Stemness Potency of Human Gingival Cells—Application in Anticancer Therapies and Clinical Trials

 , , ,
, , ,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Methods of GMSCs Isolation and Cultivation
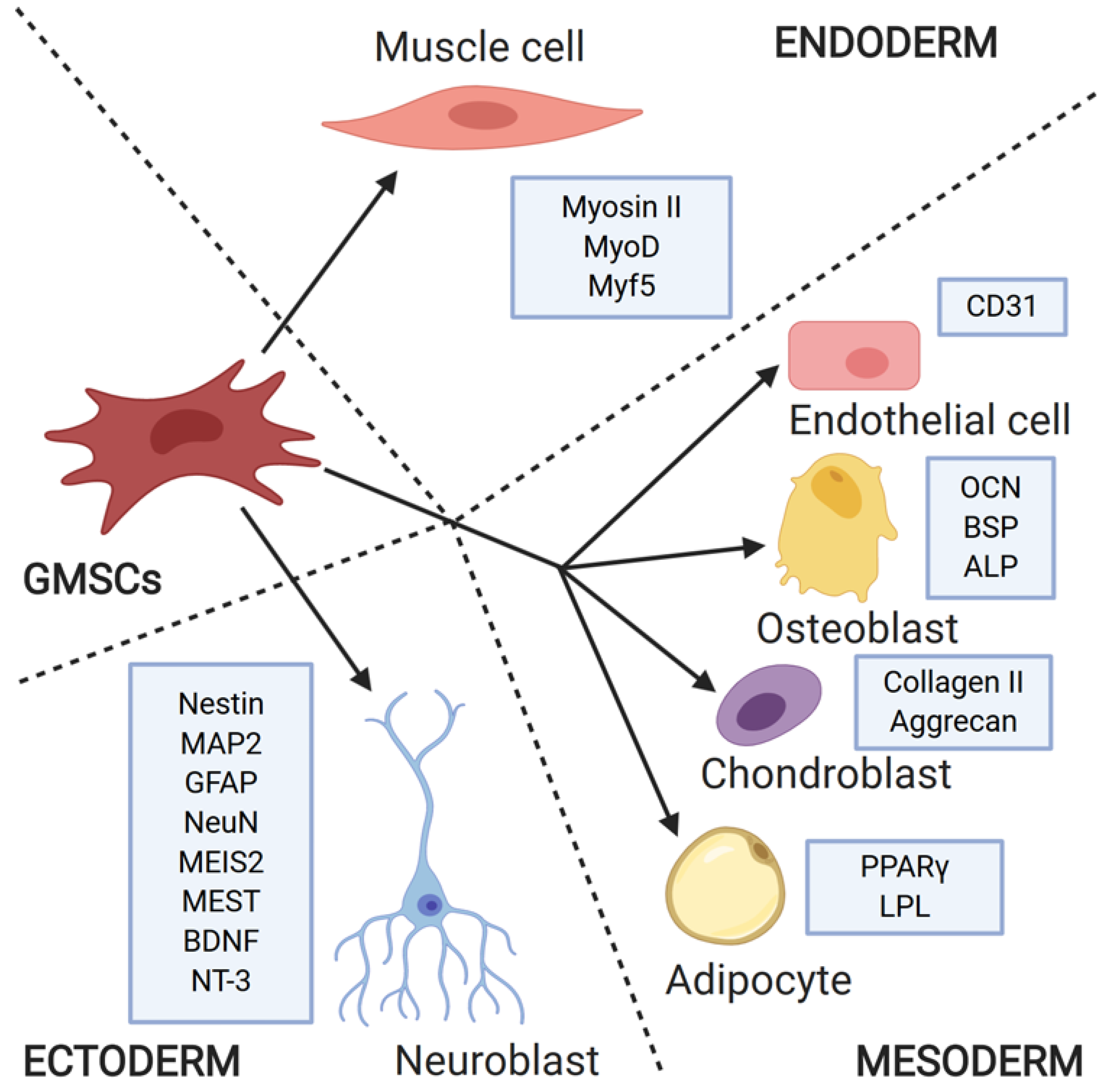
3. Stemness Properties and Differentiation Ability of GMSCs
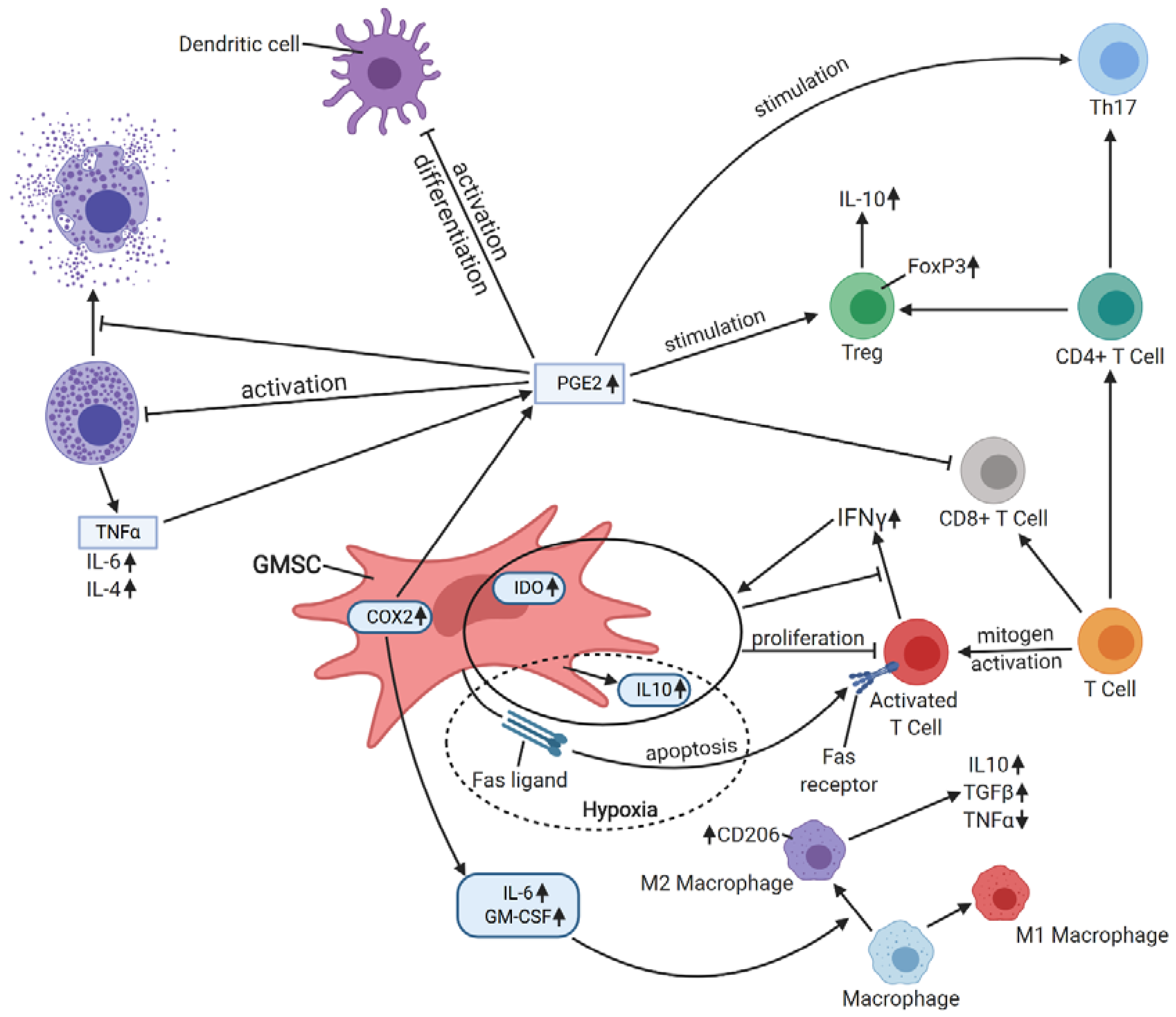
4. Immunomodulatory Properties
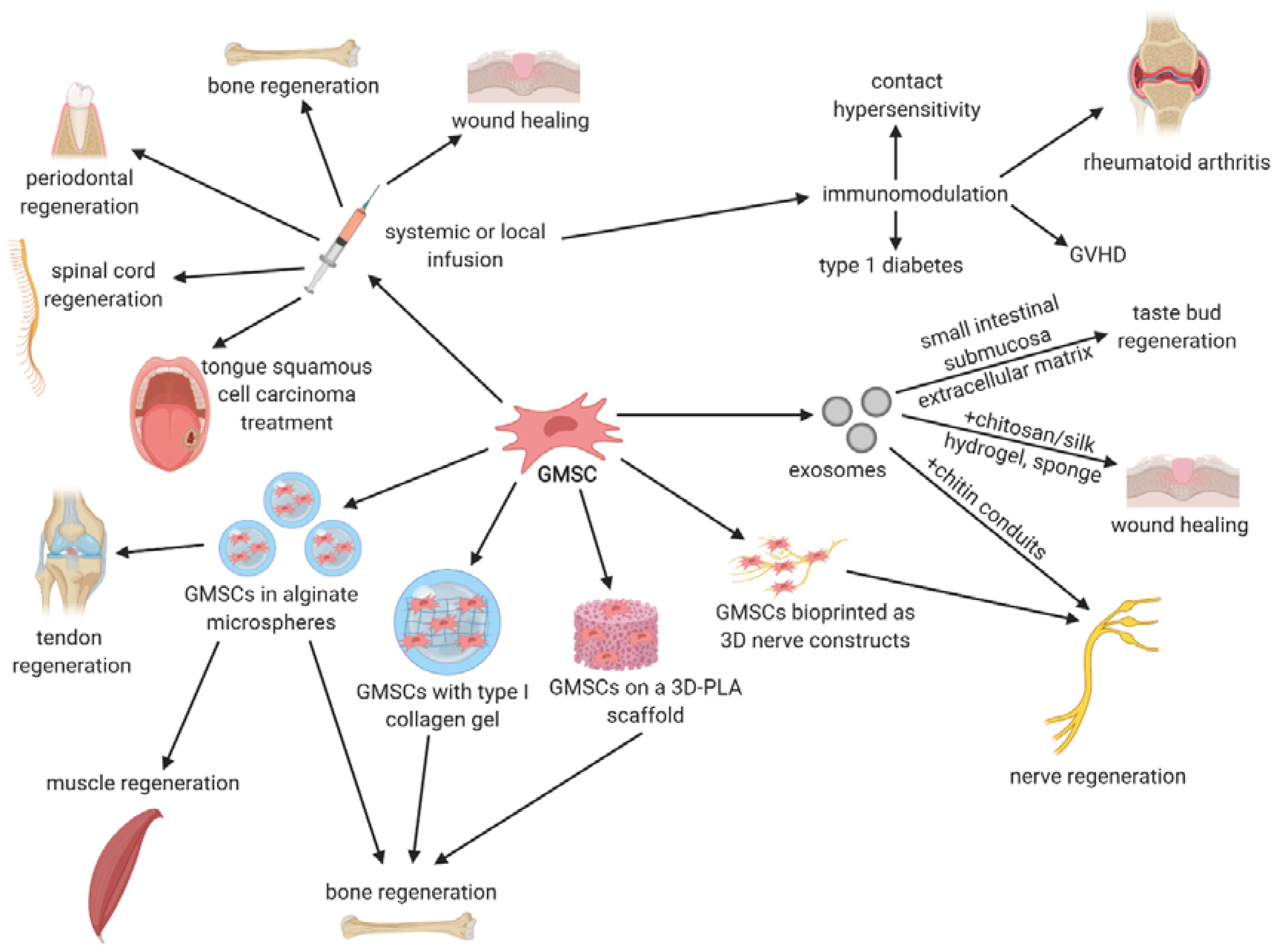
5. Preclinical Studies with GMSCs
5.1. Anticancer Therapies
5.2. Oral Mucositis
5.3. Tissue Regeneration
5.4. Immunologic- and Inflammatory-Related Disorders
6. Clinical Application in Stem Cell Therapies
7. Therapeutic Potential of GMSCs-Derived Exosomes
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal stem cells in health and disease. Nat. Rev. Immunol. 2008. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The Development of Fibroblast Colonies in Monolayer Cultures of Guinea-Pig Bone Marrow and Spleen Cells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, H.E.; Deepe, R.; Hoelscher, G.L.; Ingram, J.A.; Norton, H.J.; Scannell, B.; Loeffler, B.J.; Zinchenko, N.; Hanley, E.N.; Tapp, H. Human adipose-derived mesenchymal stem cells: Direction to a phenotype sharing similarities with the disc, gene expression profiling, and coculture with human annulus cells. Tissue Eng. Part A 2010, 16, 2843–2860. [Google Scholar] [CrossRef]
- In’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; de Groot-Swings, G.M.J.S.; Claas, F.H.J.; Fibbe, W.E.; Kanhai, H.H.H. Isolation of Mesenchymal Stem Cells of Fetal or Maternal Origin from Human Placenta. Stem Cells 2004. [Google Scholar] [CrossRef]
- In ’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.J.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 2003. [Google Scholar] [CrossRef]
- Campagnoli, C.; Roberts, I.A.G.; Kumar, S.; Bennett, P.R.; Bellantuono, I.; Fisk, N.M. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver, and bone marrow. Blood 2001. [Google Scholar] [CrossRef]
- Secco, M.; Zucconi, E.; Vieira, N.M.; Fogaça, L.L.Q.; Cerqueira, A.; Carvalho, M.D.F.; Jazedje, T.; Okamoto, O.K.; Muotri, A.R.; Zatz, M. Multipotent Stem Cells from Umbilical Cord: Cord Is Richer than Blood! Stem Cells 2008, 26, 146–150. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.; Ye, S.; Zhou, X.; Liu, D.; Ying, Q.L. Molecular basis of embryonic stem cell self-renewal: From signaling pathways to pluripotency network. Cell. Mol. Life Sci. 2015, 72, 1741–1757. [Google Scholar] [CrossRef] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000. [Google Scholar] [CrossRef] [Green Version]
- Miura, M.; Gronthos, S.; Zhao, M.; Lu, B.; Fisher, L.W.; Robey, P.G.; Shi, S. SHED: Stem cells from human exfoliated deciduous teeth. Proc. Natl. Acad. Sci. USA 2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, B.M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004. [Google Scholar] [CrossRef]
- Morsczeck, C.; Götz, W.; Schierholz, J.; Zeilhofer, F.; Kühn, U.; Möhl, C.; Sippel, C.; Hoffmann, K.H. Isolation of precursor cells (PCs) from human dental follicle of wisdom teeth. Matrix Biol. 2005, 24, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, T.; Suardita, K.; Ishii, M.; Sugiyama, M.; Igarashi, A.; Oda, R.; Nishimura, M.; Saito, M.; Nakagawa, K.; Yamanaka, K.; et al. Alveolar bone marrow as a cell source for regenerative medicine: Differences between alveolar and iliac bone marrow stromal cells. J. Bone Miner. Res. 2005, 20, 399–409. [Google Scholar] [CrossRef]
- Sonoyama, W.; Liu, Y.; Fang, D.; Yamaza, T.; Seo, B.M.; Zhang, C.; Liu, H.; Gronthos, S.; Wang, C.Y.; Shi, S.; et al. Mesenchymal stem cell-mediated functional tooth regeneration in Swine. PLoS ONE 2006. [Google Scholar] [CrossRef] [Green Version]
- Ikeda, E.; Yagi, K.; Kojima, M.; Yagyuu, T.; Ohshima, A.; Sobajima, S.; Tadokoro, M.; Katsube, Y.; Isoda, K.; Kondoh, M.; et al. Multipotent cells from the human third molar: Feasibility of cell-based therapy for liver disease. Differentiation 2008. [Google Scholar] [CrossRef]
- Zhang, Q.; Shi, S.; Liu, Y.; Uyanne, J.; Shi, Y.; Shi, S.; Le, A.D. Mesenchymal Stem Cells Derived from Human Gingiva Are Capable of Immunomodulatory Functions and Ameliorate Inflammation-Related Tissue Destruction in Experimental Colitis. J. Immunol. 2009, 183, 7787–7798. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Chen, C.; Akiyama, K.; Chai, Y.; Le, A.D.; Wang, Z.; Shi, S. Gingivae contain neural-crest- and mesoderm-derived mesenchymal stem cells. J. Dent. Res. 2013. [Google Scholar] [CrossRef] [Green Version]
- Fawzy El-Sayed, K.M.; Dörfer, C.E. Gingival Mesenchymal Stem/Progenitor Cells: A Unique Tissue Engineering Gem. Stem Cells Int. 2016. [Google Scholar] [CrossRef] [Green Version]
- Eslami, A.; Gallant-Behm, C.L.; Hart, D.A.; Wiebe, C.; Honardoust, D.; Gardner, H.; Häkkinen, L.; Larjava, H.S. Expression of integrin αvβ6 and TGF-β in Scarless vs Scar-forming wound healing. J. Histochem. Cytochem. 2009. [Google Scholar] [CrossRef] [Green Version]
- Lodi, D.; Iannitti, T.; Palmieri, B. Stem cells in clinical practice: Applications and warnings. J. Exp. Clin. Cancer Res. 2011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitrano, T.I.; Grob, M.S.; Carrión, F.; Nova-Lamperti, E.; Luz, P.A.; Fierro, F.S.; Quintero, A.; Chaparro, A.; Sanz, A. Culture and Characterization of Mesenchymal Stem Cells From Human Gingival Tissue. J. Periodontol. 2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed, K.M.F.; Paris, S.; Graetz, C.; Kassem, N.; Mekhemar, M.; Ungefroren, H.; Fändrich, F.; Dörfer, C. Isolation and characterisation of human gingival margin-derived STRO-1/MACS (+) and MACS (−) cell populations. Int. J. Oral Sci. 2015. [Google Scholar] [CrossRef] [Green Version]
- El-Bialy, T.; Alhadlaq, A.; Wong, B.; Kucharski, C. Ultrasound effect on neural differentiation of gingival stem/progenitor cells. Ann. Biomed. Eng. 2014. [Google Scholar] [CrossRef] [PubMed]
- Fournier, B.P.J.; Ferre, F.C.; Couty, L.; Lataillade, J.J.; Gourven, M.; Naveau, A.; Coulomb, B.; Lafont, A.; Gogly, B. Multipotent progenitor cells in gingival connective tissue. Tissue Eng. Part A 2010. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Ranga Rao, S.; Subbarayan, R. Passage-dependent expression of STRO-1 in human gingival mesenchymal stem cells. J. Cell. Biochem. 2019. [Google Scholar] [CrossRef]
- Gao, Y.; Zhao, G.; Li, D.; Chen, X.; Pang, J.; Ke, J. Isolation and multiple differentiation potential assessment of human gingival mesenchymal stem cells. Int. J. Mol. Sci. 2014, 15, 20982–20996. [Google Scholar] [CrossRef] [Green Version]
- Tang, L.; Li, N.; Xie, H.; Jin, Y. Characterization of mesenchymal stem cells from human normal and hyperplastic gingiva. J. Cell. Physiol. 2011. [Google Scholar] [CrossRef]
- Tomar, G.B.; Srivastava, R.K.; Gupta, N.; Barhanpurkar, A.P.; Pote, S.T.; Jhaveri, H.M.; Mishra, G.C.; Wani, M.R. Human gingiva-derived mesenchymal stem cells are superior to bone marrow-derived mesenchymal stem cells for cell therapy in regenerative medicine. Biochem. Biophys. Res. Commun. 2010. [Google Scholar] [CrossRef]
- Ge, S.; Mrozik, K.M.; Menicanin, D.; Gronthos, S.; Bartold, P.M. Isolation and characterization of mesenchymal stem cell-like cells from healthy and inflamed gingival tissue: Potential use for clinical therapy. Regen. Med. 2012. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Lee, J.E.; Yun, J.H.; Kim, I.; Ko, Y.; Park, J.B. Isolation and characterization of human mesenchymal stem cells from gingival connective tissue. J. Periodontal Res. 2015, 50, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.Z.; Nguyen, A.L.; Yu, W.H.; Le, A.D. Human oral mucosa and gingiva: A unique reservoir for mesenchymal stem Cells. J. Dent. Res. 2012, 91, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Nguyen, A.L.; Shi, S.; Hill, C.; Wilder-Smith, P.; Krasieva, T.B.; Le, A.D. Three-dimensional spheroid culture of human gingiva-derived mesenchymal stem cells enhances mitigation of chemotherapy-induced oral mucositis. Stem Cells Dev. 2012. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.M.; Chiu, H.C.; Chin, Y.T.; Lin, H.Y.; Chiang, C.Y.; Tu, H.P.; Fu, M.M.J.; Fu, E. Effects of enamel matrix derivative on the proliferation and osteogenic differentiation of human gingival mesenchymal stem cells. Stem Cell Res. Ther. 2014. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Nakata, H.; Yamamoto, M.; Kasugai, S.; Kuroda, S. Comparison of gingiva-derived and bone marrow mesenchymal stem cells for osteogenesis. J. Cell. Mol. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.I.; Ko, Y.; Park, J.B. Evaluation of the osteogenic differentiation of gingiva-derived stem cells grown on culture plates or in stem cell spheroids: Comparison of two- and three-dimensional cultures. Exp. Ther. Med. 2017. [Google Scholar] [CrossRef] [Green Version]
- Yuan, W.X.; Wang, X.X.; Zheng, D.H.; Ma, D.; Cui, Q.; Yang, F.; Zhang, J. Muscone promotes the adipogenic differentiation of human gingival mesenchymal stem cells by inhibiting the wnt/β-catenin signaling pathway. Drug Des. Devel. Ther. 2019. [Google Scholar] [CrossRef] [Green Version]
- Marynka-Kalmani, K.; Treves, S.; Yafee, M.; Rachima, H.; Gafni, Y.; Cohen, M.A.; Pitaru, S. The lamina propria of adult human oral mucosa harbors a novel stem cell population. Stem Cells 2010. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, A.; Diomede, F.; Scionti, D.; Bramanti, P.; Trubiani, O.; Mazzon, E. The Role of Hypoxia on the Neuronal Differentiation of Gingival Mesenchymal Stem Cells: A Transcriptional Study. Cell Transplant. 2019. [Google Scholar] [CrossRef] [Green Version]
- Soundara Rajan, T.; Giacoppo, S.; Scionti, D.; Diomede, F.; Grassi, G.; Pollastro, F.; Piattelli, A.; Bramanti, P.; Mazzon, E.; Trubiani, O. Cannabidiol Activates Neuronal Precursor Genes in Human Gingival Mesenchymal Stromal Cells. J. Cell. Biochem. 2017. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.R.; Subbarayan, R.; Dinesh, M.G.; Arumugam, G.; Raja, S.T.K. Differentiation of human gingival mesenchymal stem cells into neuronal lineages in 3D bioconjugated injectable protein hydrogel construct for the management of neuronal disorder. Exp. Mol. Med. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansari, S.; Chen, C.; Xu, X.; Annabi, N.; Zadeh, H.H.; Wu, B.M.; Khademhosseini, A.; Shi, S.; Moshaverinia, A. Muscle Tissue Engineering Using Gingival Mesenchymal Stem Cells Encapsulated in Alginate Hydrogels Containing Multiple Growth Factors. Ann. Biomed. Eng. 2016, 44, 1908–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.L.; Liu, W.; Wu, Y.M.; Sun, W.L.; Dörfer, C.E.; Fawzy El-Sayed, K.M. Oral Mesenchymal Stem/Progenitor Cells: The Immunomodulatory Masters. Stem Cells Int. 2020. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Ren, G.; Huang, Y.; Su, J.; Han, Y.; Li, J.; Chen, X.; Cao, K.; Chen, Q.; Shou, P.; et al. Mesenchymal stem cells: A double-edged sword in regulating immune responses. Cell Death Differ. 2012. [Google Scholar] [CrossRef]
- Davies, L.C.; Lönnies, H.; Locke, M.; Sundberg, B.; Rosendahl, K.; Götherström, C.; Le Blanc, K.; Stephens, P. Oral mucosal progenitor cells are potently immunosuppressive in a dose-independent manner. Stem Cells Dev. 2012. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.M.; Liu, J.; Zhao, J.Y.; Xiao, L.; An, S.; Gou, Y.C.; Quan, H.X.; Cheng, Q.; Zhang, Y.L.; He, W.; et al. Effects of hypoxia on the immunomodulatory properties of human gingiva-derived mesenchymal stem cells. J. Dent. Res. 2015. [Google Scholar] [CrossRef]
- Akiyama, K.; Chen, C.; Wang, D.; Xu, X.; Qu, C.; Yamaza, T.; Cai, T.; Chen, W.; Sun, L.; Shi, S. Mesenchymal-stem-cell-induced immunoregulation involves FAS-ligand-/FAS-mediated T cell apoptosis. Cell Stem Cell 2012. [Google Scholar] [CrossRef] [Green Version]
- Su, W.R.; Zhang, Q.Z.; Shi, S.H.; Nguyen, A.L.; Le, A.D. Human gingiva-derived mesenchymal stromal cells attenuate contact hypersensitivity via prostaglandin E 2-dependent mechanisms. Stem Cells 2011. [Google Scholar] [CrossRef]
- Sayed, B.A.; Christy, A.; Quirion, M.R.; Brown, M.A. The Master Switch: The Role of Mast Cells in Autoimmunity and Tolerance. Annu. Rev. Immunol. 2008. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Su, W.R.; Shi, S.H.; Wilder-Smith, P.; Xiang, A.P.; Wong, A.; Nguyen, A.L.; Kwon, C.W.; Le, A.D. Human gingiva-derived mesenchymal stem cells elicit polarization of M2 macrophages and enhance cutaneous wound healing. Stem Cells 2010. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Chen, J.; Huang, F.; Wang, J.; Su, W.; Zhou, J.; Qi, Q.; Cao, F.; Sun, B.; Liu, Z.; et al. Human gingiva tissue-derived MSC ameliorates immune-mediated bone marrow failure of aplastic anemia via suppression of Th1 and Th17 cells and enhancement of CD4 + Foxp3 + regulatory T cells differentiation. Am. J. Transl. Res. 2019, 11, 7627–7643. [Google Scholar]
- Kidd, S.; Spaeth, E.; Dembinski, J.L.; Dietrich, M.; Watson, K.; Klopp, A.; Battula, V.L.; Weil, M.; Andreeff, M.; Marini, F.C. Direct evidence of mesenchymal stem cell tropism for tumor and wounding microenvironments using in vivo bioluminescent imaging. Stem Cells 2009, 27, 2614–2623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhu, K.; Ke, C.; Yang, S.; Yang, F.; Li, Z.; Zhang, Z. Mesenchymal stem cells inhibited development of lung cancer induced by chemical carcinogens in a rat model. Am. J. Transl. Res. 2017, 9, 2891–2900. [Google Scholar] [PubMed]
- Albarenque, S.M.; Zwacka, R.M.; Mohr, A. Both human and mouse mesenchymal stem cells promote breast cancer metastasis. Stem Cell Res. 2011. [Google Scholar] [CrossRef] [PubMed]
- Walter, M.; Liang, S.; Ghosh, S.; Hornsby, P.J.; Li, R. Interleukin 6 secreted from adipose stromal cells promotes migration and invasion of breast cancer cells. Oncogene 2009, 28, 2745–2755. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Lee, W.Y.W.; Rui, Y.F.; Cheng, T.Y.; Jiang, X.H.; Li, G. Bone marrow-derived mesenchymal stem cells promote growth and angiogenesis of breast and prostate tumors. Stem Cell Res. Ther. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- El-Haibi, C.P.; Bell, G.W.; Zhang, J.; Collmann, A.Y.; Wood, D.; Scherber, C.M.; Csizmadia, E.; Mariani, O.; Zhu, C.; Campagne, A.; et al. Critical role for lysyl oxidase in mesenchymal stem cell-driven breast cancer malignancy. Proc. Natl. Acad. Sci. USA 2012, 109, 17460–17465. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.A.; Meyer, J.R.; Greco, S.J.; Corcoran, K.E.; Bryan, M.; Rameshwar, P. Mesenchymal Stem Cells Protect Breast Cancer Cells through Regulatory T Cells: Role of Mesenchymal Stem Cell-Derived TGF-β. J. Immunol. 2010, 184, 5885–5894. [Google Scholar] [CrossRef]
- François, S.; Usunier, B.; Forgue-Lafitte, M.E.; L’Homme, B.; Benderitter, M.; Douay, L.; Gorin, N.C.; Larsen, A.K.; Chapel, A. Mesenchymal Stem Cell Administration Attenuates Colon Cancer Progression by Modulating the Immune Component within the Colorectal Tumor Microenvironment. Stem Cells Transl. Med. 2019, 8, 285–300. [Google Scholar] [CrossRef] [Green Version]
- Ryu, H.; Oh, J.E.; Rhee, K.J.; Baik, S.K.; Kim, J.; Kang, S.J.; Sohn, J.H.; Choi, E.; Shin, H.C.; Kim, Y.M.; et al. Adipose tissue-derived mesenchymal stem cells cultured at high density express IFN-β and suppress the growth of MCF-7 human breast cancer cells. Cancer Lett. 2014, 352, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Zhang, Z.; Han, Y.; Song, J.; Xu, X.; Jin, J.; Su, S.; Mu, D.; Liu, X.; Xu, S.; et al. Mesenchymal stem cells derived from normal gingival tissue inhibit the proliferation of oral cancer cells in vitro and in vivo. Int. J. Oncol. 2016, 49, 2011–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, L.; Peng, R.; Leng, W.; Jia, R.; Zeng, X.; Yang, X.; Fan, M. TRAIL-expressing gingival-derived mesenchymal stem cells inhibit tumorigenesis of tongue squamous cell carcinoma. J. Dent. Res. 2015. [Google Scholar] [CrossRef]
- Sgaramella, N.; Gu, X.; Boldrup, L.; Coates, P.J.; Fahraeus, R.; Califano, L.; Tartaro, G.; Colella, G.; Spaak, L.N.; Strom, A.; et al. Searching For New Targets And Treatments In The Battle Against Squamous Cell Carcinoma Of The Head And Neck, With Specific Focus On Tumours Of The Tongue. Curr. Top. Med. Chem. 2018, 18. [Google Scholar] [CrossRef] [PubMed]
- Imamura, Y.; Tashiro, H.; Tsend-Ayush, G.; Haruta, M.; Dashdemberel, N.; Komohara, Y.; Tsuboki, J.; Takaishi, K.; Ohba, T.; Nishimura, Y.; et al. Novel therapeutic strategies for advanced ovarian cancer by using induced pluripotent stem cell-derived myelomonocytic cells producing interferon beta. Cancer Sci. 2018, 109, 3403–3410. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Xie, D.Y.; Lin, B.L.; Zhang, G.L.; Wang, P.P.; Peng, L.; Gao, Z.L. Interferon-β gene-modified human bone marrow mesenchymal stem cells attenuate hepatocellular carcinoma through inhibiting AKT/FOXO3a pathway. Br. J. Cancer 2013, 109, 1198–1205. [Google Scholar] [CrossRef] [Green Version]
- Matsuzuka, T.; Rachakatla, R.S.; Doi, C.; Maurya, D.K.; Ohta, N.; Kawabata, A.; Pyle, M.M.; Pickel, L.; Reischman, J.; Marini, F.; et al. Human umbilical cord matrix-derived stem cells expressing interferon-β gene significantly attenuate bronchioloalveolar carcinoma xenografts in SCID mice. Lung Cancer 2010, 70, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Du, L.; Liang, Q.; Ge, S.; Yang, C.; Yang, P. The growth inhibitory effect of human gingiva-derived mesenchymal stromal cells expressing interferon-β on tongue squamous cell carcinoma cells and xenograft model. Stem Cell Res. Ther. 2019. [Google Scholar] [CrossRef] [Green Version]
- Coccè, V.; Farronato, D.; Brini, A.T.; Masia, C.; Giannì, A.B.; Piovani, G.; Sisto, F.; Alessandri, G.; Angiero, F.; Pessina, A. Drug Loaded Gingival Mesenchymal Stromal Cells (GinPa-MSCs) Inhibit In Vitro Proliferation of Oral Squamous Cell Carcinoma. Sci. Rep. 2017. [Google Scholar] [CrossRef]
- Moshaverinia, A.; Xu, X.; Chen, C.; Ansari, S.; Zadeh, H.H.; Snead, M.L.; Shi, S. Application of stem cells derived from the periodontal ligament orgingival tissue sources for tendon tissue regeneration. Biomaterials 2014. [Google Scholar] [CrossRef] [Green Version]
- Moshaverinia, A.; Chen, C.; Xu, X.; Akiyama, K.; Ansari, S.; Zadeh, H.H.; Shi, S. Bone regeneration potential of stem cells derived from periodontal ligament or gingival tissue sources encapsulated in RGD-modified alginate scaffold. Tissue Eng. Part A 2014, 20, 611–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.C.; Wang, Z.G.; Ji, Q.X.; Yu, X.B.; Xu, X.Y.; Yuan, C.Q.; Deng, J.; Yang, P.S. Systemically transplanted human gingiva-derived mesenchymal stem cells contributing to bone tissue regeneration. Int. J. Clin. Exp. Pathol. 2014, 7, 4922. [Google Scholar]
- Wang, F.; Yu, M.; Yan, X.; Wen, Y.; Zeng, Q.; Yue, W.; Yang, P.; Pei, X. Gingiva-derived mesenchymal stem cell-mediated therapeutic approach for bone tissue regeneration. Stem Cells Dev. 2011. [Google Scholar] [CrossRef] [PubMed]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.F.; Mazzon, E.; Trubiani, O. Biotherapeutic effect of gingival stem cells conditioned medium in bone tissue restoration. Int. J. Mol. Sci. 2018, 19, 329. [Google Scholar] [CrossRef] [Green Version]
- Shi, A.; Heinayati, A.; Bao, D.; Liu, H.; Ding, X.; Tong, X.; Wang, L.; Wang, B.; Qin, H. Small molecule inhibitor of TGF-β signaling enables robust osteogenesis of autologous GMSCs to successfully repair minipig severe maxillofacial bone defects. Stem Cell Res. Ther. 2019. [Google Scholar] [CrossRef] [Green Version]
- Sun, W.; Wang, Z.; Xu, Q.; Sun, H.; Liu, X.; Yang, J.; Hong, R. The treatment of systematically transplanted gingival mesenchymal stem cells in periodontitis in mice. Exp. Ther. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, Z.; Song, W.; Sun, W.; Hong, R.; Pothukuchi, A.; Xu, Q. Systematically transplanted human gingiva-derived mesenchymal stem cells regulate lipid metabolism and inflammation in hyperlipidemic mice with periodontitis. Exp. Ther. Med. 2019. [Google Scholar] [CrossRef]
- Qiu, J.; Wang, X.; Zhou, H.; Zhang, C.; Wang, Y.; Huang, J.; Liu, M.; Yang, P.; Song, A. Enhancement of periodontal tissue regeneration by conditioned media from gingiva-derived or periodontal ligament-derived mesenchymal stem cells: A comparative study in rats. Stem Cell Res. Ther. 2020. [Google Scholar] [CrossRef]
- Zhang, Q.; Nguyen, P.D.; Shi, S.; Burrell, J.C.; Cullen, D.K.; Le, A.D. 3D bio-printed scaffold-free nerve constructs with human gingiva-derived mesenchymal stem cells promote rat facial nerve regeneration. Sci. Rep. 2018. [Google Scholar] [CrossRef] [Green Version]
- Mammana, S.; Gugliandolo, A.; Cavalli, E.; Diomede, F.; Iori, R.; Zappacosta, R.; Bramanti, P.; Conti, P.; Fontana, A.; Pizzicannella, J.; et al. Human gingival mesenchymal stem cells pretreated with vesicular moringin nanostructures as a new therapeutic approach in a mouse model of spinal cord injury. J. Tissue Eng. Regen. Med. 2019. [Google Scholar] [CrossRef] [Green Version]
- Rajan, T.S.; Diomede, F.; Bramanti, P.; Trubiani, O.; Mazzon, E. Conditioned medium from human gingival mesenchymal stem cells protects motor-neuron-like NSC-34 cells against scratch-injury-induced cell death. Int. J. Immunopathol. Pharmacol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhao, Y.; Ge, L. Therapeutic effects of human gingiva-derived mesenchymal stromal cells on murine contact hypersensitivity via prostaglandin E2-EP3 signaling. Stem Cell Res. Ther. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Su, W.; Lin, X.; Guo, Z.; Wang, J.; Zhang, Q.; Brand, D.; Ryffel, B.; Huang, J.; Liu, Z.; et al. Adoptive transfer of human gingiva-derived mesenchymal stem cells ameliorates collagen-induced arthritis via suppression of Th1 and Th17 cells and enhancement of regulatory T cell differentiation. Arthritis Rheum. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, Y.; Shi, S. Transplantation of gingiva-derived mesenchymal stem cells ameliorates collagen-induced arthritis. Arthritis Res. Ther. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Wu, W.; Gu, J.; Zhang, X.; Dang, J.; Wang, J.; Zheng, Y.; Huang, F.; Yuan, J.; Xue, Y.; et al. Human gingival tissue-derived MSC suppress osteoclastogenesis and bone erosion via CD39-adenosine signal pathway in autoimmune arthritis. EBioMedicine 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Chen, M.; Chen, W.; Gu, J.; Yuan, J.; Xue, Y.; Dang, J.; Su, W.; Wang, J.; Zadeh, H.H.; et al. Human gingiva-derived mesenchymal stem cells inhibit xeno-graft-versus-host disease via CD39-CD73-adenosine and IDO signals. Front. Immunol. 2017. [Google Scholar] [CrossRef] [Green Version]
- Ni, X.; Xia, Y.; Zhou, S.; Peng, H.; Wu, X.; Lu, H.; Wang, H.; Liu, R.; Blazar, B.R.; Gu, J.; et al. Reduction in murine acute GVHD severity by human gingival tissue-derived mesenchymal stem cells via the CD39 pathways. Cell Death Dis. 2019. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhou, L.; Dang, J.; Zhang, X.; Wang, J.; Chen, Y.; Liang, J.; Li, D.; Ma, J.; Yuan, J.; et al. Human Gingiva-Derived Mesenchymal Stem Cells Ameliorate Streptozoticin-induced T1DM in mice via Suppression of T effector cells and Up-regulating Treg Subsets. Sci. Rep. 2017. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020. [Google Scholar] [CrossRef]
- Lane, R.E.; Korbie, D.; Anderson, W.; Vaidyanathan, R.; Trau, M. Analysis of exosome purification methods using a model liposome system and tunable-resistive pulse sensing. Sci. Rep. 2015. [Google Scholar] [CrossRef] [Green Version]
- Tschuschke, M.; Kocherova, I.; Bryja, A.; Mozdziak, P.; Angelova Volponi, A.; Janowicz, K.; Sibiak, R.; Piotrowska-Kempisty, H.; Iżycki, D.; Bukowska, D.; et al. Inclusion Biogenesis, Methods of Isolation and Clinical Application of Human Cellular Exosomes. J. Clin. Med. 2020, 9, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, F.; Zhang, D.; Fang, T.; Lu, C.; Wang, B.; Ding, X.; Wei, S.; Zhang, Y.; Pi, W.; Xu, H.; et al. Exosomes from human gingiva-derived mesenchymal stem cells combined with biodegradable chitin conduits promote rat sciatic nerve regeneration. Stem Cells Int. 2019. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Qian, Z.; Liu, D.; Sun, J.; Wang, X.; Liu, H.; Xu, J.; Guo, X. GMSC-derived exosomes combined with a chitosan/silk hydrogel sponge accelerates wound healing in a diabetic rat skin defect model. Front. Physiol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shi, S.; Xu, Q.; Zhang, Q.; Shanti, R.M.; Le, A.D. SIS-ECM Laden with GMSC-Derived Exosomes Promote Taste Bud Regeneration. J. Dent. Res. 2019. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolation Method | Author | Publication Date | Composition of Culture Medium | Enzymes Used for Digestion | Time of Digestion |
|---|---|---|---|---|---|
| Explant method | El-Bialy et al. [24] | 2014 | DMEM, 10% FBS, 100 U/mL penicillin, 100 µg/mL streptomycin | – | – |
| Fawzy El-Sayed et al. [23] | 2015 | α-MEM, 15% FCS, 400 mM/mL L-glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin, 1% amphotericin | – | – | |
| Fournier et al. [25] | 2010 | DMEM, 20% FCS, 100 µg/mL penicillin, 100 µg/mL streptomycin, 2 ng/mL amphotericin | – | – | |
| Mitrano et al. [22] | 2010 | α-MEM, 10% FBS, 1% penicillin, streptomycin, amphotericin | – | – | |
| Enzymatic method | Gao et al. [28] | 2014 | α-MEM, 10% FBS | 4 mg/mL collagenase IV | 2 h at 37 °C |
| Ge et al. [31] | 2012 | α-MEM, 20% FCS, 2 mM L-glutamine, 100 µM L-ascorbate-2-phosphate, 1 mM sodium pyruvate, 50 U/mL penicillin, 50 µg/mL streptomycin, 2.5 µg/mL amphotericin | 3 mg/mL collagenase I, 4 mg/mL Dispase II | 50 min at 37 °C | |
| Jin et al. [32] | 2015 | α-MEM, 15% FBS, 100 U/mL penicillin, 100 µg/mL streptomycin, 200 mM L-glutamine, 10 mM ascorbic acid 2-phosphate | 2 mg/mL collagenase IV, 1 mg/mL Dispase | 30 at 37 °C (fraction discarded) 90 min at 37 °C (fraction seeded) | |
| Tang et al. [29] | 2011 | DMEM, 10% FBS, 0.292 mg/mL glutamine, 100 U/mL penicillin, 100 µg/mL streptomycin | 0.66 mg/mL collagenase I | 50 min at 37 °C | |
| Tomar et al. [30] | 2010 | α-MEM, 10% FCS | 0.1% collagenase, 0.2% Dispase | 15 min at 37 °C (fraction discarded) 5 min at 37 °C 10 min at 37 °C 15 min at 37 °C (fractions pooled and seeded) | |
| Zhang et al. [17] | 2009 | α-MEM, 10% FBS, 100 U/mL penicillin, 100 µg/mL streptomycin, 2 mM L-glutamine, 100 mM nonessential amino acids | 4 mg/mL collagenase IV | 2 h at 37 °C |
| Author | Publication Date | Type of Cancer | Employed Method | Obtained Results |
|---|---|---|---|---|
| Coccè et al. [69] | 2017 | TSCC line (SCC154) | Drug loading of GMSCs with paclitaxel, doxorubicin, and gemcitabine | Growth inhibition of cancer cells in response to drug release by GMSCs |
| Du et al. [68] | 2019 | TSCC line (CAL27); TSCC xenograft model in BALB/c nude mice | Transfection of GMSCs with IFNβ | Growth inhibition and apoptosis of cancer cells in vitro and inhibition of tumor cell proliferation in vivo |
| Ji et al. [62] | 2016 | TSCC lines (CAL27, WSU-HN6) | Direct and Transwell coculture of GMSCs and cancer cells; simultaneous subcutaneous injection of cancer cells with GMSCs | Growth inhibition of cancer cells in vitro and in vivo |
| Xia et al. [63] | 2014 | TSCC line (TCA8113, CAL27) | Transfection of GMSCs with TRAIL | Tumor cell necrosis and apoptosis in vitro and in vivo |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefańska, K.; Mehr, K.; Wieczorkiewicz, M.; Kulus, M.; Angelova Volponi, A.; Shibli, J.A.; Mozdziak, P.; Skowroński, M.T.; Antosik, P.; Jaśkowski, J.M.; et al. Stemness Potency of Human Gingival Cells—Application in Anticancer Therapies and Clinical Trials. Cells 2020, 9, 1916. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9081916
Stefańska K, Mehr K, Wieczorkiewicz M, Kulus M, Angelova Volponi A, Shibli JA, Mozdziak P, Skowroński MT, Antosik P, Jaśkowski JM, et al. Stemness Potency of Human Gingival Cells—Application in Anticancer Therapies and Clinical Trials. Cells. 2020; 9(8):1916. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9081916
Chicago/Turabian StyleStefańska, Katarzyna, Katarzyna Mehr, Maria Wieczorkiewicz, Magdalena Kulus, Ana Angelova Volponi, Jamil A. Shibli, Paul Mozdziak, Mariusz T. Skowroński, Paweł Antosik, Jędrzej M. Jaśkowski, and et al. 2020. "Stemness Potency of Human Gingival Cells—Application in Anticancer Therapies and Clinical Trials" Cells 9, no. 8: 1916. https://0-doi-org.brum.beds.ac.uk/10.3390/cells9081916