1. Introduction
Major threats to ecosystem functioning include the loss of biodiversity and habitat fragmentation due to human activities [
1]. As a result, wildlife populations may become increasingly isolated from one another [
2,
3,
4], increasing the likelihood of inbreeding and genetic drift-induced differentiation [
2,
4,
5]. Broader ecological impacts of isolation include higher levels of intra- and interspecific competition in isolated habitat patches [
6,
7], and a resulting displacement of individuals into high-risk areas with increased anthropogenic effects [
8,
9].
Ensuring inter-patch connectivity in fragmented environments is paramount to mitigating the effects of isolation and improving species survival [
3]. Two broad definitions of connectivity exist: structural and functional [
10]. A structural corridor describes the physical environment and excludes the behavioural response of individuals [
10]. Alternatively, a corridor becomes functional when it enables individuals to move along these corridors, promoting inter-path connectivity and enhancing genetic exchange [
10,
11,
12]. Therefore, the ability of individuals to move across the landscape depends on complex interactions. For example, human-associated landscapes often contain barriers to animal movement such as linear transport lines [
13], transformed landscapes (e.g., cultivated areas [
14]) and human-caused mortality, either accidentally (e.g., vehicle collisions) or intentionally (e.g., hunting, human–wildlife conflict; [
15,
16]). Similarly, natural landscapes can also contain barriers (e.g., steep ravines, large lakes [
17]), while prey availability and distribution also contribute to species distribution (e.g., resource dispersion hypothesis, [
18]).
Understanding the factors driving habitat selection provides opportunities to predict species distribution, and to identify potential functional corridors linking populations [
19]. We used a multivariate approach, maximum entropy (Maxent), to correlate species presence data and environmental covariates to predict habitat distribution of species across the landscape [
20]. The isolation by resistance hypothesis (IBR) [
21] predicts a positive correlation between the isolation of populations and the level of resistance separating these populations. Low-quality habitat and non-habitat may have scarce resources and a high mortality risk, resulting in landscapes with higher resistance, while high-quality habitat contains abundant resources and low mortality risk, providing a permeable landscape. By incorporating species habitat distribution, derived using data from the target species, as a resistance landscape into corridor modelling, functional corridors can be delineated using circuit theory [
21,
22]. Circuit theory considers movement based on random walk algorithms and assumes the individual has no insight into landscape heterogeneity, thereby predicting many potential routes for current flow where there is the least resistance [
21,
22]. Incorporating spatial heterogeneity of landscapes along with random walk considerations gives IBR a strong theoretical foundation for predicting functional corridors [
19,
23,
24].
Their large spatial requirements, low reproductive rates, strong association with high species richness, and their regularity roles in terrestrial ecosystems make large carnivores a useful focal species to predict landscape connectivity at regional scale for sympatric species [
25,
26,
27]. Despite being highly adaptable and able to occupy a wide range of environments, leopard (
Panthera pardus) survival is threatened by habitat loss and fragmentation [
15,
25]. These threats are further compounded by human-caused mortality due to human-carnivore conflict, and hunting [
15,
28,
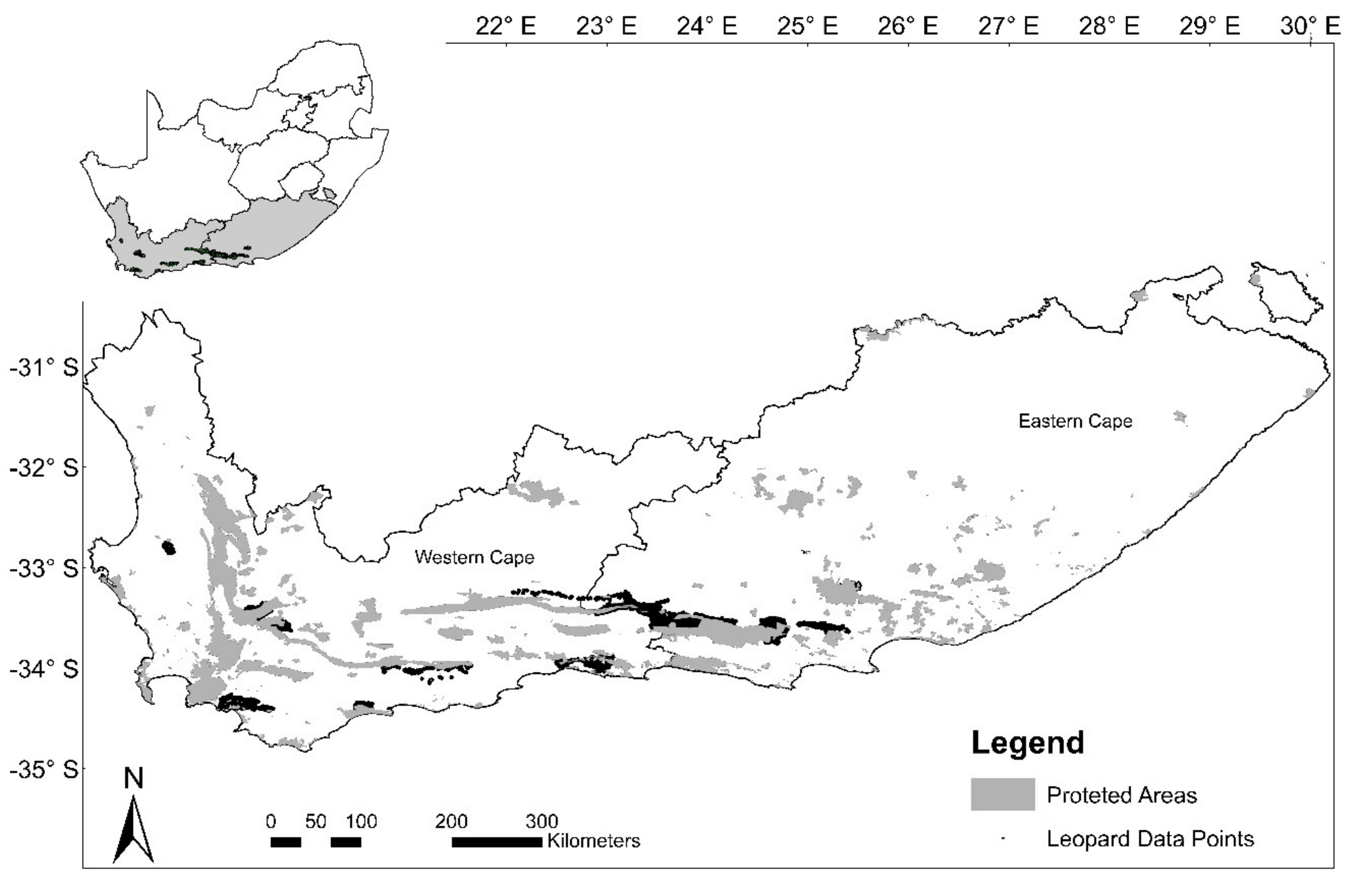
29]. Leopards in the Eastern and Western Cape provinces of South Africa are reported to have a discrete population structure, low population densities, and are persecuted due to high levels of human-carnivore conflict [
4,
30,
31,
32]. In the face of these threats, identifying corridors connecting habitat patches and reducing human–carnivore conflict can promote gene flow and the long-term survival of the leopard and sympatric species. In this study, we aim to identify leopard habitat, predict potential landscape corridors linking habitat patches, and test functionality between predicted habitat patches to determine whether leopard populations in the region can connect despite modified landscapes. First, we evaluate the relationship between leopard presence and anthropogenic and natural covariates, and use the model with the best fit to predict leopard habitat. Second, we employ circuit theory [
33], to predict potential corridors linking 14 known leopard populations using the modelled leopard habitat distribution as a surface layer. Finally, while incorporating leopard occurrence data to random walk algorithms using circuit theory improves the identification of functional corridors [
21], we used camera trap surveys to validate and test the functionality of five predicted corridors linking potentially isolated leopard populations in the Western Cape.
4. Discussion
Wide-ranging mammals depend on large enough habitat patches and habitat connectivity to persist in fragmented landscapes [
25,
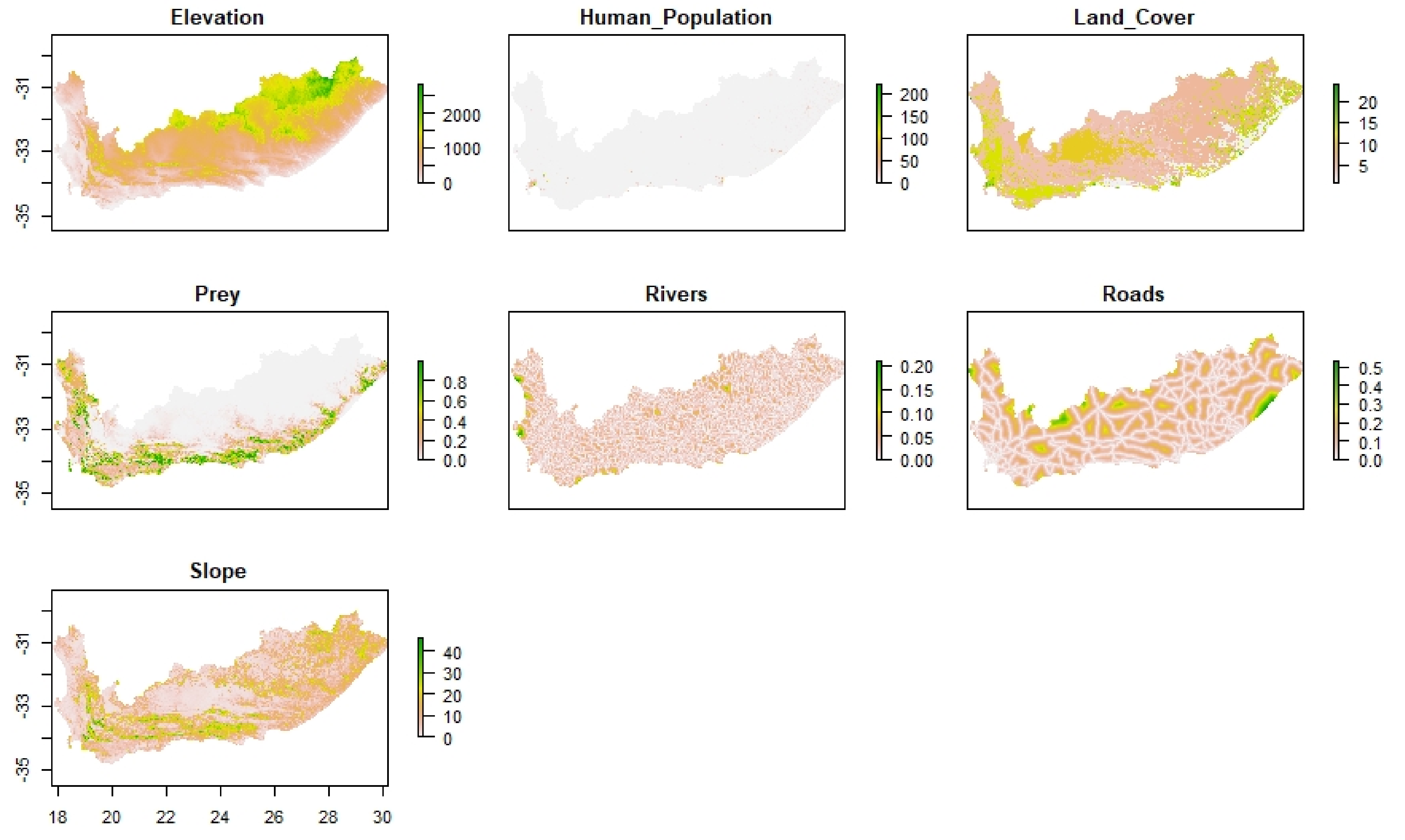
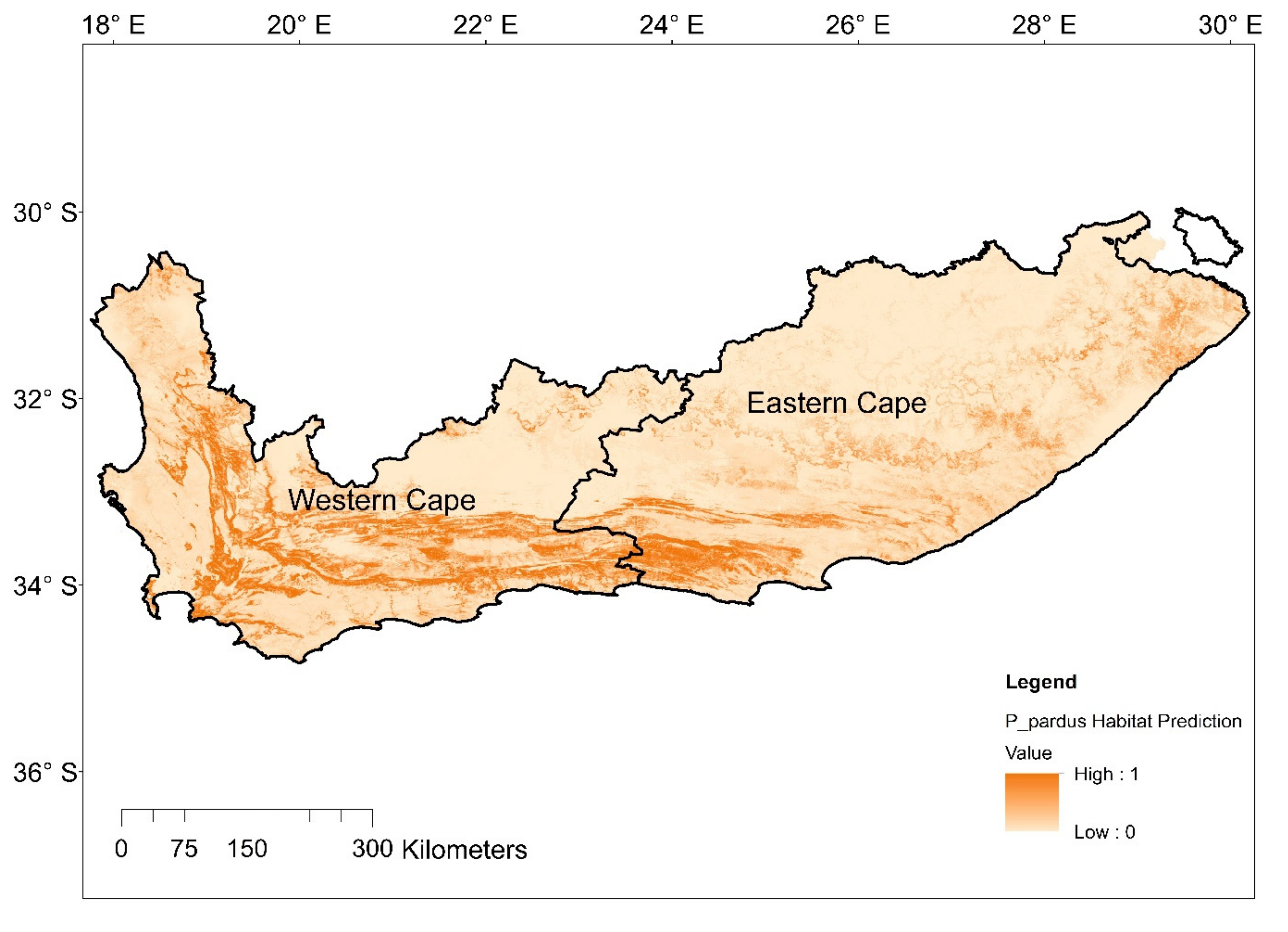
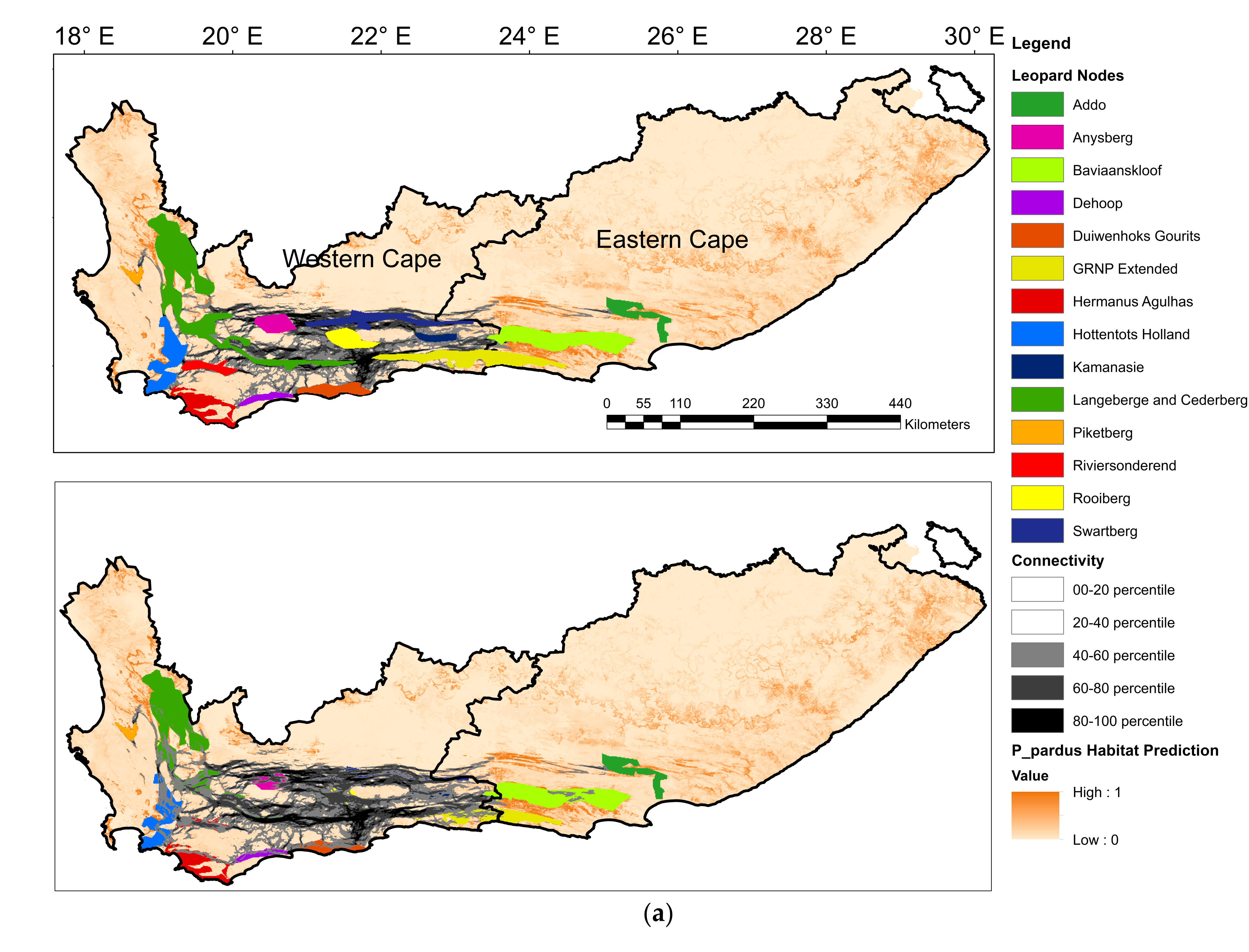
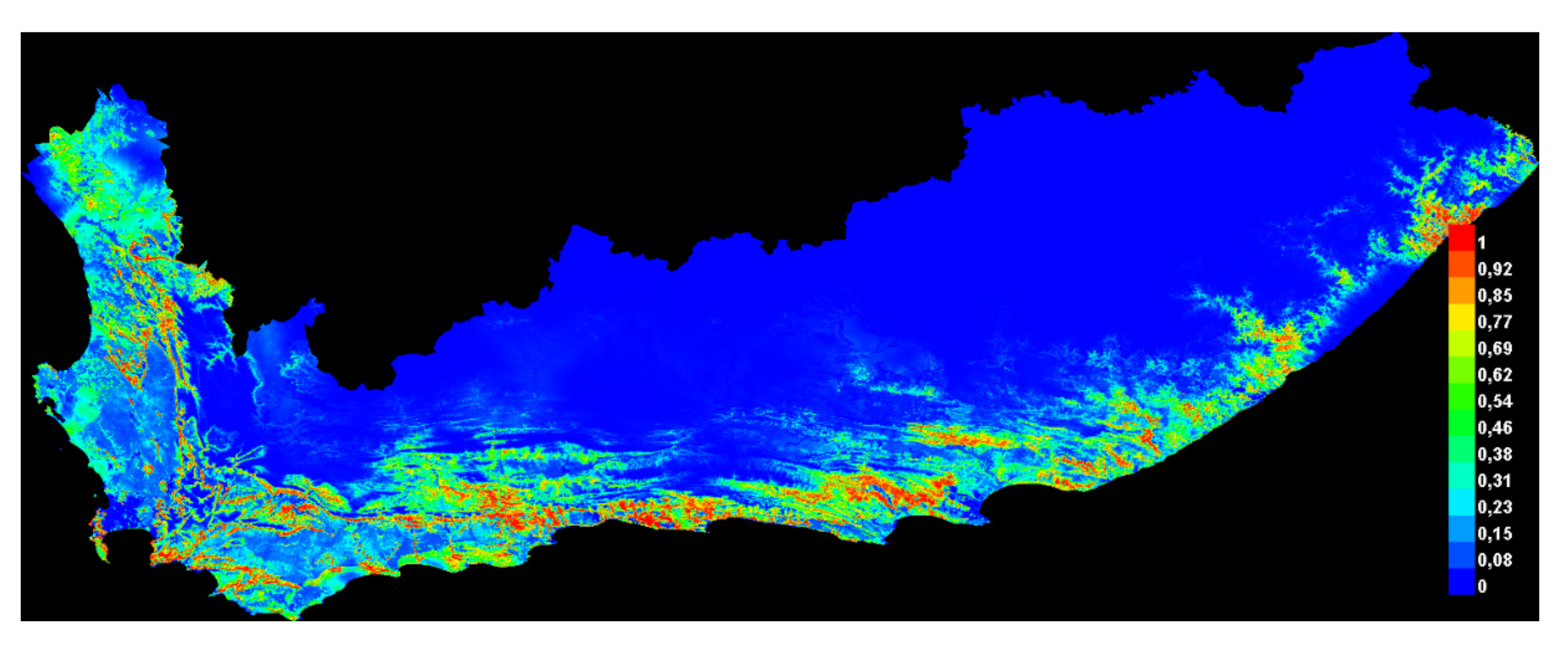
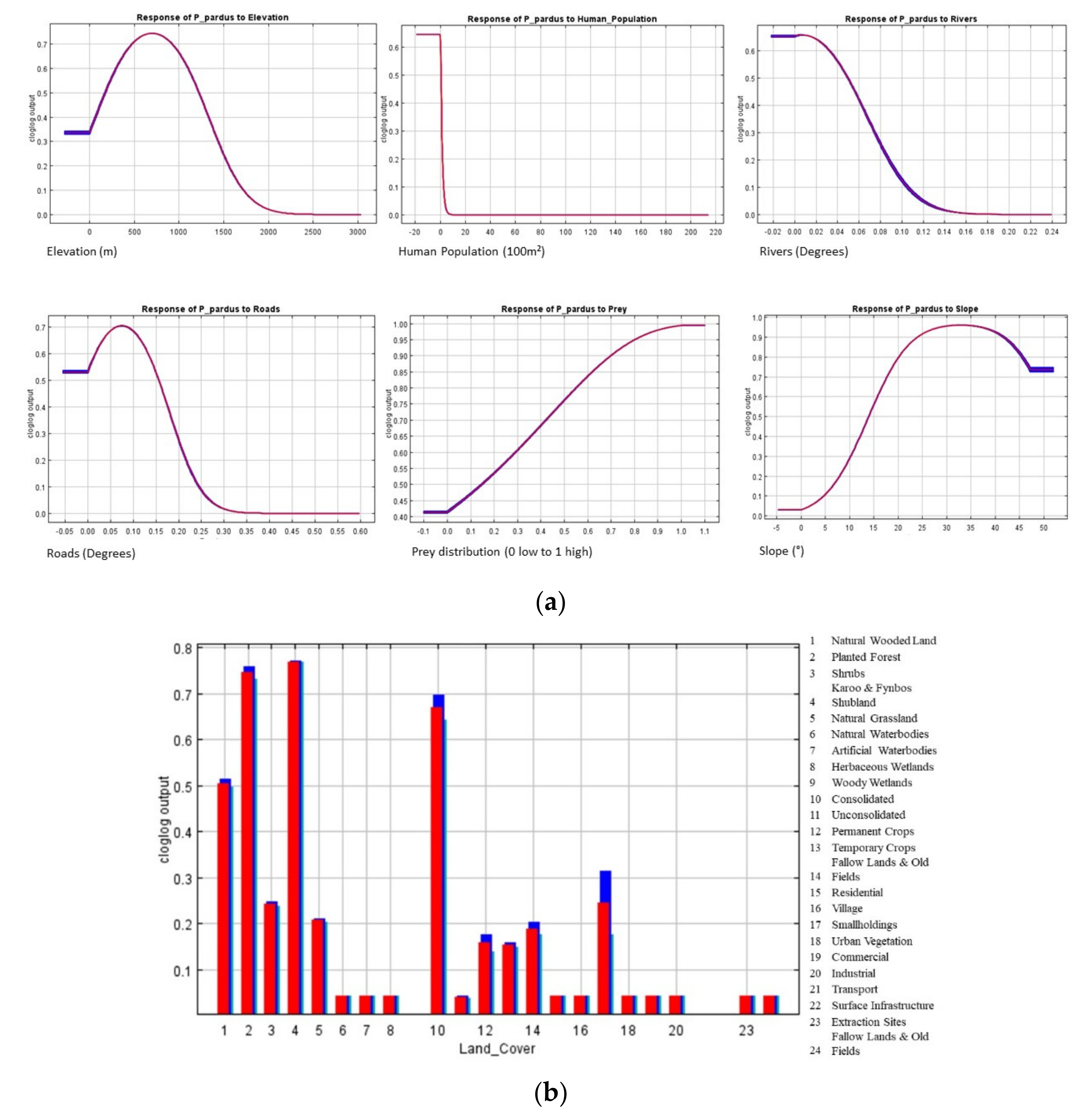
29]. We combined species data and environmental variables to predict leopard habitat distribution and delineate landscape connectivity. We found that leopard habitat in the Eastern Cape and Western Cape was strongly correlated to moderate slopes and areas of natural land-cover such as forests, shrublands as well as forest plantations. Modified landscapes, such as cultivated land, and highly human populated areas appeared to obstruct leopard presence. As a result, high-quality leopard habitat appeared to be continuous along the east and west direction of the prominent mountain ranges expanding across the two provinces and continued in a north and south directions along the Cederberg Mountain chain in the west (
Figure 5). Mountainous, rugged areas offer important habitat for leopard and other large carnivores [
39,
62,
63,
64]. However, leopards occupy non-mountainous areas in regions such as Kwa-Zulu Natal, South Africa, and elsewhere in Africa [
65,
66]. Therefore, it is likely that mountainous areas are strongly selected because these areas offer refugia from human activity [
15,
39]. The distributional range estimated in this study appeared to correspond well with that described by Swanepoel et al. [
39].
The connectivity model using Circuitscape is a useful addition to habitat prediction in delineating potential functional corridors linking habitat patches across complex landscapes. This allowed the identification of leopard nodes that may be in risk of isolation due to low predicted connectivity. The easternmost leopard node (Addo) appeared to have severed connectivity to neighbouring leopard habitat patches. The nearest node to Addo was the Baviaanskloof node, where potential barriers such as cultivated areas and human population density may contribute to the apparent connectivity impediment. Connectivity was also restricted between Addo and habitat in the northwest direction where connectivity to the Swartberg node also appeared to be severed, likely due to a combination of transformed landscapes and limited slopes being present in this area. The Hermanus-Agulhas and De Hoop (HA and DH) nodes showed limited connectivity potential to leopard populations along the Langeberg–Cederberg and Riviersonderend nodes.
The genetic population structure of leopards in the Eastern Cape and Western Cape is reportedly represented by three sub-structured populations [
4]. One of these populations occupies the HA and DH node, where gene flow to the other two genetic populations to the north and northeast was reported to be very low [
4]. This provides support of our findings of limited leopard habitat and connectivity between the HA and DH nodes to the northern nodes [
4]. Human-associated landscape features are known to fragment natural landscapes and restrict mammalian connectivity [
4,
15,
25,
27]. The vast cultivated landscapes between the Langeberg–Cederberg node and the HA and DH nodes appear to fragment habitat and restrict connectivity in the region.
Identifying and promoting movement along corridors can offset the negative effects of habitat fragmentation by connecting isolated populations [
24,
59,
67]. We identified narrow, linear corridors running through the highly transformed area between the Langeberg habitat to the coastal Duiwenhoks habitat. These areas were also identified as suitable habitat based on our prediction modelling, and were characterised by large river courses (Breede, Duiwenhoks, Goukou and Gourits), moderate slopes with some remnant riparian vegetation, surrounded by large areas of transformed, cultivated lands. In some places, these corridors become extremely narrow (<20 m) between the river and the transformed agricultural surrounds. These narrow areas caused breaks in habitat prediction, since the model evaluates suitability based on the average conditions at approximately a 1 km
2 scale, but these were identified as potential corridors in the Circuitscape modelling. These corridors may play a critical role in the region, as these sites represent the last potential habitat corridors linking disjunct leopard populations in the region.
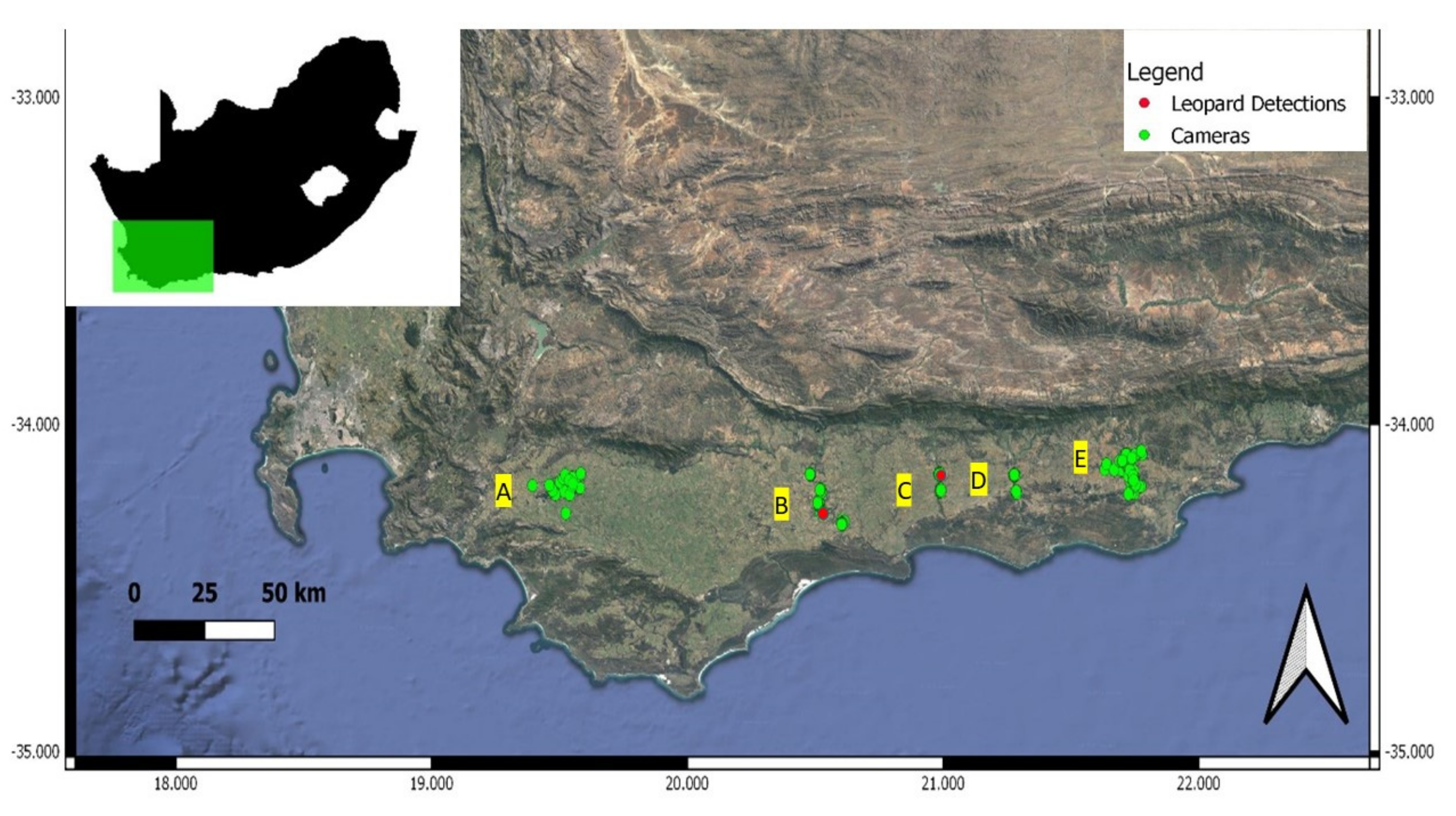
While incorporating target specific occurrence data to derive species habitat distribution improves the identification of functional corridors [
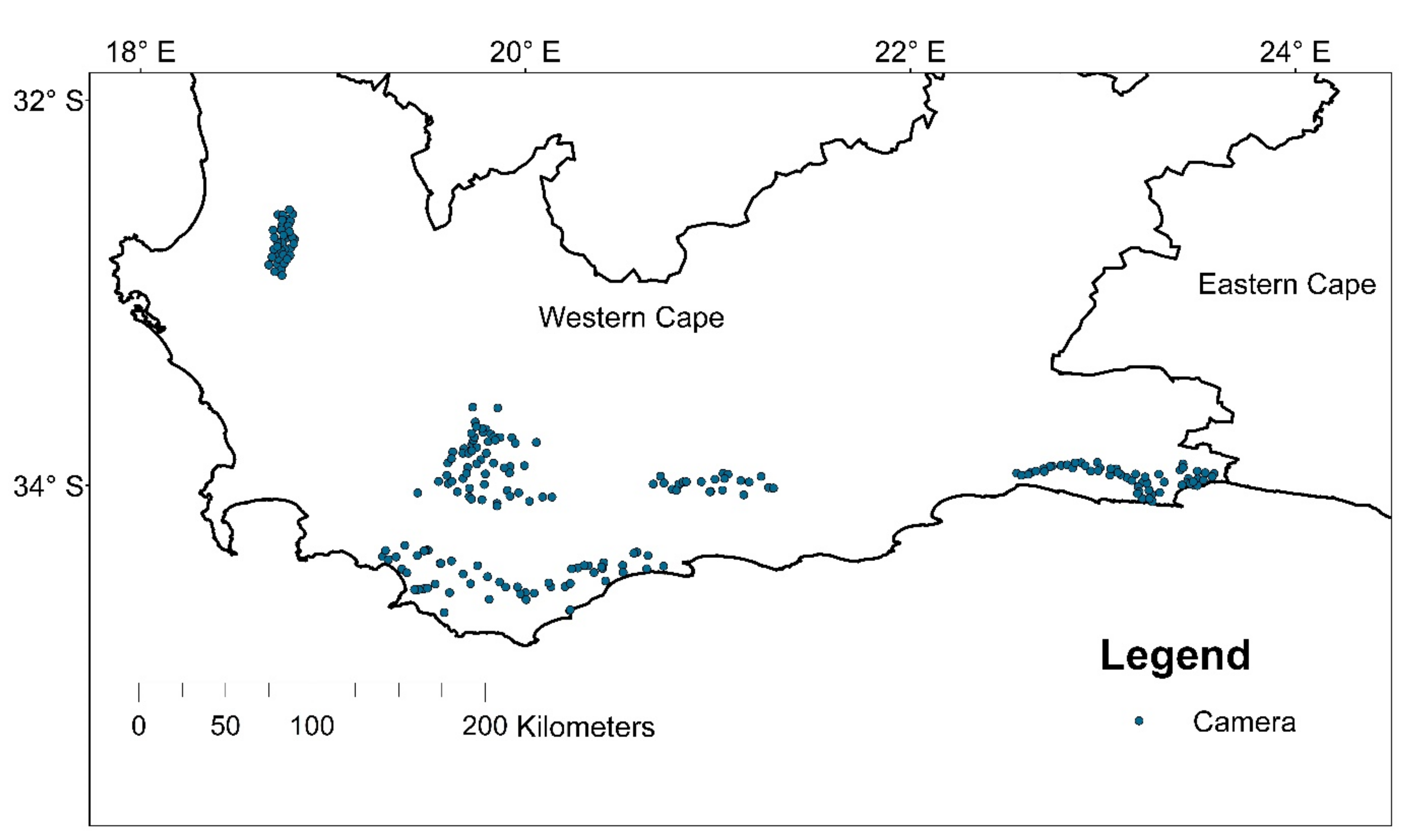
21], validating corridors using independent data contributes to confirming targeted conservation management efforts [
68]. Using camera traps, we detected leopard on two of the river corridors delineated by Maxent and Circuitscape, indicating their use by leopards to some degree. The low leopard detection rates along the river corridors may be expected, as gene flow from source populations in the area is reported to be low [
4] and could further be compounded by the naturally low dispersal rates of leopards [
69], as well as narrow strips of natural landscapes offering limited sites to place cameras to optimize leopard detection. The river corridors hosted between 23 and 32 mammalian wildlife species, confirming the high ecological value these corridors have in the region. While leopards might rarely use these corridors, these areas may contribute to the long-term genetic rigor and survival of the species in the region. This has implications for the conservation of river courses and associated riparian vegetation, particularly where the surroundings are highly modified. The existence of reduced genetic heterogeneity found for leopards in the region [
4] suggests, however, that these predicted corridors do not significantly overcome the gene flow barriers to ensure populations are adequately genetically connected. Ensuring corridors facilitate the successful movement and gene flow among leopard populations is key to local population persistence. Dispersal rates are driven by complex factors such has population density [
69,
70]. When populations are near saturation, and no space is available for maturing animals to occupy, dispersal rates increase [
69,
70]. Therefore, where populations suffer from stressors that depress the population, dispersal rates would be lower [
70]. Obtaining data from dispersal or the movements of translocated individuals in the area would contribute to better understanding the likelihood of corridors to contribute to leopard meta-population dynamics in the region [
71].
Management Implications
Natural landscapes have diminished substantially over the past century, and the leopard is only found in 25% of its historical distribution [
28]. This loss of range can be attributed to both extensive habitat transformation [
72], which impacts both leopards and prey species, resulting in a loss of available prey [
28,
40], and to high levels of human caused mortality, principally via human–wildlife conflict and unsustainable leopard harvesting [
15,
73]. Protected areas are insufficient to contribute to leopard conservation in South Africa [
15,
39]. Therefore, species management practices have important implications in conserving meta-population dynamics, and managing the genetic viability of isolated populations may become increasingly necessary as altered habitat expands [
74,
75].
Connectivity is the most frequently cited recommendation to conserve species and ecosystem functioning [
76,
77]. Our study identified habitat permeability estimates, which highlight areas requiring focused conservation efforts where connectivity might need to be increased. We raise particular concern regarding the limited evidence of connectivity between the HA and DH populations and their counterparts along the Langeberg and Riviersonderend nodes. This may require active management interventions such as maintaining and expanding riparian zones, particularly in transformed areas. Further research is required to determine the presence of leopard populations beyond Addo to the east to predict connectivity potential eastward. However, our study also highlights the limited connectivity between Addo and populations to the west. Human-caused mortality is a major threat to leopard survival and reducing human–carnivore conflict in and around protected areas, predicted leopard habitat, and corridors will contribute to leopard conservation [
15]. Therefore, we encourage conservationists and governmental institutions to advocate for non-lethal conflict mitigation strategies [
78]. This can promote community tolerance and raise awareness of the regulatory-role carnivores play in ecosystems [
25,
26,
27]. Restoring degraded landscapes through strategic land-use planning adjacent to areas such as protected areas, leopard habitat and corridors, would be a valuable conservation effort. Where connectivity is constrained and cannot be overcome with available habitat connections, and genetic structuring is apparent, and populations densities are low, active local translocations could be prudent to bolster genetic heterogeneity [
74,
79,
80].
Over the last three decades, connectivity projects were employed at state level in the USA [
81], national level in Bhutan [
82] and continental scale in Europe (Natura 2000, EU Habitat Directive). However, no regional connectivity maps exist for carnivore conservation in South Africa. Coordinated planning between municipalities, provinces, and national regions with inputs from private, non-governmental and government conservation agencies should be adopted in South Africa to reduce and perhaps reverse the negative effects of habitat loss. Our models provide the first interprovincial connectivity map for leopard in South Africa. Our results can assist environmental managers in identifying sensitive and important biodiversity corridors to ensure species persistence. We recommend species management plans address the connectivity of leopard populations at provincial, national and even pan-African scales. This should be considered in transport and large development projects in South Africa to promote species conservation (e.g., Trans-Canada Highway, Banff National Park [
83].


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}









