
First Report of the Parasitic Nematode Pseudoterranova spp. Found in Mediterranean Monk Seal (Monachus monachus) in Greece: Conservation Implications


 ,
,  and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Parasite Collection and Molecular Techniques
2.2. Statistical Methods
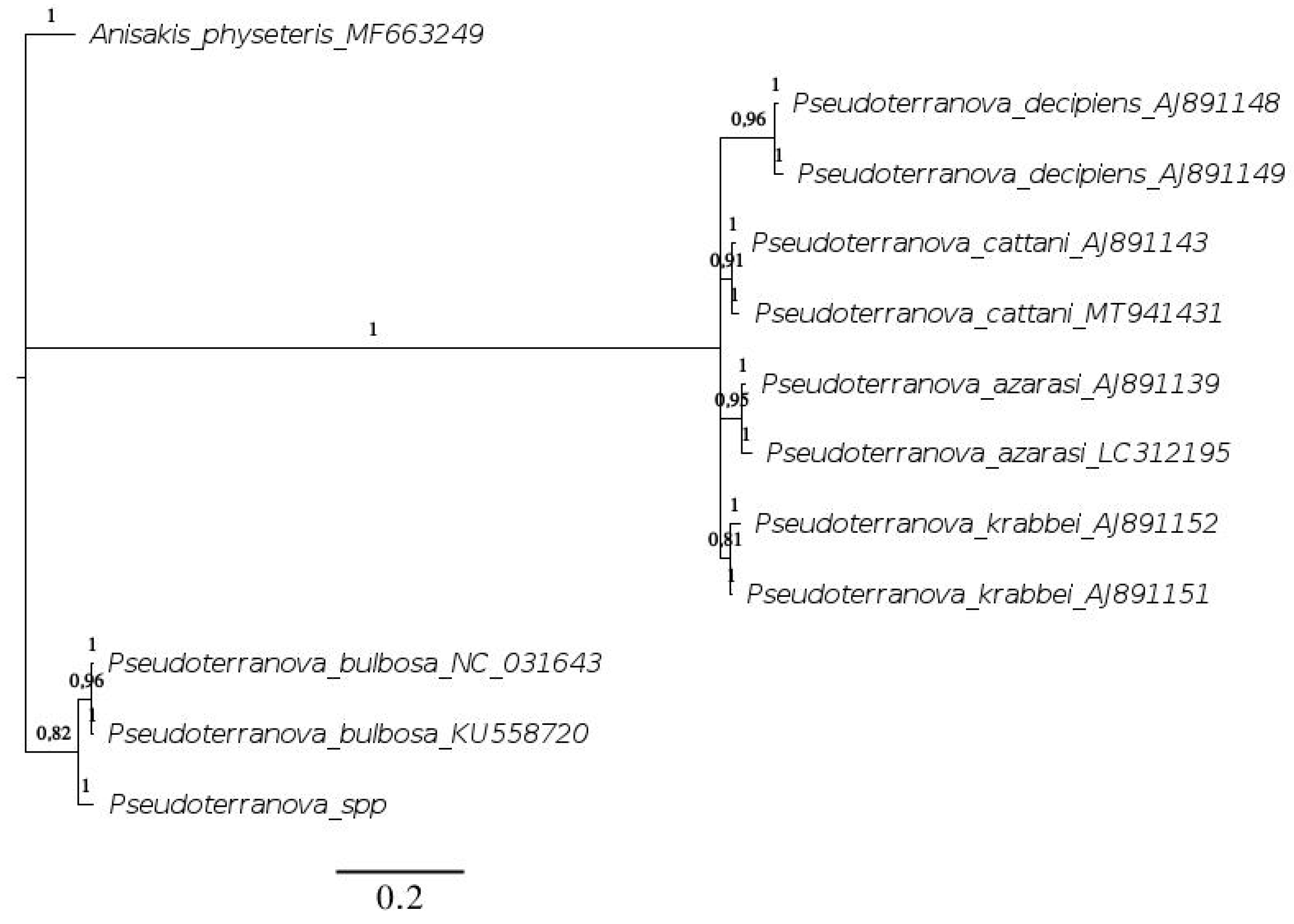
3. Results
4. Discussion
4.1. Parasitic Species Identification
4.2. Parasitic Dispersal
4.3. Genetic Relationships
5. Management and Conservation Implications
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Karamanlidis, A.A.; Dendrinos, P.; de Larrinoa, P.F.; Gücü, A.C.; Johnson, W.M.; Kiraç, C.O.; Pires, R. The Mediterranean Monk Seal M Onachus Monachus: Status, Biology, Threats, and Conservation Priorities: The Mediterranean Monk Seal. Mammal Rev. 2016, 46, 92–105. [Google Scholar] [CrossRef]
- Pires, R.; Neves, H.C.; Karamanlidis, A.A. The Critically Endangered Mediterranean Monk Seal Monachus monachus in the Archipelago of Madeira: Priorities for Conservation. Oryx 2008, 42, 278–285. [Google Scholar] [CrossRef] [Green Version]
- González, L.M.; Cedenilla, M.A.; de Larrinoa, P.F.; Layna, J.F.; Aparicio, F. Changes in the Breeding Variables of the Mediterranean Monk Seal (Monachus monachus) Colony of Cabo Blanco Peninsula after a Mass Mortality Episode. Mammalia 2002, 66, 173–182. [Google Scholar] [CrossRef]
- Martínez-Jauregui, M.; Tavecchia, G.; Cedenilla, M.; Coulson, T.; Fernández de Larrinoa, P.; Muñoz, M.; González, L. Population Resilience of the Mediterranean Monk Seal Monachus monachus at Cabo Blanco Peninsula. Mar. Ecol. Prog. Ser. 2012, 461, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Dendrinos, P.; Karamanlidis, A.A.; Kotomatas, S.; Paravas, V.; Adamantopoulou, S. Report of a New Mediterranean Monk Seal (Monachus monachus) Breeding Colony in the Aegean Sea, Greece. Aquat. Mamm. 2008, 34, 355–361. [Google Scholar] [CrossRef]
- Gucu, A.C.; Gucu, G.; Orek, H. Habitat Use and Preliminary Demographic Evaluation of the Critically Endangered Mediterranean Monk Seal (Monachus monachus) in the Cilician Basin (Eastern Mediterranean). Biol. Conserv. 2004, 116, 417–431. [Google Scholar] [CrossRef]
- Güçlüsoy, H. Status of the Mediterranean Monk Seal, Monachus monachus (Hermann, 1779) in the Coastal Waters of Turkey. Ege J. Fish. Aquat. Sci. 2004, 21, 201–210. [Google Scholar]
- Adamantopoulou, S.; Androukaki, E.; Dendrinos, P.; Kotomatas, S.; Paravas, V.; Psaradellis, M.; Tounta, E.; Karamanlidis, A. Movements of Mediterranean Monk Seals (Monachus monachus) in the Eastern Mediterranean Sea. Aquat. Mamm. 2011, 37, 256. [Google Scholar] [CrossRef]
- Pool, R.; Chandradeva, N.; Gkafas, G.; Raga, J.A.; Fernández, M.; Aznar, F.J. Transmission and Predictors of Burden of Lungworms of the Striped Dolphin (Stenella coeruleoalba) in the Western Mediterranean. J. Wildl. Dis. 2019, 56, 186–191. [Google Scholar] [CrossRef]
- Golani, D. Impact of Red Sea Fish Migrants through the Suez Canal on the Aquatic Environment of the Eastern Mediterranean. Bull. Ser. Yale Sch. For. Environ. Stud. 1998, 103, 375–387. [Google Scholar]
- Rilov, G.; Galil, B. Marine Bioinvasions in the Mediterranean Sea–History, Distribution and Ecology. In Biological Invasions in Marine Ecosystems; Springer: Berlin/Heidelberg, Germany, 2009; pp. 549–575. [Google Scholar]
- Özbilgin, Y.; Kalecik, E.; Gücü, A.C. First Record of Humpback Dolphins in Mersin Bay, the Eastern Mediterranean, Turkey. Turk. J. Fish. Aquat. Sci. 2018, 18, 187–190. [Google Scholar] [CrossRef]
- Frantzis, A. A Long and Deep Step in Range Expansion of an Alien Marine Mammal in the Mediterranean: First Record of the Indian Ocean Humpback Dolphin Sousa plumbea (G. Cuvier, 1829) in the Greek Seas. BioInvasions Rec. 2018, 7, 83–87. [Google Scholar] [CrossRef]
- Colloca, F.; Scannella, D.; Geraci, M.L.; Falsone, F.; Batista, G.; Vitale, S.; Di Lorenzo, M. British Sharks in Sicily: Records of Long Distance Migration of Tope Shark (Galeorhinus Galeus) from North-Eastern Atlantic to Mediterranean Sea. Mediterr. Mar. Sci. 2019, 20, 309–313. [Google Scholar] [CrossRef]
- Scheinin, A.P.; Kerem, D.; MacLeod, C.D.; Gazo, M.; Chicote, C.A.; Castellote, M. Gray Whale (Eschrichtius robustus) in the Mediterranean Sea: Anomalous Event or Early Sign of Climate-Driven Distribution Change? Mar. Biodivers. Rec. 2011, 4, e28. [Google Scholar] [CrossRef] [Green Version]
- Gauffier, P.; Verborgh, P.; Giménez, J.; Esteban, R.; Sierra, J.M.S.; de Stephanis, R. Contemporary Migration of Fin Whales through the Strait of Gibraltar. Mar. Ecol. Prog. Ser. 2018, 588, 215–228. [Google Scholar] [CrossRef]
- Kapiris, K.; Apostolidis, C.; Baldacconi, R.; Başusta, N.; Bilecenoglu, M.; Bitar, G.; Bobori, D.; Boyaci, Y.Ö.; Dimitriadis, C.; Djurović, M. New Mediterranean Biodiversity Records (April, 2014). Mediterr. Mar. Sci. 2014, 15, 198–212. [Google Scholar] [CrossRef] [Green Version]
- Beqiraj, S.; Kashta, L. The Establishment of Blue Crab Callinectes sapidus Rathbun, 1896 in the Lagoon of Patok, Albania (South-East Adriatic Sea). Aquat. Invasions 2010, 5, 219–221. [Google Scholar] [CrossRef]
- Deval, M.C.; Kaya, Y.; Güven, O.; Gökoğlu, M.; Froglia, C. An Unexpected Find of the Western Atlantic Shrimp, Farfantepenaeus Aztecus (Ives, 1891) (Decapoda, Penaeidae) in Antalya Bay, Eastern Mediterranean Sea. Crustaceana 2010, 83, 1531–1537. [Google Scholar]
- Kampouris, T.E.; Gkafas, G.A.; Sarantopoulou, J.; Exadactylos, A.; Batjakas, I.E. An American in the Aegean: First Record of the American Lobster Homarus Americanus, H. Milne Edwards, 1837 from the Eastern Mediterranean Sea. BioInvasions Rec. 2021, in press. [Google Scholar] [CrossRef]
- Amilhat, E.; Aarestrup, K.; Faliex, E.; Simon, G.; Westerberg, H.; Righton, D. First Evidence of European Eels Exiting the Mediterranean Sea during Their Spawning Migration. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dunn, A.M.; Hatcher, M.J. Parasites and Biological Invasions: Parallels, Interactions, and Control. TRENDS Parasitol. 2015, 31, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, E.; Loukopoulos, P.; Komnenou, A.; Androukaki, E.; Karamanlidis, A. First Report of Acanthocheilonema Spirocauda in the Mediterranean Monk Seal (Monachus monachus). J. Wildl. Dis. 2010, 46, 570–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ELEY, T.J. Dipetalonema spirocauda in Alaskan marine mammals. J. Wildl. Dis. 1981, 17, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Measures, L.N.; Gosselin, J.-F.; Bergeron, E. Heartworm, Acanthocheilonema spirocauda (Leidy, 1858), Infections in Canadian Phocid Seals. Available online: https://cdnsciencepub.com/doi/10.1139/f96-342 (accessed on 27 December 2021).
- Komnenou, A.T.; Gkafas, G.A.; Kofidou, E.; Sarantopoulou, J.; Exadactylos, A.; Tounta, E.; Koemtzopoulos, K.; Dendrinos, P.; Karamanlidis, A.A.; Gulland, F.; et al. First Report of Uncinaria hamiltoni in Orphan Eastern Mediterranean Monk Seal Pups in Greece and Its Clinical Significance. Pathogens 2021, 10, 1581. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a Healthy Ecosystem One That Is Rich in Parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Thompson, R.; Lymbery, A.; Smith, A. Parasites, Emerging Disease and Wildlife Conservation. Int. J. Parasitol. 2010, 40, 1163–1170. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.M.; May, R.M. Population Biology of Infectious Diseases: Part, I. Nature 1979, 280, 361–367. [Google Scholar] [CrossRef]
- Vos, J.G.; Bossart, G.; Fournier, M.; O’Shea, T. Toxicology of Marine Mammals; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar]
- Stroud, R.K.; Dailey, M.D. Parasites and Associated Pathology Observed in Pinnipeds Stranded along the Oregon Coast. J. Wildl. Dis. 1978, 14, 292–298. [Google Scholar] [CrossRef] [Green Version]
- Salman, A.; Bilecenoglu, M.; Güçlüsoy, H. Stomach Contents of Two Mediterranean Monk Seals (Monachus monachus) from the Aegean Sea, Turkey. J. Mar. Biol. Assoc. UK 2001, 81, 719–720. [Google Scholar] [CrossRef]
- Pierce, G.J.; Hernandez-Milian, G.; Santos, M.B.; Dendrinos, P.; Psaradellis, M.; Tounta, E.; Androukaki, E.; Edridge, A. Diet of the Monk Seal (Monachus monachus) in Greek Waters. Aquat. Mamm. 2011, 37, 284. [Google Scholar] [CrossRef]
- Kıraç, C.O.; Ok, M. Diet of a Mediterranean Monk Seal Monachus monachus in a Transitional Post-Weaning Phase and Its Implications for the Conservation of the Species. Endanger. Species Res. 2019, 39, 315–320. [Google Scholar] [CrossRef]
- Karamanlidis, A.A.; Kallianiotis, A.; Psaradellis, M.; Adamantopoulou, S. Stomach Contents of a Subadult Mediterranean Monk Seal (Monachus monachus) from the Aegean Sea. Aquat. Mamm. 2011, 37, 280. [Google Scholar] [CrossRef]
- Mattiucci, S.; Cipriani, P.; Levsen, A.; Paoletti, M.; Nascetti, G. Molecular Epidemiology of Anisakis and Anisakiasis: An Ecological and Evolutionary Road Map. In Advances in Parasitology; Elsevier: Amsterdam, The Netherlands, 2018; Volume 99, pp. 93–263. [Google Scholar] [CrossRef]
- Campana-Rouget, Y. Sur deux nouveaux genres de Spirurides parasites de Poissons; Discussion systématique des genres voisins. Ann. Parasitol. Hum. Comparée 1955, 30, 346–362. [Google Scholar] [CrossRef]
- Schnapp, B.; Hellwing, S.; Ghizelea, G. Contributions to the Knowledge of the Black Sea Seal (Monachus monachus) Herm. Trav. Mus. D’Histoire Nat. Gr. Antipa 1962, 3, 383–400. [Google Scholar]
- Blagoveshtchensky, D. New Forms of Lice (Siphunculata) Parasites of Pinnipeds and Hares. Rev. Entom URSS 1966, 45, 806–813. [Google Scholar]
- Mackiewicz, J.S. Cestode Transmission Patterns. J. Parasitol. 1988, 74, 60–71. [Google Scholar] [CrossRef]
- Paggi, L.; Nascetti, G.; Cianchi, R.; Orecchia, P.; Mattiucci, S.; D’Amelio, S.; Berland, B.; Brattey, J.; Smith, J.; Bullini, L. Genetic Evidence for Three Species within Pseudoterranova decipiens (Nematoda, Ascaridida, Ascaridoidea) in the North Atlantic and Norwegian and Barents Seas. Int. J. Parasitol. 1991, 21, 195–212. [Google Scholar] [CrossRef]
- Paggi, L.; Mattiucci, S.; Gibson, D.I.; Berland, B.; Nascetti, G.; Cianchi, R.; Bullini, L. Pseudoterranova Decipiens Species A and B (Nematoda, Ascaridoidea): Nomenclatural Designation, Morphological Diagnostic Characters and Genetic Markers. Syst. Parasitol. 2000, 45, 185–197. [Google Scholar] [CrossRef]
- Lunneryd, S.-G.; Boström, M.K.; Aspholm, P.E. Sealworm (Pseudoterranova decipiens) Infection in Grey Seals (Halichoerus grypus), Cod (Gadus morhua) and Shorthorn Sculpin (Myoxocephalus scorpius) in the Baltic Sea. Parasitol. Res. 2015, 114, 257–264. [Google Scholar] [CrossRef] [Green Version]
- McClelland, G. The Trouble with Sealworms (Pseudoterranova decipiens Species Complex, Nematoda): A Review. Parasitology 2002, 124, 183–203. [Google Scholar] [CrossRef]
- Martell, D.; McClelland, G. Transmission of Pseudoterranova decipiens (Nematoda: Ascaridoidea) via Benthic Macrofauna to Sympatric Flatfishes (Hippoglossoides platessoides, Pleuronectes ferrugineus, P. americanus) on Sable Island Bank, Canada. Mar. Biol. 1995, 122, 129–135. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol Biotechnol 1994, 3, 294–299. [Google Scholar] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Tavaré, S. Some Probabilistic and Statistical Problems in the Analysis of DNA Sequences. Lect. Math. Life Sci. 1986, 17, 57–86. [Google Scholar]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Cao, Z.; Zhu, X.Z.Q.; Weng, Y.-B.; Lin, R.-Q.; Li, M.-W.; Zou, F.C.; He, F. Polymorphisms in Mitochondrial LsrRNA and Cox1 Genes within and between Members of Pseudoterranova decipiens Complex. Chin. Vet. J. 2005, 25, 600–603. [Google Scholar]
- Ebmer, D.; Navarrete, M.J.; Muñoz, P.; Flores, L.M.; Gärtner, U.; Brabec, J.; Poppert, S.; Taubert, A.; Hermosilla, C. Anthropozoonotic Parasites Circulating in Synanthropic and Pacific Colonies of South American Sea Lions (Otaria flavescens): Non-Invasive Techniques Data and a Review of the Literature. Front. Mar. Sci. 2020, 847. [Google Scholar] [CrossRef]
- Tokoro, M.; Yamaguchi, H.; Ito, Y.; Mizuno, T.; Mitsuboshi, A.; Kasai, M. Extra-Gastrointestinal Anisakidosis Caused by Pseudoterranova azarasi Manifesting as Strangulated Inguinal Hernia. Parasitol. Int. 2017, 66, 810–812. [Google Scholar]
- Liu, G.-H.; Nadler, S.A.; Liu, S.-S.; Podolska, M.; D’Amelio, S.; Shao, R.; Gasser, R.B.; Zhu, X.-Q. Mitochondrial Phylogenomics Yields Strongly Supported Hypotheses for Ascaridomorph Nematodes. Sci. Rep. 2016, 6, 39248. [Google Scholar] [CrossRef] [PubMed]
- Quiazon, K.M.A.; Hill-Spanik, K.M.; Denson, M.R.; de Buron, I. Identification and Distribution of Ascaridoid Larvae in Marine Fishes from Inshore and Offshore Waters along the Coast of South Carolina, USA; Grice Marine Lab, College of Charleston: 205 Fort Johnson Road, Charleston, SC, USA, 2017. [Google Scholar]
- Mattiucci, S.; Nascetti, G. Chapter 2 Advances and Trends in the Molecular Systematics of Anisakid Nematodes, with Implications for Their Evolutionary Ecology and Host–Parasite Co-Evolutionary Processes. In Advances in Parasitology; Academic Press: Cambridge, MA, USA, 2008; Volume 66, pp. 47–148. [Google Scholar] [CrossRef]
- Cavallero, S.; Scribano, D.; D’Amelio, S. First Case Report of Invasive Pseudoterranoviasis in Italy. Parasitol. Int. 2016, 65, 488–490. [Google Scholar] [CrossRef] [PubMed]
- Brattey, J.; Stenson, G.B. Host Specificity and Abundance of Parasitic Nematodes (Ascaridoidea) from the Stomachs of Five Phocid Species from Newfoundland and Labrador. Can. J. Zool. 1993, 71, 2156–2166. [Google Scholar] [CrossRef]
- Mattiucci, S.; Paggi, L.; Nascetti, G.; Ishikura, H.; Kikuchi, K.; Sato, N.; Cianchi, R.; Bullini, L. Allozyme and Morphological Identification of Shape Anisakis, Contracaecum and Pseudoterranova from Japanese Waters (Nematoda, Ascaridoidea). Syst. Parasitol. 1998, 40, 81–92. [Google Scholar] [CrossRef]
- Nadler, S.A.; D’Amelio, S.; Dailey, M.D.; Paggi, L.; Siu, S.; Sakanari, J.A. Molecular Phylogenetics and Diagnosis of Anisakis, Pseudoterranova, and Contracaecum from Northern Pacific Marine Mammals. J. Parasitol. 2005, 91, 1413–1429. [Google Scholar] [CrossRef] [Green Version]
- Paggi, L.; Mattiucci, S.; D’Amelio, S.; Nascetti, G. Nematodi Del Genere Anisakis in Pesci, Cefalopodi e Cetacei Del Mar Mediterraneo e Dell’Oceano Atlantico e Pacifico. Biol. Mar. Mediterr. 1998, 5, 1585–1592. [Google Scholar]
- Yamaguti, S.; Arima, S. Porrocaecum azarasi n. sp. (Nematoda) from the Japanese Seal. Trans. Sapporo Nat. Hist. Soc. 1942, 17, 113–116. [Google Scholar]
- George-Nascimento, M.; Urrutia, X. Pseudoterranova cattani sp. nov. (Ascaridoidea: Anisakidae), Un Parásito Del Lobo Marino Común Otaria Byronia De Blainville En Chile. Rev. Chil. Hist. Nat. 2000, 73, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Finley, K.J.; Evans, C. Summer Diet of the Bearded Seal (Erignathus barbatus) in the Canadian High Arctic. Arctic 1983, 36, 82–89. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, C.; Dunlop, E.S.; Opdal, A.F.; Fiksen, Ø. The Evolution of Spawning Migrations: State Dependence and Fishing-induced Changes. Ecology 2008, 89, 3436–3448. [Google Scholar] [CrossRef]
- Morey, G.; Morales-Nin, B.; Riera, F.; Grau, A.; Geffen, A.J.; Pérez-Mayol, S.; Chang, M.-Y.; Grau, A.M. Atlantic Cod Gadus morhua in the Mediterranean: A Surprising Immigrant. Mar. Ecol. Prog. Ser. 2012, 467, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Holmes, J.C. Helminth Communities in Marine Fishes. In Parasite Communities: Patterns and Processes; Springer: Berlin/Heidelberg, Germany, 1990; pp. 101–130. [Google Scholar]
- Alt, K.G.; Kochmann, J.; Klimpel, S.; Cunze, S. Improving Species Distribution Models of Zoonotic Marine Parasites. Sci. Rep. 2019, 9, 9851. [Google Scholar]
- Marcogliese, D.J. Food Webs and the Transmission of Parasites to Marine Fish. Parasitology 2002, 124, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Learmonth, J.A.; MacLeod, C.D.; Santos, M.B.; Pierce, G.J.; Crick, H.; Robinson, R. Potential Effects of Climate Change on Marine Mammals. Oceanogr. Mar. Biol. 2006, 44, 431. [Google Scholar]
- Fiorenza, E.A.; Wendt, C.A.; Dobkowski, K.A.; King, T.L.; Pappaionou, M.; Rabinowitz, P.; Samhouri, J.F.; Wood, C.L. It’sa Wormy World: Meta-analysis Reveals Several Decades of Change in the Global Abundance of the Parasitic Nematodes Anisakis spp. and Pseudoterranova spp. in Marine Fishes and Invertebrates. Glob. Change Biol. 2020, 26, 2854–2866. [Google Scholar] [CrossRef] [PubMed]
- Hermosilla, C.; Hirzmann, J.; Silva, L.; Brotons, J.; Cerdà, M.; Prenger-Berninghoff, E.; Ewers, C.; Taubert, A. Occurrence of Anthropozoonotic Parasitic Infections and Faecal Microbes in Free-Ranging Sperm Whales (Physeter macrocephalus) from the Mediterranean Sea. Parasitol. Res. 2018, 117, 2531–2541. [Google Scholar] [CrossRef]
- Stewardson, C.L.; Fourie, H. Endoparasites of the Cape Fur Seal Arctocephalus Pusillus Pusillus from the Eastern Cape Coast of South Africa. Trans. R. Soc. South. Afr. 1998, 53, 33–51. [Google Scholar] [CrossRef]
- Mattiucci, S.; Paggi, L.; Nascetti, G.; Abollo, E.; Webb, S.; Pascual, S.; Cianchi, R.; Bullini, L. Genetic Divergence and Reproductive Isolation between Anisakis brevispiculata and Anisakis physeteris (Nematoda: Anisakidae) s. Int. J. Parasitol. 2001, 31, 9–14. [Google Scholar] [CrossRef]
- Farjallah, S.; Slimane, B.B.; Busi, M.; Paggi, L.; Amor, N.; Blel, H.; Said, K.; D’Amelio, S. Occurrence and Molecular Identification of Anisakis spp. from the North African Coasts of Mediterranean Sea. Parasitol. Res. 2008, 102, 371–379. [Google Scholar] [CrossRef]
- Palomba, M.; Mattiucci, S.; Crocetta, F.; Osca, D.; Santoro, M. Insights into the Role of Deep-Sea Squids of the Genus Histioteuthis (Histioteuthidae) in the Life Cycle of Ascaridoid Parasites in the Central Mediterranean Sea Waters. Sci. Rep. 2021, 11, 7135. [Google Scholar] [CrossRef]
- Frantzis, A.; Swift, R.; Gillespie, D.; Menhennett, C.; Gordon, J.; Gialinakis, S. Sperm Whale Presence off Southwest Crete, Greece, Eastern Mediterranean. Eur Res. Cet 1999, 13, 214–217. [Google Scholar]
- Shamsi, S.; Ghadam, M.; Suthar, J.; Mousavi, H.E.; Soltani, M.; Mirzargar, S. Occurrence of Ascaridoid Nematodes in Selected Edible Fish from the Persian Gulf and Description of Hysterothylacium Larval Type XV and Hysterothylacium persicum n. sp. (Nematoda: Raphidascarididae). Int. J. Food Microbiol. 2016, 236, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Berland, B. Nematodes from Some Norwegian Marine Fishes. Sarsia 1961, 2, 1–50. [Google Scholar] [CrossRef]
- Baptista-Fernandes, T.; Rodrigues, M.; Castro, I.; Paixão, P.; Pinto-Marques, P.; Roque, L.; Belo, S.; Ferreira, P.M.; Mansinho, K.; Toscano, C. Human Gastric Hyperinfection by Anisakis simplex: A Severe and Unusual Presentation and a Brief Review. Int. J. Infect. Dis. 2017, 64, 38–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasawa, K. The Life Cycle of Anisakis simplex: A Review. Intest. Anisakiasis Jpn. 1990, 31–40. [Google Scholar] [CrossRef]
- Portes Santos, C.; Lodi, L. Occurrence of Anisakis Physeteris Baylis, 1923 and Pseudoterranova sp. (Nematoda) in Pygmy Sperm Whale Kogia Breviceps (De Blainvillei, 1838) (Physeteridae) in Northeastern Coast of Brazil. Mem. Inst. Oswaldo Cruz 1998, 93, 187–188. [Google Scholar] [CrossRef]
- Solís, D.G.; Vidal-Martínez, V.; Antochiw-Alonso, D.; Ortega-Argueta, A. Anisakid Nematodes from Stranded Pygmy Sperm Whales, Kogia breviceps (Kogiidae), in Three Localities of the Yucatan Peninsula, Mexico. J. Parasitol. 2006, 92, 1120–1122. [Google Scholar] [CrossRef]
- Holt, R.D.; Dobson, A.P.; Begon, M.; Bowers, R.G.; Schauber, E.M. Parasite Establishment in Host Communities; Wiley: Hoboken, NJ, USA, 2003. [Google Scholar]
- Langwig, K.E.; Voyles, J.; Wilber, M.Q.; Frick, W.F.; Murray, K.A.; Bolker, B.M.; Collins, J.P.; Cheng, T.L.; Fisher, M.C.; Hoyt, J.R. Context-dependent Conservation Responses to Emerging Wildlife Diseases. Front. Ecol. Environ. 2015, 13, 195–202. [Google Scholar] [CrossRef] [Green Version]
- Karamanlidis, A.A.; Androukaki, E.; Adamantopoulou, S.; Chatzispyrou, A.; Johnson, W.M.; Kotomatas, S.; Papadopoulos, A.; Paravas, V.; Paximadis, G.; Pires, R. Assessing Accidental Entanglement as a Threat to the Mediterranean Monk Seal Monachus monachus. Endanger. Species Res. 2008, 5, 205–213. [Google Scholar] [CrossRef]
- Formigaro, C.; Karamanlidis, A.A.; Dendrinos, P.; Marsili, L.; Silvi, M.; Zaccaroni, A. Trace Element Concentrations in the Mediterranean Monk Seal (Monachus monachus) in the Eastern Mediterranean Sea. Sci. Total Environ. 2017, 576, 528–537. [Google Scholar] [CrossRef]
- Karamanlidis, A.A.; Gaughran, S.; Aguilar, A.; Dendrinos, P.; Huber, D.; Pires, R.; Schultz, J.; Skrbinšek, T.; Amato, G. Shaping Species Conservation Strategies Using MtDNA Analysis: The Case of the Elusive Mediterranean Monk Seal (Monachus monachus). Biol. Conserv. 2016, 193, 71–79. [Google Scholar] [CrossRef]
- Luong, L.; Heath, B.; Polak, M. Host Inbreeding Increases Susceptibility to Ectoparasitism. J. Evol. Biol. 2007, 20, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Gkafas, G.A.; de Jong, M.; Exadactylos, A.; Raga, J.A.; Aznar, F.J.; Hoelzel, A.R. Sex-Specific Impact of Inbreeding on Pathogen Load in the Striped Dolphin. Proc. R. Soc. B 2020, 287, 20200195. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Whitehouse, K.; Petetti, L.; Duignan, P.; Castinel, A. Hookworm Infection, Anaemia and Genetic Variability of the New Zealand Sea Lion. Proc. R. Soc. B Biol. Sci. 2009, 276, 3523–3529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karamanlidis, A.A.; Skrbinšek, T.; Amato, G.; Dendrinos, P.; Gaughran, S.; Kasapidis, P.; Kopatz, A.; Stronen, A.V. Genetic and Demographic History Define a Conservation Strategy for Earth’s Most Endangered Pinniped, the Mediterranean Monk Seal Monachus monachus. Sci. Rep. 2021, 11, 373. [Google Scholar]
- Harvell, D.; Altizer, S.; Cattadori, I.M.; Harrington, L.; Weil, E. Climate Change and Wildlife Diseases: When Does the Host Matter the Most? Ecology 2009, 90, 912–920. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Classification | GenBank Accession Numbers | Reference |
|---|---|---|
| Pseudoterranova decipiens | AJ891148 and AJ891149 | [52] |
| Pseudoterranova cattani | AJ891143 and MT941431 | [52,53] |
| Pseudoterranova azarasi | AJ891139 | |
| LC312195 | [54] | |
| Pseudoterranova krabbei | AJ891152 and AJ891151 | [52] |
| Pseudoterranova bulbosa | NC031643 and KU558720 | [55] |
| Pseudoterranova spp. | OM037608 | this study |
| Anisakis physeteris | MF663249 | [56] |
| A. Physeteris_MF663249 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| P. bulbosa_NC_031643 | 0.156 | ||||||||||
| P. bulbosa_KU558720 | 0.156 | 0.000 | |||||||||
| P. decipiens_AJ891148 | 2.682 | 1.994 | 1.994 | ||||||||
| P. decipiens_AJ891149 | 2.937 | 2.093 | 2.093 | 0.016 | |||||||
| P. cattani_AJ891143 | 2.600 | 1.836 | 1.836 | 0.112 | 0.119 | ||||||
| P. cattani_MT941431 | 2.378 | 1.885 | 1.885 | 0.113 | 0.120 | 0.016 | |||||
| P. azarasi_AJ891139 | 3.088 | 2.058 | 2.058 | 0.138 | 0.142 | 0.070 | 0.077 | ||||
| P. azarasi_LC312195 | 2.006 | 1.580 | 1.580 | 0.078 | 0.085 | 0.063 | 0.057 | 0.012 | |||
| P. krabbei_AJ891152 | 2.677 | 1.844 | 1.844 | 0.120 | 0.124 | 0.064 | 0.071 | 0.061 | 0.051 | ||
| P. krabbei_AJ891151 | 2.677 | 1.726 | 1.726 | 0.109 | 0.120 | 0.049 | 0.055 | 0.055 | 0.044 | 0.016 | |
| Pseudoterranova spp. | 0.162 | 0.042 | 0.042 | 1.994 | 2.093 | 1.880 | 1.913 | 2.058 | 1.505 | 1.844 | 1.726 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koitsanou, E.; Sarantopoulou, J.; Komnenou, A.; Exadactylos, A.; Dendrinos, P.; Papadopoulos, E.; Gkafas, G.A. First Report of the Parasitic Nematode Pseudoterranova spp. Found in Mediterranean Monk Seal (Monachus monachus) in Greece: Conservation Implications. Conservation 2022, 2, 122-133. https://0-doi-org.brum.beds.ac.uk/10.3390/conservation2010010
Koitsanou E, Sarantopoulou J, Komnenou A, Exadactylos A, Dendrinos P, Papadopoulos E, Gkafas GA. First Report of the Parasitic Nematode Pseudoterranova spp. Found in Mediterranean Monk Seal (Monachus monachus) in Greece: Conservation Implications. Conservation. 2022; 2(1):122-133. https://0-doi-org.brum.beds.ac.uk/10.3390/conservation2010010
Chicago/Turabian StyleKoitsanou, Efstathia, Joanne Sarantopoulou, Anastasia Komnenou, Athanasios Exadactylos, Panagiotis Dendrinos, Elias Papadopoulos, and Georgios A. Gkafas. 2022. "First Report of the Parasitic Nematode Pseudoterranova spp. Found in Mediterranean Monk Seal (Monachus monachus) in Greece: Conservation Implications" Conservation 2, no. 1: 122-133. https://0-doi-org.brum.beds.ac.uk/10.3390/conservation2010010