauchy, R.T.; Xiang, S.; Brimer, S.; Mao, L.; Hauch, A.; Lundberg, P.W.; Summers, W.; Yuan, L.; et al. Melatonin: An inhibitor of breast cancer. Endocr. Relat. Cancer 2015, 22, R183–R204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levi, B.; Longaker, M.T. Concise review: Adipose-derived stromal cells for skeletal regenerative medicine. Stem Cells 2011, 29, 576–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luchetti, F.; Canonico, B.; Bartolini, D.; Arcangeletti, M.; Ciffolilli, S.; Murdolo, G.; Piroddi, M.; Papa, S.; Reiter, R.J.; Galli, F. Melatonin regulates mesenchymal stem cell differentiation: A review. J. Pineal Res. 2014, 56, 382–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Su, P.; Xu, C.; Chen, C.; Liang, A.; Du, K.; Peng, Y.; Huang, D. Melatonin inhibits adipogenesis and enhances osteogenesis of human mesenchymal stem cells by suppressing ppargamma expression and enhancing runx2 expression. J. Pineal Res. 2010, 49, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Radio, N.M.; Doctor, J.S.; Witt-Enderby, P.A. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via mt2 melatonin receptors and the mek/erk (1/2) signaling cascade. J. Pineal Res. 2006, 40, 332–342. [Google Scholar] [CrossRef]
- Arendt, J. Melatonin and human rhythms. Chronobiol. Int. 2006, 23, 21–37. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Ladizesky, M.G.; Boggio, V.; Cutrera, R.A.; Mautalen, C. Melatonin effects on bone: Experimental facts and clinical perspectives. J. Pineal Res. 2003, 34, 81–87. [Google Scholar] [CrossRef]
- Park, K.H.; Kang, J.W.; Lee, E.M.; Kim, J.S.; Rhee, Y.H.; Kim, M.; Jeong, S.J.; Park, Y.G.; Kim, S.H. Melatonin promotes osteoblastic differentiation through the bmp/erk/wnt signaling pathways. J. Pineal Res. 2011, 51, 187–194. [Google Scholar] [CrossRef]
- Tan, D.X.; Manchester, L.C.; Fuentes-Broto, L.; Paredes, S.D.; Reiter, R.J. Significance and application of melatonin in the regulation of brown adipose tissue metabolism: Relation to human obesity. Obes. Rev. 2011, 12, 167–188. [Google Scholar] [CrossRef]
- Nakade, O.; Koyama, H.; Ariji, H.; Yajima, A.; Kaku, T. Melatonin stimulates proliferation and type i collagen synthesis in human bone cells in vitro. J. Pineal Res. 1999, 27, 106–110. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D. How important is the pineal gland in children. In Adv. Pineal Research; Reiter, R.J., Pang, S.F., Eds.; John Libbey: London, UK, 1989; pp. 291–297. [Google Scholar]
- Ho, H.Y.; Lin, C.W.; Chien, M.H.; Reiter, R.J.; Su, S.C.; Hsieh, Y.H.; Yang, S.F. Melatonin suppresses tpa-induced metastasis by downregulating matrix metalloproteinase-9 expression through jnk/sp-1 signaling in nasopharyngeal carcinoma. J. Pineal Res. 2016, 61, 479–492. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.H.; Su, S.C.; Lin, C.W.; Hsieh, Y.H.; Lin, Y.C.; Chien, M.H.; Reiter, R.J.; Yang, S.F. Melatonin attenuates osteosarcoma cell invasion by suppression of c-c motif chemokine ligand 24 through inhibition of the c-jun n-terminal kinase pathway. J. Pineal Res. 2018, 65, e12507. [Google Scholar] [CrossRef] [PubMed]
- Hoang, B.X.; Shaw, D.G.; Pham, P.T.; Levine, S.A. Neurobiological effects of melatonin as related to cancer. Eur. J. Cancer Prev. 2007, 16, 511–516. [Google Scholar] [CrossRef]
- Su, S.C.; Hsieh, M.J.; Yang, W.E.; Chung, W.H.; Reiter, R.J.; Yang, S.F. Cancer metastasis: Mechanisms of inhibition by melatonin. J. Pineal Res. 2017, 62. [Google Scholar] [CrossRef]
- Reiter, R.J.; Rosales-Corral, S.A.; Tan, D.X.; Acuna-Castroviejo, D.; Qin, L.; Yang, S.F.; Xu, K. Melatonin, a full service anti-cancer agent: Inhibition of initiation, progression and metastasis. Int. J. Mol. Sci. 2017, 18, 843. [Google Scholar] [CrossRef]
- Yu, Q.; Miller, S.C.; Osmond, D.G. Melatonin inhibits apoptosis during early b-cell development in mouse bone marrow. J. Pineal Res. 2000, 29, 86–93. [Google Scholar] [CrossRef]
- Bizzarri, M.; Proietti, S.; Cucina, A.; Reiter, R.J. Molecular mechanisms of the pro-apoptotic actions of melatonin in cancer: A review. Exp. Opin. Targets 2013, 17, 1483–1496. [Google Scholar] [CrossRef]
- Hardeland, R. Antioxidative protection by melatonin: Multiplicity of mechanisms from radical detoxification to radical avoidance. Endocrine 2005, 27, 119–130. [Google Scholar] [CrossRef]
- Maestroni, G.J. The immunoneuroendocrine role of melatonin. J. Pineal Res. 1993, 14, 1–10. [Google Scholar] [CrossRef]
- Garcia-Maurino, S.; Gonzalez-Haba, M.G.; Calvo, J.R.; Rafii-El-Idrissi, M.; Sanchez-Margalet, V.; Goberna, R.; Guerrero, J.M. Melatonin enhances il-2, il-6, and ifn-gamma production by human circulating cd4+ cells: A possible nuclear receptor-mediated mechanism involving t helper type 1 lymphocytes and monocytes. J. Immunol. 1997, 159, 574–581. [Google Scholar] [PubMed]
- Blask, D.E.; Brainard, G.C.; Dauchy, R.T.; Hanifin, J.P.; Davidson, L.K.; Krause, J.A.; Sauer, L.A.; Rivera-Bermudez, M.A.; Dubocovich, M.L.; Jasser, S.A.; et al. Melatonin-depleted blood from premenopausal women exposed to light at night stimulates growth of human breast cancer xenografts in nude rats. Cancer Res. 2005, 65, 11174–11184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leon-Blanco, M.M.; Guerrero, J.M.; Reiter, R.J.; Calvo, J.R.; Pozo, D. Melatonin inhibits telomerase activity in the mcf-7 tumor cell line both in vivo and in vitro. J. Pineal Res. 2003, 35, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Leon, J.; Casado, J.; Jimenez Ruiz, S.M.; Zurita, M.S.; Gonzalez-Puga, C.; Rejon, J.D.; Gila, A.; Munoz de Rueda, P.; Pavon, E.J.; Reiter, R.J.; et al. Melatonin reduces endothelin-1 expression and secretion in colon cancer cells through the inactivation of foxo-1 and nf-kappabeta. J. Pineal Res. 2014, 56, 415–426. [Google Scholar] [CrossRef]
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Lu, K.H.; Chen, P.N.; Lue, K.H.; Lai, M.T.; Lin, M.S.; Hsieh, Y.S.; Chu, S.C. 2’-hydroxyflavanone induces apoptosis of human osteosarcoma 143 b cells by activating the extrinsic trail- and intrinsic mitochondria-mediated pathways. Nutr. Cancer 2014, 66, 625–635. [Google Scholar] [CrossRef]
- Degterev, A.; Boyce, M.; Yuan, J. A decade of caspases. Oncogene 2003, 22, 8543–8567. [Google Scholar] [CrossRef] [Green Version]
- Davis, R.J. Signal transduction by the jnk group of map kinases. Cell 2000, 103, 239–252. [Google Scholar] [CrossRef] [Green Version]
- Karin, M.; Cao, Y.; Greten, F.R.; Li, Z.W. Nf-kappab in cancer: From innocent bystander to major culprit. Nat. Rev. Cancer 2002, 2, 301–310. [Google Scholar] [CrossRef]
- Toma, C.D.; Svoboda, M.; Arrich, F.; Ekmekcioglu, C.; Assadian, O.; Thalhammer, T. Expression of the melatonin receptor (mt) 1 in benign and malignant human bone tumors. J. Pineal Res. 2007, 43, 206–213. [Google Scholar] [CrossRef]
- Peng, T.I.; Hsiao, C.W.; Reiter, R.J.; Tanaka, M.; Lai, Y.K.; Jou, M.J. Mtdna t8993g mutation-induced mitochondrial complex v inhibition augments cardiolipin-dependent alterations in mitochondrial dynamics during oxidative, ca(2+), and lipid insults in narp cybrids: A potential therapeutic target for melatonin. J. Pineal Res. 2012, 52, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Panzer, A.; Lottering, M.L.; Bianchi, P.; Glencross, D.K.; Stark, J.H.; Seegers, J.C. Melatonin has no effect on the growth, morphology or cell cycle of human breast cancer (mcf-7), cervical cancer (hela), osteosarcoma (mg-63) or lymphoblastoid (tk6) cells. Cancer Lett. 1998, 122, 17–23. [Google Scholar] [CrossRef]
- Liu, L.; Xu, Y.; Reiter, R.J. Melatonin inhibits the proliferation of human osteosarcoma cell line mg-63. Bone 2013, 55, 432–438. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Xu, Y.; Reiter, R.J.; Pan, Y.; Chen, D.; Liu, Y.; Pu, X.; Jiang, L.; Li, Z. Inhibition of erk1/2 signaling pathway is involved in melatonin’s antiproliferative effect on human mg-63 osteosarcoma cells. Cell Physiol. Biochem. 2016, 39, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Cai, L.; Jiang, P.; Wang, J.; Gao, C.; Feng, H.; Wang, C.; Pan, H.; Yang, Y. Sirt1 inhibition by melatonin exerts antitumor activity in human osteosarcoma cells. Eur. J. Pharm. 2013, 715, 219–229. [Google Scholar] [CrossRef] [PubMed]
- Jung-Hynes, B.; Reiter, R.J.; Ahmad, N. Sirtuins, melatonin and circadian rhythms: Building a bridge between aging and cancer. J. Pineal Res. 2010, 48, 9–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valastyan, S.; Weinberg, R.A. Tumor metastasis: Molecular insights and evolving paradigms. Cell 2011, 147, 275–292. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, Y.S.; Chu, S.C.; Yang, S.F.; Chen, P.N.; Liu, Y.C.; Lu, K.H. Silibinin suppresses human osteosarcoma mg-63 cell invasion by inhibiting the erk-dependent c-jun/ap-1 induction of mmp-2. Carcinogenesis 2007, 28, 977–987. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H.L.; Lin, C.W.; Yang, J.S.; Hsieh, M.J.; Yang, S.F.; Lu, K.H. Zoledronate blocks geranylgeranylation not farnesylation to suppress human osteosarcoma u2os cells metastasis by emt via rho a activation and fak-inhibited jnk and p38 pathways. Oncotarget 2016, 7, 9742–9758. [Google Scholar] [CrossRef] [Green Version]
- Kahlert, U.D.; Joseph, J.V.; Kruyt, F.A.E. Emt- and met-related processes in nonepithelial tumors: Importance for disease progression, prognosis, and therapeutic opportunities. Mol. Oncol. 2017, 11, 860–877. [Google Scholar] [CrossRef]
- Kim, E.H.; Kim, M.S.; Lee, K.H.; Koh, J.S.; Jung, W.G.; Kong, C.B. Zoledronic acid is an effective radiosensitizer in the treatment of osteosarcoma. Oncotarget 2016, 7, 70869–70880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daw, N.C.; Chou, A.J.; Jaffe, N.; Rao, B.N.; Billups, C.A.; Rodriguez-Galindo, C.; Meyers, P.A.; Huh, W.W. Recurrent osteosarcoma with a single pulmonary metastasis: A multi-institutional review. Br. J. Cancer 2015, 112, 278–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung-Hynes, B.; Ahmad, N. Sirt1 controls circadian clock circuitry and promotes cell survival: A connection with age-related neoplasms. FASEB J. 2009, 23, 2803–2809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung-Hynes, B.; Ahmad, N. Role of p53 in the anti-proliferative effects of sirt1 inhibition in prostate cancer cells. Cell Cycle 2009, 8, 1478–1483. [Google Scholar] [CrossRef]
- Qu, H.; Xue, Y.; Lian, W.; Wang, C.; He, J.; Fu, Q.; Zhong, L.; Lin, N.; Lai, L.; Ye, Z.; et al. Melatonin inhibits osteosarcoma stem cells by suppressing sox9-mediated signaling. Life Sci. 2018, 207, 253–264. [Google Scholar] [CrossRef]
- Fan, L.L.; Sun, G.P.; Wei, W.; Wang, Z.G.; Ge, L.; Fu, W.Z.; Wang, H. Melatonin and doxorubicin synergistically induce cell apoptosis in human hepatoma cell lines. World J. Gastroenterol. 2010, 16, 1473–1481. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Z.; Chua, C.C.; Ma, Y.S.; Youngberg, G.A.; Hamdy, R.; Chua, B.H. Melatonin as an effective protector against doxorubicin-induced cardiotoxicity. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H254–H263. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.P.; Yang, Z.P. Effects of melatonin combined with cis-platinum or methotrexate on the proliferation of osteosarcoma cell line saos-2. Acta Acad. Med. Sin. 2015, 37, 215–220. [Google Scholar]
- Nataraj, V.; Rastogi, S.; Khan, S.A.; Sharma, M.C.; Agarwala, S.; Vishnubhatla, S.; Bakhshi, S. Prognosticating metastatic osteosarcoma treated with uniform chemotherapy protocol without high dose methotrexate and delayed metastasectomy: A single center experience of 102 patients. Clin. Transl. Oncol. 2016, 18, 937–944. [Google Scholar] [CrossRef]
- Liu, L.; Zhu, Y.; Xu, Y.; Reiter, R.J. Melatonin delays cell proliferation by inducing g1 and g2/m phase arrest in a human osteoblastic cell line hfob 1.19. J. Pineal Res. 2011, 50, 222–231. [Google Scholar] [CrossRef]
- Altindal, D.C.; Gumusderelioglu, M. Melatonin releasing plga micro/nanoparticles and their effect on osteosarcoma cells. J. Microencapsul. 2016, 33, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Iguchi, H.; Kato, K.I.; Ibayashi, H. Melatonin serum levels and metabolic clearance rate in patients with liver cirrhosis. J. Clin. Endocrinol. Metab. 1982, 54, 1025–1027. [Google Scholar] [CrossRef] [PubMed]
- Topal, B.; Cetin Altindal, D.; Gumusderelioglu, M. Melatonin/hpbetacd complex: Microwave synthesis, integration with chitosan scaffolds and inhibitory effects on mg-63cells. Int. J. Pharm. 2015, 496, 801–811. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
The process of the melatonin’s biosynthesis.

Figure 2.
Multiple functions of melatonin relating to the bone.

Figure 3.
A summary of various signaling pathways involved in melatonin on human osteosarcoma. MT1: melatonin receptor 1; ERK: extracellular signal-regulated protein kinase; JNK: c-Jun N-terminal kinase; ROS: reactive oxygen species; GSH: glutathione; Bcl-2: B-cell lymphoma/leukemia 2; BAX: Bcl-2-associated X protein; SIRT1: sirtuin 1; EMT: epithelial–mesenchymal transition; CCL24: C-C motif chemokine ligand 24; and MG-63, 143B, HOS, U2OS, and SOSP-9607: human osteosarcoma cell lines; Stem: osteosarcoma stem cells.
Figure 3.
A summary of various signaling pathways involved in melatonin on human osteosarcoma. MT1: melatonin receptor 1; ERK: extracellular signal-regulated protein kinase; JNK: c-Jun N-terminal kinase; ROS: reactive oxygen species; GSH: glutathione; Bcl-2: B-cell lymphoma/leukemia 2; BAX: Bcl-2-associated X protein; SIRT1: sirtuin 1; EMT: epithelial–mesenchymal transition; CCL24: C-C motif chemokine ligand 24; and MG-63, 143B, HOS, U2OS, and SOSP-9607: human osteosarcoma cell lines; Stem: osteosarcoma stem cells.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Cytotoxic activity of melatonin in human osteosarcoma in vitro and in vivo.
| Action | Pathway | Cell Line/In Vivo | Dose | Reference |
|---|---|---|---|---|
| No effect on the growth, morphology or cell cycle | MG-63 | 10−5–10−13 M | Panzer A, et al. 1998. [64] | |
| Abundant expression of MT1-mRNA | HOS, MG-63 | Toma CD, et al. 2007. [62] | ||
| Rescues cardiolipin-dependent mitochondrial dynamics-associated mitochondrial pathologies | Reduces mitochondrial ROS, cell death, and depletion of cardiolipin to improve retardation of mitochondrial movement and dynamics | 143B | 100 μM | Peng TI, et al. 2012. [63] |
| Inhibits proliferation | Down-regulates cyclin D1 and CDK4, (G1 phase), and cyclin B1 and CDK1 (G2/M phase) | MG-63 | 4(−10) mM | Liu L, et al. 2013. [65] |
| Induces apoptosis, increases ROS, and decreases GSH | Down-regulates SIRT1 and mitochondrial apoptotic pathway, and up-regulates acetylated-p53 | SOSP-9607 | (250−)1000 μM | Cheng Y, et al. 2013. [67] |
| Reduces adhesion and migration | 12.5–50 μM | |||
| Inhibits proliferation, induces G1 and G2/M phase arrest, and down-regulates cyclin D1, CDK4, cyclin B1 and CDK1 | Inhibits the ERK1/2 pathway | MG-63 | 4 mM | Liu L, et al. 2016. [66] |
MT1: melatonin receptor 1; ROS: reactive oxygen species; CDK: cyclin-dependent kinase; GSH: glutathione; SIRT1: sirtuin 1; ERK: extracellular signal-regulated protein kinase; and MG-63, HOS, 143B, and SOSP-9607: human osteosarcoma cell lines.
Table 2.
Anti-metastatic effects of melatonin in human osteosarcoma in vitro and in vivo.
| Action | Pathway | Cell Line/In Vivo | Dose | Reference |
|---|---|---|---|---|
| Induces apoptosis, increases ROS, and decreases GSH | Down-regulates SIRT1 and mitochondrial apoptotic pathway, and up-regulates acetylated-p53 | SOSP-9607 | (250−)1000 μM | Cheng Y, et al. 2013. [67] |
| Reduces adhesion and migration | 12.5–50 μM | |||
| Reduces the number (anti-proliferation) | Suppresses EMT via downregulation of SOX9, via the ERK pathway | Stem cell | >0.5 mM | Qu H, et al. 2018. [77] |
| Suppresses migration and invasion | U2OS, HOS | 0.5 mM | ||
| Inhibits initiation and metastasis in vivo | Mice model | 100 mg/kg | ||
| Inhibits motility, migration and invasiveness, and the CCL24 gene expression | Attenuates invasion and migration by suppression of CCL24 through inhibition of the JNK pathway | U2OS, HOS | 2 mM | Lu KH, et al. 2018. [45] |
| Recombinant human CCL24 enhances migration and silencing of CCL24 attenuates migration and invasion | ||||
| Suppresses CCL24 and anti-metastasis through inhibition of the JNK pathway |
ROS: reactive oxygen species; GSH: glutathione; SIRT1: sirtuin 1; EMT: epithelial–mesenchymal transition; SOX9: a transcription factor; CCL24: C-C motif chemokine ligand 24; JNK: c-Jun N-terminal kinase; and SOSP-9607, U2OS, and HOS: human osteosarcoma cell lines; Stem: osteosarcoma stem cells.
Table 3.
Synergistic effects and micro/nanoparticles delivery and inclusion complex systems of melatonin in human osteosarcoma in vitro and in vivo.
Table 3.
Synergistic effects and micro/nanoparticles delivery and inclusion complex systems of melatonin in human osteosarcoma in vitro and in vivo.
| Action | Pathway | Cell Line/In Vivo | Dose | Reference |
|---|---|---|---|---|
| Inhibits cell activity, blocks the cell cycle at G1-stage, and induces apoptosis | Blocks the cell cycle at G1-stage | SaOS-2 | 0.5–5 mM | Wang YP, et al. 2015. [80] |
| An antagonistic effect with lower concentration of cis-platinum | 1 mM | |||
| A synergistic effect with methotrexate or higher concentration of cis-platinum | ||||
| Melatonin releasing PLGA micro/nanoparticles increases the inhibitory effect | Increases the inhibitory effect of proliferation | MG-63 | Altındal DÇ, et al. 2016. [83] | |
| Melatonin/HPβCD inclusion complex loaded into chitosan scaffolds causes cell death | Melatonin/HPβCD system reduces the proportion in the G2/M phase rather than S phase | MG-63 | 9 mM | Topal B, et al. 2015. [85] |
PLGA: poly(d,l-lactide-co-glycolide); HPβCD: 2-hydroxypropyl-β-cyclodextrin; MG-63, SaOS-2: human osteosarcoma cell lines.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lu, K.-H.; Lin, R.-C.; Yang, J.-S.; Yang, W.-E.; Reiter, R.J.; Yang, S.-F. Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma. Cells 2019, 8, 1618. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121618
AMA Style
Lu K-H, Lin R-C, Yang J-S, Yang W-E, Reiter RJ, Yang S-F. Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma. Cells. 2019; 8(12):1618. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121618
Chicago/Turabian StyleLu, Ko-Hsiu, Renn-Chia Lin, Jia-Sin Yang, Wei-En Yang, Russel J. Reiter, and Shun-Fa Yang. 2019. "Molecular and Cellular Mechanisms of Melatonin in Osteosarcoma" Cells 8, no. 12: 1618. https://0-doi-org.brum.beds.ac.uk/10.3390/cells8121618
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.