
Isothermal Amplification and Lateral Flow Nucleic Acid Test for the Detection of Shiga Toxin-Producing Bacteria for Food Monitoring


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Bacterial Lysis and DNA Collection in Media
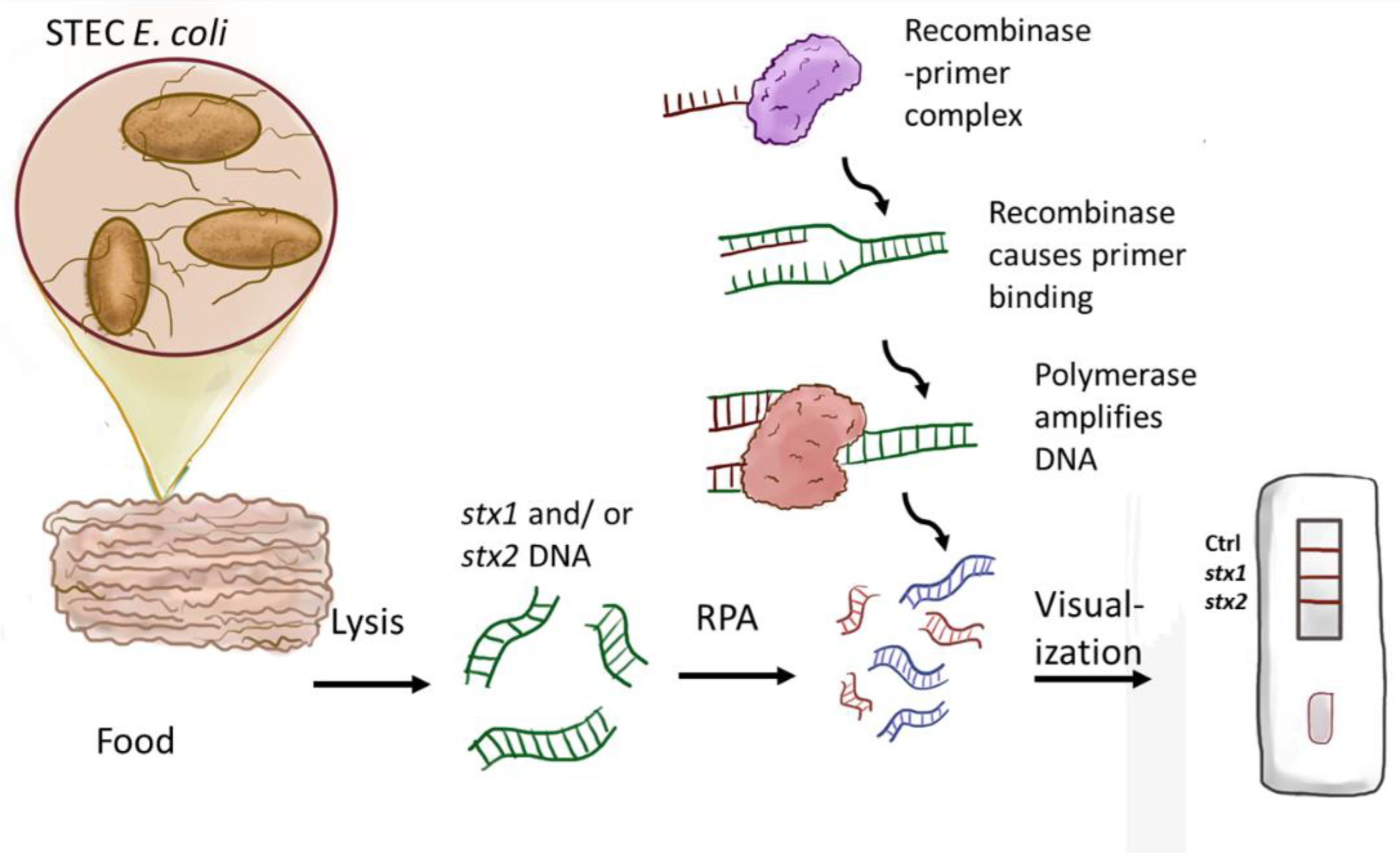
2.3. Shiga Toxin Gene Amplification Using RPA
2.4. Lateral Flow Assay
2.5. Solid Food Sample Handling and Spiked Sample Studies
3. Results
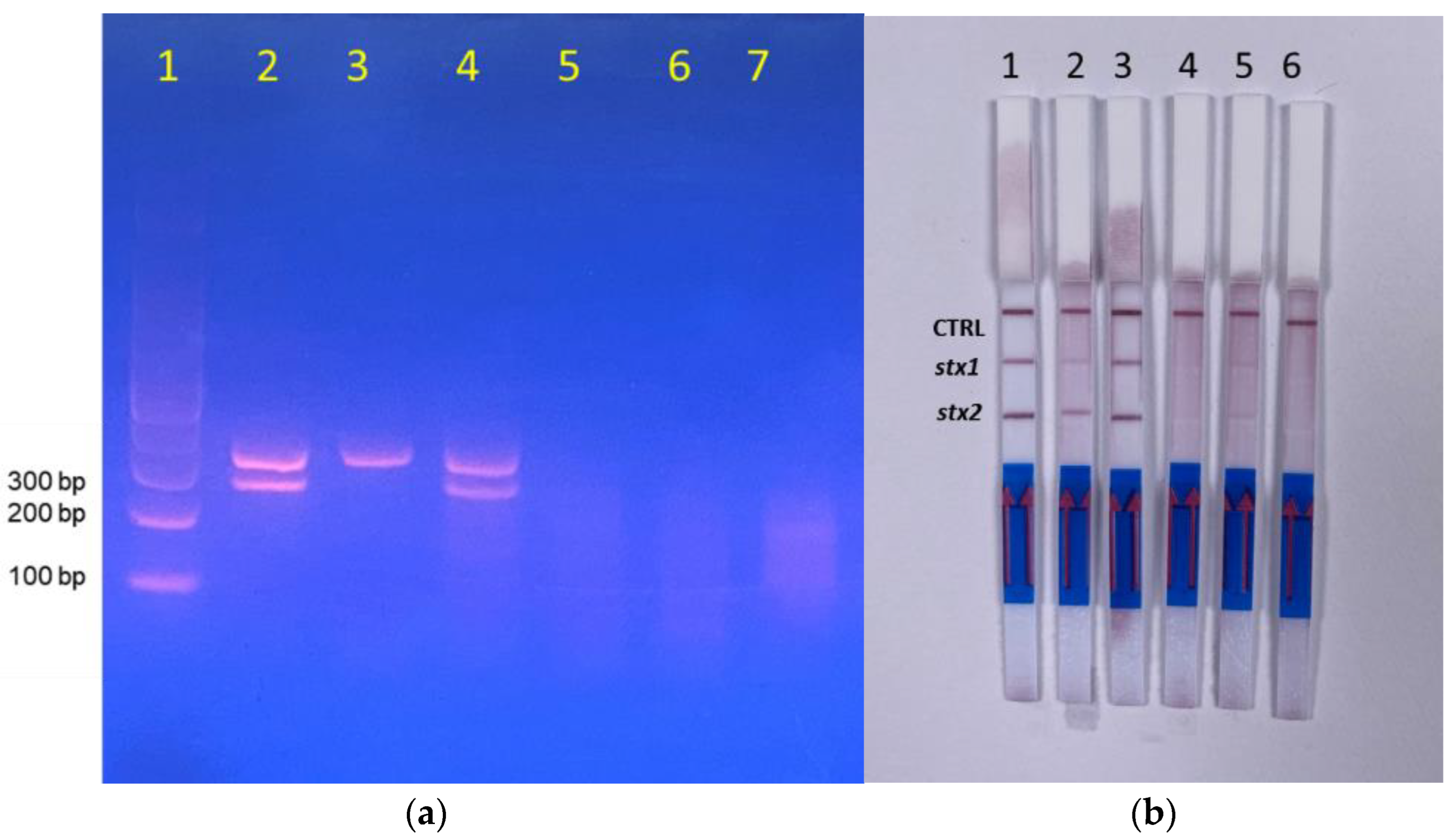
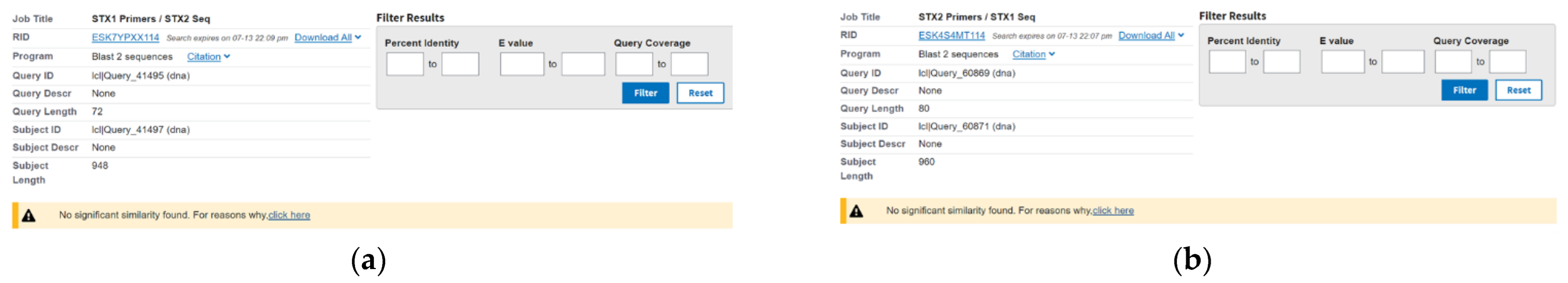
3.1. RPA Optimization and Primer Selection
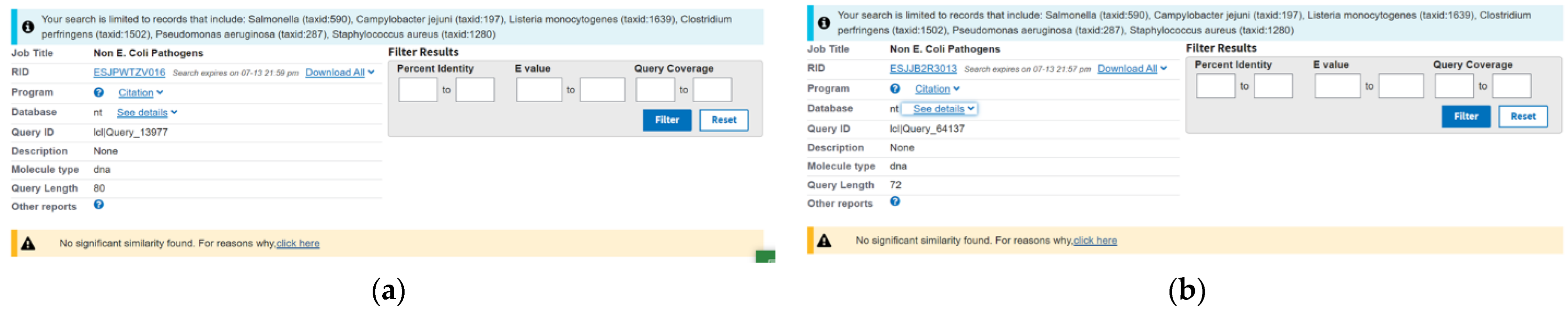
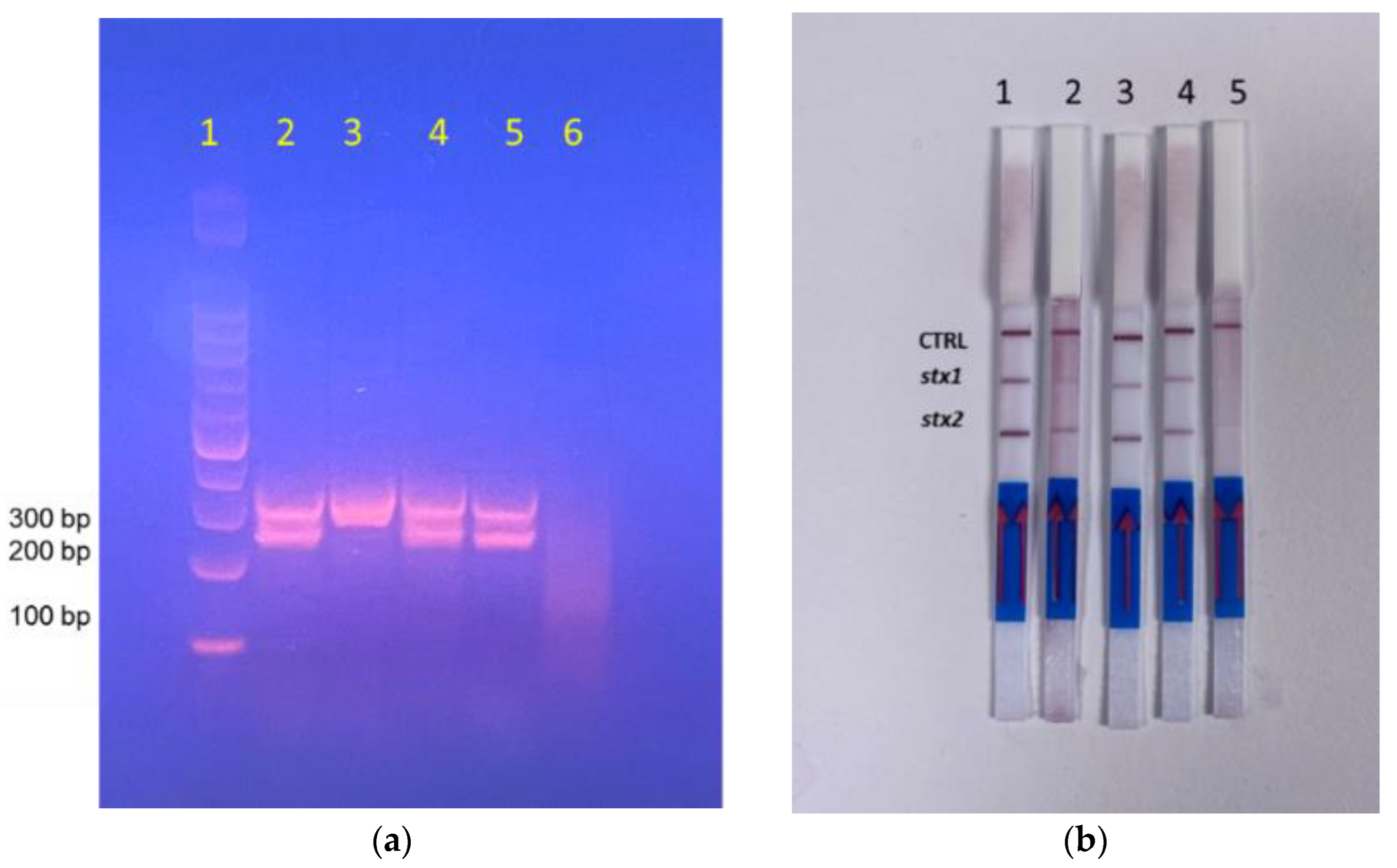
3.2. Specificity
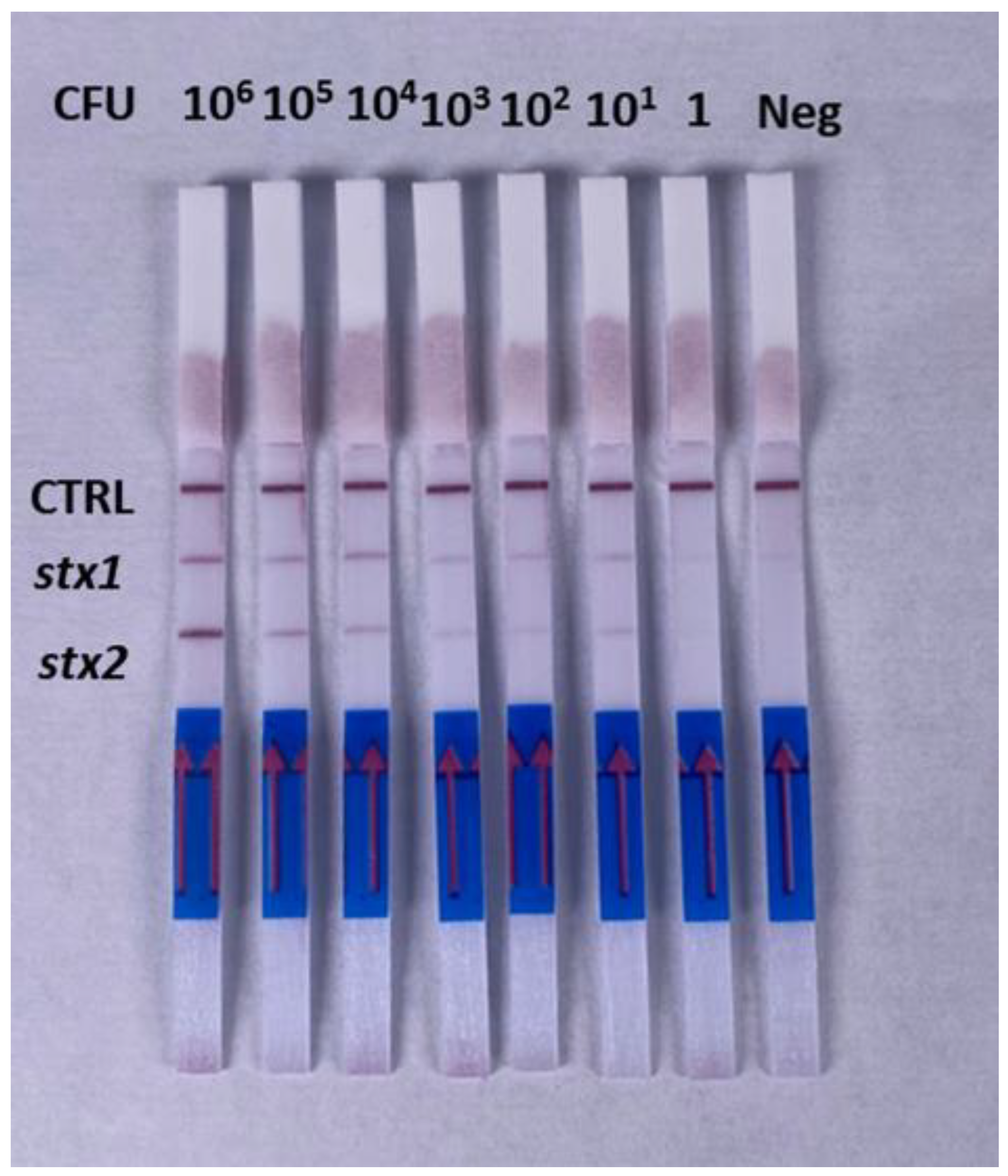
3.3. Evaluation of Sensitivity in Media
3.4. Evaluation of Sensitivity in Spiked Food Samples
3.5. Evaluation of a Mixed Sample
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Scharff, R.L. Economic Burden from Health Losses due to Foodborne Illness in the United States. J. Food Prot. 2012, 75, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Bartsch, S.M.; Asti, L.; Nyathi, S.; Spiker, M.L.; Lee, B.Y. Estimated Cost to a Restaurant of a Foodborne Illness Outbreak. Public Health Rep. 2018, 133, 274–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrucci, S.; Costa, C.; Broyles, D.; Dikici, E.; Daunert, S.; Deo, S. On-site detection of food and waterborne bacteria—Current technologies, challenges, and future directions. Trends Food Sci. Technol. 2021, 115, 409–421. [Google Scholar] [CrossRef] [PubMed]
- Farrokh, C.; Jordan, K.; Auvray, F.; Glass, K.; Oppegaard, H.; Raynaud, S.; Thevenot, D.; Condron, R.; de Reu, K.; Govaris, A.; et al. Review of Shiga toxin-producing Escherichia coli (STEC) and their significance in dairy production. Int. J. Food Microbiol. 2013, 162, 190–212. [Google Scholar] [CrossRef] [PubMed]
- Scheutz, F.; Teel, L.D.; Beutin, L.; Piérard, D.; Buvens, G.; Karch, H.; Mellmann, A.; Caprioli, A.; Tozzoli, R.; Morabito, S.; et al. Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 2012, 50, 2951–2963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melton-Celsa, A.R. Shiga Toxin (Stx) Classification, Structure, and Function. Microbiol. Spectr. 2014, 2, 2–4. [Google Scholar] [CrossRef] [Green Version]
- Mauro, S.A.; Koudelka, G.B. Shiga toxin: Expression, distribution, and its role in the environment. Toxins 2011, 3, 608–625. [Google Scholar] [CrossRef]
- Los, J.; Los, M.; Wegrzyn, A.; Wegrzyn, G. Altruism of Shiga toxin-producing Escherichia coli: Recent hypothesis versus experimental results. Front. Cell. Infect. Microbiol. 2013, 2, 166. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rubio, L.; Haarmann, N.; Schwidder, M.; Muniesa, M.; Schmidt, H. Bacteriophages of Shiga Toxin-Producing Escherichia coli and Their Contribution to Pathogenicity. Pathogens 2021, 10, 404. [Google Scholar] [CrossRef]
- Mcgannon, C.M.; Fuller, C.A.; Weiss, A.A. Different classes of antibiotics differentially influence Shiga toxin production. Antimicrob. Agents Chemother. 2010, 54, 3790–3798. [Google Scholar] [CrossRef] [Green Version]
- Chan, Y.S.; Ng, T.B. Shiga toxins: From structure and mechanism to applications. Appl. Microbiol. Biotechnol. 2016, 100, 1597–1610. [Google Scholar] [CrossRef] [PubMed]
- Tahamtan, Y.; Hayati, M.; Namavari, M. Prevalence and distribution of the Stx, Stx genes in Shiga toxin producing E. coli (STEC) isolates from cattle. Iran. J. Microbiol. 2010, 2, 8–13. [Google Scholar] [PubMed]
- Zhang, P.; Essendoubi, S.; Keenliside, J.; Reuter, T.; Stanford, K.; King, R.; Lu, P.; Yang, X. Genomic analysis of Shiga toxin-producing Escherichia coli O157:H7 from cattle and pork-production related environments. NPJ Sci. Food 2021, 5, 21. [Google Scholar] [CrossRef] [PubMed]
- Valilis, E.; Ramsey, A.; Sidiq, S.; Dupont, H.L. Non-O157 Shiga toxin-producing Escherichia coli—A poorly appreciated enteric pathogen: Systematic review. Int. J. Infect. Dis. 2018, 76, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Deisingh, A.K.; Thompson, M. Strategies for the detection of Escherichia coli O157:H7 in foods. J. Appl. Microbiol. 2004, 96, 419–429. [Google Scholar] [CrossRef] [Green Version]
- Han, X.; Liu, Y.; Yin, J.; Yue, M.; Mu, Y. Microfluidic devices for multiplexed detection of foodborne pathogens. Food Res. Int. 2021, 143, 110246. [Google Scholar] [CrossRef]
- Sohrabi, H.; Majidi, M.R.; Khaki, P.; Jahanban-Esfahlan, A.; De La Guardia, M.; Mokhtarzadeh, A. State of the art: Lateral flow assays toward the point-of-care foodborne pathogenic bacteria detection in food samples. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1868–1912. [Google Scholar] [CrossRef]
- Shin, J.H.; Hong, J.; Go, H.; Park, J.; Kong, M.; Ryu, S.; Kim, K.-P.; Roh, E.; Park, J.-K. Multiplexed detection of foodborne pathogens from contaminated lettuces using a handheld multistep lateral flow assay device. J. Agric. Food Chem. 2018, 66, 290–297. [Google Scholar] [CrossRef]
- Lin, L.; Zheng, Y.; Huang, H.; Zhuang, F.; Chen, H.; Zha, G.; Yang, P.; Wang, Z.; Kong, M.; Wei, H.; et al. A visual method to detect meat adulteration by recombinase polymerase amplification combined with lateral flow dipstick. Food Chem. 2021, 354, 129526. [Google Scholar] [CrossRef]
- Lalremruata, A.; Nguyen, T.T.; Mccall, M.B.B.; Mombo-Ngoma, G.; Agnandji, S.T.; Adegnika, A.A.; Lell, B.; Ramharter, M.; Hoffman, S.L.; Kremsner, P.G.; et al. Recombinase Polymerase Amplification and Lateral Flow Assay for Ultrasensitive Detection of Low-Density Plasmodium Falciparum Infection from Controlled Human Malaria Infection Studies and Naturally Acquired Infections. J. Clin. Microbiol. 2020, 58, e01879-19. [Google Scholar] [CrossRef]
- Petrucci, S.; Costa, C.; Broyles, D.; Kaur, A.; Dikici, E.; Daunert, S.; Deo, S.K. Monitoring Pathogenic Viable E. coli O157:H7 in Food Matrices Based on the Detection of RNA Using Isothermal Amplification and a Paper-Based Platform. Anal. Chem. 2022, 94, 2485–2492. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Macdonald, J.; von Stetten, F. Review: A comprehensive summary of a decade development of the recombinase polymerase amplification. Analyst 2019, 144, 31–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piepenburg, O.; Williams, C.H.; Stemple, D.L.; Armes, N.A. DNA Detection Using Recombination Proteins. PLoS Biol. 2006, 4, e204. [Google Scholar] [CrossRef]
- Lobato, I.M.; O’Sullivan, C.K. Recombinase polymerase amplification: Basics, applications and recent advances. Trends Anal. Chem. TRAC 2018, 98, 19–35. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Won, J.; Choi, B.Y.; Lee, C.J. Optimization of primer sets and detection protocols for SARS-CoV-2 of coronavirus disease 2019 (COVID-19) using PCR and real-time PCR. Exp. Mol. Med. 2020, 52, 963–977. [Google Scholar] [CrossRef] [PubMed]
- Barra, G.B.; Rita, T.H.S.; Mesquita, P.G.; Jacomo, R.H.; Nery, L.F.A. Analytical Sensitivity and Specificity of Two RT-qPCR Protocols for SARS-CoV-2 Detection Performed in an Automated Workflow. Genes 2020, 11, 1183. [Google Scholar] [CrossRef] [PubMed]
- Etcheverría, A.I.; Padola, N.L. Shiga toxin-producing Escherichia coli: Factors involved in virulence and cattle colonization. Virulence 2013, 4, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Edson, D.C.; Empson, S.; Massey, L.D. Pathogen detection in food microbiology laboratories: An analysis of qualitative proficiency test data, 1999–2007. J. Food Saf. 2009, 29, 521–530. [Google Scholar] [CrossRef]
- Massih, M.A.; Planchon, V.; Polet, M.; Dierick, K.; Mahillon, J. Analytical performances of food microbiology laboratories—Critical analysis of 7 years of proficiency testing results. J. Appl. Microbiol. 2016, 120, 346–354. [Google Scholar] [CrossRef] [Green Version]
- Rahal, E.; Kazzi, N.; Nassar, F.; Matar, G. Escherichia coli O157:H7—Clinical aspects and novel treatment approaches. Front. Cell. Infect. Microbiol. 2012, 2, 138. [Google Scholar] [CrossRef] [Green Version]
- Gyles, C.L. Shiga toxin-producing Escherichia coli: An overview1. J. Anim. Sci. 2007, 85, e45–e62. [Google Scholar] [CrossRef]
- Momtaz, H.; Jamshidi, A. Shiga toxin-producing Escherichia coli isolated from chicken meat in Iran: Serogroups, virulence factors, and antimicrobial resistance properties. Poult. Sci. 2013, 92, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Zarei, O.; Shokoohizadeh, L.; Hossainpour, H.; Alikhani, M.Y. The prevalence of Shiga toxin-producing Escherichia coli and enteropathogenic Escherichia coli isolated from raw chicken meat samples. Int. J. Microbiol. 2021, 2021, 3333240. [Google Scholar] [CrossRef]
- Doane, C.A.; Pangloli, P.; Richards, H.A.; Mount, J.R.; Golden, D.A.; Draughon, F.A. Occurrence of Escherichia coli O157:H7 in diverse farm environments. J. Food Prot. 2007, 70, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, G.L.; Hollingsworth, J.; Morris, J.G., Jr. Emerging foodborne pathogens: Escherichia coli O157:H7 as a model of entry of a new pathogen into the food supply of the developed world. Epidemiol. Rev. 1996, 18, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Kuiper, H.A.; Paoletti, C. Food And Feed Safety Assessment: The Importance of Proper Sampling. J. AOAC Int. 2019, 98, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Faulds, N.; Evans, K.; Williams, J.; Leonte, A.-M.; Crabtree, D.; Church, K.; Leak, D.; Sohier, D.; Palomäki, J.-P.; Heikkinen, P.; et al. Validation of the Thermo Scientific Suretect™ Escherichia coli O157:H7 And STEC Screening PCR Assay and Suretect™ Escherichia coli STEC Identification PCR Assay for the Detection of Escherichia coli O157:H7 and the Escherichia Coli STEC Serotypes (O26, O45, O103, O111, O121, O145) From Fresh Raw Spinach, Fresh Baby Leaves, Fresh Cut Tomatoes, Frozen Raw Beef, Raw Beef Trim, and Beef Carcass Sponges: AOAC Performance Tested MethodSM 012102. J. AOAC Int. 2022, 105, 521–548. [Google Scholar] [CrossRef]
- Berenger, B.M.; Chui, L.; Ferrato, C.; Lloyd, T.; Li, V.; Pillai, D.R. Performance of four commercial real-time PCR assays for the detection of bacterial enteric pathogens in clinical samples. Int. J. Infect. Dis. 2022, 114, 195–201. [Google Scholar] [CrossRef]
- Parma, Y.R.; Chacana, P.A.; Lucchesi, P.M.A.; Rogé, A.; Granobles Velandia, C.V.; Krüger, A.; Parma, A.E.; Fernández-Miyakawa, M.E. Detection of Shiga toxin-producing Escherichia coli by sandwich enzyme-linked immunosorbent assay using chicken egg yolk IgY antibodies. Front. Cell. Infect. Microbiol. 2012, 2, 84. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Bielaszewska, M.; Pulz, M.; Becker, K.; Friedrich, A.W.; Karch, H.; Kuczius, T. New Immuno-PCR Assay for Detection of Low Concentrations of Shiga Toxin 2 and Its Variants. J. Clin. Microbiol. 2008, 46, 1292–1297. [Google Scholar] [CrossRef] [Green Version]
- Reischl, U.; Youssef, M.T.; Kilwinski, J.; Lehn, N.; Zhang, W.L.; Karch, H.; Strockbine, N.A. Real-time fluorescence PCR assays for detection and characterization of Shiga toxin, intimin, and enterohemolysin genes from Shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 2002, 40, 2555–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murinda, S.E.; Ibekwe, A.M.; Zulkaffly, S.; Cruz, A.; Park, S.; Razak, N.; Paudzai, F.M.; Samad, A.L.; Baquir, K.; Muthaiyah, K.; et al. Real-time isothermal detection of Shiga toxin-producing Escherichia coli using recombinase polymerase amplification. Foodborne Pathog. Dis. 2014, 11, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Verstraete, K.; Van Coillie, E.; Werbrouck, H.; Van Weyenberg, S.; Herman, L.; Del-Favero, J.; De Rijk, P.; De Zutter, L.; Joris, M.-A.; Heyndrickx, M.; et al. A qPCR assay to detect and quantify Shiga toxin-producing E. coli (STEC) in cattle and on farms: A potential predictive tool for STEC culture-positive farms. Toxins 2014, 6, 1201–1221. [Google Scholar] [CrossRef] [PubMed]
- Jinneman, K.C.; Yoshitomi, K.J.; Weagant, S.D. Multiplex real-time PCR method to identify Shiga toxin genes stx1 and stx2 and Escherichia coli O157:H7/H− serotype. Appl. Environ. Microbiol. 2003, 69, 6327–6333. [Google Scholar] [CrossRef] [Green Version]
- Shan, S.; Huang, Y.; Huang, Z.; Long, Z.; Liu, C.; Zhao, X.; Xing, K.; Xiao, X.; Liu, J.; Huang, Y.; et al. Detection of stx1 and stx2 and subtyping of Shiga toxin-producing Escherichia coli using asymmetric PCR combined with lateral flow immunoassay. Food Control 2021, 126, 108051. [Google Scholar] [CrossRef]
- Singh, P.; Liu, Y.; Bosilevac, J.M.; Mustapha, A. Detection of Shiga toxin-producing Escherichia coli, stx1, stx2 and Salmonella by two high resolution melt curve multiplex real-time PCR. Food Control 2019, 96, 251–259. [Google Scholar] [CrossRef]
- Capobianco, J.A.; Clark, M.; Cariou, A.; Leveau, A.; Pierre, S.; Fratamico, P.; Strobaugh, T.P.; Armstrong, C.M. Detection of Shiga toxin-producing Escherichia coli (STEC) in beef products using droplet digital PCR. Int. J. Food Microbiol. 2020, 319, 108499. [Google Scholar] [CrossRef]
- De Boer, E.; Beumer, R.R. Methodology for detection and typing of foodborne microorganisms. Int. J. Food Microbiol. 1999, 50, 119–130. [Google Scholar] [CrossRef]
- Ching, K.H.; He, X.; Stanker, L.H.; Lin, A.V.; Mcgarvey, J.A.; Hnasko, R. Detection of Shiga toxins by lateral flow assay. Toxins 2015, 7, 1163–1173. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Katani, R.; Li, L.; Hegde, N.; Roberts, E.L.; Kapur, V.; Debroy, C. Rapid Detection of Escherichia coli o157 and Shiga Toxins by Lateral Flow Immunoassays. Toxins 2016, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Boone, J.T.; Campbell, D.E.; Dandro, A.S.; Chen, L.; Herbein, J.F. A Rapid Immunoassay for Detection of Shiga Toxin-Producing Escherichia coli Directly from Human Fecal Samples and Its Performance in Detection of Toxin Subtypes. J. Clin. Microbiol. 2016, 54, 3056–3063. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| stx1 F | 5′-FAM-TTTTATCGCTTTGCTGATTTTTCACATGTT-3′ |
| stx1 R | 5′-DIG-CAAACCGTAACATCGCTCTTGCCACAGACT-3′ |
| stx2 F | 5′-FAM-CAGAGATATCGACCCCTCTTGAACATATAT-3′ |
| stx2 R | 5′-Biotin-GTATAACTGCTGTCCGTTGTCATGGAAACC-3′ |
| Methodology | Assay Run Time | Limit of Detection | Reference |
|---|---|---|---|
| Immuno-PCR | Not stated | 10 pg/mL purified toxin | [40] |
| Real-time PCR | ~1 h | 5 × 103 CFU/mL | [41] |
| Real-time RPA | 5–10 min | ~5–50 CFU/mL | [42] |
| qPCR | Not stated | <2.7–3.7 log copies g−1 feces | [43] |
| Multiplex Real-Time PCR | Not stated | 6 CFU/mL | [44] |
| Asymmetric PCR and LFA | Not stated | Not stated but 10 × PCR | [45] |
| Multiplex melting curve PCR | >6 h (because of enrichment step) | 1 CFU/mL with enrichment of 6 h | [46] |
| Droplet digital PCR | Not stated | Not stated | [47] |
| Culture-Based | 1–3 days | 1 cell | [48] |
| LFA-immunoassay | <10 min | 0.1 ng/mL toxin | [49] |
| LFA-immunoassay | ~3 h | 105 | [50] |
| Immunoassay | <30 min | 3.3 × 101 CFU/g to 1.3 × 103 CFU/g when induced with antibiotic | [51] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrucci, S.; Dikici, E.; Daunert, S.; Deo, S.K. Isothermal Amplification and Lateral Flow Nucleic Acid Test for the Detection of Shiga Toxin-Producing Bacteria for Food Monitoring. Chemosensors 2022, 10, 210. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060210
Petrucci S, Dikici E, Daunert S, Deo SK. Isothermal Amplification and Lateral Flow Nucleic Acid Test for the Detection of Shiga Toxin-Producing Bacteria for Food Monitoring. Chemosensors. 2022; 10(6):210. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060210
Chicago/Turabian StylePetrucci, Sabrina, Emre Dikici, Sylvia Daunert, and Sapna K. Deo. 2022. "Isothermal Amplification and Lateral Flow Nucleic Acid Test for the Detection of Shiga Toxin-Producing Bacteria for Food Monitoring" Chemosensors 10, no. 6: 210. https://0-doi-org.brum.beds.ac.uk/10.3390/chemosensors10060210