Comment Concerning the Effects of Light Intensity on Melatonin Suppression in the Review “Light Modulation of Human Clocks, Wake, and Sleep” by A. Prayag et al.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:ANNEX
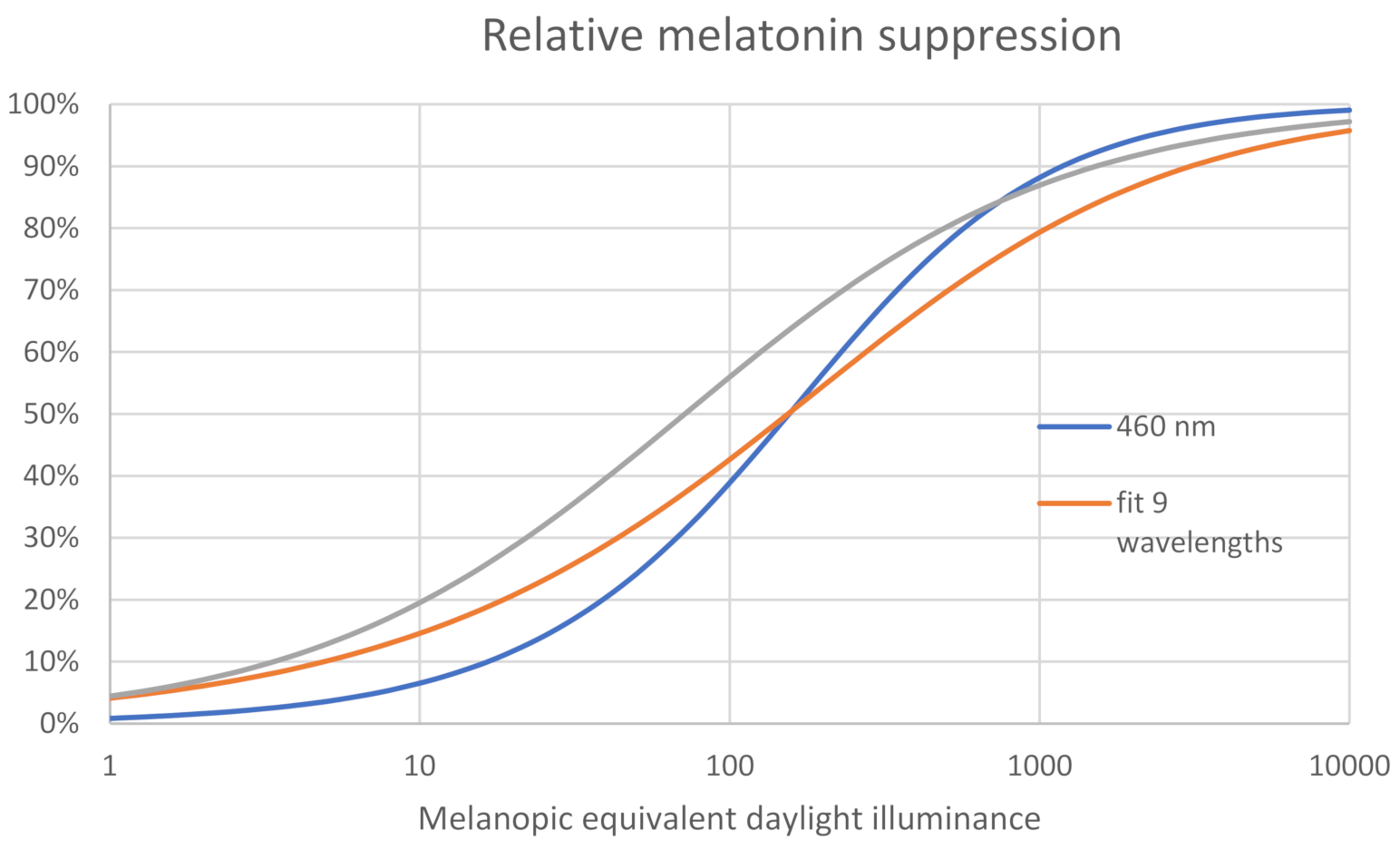
Dose–Response Curves for Circadian Phase Shift and Melatonin Suppression vs. MEDI
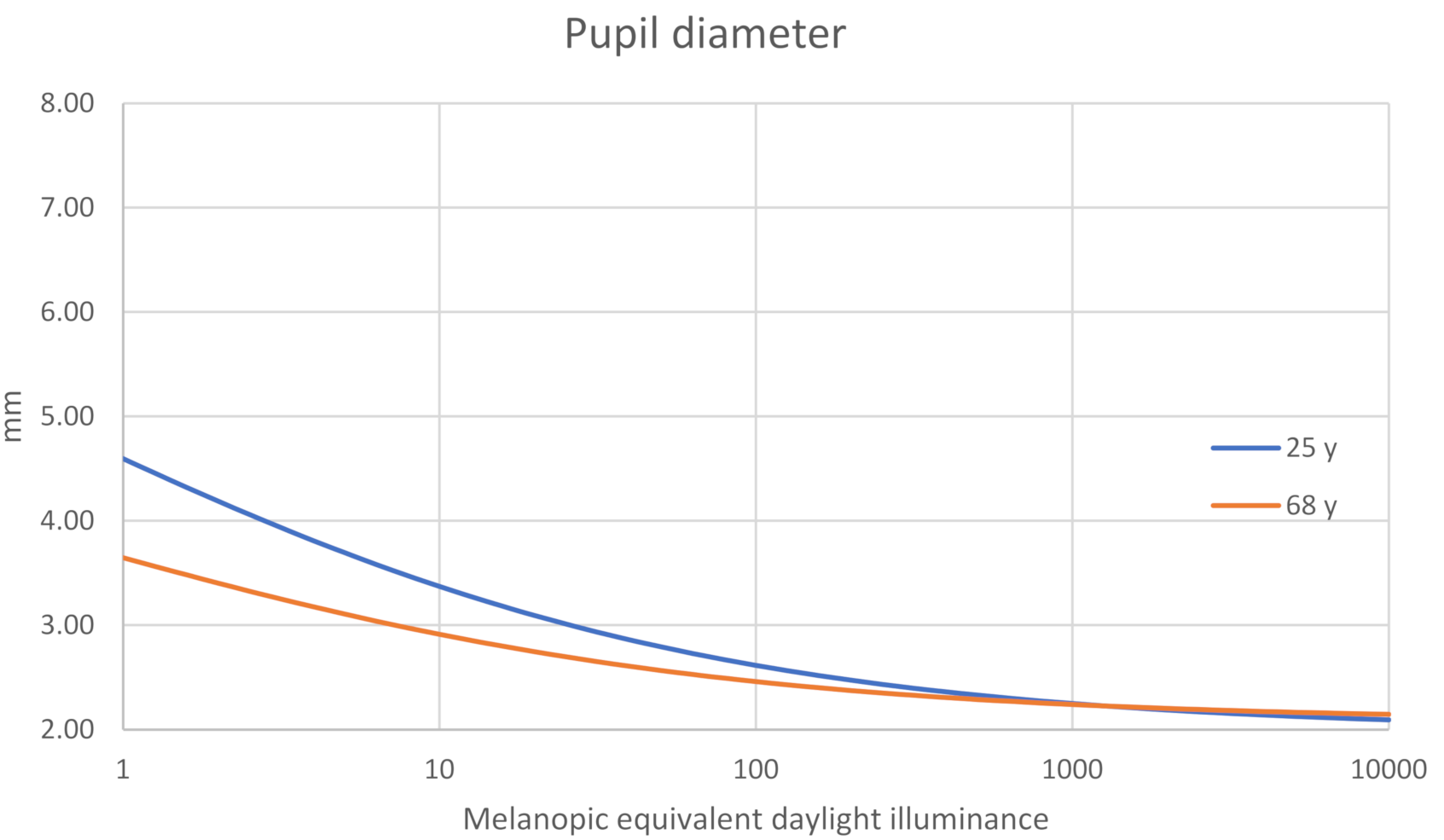
Pupil Size vs. MEDI
Single Wavelength Dilated Pupil Irradiance Converted to MEDI for Non-Dilated Pupil
About the Cone Contribution to the Spectral Response
Spectral Response of Circadian Phase Shift
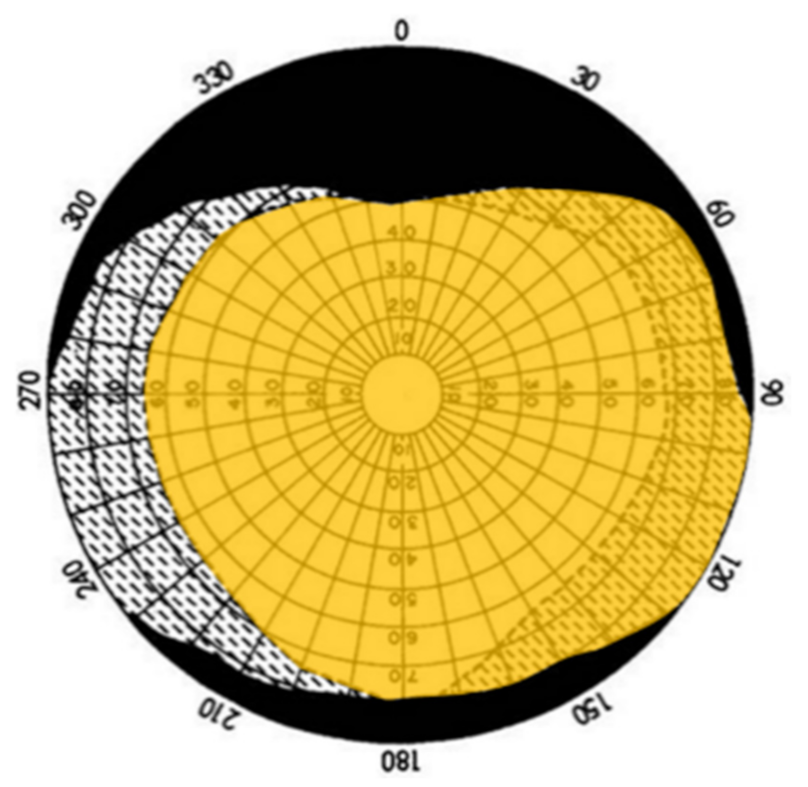
Spectral Response of Pupil Constriction
Inter-Individual Pupil Size Differences
Some Observations
- Spectral summation is not sufficiently statistical proven, but seems sufficiently appropriate.
- Typical viewing conditions differ from a ganzfeld and, thus, there could be a very small L+M-S contribution, but this is negligible unless the lighting itself fluctuates rapidly (peak at 0.2 Hz).
- The formula for pupil size with age in the illumination range of interest is an extrapolation since experimental data is lacking.
- Inter-individual pupil area differences are important (the 90 percentile needs 56% more illumination than the average).
- Many papers are lacking necessary data (such as average pupil area, illumination spectra, etc.).
- A logistic function might not be the physiological correct fit.
- Individual responses in typical living environments with other light spectra could deviate substantially from what the curves predict.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Prayag, A.S.; Münch, M.; Aeschbach, D.; Chellappa, S.L.; Gronfier, C. Light Modulation of Human Clocks, Wake, and Sleep. Clocks Sleep 2019, 1, 193–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeitzer, J.M.; Khalsa, S.B.S.; Boivin, D.B.; Duffy, J.F.; Shanahan, T.L.; Kronauer, R.E.; Czeisler, C.A. Temporal dynamics of late-night photic stimulation of the human circadian timing system. Am. J. Physiol. Integr. Comp. Physiol. 2005, 289, R839–R844. [Google Scholar] [CrossRef] [Green Version]
- Cajochen, C.; Zeitzer, J.M.; Czeisler, C.A.; Dijk, D.J. Dose-response relationship for light intensity and ocular and electroencephalographic correlates of human alertness. Behav. Brain Res. 2000, 115, 75–83. [Google Scholar] [CrossRef]
- Souman, J.L.; Tinga, A.M.; Pas, S.F.T.; Van Ee, R.; Vlaskamp, B.N. Acute alerting effects of light: A systematic literature review. Behav. Brain Res. 2018, 337, 228–239. [Google Scholar] [CrossRef] [PubMed]
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.M.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action Spectrum for Melatonin Regulation in Humans: Evidence for a Novel Circadian Photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brainard, G.C.; Sliney, D.; Hanifin, J.P.; Glickman, G.; Byrne, B.; Greeson, J.M.; Jasser, S.; Gerner, E.; Rollag, M.D. Sensitivity of the Human Circadian System to Short-Wavelength (420-nm) Light. J. Biol. Rhythm. 2008, 23, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Prayag, A.S.; Najjar, R.P.; Gronfier, C. Melatonin suppression is exquisitely sensitive to light and primarily driven by melanopsin in humans. J. Pineal Res. 2019, 66, e12562. [Google Scholar] [CrossRef]
- Van de Werken, M.; Gimenez, M.C.; de Vries, B.; Beersma, D.G.M.; Gordijn, M.C.M. Short-wavelength attenuated polychromatic white light during work at night: Limited melatonin suppression without substantial decline of alertness. Chronobiol. Int. 2013, 30, 843–854. [Google Scholar] [CrossRef] [Green Version]
- Souman, J.L.; Borra, T.; De Goijer, I.; Schlangen, L.J.M.; Vlaskamp, B.N.S.; Lucassen, M.P. Spectral Tuning of White Light Allows for Strong Reduction in Melatonin Suppression without Changing Illumination Level or Color Temperature. J. Biol. Rhythm. 2018, 33, 420–431. [Google Scholar] [CrossRef]
- Mark, R.N.; Plitnick, R.B.S.; Figueiro, M.G. Effect of White Light Devoid of “Cyan” Spectrum Radiation on Nighttime Melatonin Suppression Over a 1-h Exposure Duration. J. Biol. Rhythm. 2019, 34, 195–204. [Google Scholar]
- CIE S 026/E:2018. CIE System for Metrology of Optical Radiation for ipRGC-Influenced Responses to Light; CIE: Vienna, Austria, 2018. [Google Scholar]
- McDougal, D.H.; Gamlin, P.D. The influence of intrinsically-photosensitive retinal ganglion cells on the spectral sensitivity and response dynamics of the human pupillary light reflex. Vis. Res. 2010, 50, 72–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gooley, J.J.; Mien, I.H.; Hilarie, M.A.S.; Yeo, S.-C.; Chua, E.C.-P.; van Reen, E.; Hanley, C.J.; Hull, J.T.; Czeisler, C.A.; Lockley, S.W. Melanopsin and rod-cone photoreceptors play different roles in mediating pupillary light responses during exposure to continuous light in humans. J. Neurosci. 2012, 32, 14242–14253. [Google Scholar] [CrossRef] [PubMed]
- Barrionuevo, P.A.; Nicandro, N.; McAnany, J.J.; Zele, A.J.; Gamlin, P.D.; Cao, D. Assessing Rod, Cone, and Melanopsin Contributions to Human Pupil Flicker Responses. Investig. Opthalmol. Vis. Sci. 2014, 55, 719–727. [Google Scholar] [CrossRef] [Green Version]
- Watson, A.B.; Yellott, J.I. A unified formula for light-adapted pupil size. J. Vis. 2012, 12. [Google Scholar] [CrossRef]
- Park, J.C.; McAnany, J.J. Effect of stimulus size and luminance on the rod-, cone-, and melanopsin-mediated pupillary light reflex. J. Vis. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Dreyfuss, H. The Measure of Man;.Human Factors. In Design, The Visual Fields: A Textbook and Atlas of Clinical Perimetry; Whitney Library of Design: New York, NY, USA, 1960. [Google Scholar]
- Ruch, T.C.; Fulton, J.F. Medical Physiology and Biophysics, 18th ed.; W. B. Saunders & Co.: Philadelphia, PA, USA, 1960. [Google Scholar]
- Stanley, P.A.; Davies, A.K. The effect of field of view size on steady-state pupil diameter. Ophthal. Physiol. Opt. 1995, 15, 601–603. [Google Scholar] [CrossRef]
- Optical Society of America. Chapter 24: Optics of the Eye: Part 7: Vision. In Handbook of Optics. Volume 1: Fundamentals, Techniques, and Design, 2nd ed.; Bass, M., Ed.; McGraw-Hill Professional Publishing: New York, NY, USA, 1995. [Google Scholar]
- Pflibsen, K.P.; Pomerantzeff, O.; Ross, R.N. Retinal illuminance using a wide-angle model of the eye. J. Opt. Soc. Am. A 1988, 5, 146–150. [Google Scholar] [CrossRef]
- Dacey, D.M.; Liao, H.-W.; Peterson, B.B.; Robinson, F.R.; Smith, V.C.; Pokorny, J.; Yau, K.-W.; Gamlin, P.D. Melanopsin-expressing ganglion cells in primate retina signal colour and irradiance and project to the LGN. Nat. Cell Biol. 2005, 433, 749–754. [Google Scholar] [CrossRef]
- Gooley, J.J.; Rajaratnam, S.M.W.; Brainard, G.C.; Kronauer, R.E.; Czeisler, C.A.; Lockley, S.W. Spectral Responses of the Human Circadian System Depend on the Irradiance and Duration of Exposure to Light. Sci. Translat. Med. 2010, 2, 31ra33. [Google Scholar] [CrossRef] [Green Version]
- Spitschan, M.; Jain, S.; Brainard, D.H.; Aguirre, G.K. Opponent melanopsin and S-cone signals in the human pupillary light response. Proc. Natl. Acad. Sci. USA 2014, 111, 15568–15572. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, S.; Ishibashi, K.; Aritake, S.; Enomoto, M.; Hida, A.; Tamura, M.; Kozaki, T.; Motohashi, Y.; Mishima, K. Inter-individual difference in pupil size correlates to suppression of melatonin by exposure to light. Neurosci. Lett. 2008, 440, 23–26. [Google Scholar] [CrossRef]
- Winn, B.; Whitaker, D.; Elliott, D.B.; Phillips, N.J. Factors affecting light-adapted pupil size in normal human subjects. Investig. Ophthalmol. Vis. Sci. 1994, 35, 1132–1137. [Google Scholar]
- Farrell, R.J.; Booth, J.M. Design Handbook for Imagery Interpretation Equipment; Boeing Aerospace Company: Seattle, WA, SUA, 1975. [Google Scholar]
- Yu, M.; Kautz, M.A.; Thomas, M.L.; Johnson, D.; Hotchkiss, E.R.; Russo, M.B. Operational implications of varying ambient light levels and time-of-day effects on saccadic velocity and pupillary light reflex. Ophthalmic Physiol. Opt. 2007, 27, 130–141. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bracke, P.; Van de Putte, E.; Ryckaert, W.R. Comment Concerning the Effects of Light Intensity on Melatonin Suppression in the Review “Light Modulation of Human Clocks, Wake, and Sleep” by A. Prayag et al. Clocks & Sleep 2021, 3, 181-188. https://0-doi-org.brum.beds.ac.uk/10.3390/clockssleep3010011
Bracke P, Van de Putte E, Ryckaert WR. Comment Concerning the Effects of Light Intensity on Melatonin Suppression in the Review “Light Modulation of Human Clocks, Wake, and Sleep” by A. Prayag et al. Clocks & Sleep. 2021; 3(1):181-188. https://0-doi-org.brum.beds.ac.uk/10.3390/clockssleep3010011
Chicago/Turabian StyleBracke, Peter, Eowyn Van de Putte, and Wouter R. Ryckaert. 2021. "Comment Concerning the Effects of Light Intensity on Melatonin Suppression in the Review “Light Modulation of Human Clocks, Wake, and Sleep” by A. Prayag et al." Clocks & Sleep 3, no. 1: 181-188. https://0-doi-org.brum.beds.ac.uk/10.3390/clockssleep3010011