Effect of Type I Antifreeze Proteins on the Freezing and Melting Processes of Cryoprotective Solutions Studied by Site-Directed Spin Labeling Technique


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Syntheses of the Spin Labeled HPLC6 Peptides
2.3. EPR Experiments and Line-Shape Simulations
2.4. Cryo-Photo Microscopic Method
3. Results and Discussion
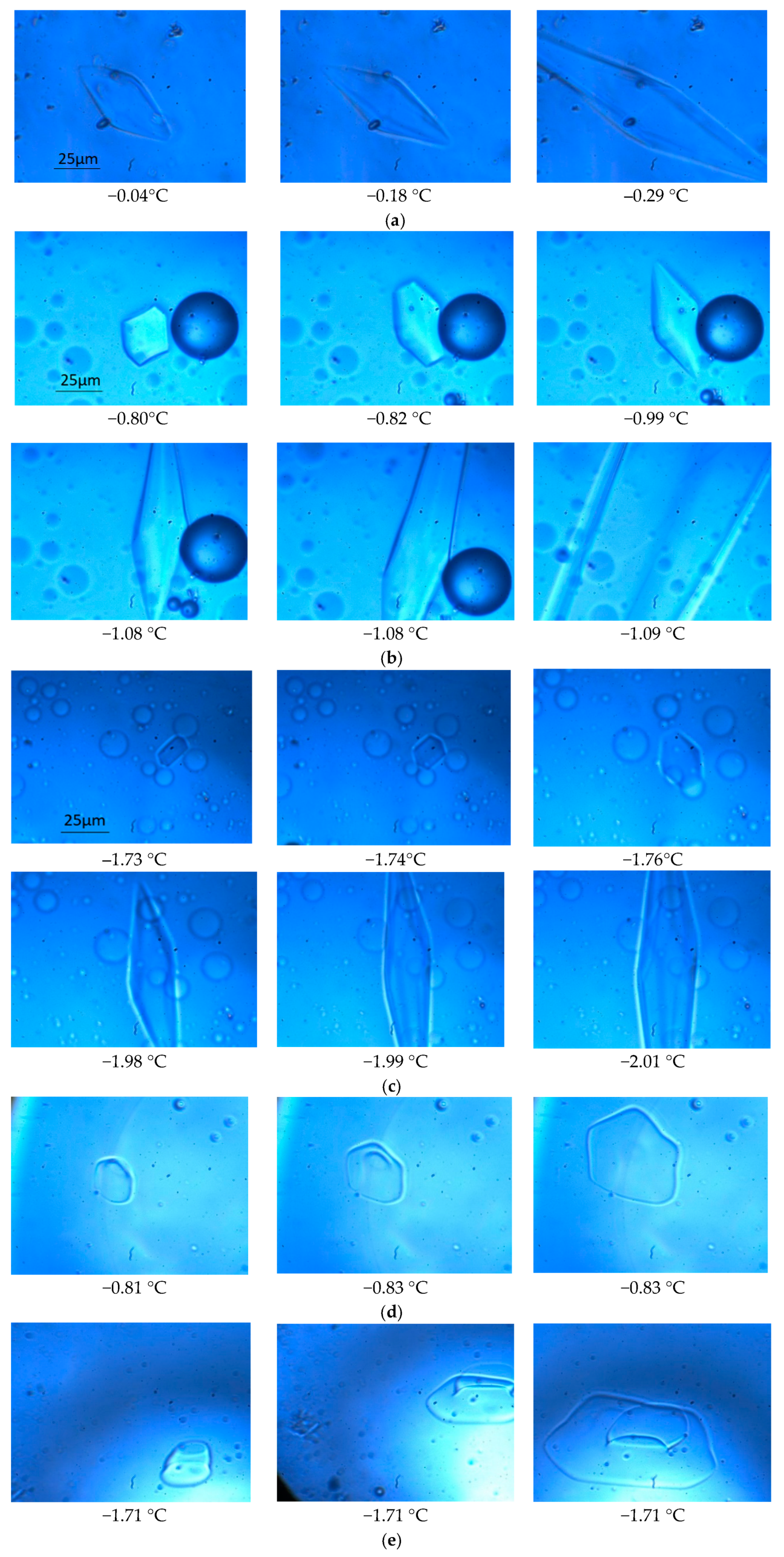
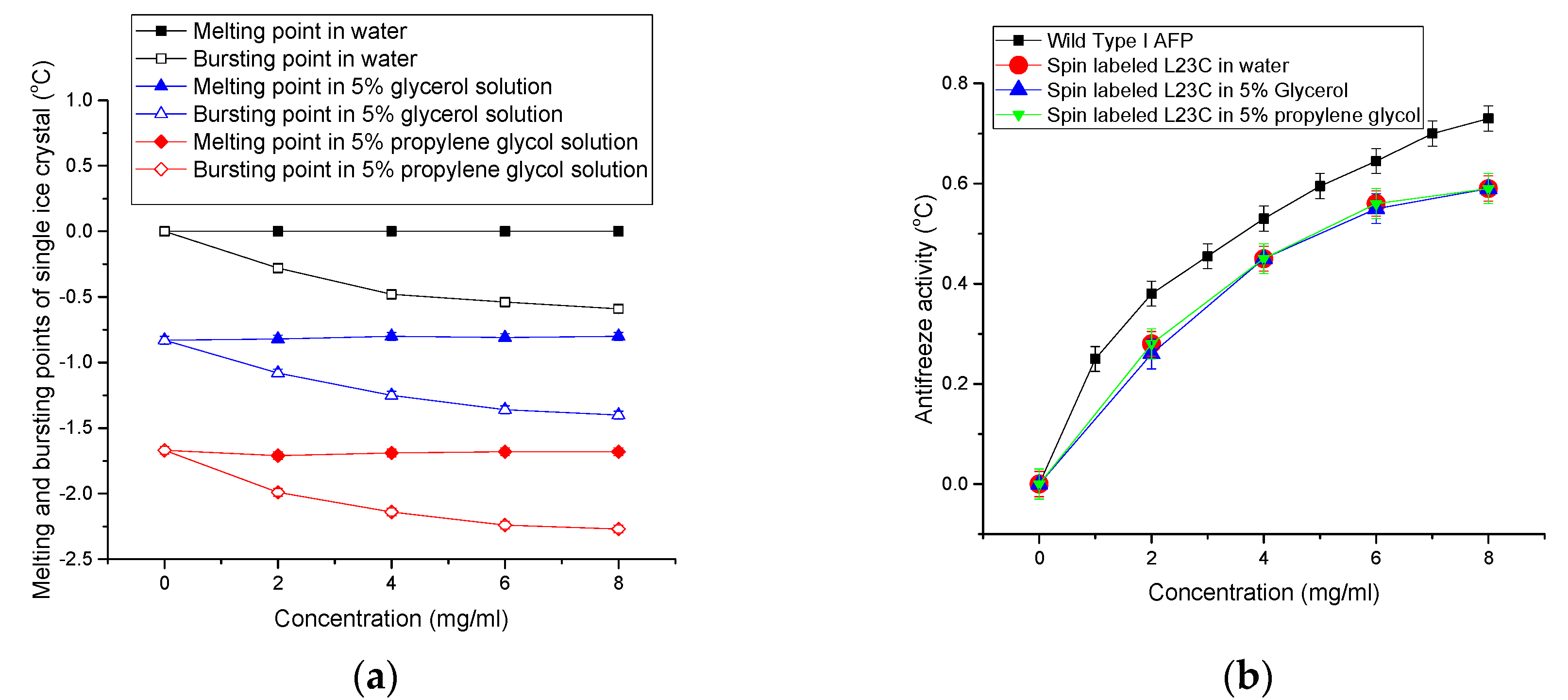
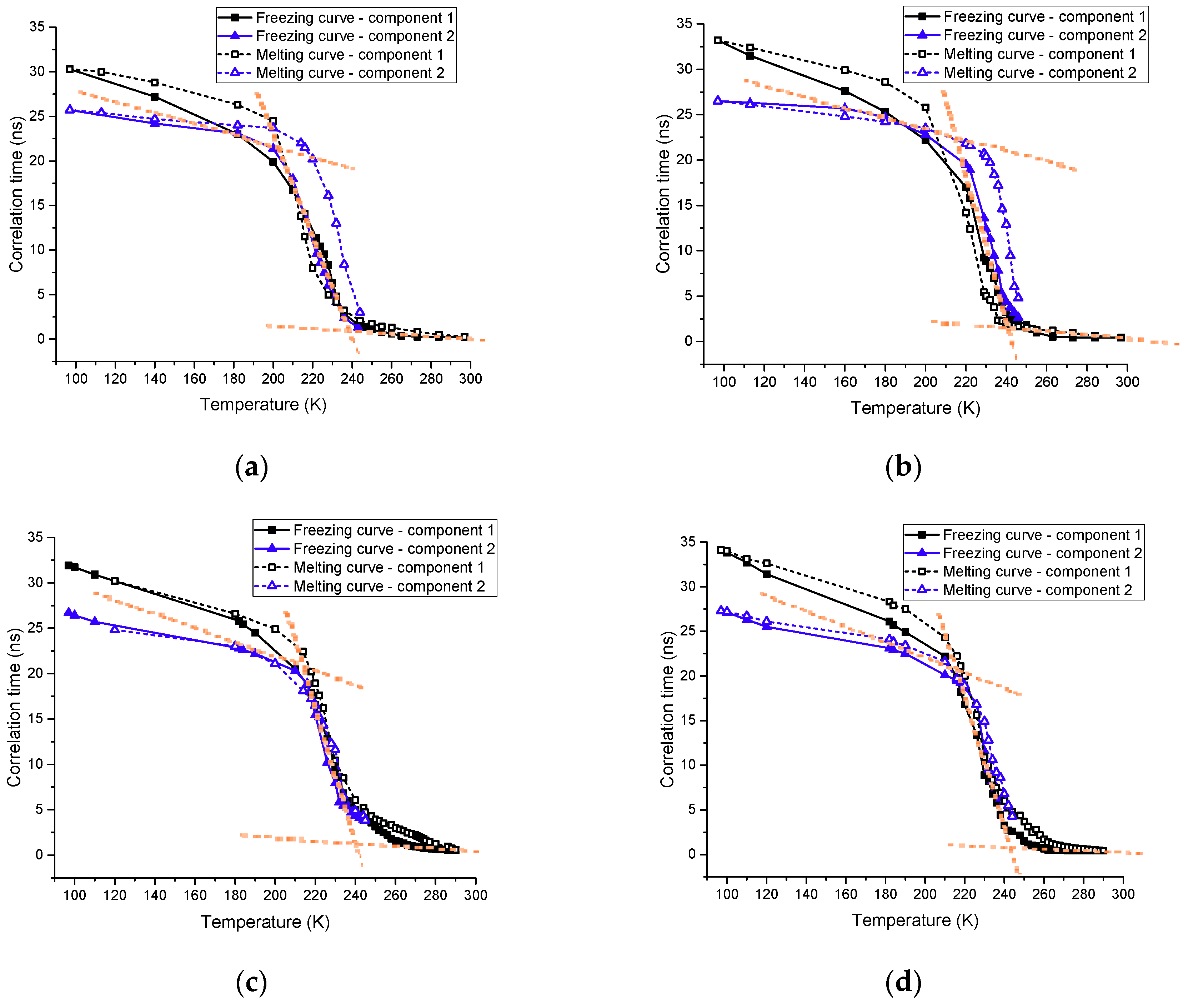
3.1. The Growths of Ice Crystals in the Spin-Labeled L23C and A17C CPA Solutions
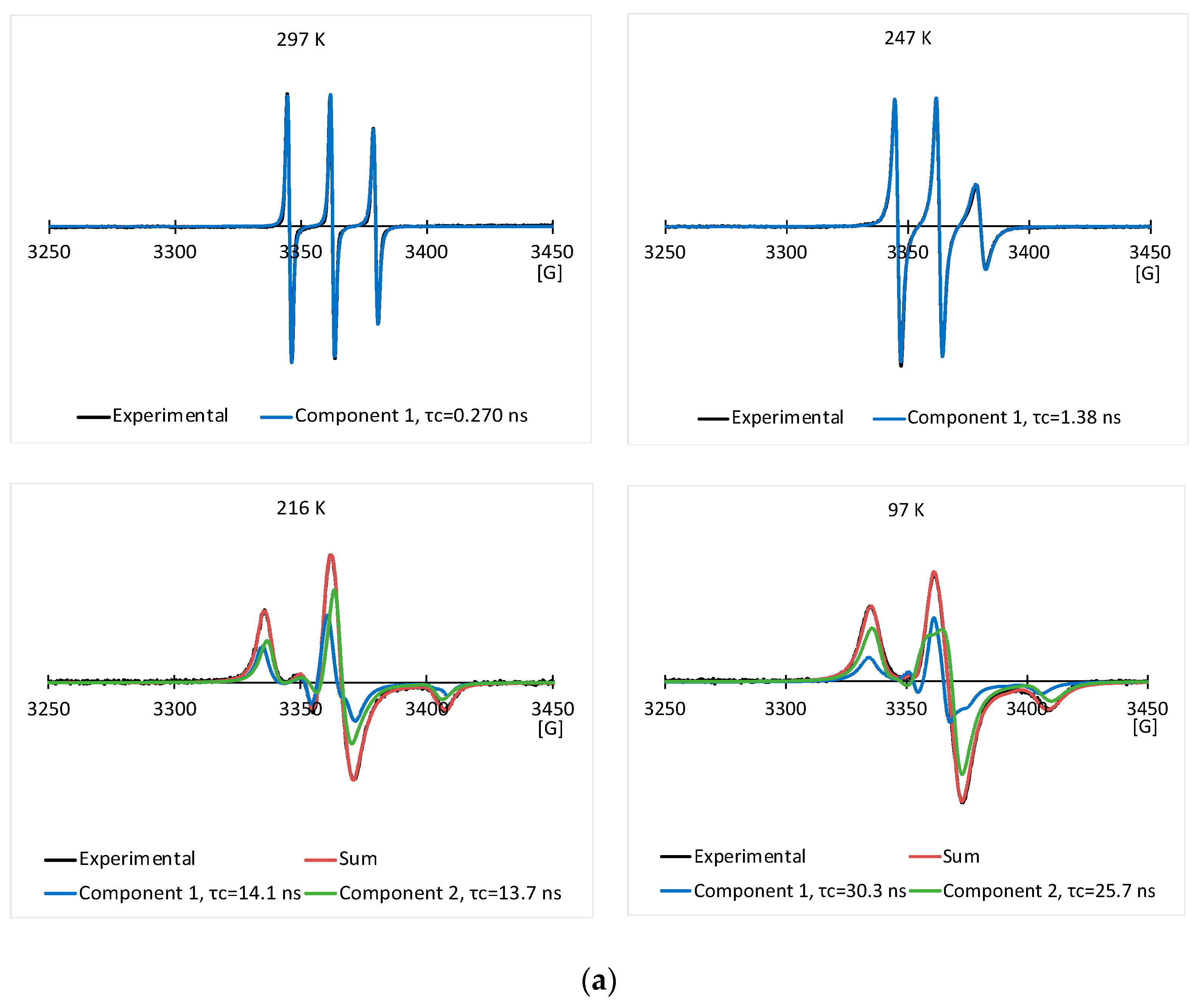
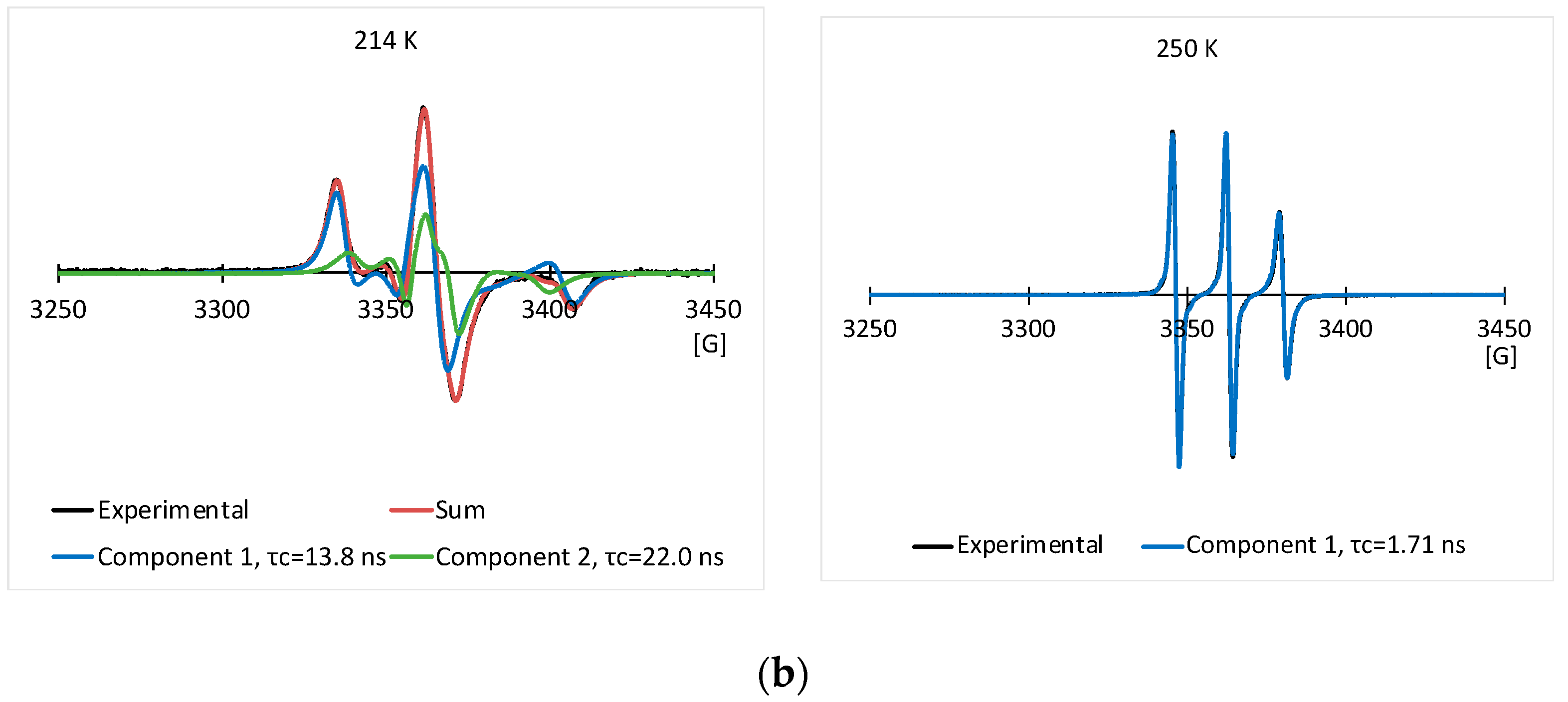
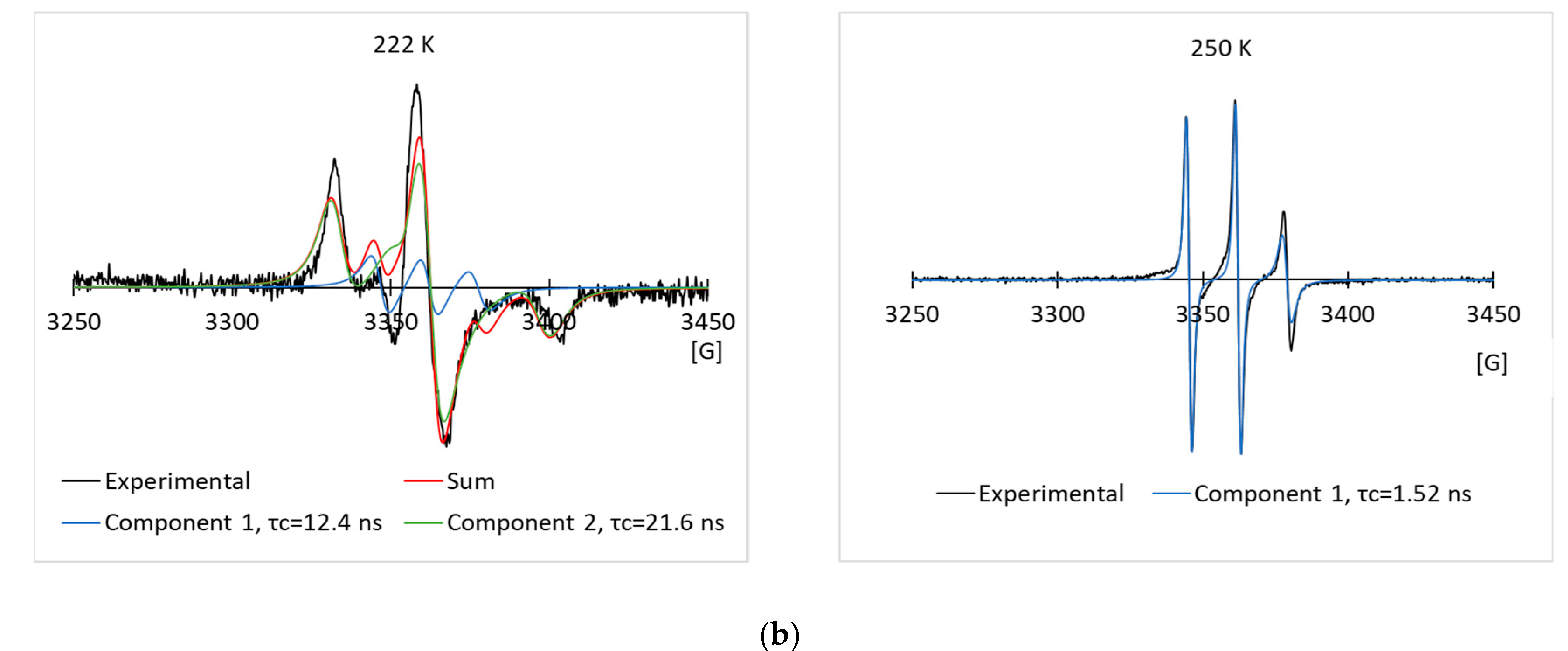
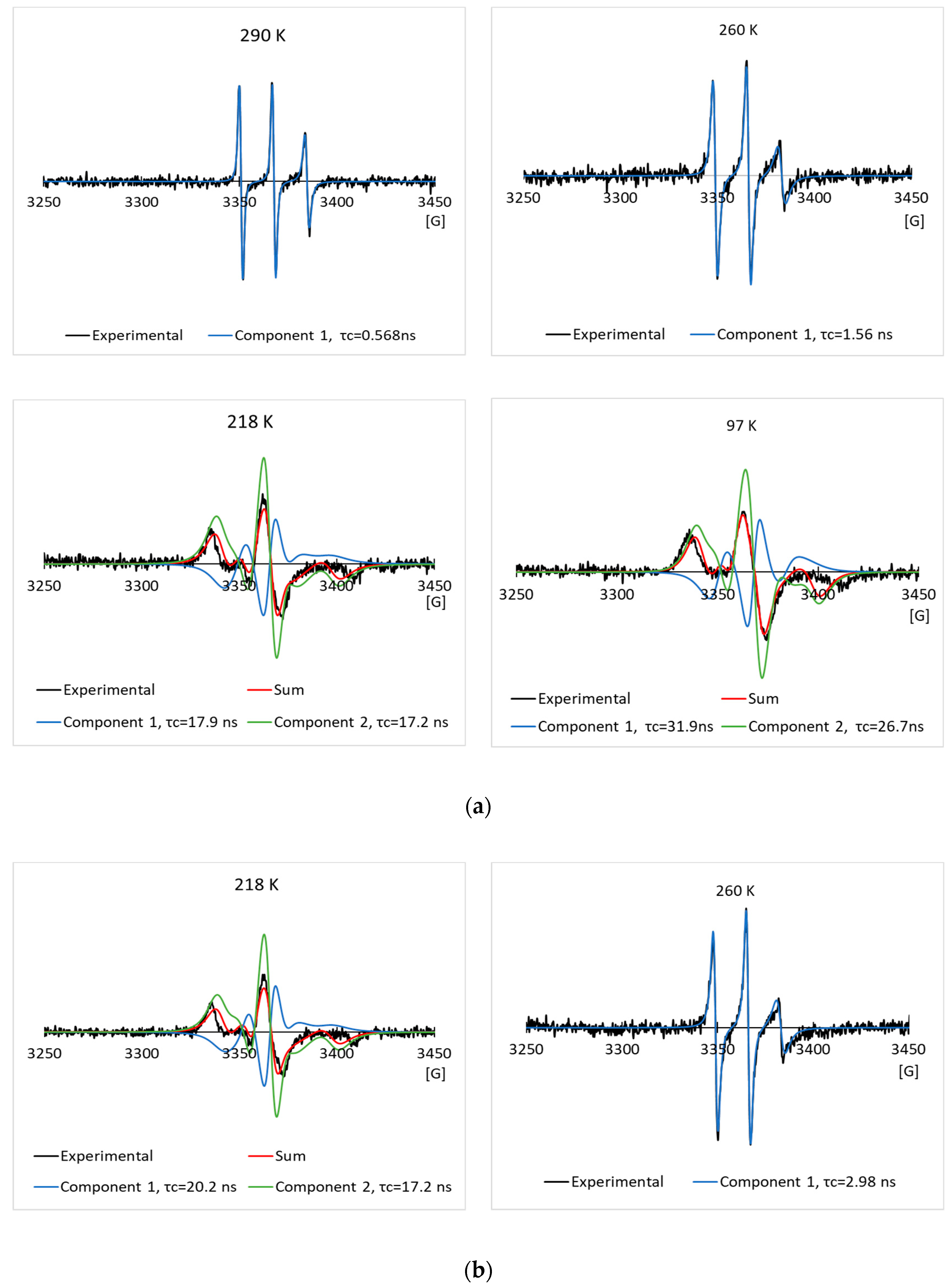
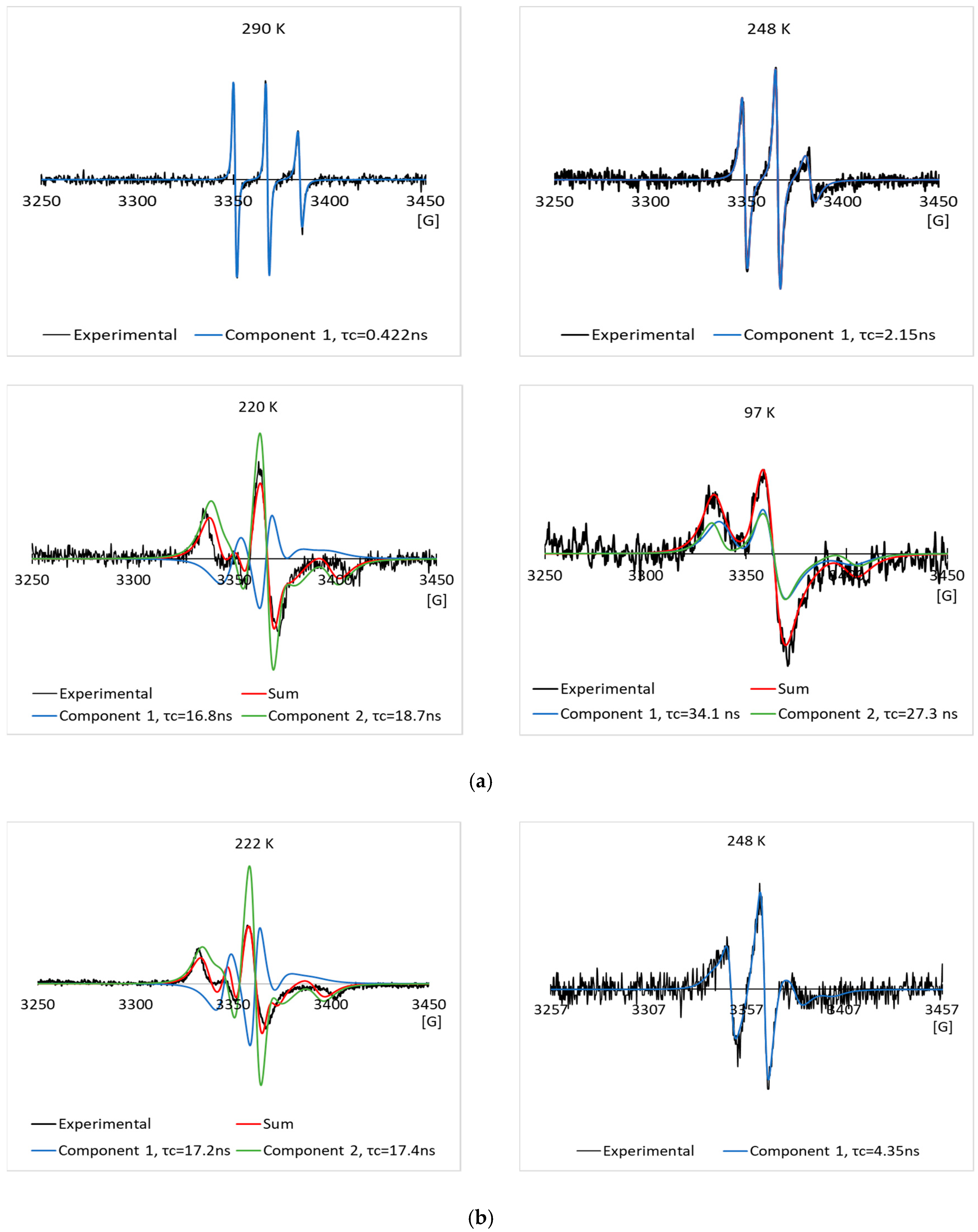
3.2. EPR Results of the Spin Labeled Type I AFP in the CPA Solutions
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- DeVries, A.L.; Komatsu, S.K.; Feeney, R.E. Chemical and physical properties of freezing point-depressing glycolproteins from Antarctic fihes. J. Biol. Chem. 1970, 245, 2901–2908. [Google Scholar] [PubMed]
- DeVries, A.L. Antifreeze peptides and glycopeptides in cold-water fishes. Annu. Rev. Physiol. 1983, 45, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Duman, J.G.; DeVries, A.L. Freezing behavior of aqueous solutions of glycoproteins from the blood of an Antarctic fish. Cryobiology 1972, 9, 469–472. [Google Scholar] [CrossRef]
- DeVries, A.L.; Wohlschlag, D.E. Freezing resistance in some Antarctic fishes. Science 1969, 163, 1073–1075. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, S.; DeVries, A.L.; Feeney, R.E. Studies of the structure of freezing point-depressing glycoproteins from an Antarctic fish. J. Biol. Chem. 1970, 245, 2909–2913. [Google Scholar] [PubMed]
- Feeney, R.E. A biological antifreeze. Am. Sci. 1974, 62, 712–719. [Google Scholar] [PubMed]
- Feeney, R.E.; Yeh, Y. Antifreeze proteins from fish bloods. Adv. Protein. Chem. 1978, 32, 191–282. [Google Scholar] [PubMed]
- Davies, P.L.; Hew, C.L. Biochemistry of fish antifreeze proteins. FASEB J. 1990, 4, 2460–2468. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.H.; DeVries, A.L. Structures of antifreeze peptides from the antarctic eel pout, Austrolycicthys brachycephalus. Biochim. Biophys. Acta 1989, 997, 55–64. [Google Scholar] [CrossRef]
- Duman, J.G.; Wu, D.W.; Olsen, T.M.; Urrutia, M.; Turaman, D. Thermal-Hysteresis Proteins. Adv. Low Temp. Biol. 1993, 2, 131. [Google Scholar]
- Knight, C.A.; DeVries, A.L.; Oolman, L.D. Fish antifreeze protein and the freezing and recrystallization of ice. Nature 1984, 308, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Duman, J.G.; DeVries, A.L. Freezing resistance in winter flounder Pseudopleuronectus americanus. Nature 1974, 247, 237–238. [Google Scholar] [CrossRef]
- Duman, J.G.; DeVries, A.L. Isolation, characterization and physical properties of protein antifreeze from the winter flounder Pseudopleunectus americanus. Comp. Biochem. Physiol. 1976, 54, 375–380. [Google Scholar]
- Hew, C.L.; Joshi, S.; Wang, N.-C.; Kao, M.-H.; Ananthanarayanan, V.S. Structure of shorthorn sculpin antifreeze polypeptides. Eur. J. Biochem. 1985, 151, 167–172. [Google Scholar] [CrossRef]
- Sönnichsen, F.D.; Davies, P.L.; Sykes, B.D. NMR structural studies on antifreeze proteins. Biochem. Cell Biol. 1998, 76, 284–293. [Google Scholar] [CrossRef]
- Harding, M.M.; Ward, L.G.; Haymet, A.D. Type I ‘antifreeze’ proteins. Structure-activity studies and mechanisms of ice growth inhibition. Eur. J. Biochem. 1999, 264, 653–665. [Google Scholar] [CrossRef] [PubMed]
- Mao, Y.; Lin, W.; Ba, Y. Antifreeze protein NMR sensor to detect water molecular reorientation in the surface of ice. J. Chem. Phys. 2009, 131, 101102. [Google Scholar] [CrossRef] [Green Version]
- Mao, Y.; Jeong, M.; Wang, T.; Ba, Y. Threonine side chain conformational population distribution of a type I antifreeze protein on interacting with ice surface studied via 13C–15N dynamic REDOR NMR. Solid State Nucl. Magn. Reson. 2011, 39, 7–13. [Google Scholar] [CrossRef]
- Mao, Y.; Ba, Y. Insight into the Binding of Antifreeze Proteins to Ice Surfaces via 13C Spin Lattice Relaxation Solid State NMR. Biophys. J. 2006, 91, 1059–1068. [Google Scholar] [CrossRef]
- Sicheri, F.; Yang, D.S. Ice-binding structure and mechanism of an antifreeze protein from winter flounder. Nature 1995, 375, 427–431. [Google Scholar] [CrossRef]
- Yang, D.S.; Sax, M.; Chakrabartty, A.; Hew, C.L. Crystal structure of an antifreeze polypeptide and its mechanistic implications. Nature 1988, 333, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Sicheri, F.; Yang, D.S.C. Structure determination of a lone a-helical antifreeze protein from winter flounder. Acta Cryst. D 1996, 52, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Gronwald, W.; Chao, H.; Reddy, D.V.; Davies, P.L.; Sykes, B.D.; Sönnichsen, F.D. NMR characterization of side chain flexibility and backbone structure in the type I antifreeze protein at near freezing temperatures. Biochemistry 1996, 35, 16698–16704. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.A.; Cheng, C.C.; DeVries, A.L. Adsorption of alpha-helical antifreeze peptides on specific ice crystal surface planes. Biophys. J. 1991, 59, 409–418. [Google Scholar] [CrossRef] [Green Version]
- Wen, D.; Laursen, R.A. A model for binding of an antifreeze polypeptide to ice. Biophys. J. 1992, 63, 1659–1662. [Google Scholar] [CrossRef] [Green Version]
- Chao, H.; DeLuca, C.I.; Davies, P.L. Mixing antifreeze protein types changes ice crystal morphology without affecting antifreeze activity. FEBS Lett. 1995, 357, 183–186. [Google Scholar] [Green Version]
- Haymet, A.D.J.; Ward, L.G.; Harding, M.M. Winter Flounder “Antifreeze” Proteins: Synthesis and Ice Growth Inhibition of Analogues that Probe the Relative Importance of Hydrophobic and Hydrogen-Bonding Interactions. J. Am. Chem. Soc. 1999, 121, 941–948. [Google Scholar] [CrossRef]
- Baardsnes, J.; Kondejewski, L.H.; Hodges, R.S.; Chao, H.; Kay, C.; Davies, P.L. New ice-binding face for type I antifreeze protein. FEBS Lett. 1999, 463, 87–91. [Google Scholar] [CrossRef]
- Chao, H.; Houston, M.E.; Hodges, R.S.; Kay, C.M.; Sykes, B.D.; Loewen, M.C.; Davies, P.L.; Sönnichsen, F.D.; et al. A diminished role for hydrogen bonds in antifreeze protein binding to ice. Biochemistry 1997, 36, 14652–14660. [Google Scholar] [CrossRef]
- Haymet, A.D.; Ward, L.G.; Harding, M.M.; Knight, C.A. Valine substituted winter flounder ‘antifreeze’: preservation of ice growth hysteresis. FEBS Lett. 1998, 430, 301–306. [Google Scholar] [CrossRef]
- Zhang, W.; Laursen, R.A. Structure-function relationships in a type I antifreeze polypeptide. The role of threonine methyl and hydroxyl groups in antifreeze activity. J. Biol. Chem. 1998, 273, 34806–34812. [Google Scholar] [CrossRef] [PubMed]
- Haymet, A.D.; Ward, L.G.; Harding, M.M. Hydrophobic analogues of the winter flounder ‘antifreeze’ protein. FEBS Lett. 2001, 491, 285–288. [Google Scholar] [CrossRef]
- Loewen, M.C.; Chao, H.; Houston, M.E.; Baardsnes, J.; Hodges, R.S.; Kay, C.M.; Sykes, B.D.; Sönnichsen, F.D.; Davies, P.L. Alternative roles for putative ice-binding residues in type I antifreeze protein. Biochemistry 1999, 38, 4743–4749. [Google Scholar] [CrossRef] [PubMed]
- Flores, A.; Quon, J.C.; Perez, A.F.; Ba, Y. Mechanisms of Antifreeze Proteins Investigated via the Site-Directed Spin Labeling Technique. Eur. Biophys. J. 2018, 47, 611–630. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A. Glycerol Is a Colligative Antifreeze in Some Northern Fishes. J. Exp. Zool. 1992, 262, 347–352. [Google Scholar] [CrossRef]
- Rubinsky, B.; Arav, A.; Hong, J.-S.; Lee, C.Y. Freezing of Mammalian Livers with Glycerol and Antifreeze Proteins. Biochem. Biophys. Res. Commun. 1994, 200, 732–741. [Google Scholar] [CrossRef]
- Guiberta, E.E.; Petrenkob, A.Y.; Balabana, C.L.; Somovb, A.Y.; Rodrigueza, J.V.; Fullerc, B.J. Organ Preservation: Current Concepts and New Strategies for the Next Decade. Transfus. Med. Hemotherapy 2011, 38, 125–142. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, S.; Prajapati, B.G. A Review on Cryoprotectant and its Modern Implication in Cryonics. Asian J. Pharm. 2016, 10, 154–159. [Google Scholar]
- Wood, D.L. Antifreeze Compositions. U.S. Patent 4,455,248, 19 June 1984. [Google Scholar]
- Toxicological Profile for Propylene Glycol; Agency for Toxic Substances and Disease Registry (ATSDR): Atlanta, GA, USA, 1997; pp. 90–94.
- Hubbell, W.L.; Gross, A.; Langen, R.; Lietzow, M.A. Recent advances in site-directed spin labeling of proteins. Curr. Opin. Struct. Biol. 1998, 8, 649–656. [Google Scholar] [CrossRef]
- Hubbell, W.L.; Altenbach, C. Site-directed spin labeling of membrane proteins. Methods Physiol. Ser. 1994, 1, 224–248. [Google Scholar]
- Hubbell, W.L.; Altenbach, C.; Langen, R.; Oh, K.J.; Cascio, D.; Columbus, L.; Hideg, K.; Cai, K.; Klein-Seetharaman, J.; Kim, J.; et al. Nitroxide’s View of Membrane Protein Structure and Conformational Switching; American Chemical Society: Washington, NY, USA, 2000; p. PHYS-454. [Google Scholar]
- Galiano, L.; Blackburn, M.E.; Veloro, A.M.; Bonora, M.; Fanucci, G.E. Solute Effects on Spin Labels at an Aqueous-Exposed Site in the Flap Region of HIV-1 Protease. J. Phys. Chem. B. 2009, 113, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Steinhoff, H.-J.; Suess, B. Molecular mechanisms of gene regulation studied by site-directed spin labeling. Methods 2003, 29, 188–195. [Google Scholar] [CrossRef]
- Longhi, S.; Belle, V.; Fournel, A.; Guigliarelli, B.; Carriére, F. Probing structural transitions in both structured and disordered proteins using site-directed spin-labeling EPR spectroscopy. J. Pept. Sci. 2011, 17, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Štrancar, J.; Kavalenka, A.; Urbančič, I.; Ljubetič, A.; Hemminga, M.A. SDSL-ESR-based protein structure characterization. Eur. Biophys. J. 2010, 39, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Klare, J.P.; Steinhoff, H.-J. Spin labeling EPR. Photosynth Res. 2009, 102, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Hideg, K.; Hankovszky, O.H. Chemistry of Spin-labeled Amino Acids and Peptides. In Spin Labeling Theory and Applications; Berliner, L.J., Reuben, J., Eds.; Plenum Press: New York, NY, USA; London, UK, 1989; pp. 427–487. [Google Scholar]
- Chao, H.; Sönnichsen, F.D.; DeLuca, C.I.; Sykes, B.D.; Davies, P.L. Structure-function relationship in the globular type III antifreeze protein: identification of a cluster of surface residues required for binding to ice. Protein Sci. 1994, 3, 1760–1769. [Google Scholar] [CrossRef]
- DeLuca, C.I.; Chao, H.; Sonnichsen, F.D.; Sykes, B.D.; Davies, P.L. Effect of type III antifreeze protein dilution and mutation on the growth inhibition of ice. Biophys. J. 1996, 71, 2346–2355. [Google Scholar] [CrossRef] [Green Version]
- Deluca, C.I.; Davies, P.L.; Ye, Q.; Jia, Z. The effects of steric mutations on the structure of type III antifreeze protein and its interaction with ice. J. Mol. Biol. 1998, 275, 515–525. [Google Scholar] [CrossRef]
- Graether, S.P.; DeLuca, C.I.; Baardsnes, J.; Hill, G.A.; Davies, P.L.; Jia, Z. Quantitative and qualitative analysis of type III antifreeze protein structure and function. J. Biol. Chem. 1999, 274, 11842–11847. [Google Scholar] [CrossRef]
- Altenbach, C. Multi-Component EPR Fitting Version 742, LabVIEW Software: Austin, TX, USA.
- Takacs, I.M.; Mot, A.; Silaghi-Dumitrescu, R.; Damian, G. Hemoglobin Side Chains by Spin Labeled EPR Spectroscopy. Studia UBB Phys. 2013, 58, 49–58. [Google Scholar]
- Couto, S.G.; Nonato, M.C.; Costa-Filho, A.J. Site directed spin labeling studies of Escherichia coli dihydroorotate dehydrogenase N-terminal extension. Biochem. Biophys. Res. Commun. 2011, 414, 487–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, Y.; Ba, Y. Ice-Surface Adsorption Enhanced Colligative Effect of Antifreeze Proteins in Ice Growth Inhibition. J. Chem. Phys. 2006, 125, 091102. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perez, A.F.; Taing, K.R.; Quon, J.C.; Flores, A.; Ba, Y. Effect of Type I Antifreeze Proteins on the Freezing and Melting Processes of Cryoprotective Solutions Studied by Site-Directed Spin Labeling Technique. Crystals 2019, 9, 352. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst9070352
Perez AF, Taing KR, Quon JC, Flores A, Ba Y. Effect of Type I Antifreeze Proteins on the Freezing and Melting Processes of Cryoprotective Solutions Studied by Site-Directed Spin Labeling Technique. Crystals. 2019; 9(7):352. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst9070352
Chicago/Turabian StylePerez, Adiel F., Kyle R. Taing, Justin C. Quon, Antonia Flores, and Yong Ba. 2019. "Effect of Type I Antifreeze Proteins on the Freezing and Melting Processes of Cryoprotective Solutions Studied by Site-Directed Spin Labeling Technique" Crystals 9, no. 7: 352. https://0-doi-org.brum.beds.ac.uk/10.3390/cryst9070352