
Biogeography of Iberian Ants (Hymenoptera: Formicidae)


Department of Zoology, University of Granada, Campus de Fuentenueva s/n, 18071 Granada, Spain
*
Author to whom correspondence should be addressed.
Diversity 2021, 13(2), 88; https://0-doi-org.brum.beds.ac.uk/10.3390/d13020088
Submission received: 6 December 2020
/
Revised: 10 February 2021
/
Accepted: 12 February 2021
/
Published: 19 February 2021
(This article belongs to the Special Issue Diversity, Biogeography and Community Ecology of Ants)
Abstract
:Ants are highly diverse in the Iberian Peninsula (IP), both in species richness (299 cited species) and in number of endemic species (72). The Iberian ant fauna is one of the richest in the broader Mediterranean region, it is similar to the Balkan Peninsula but lower than Greece or Israel, when species richness is controlled by the surface area. In this first general study on the biogeography of Iberian ants, we propose seven chorological categories for grouping thems. Moreover, we also propose eight biogeographic refugium areas, based on the criteria of “refugia-within-refugium” in the IP. We analysed species richness, occurrence and endemism in all these refugium areas, which we found to be significantly different as far as ant similarity was concerned. Finally, we collected published evidence of biological traits, molecular phylogenies, fossil deposits and geological processes to be able to infer the most probable centre of origin and dispersal routes followed for the most noteworthy ants in the IP. As a result, we have divided the Iberian myrmecofauna into four biogeographical groups: relict, Asian-IP disjunct, Baetic-Rifan and Alpine. To sum up, our results support biogeography as being a significant factor for determining the current structure of ant communities, especially in the very complex and heterogenous IP. Moreover, the taxonomic diversity and distribution patterns we describe in this study highlight the utility of Iberian ants for understanding the complex evolutionary history and biogeography of the Iberian Peninsula.
1. Introduction
Ants bring together a number of traits that make up an interesting subject for biogeographical studies: they are conspicuous and one of the most well-known taxa of the terrestrial invertebrates [1], near-ubiquitous in terrestrial ecosystems [2]. They are an ecologically dominant faunal group [3] and highly diverse but still accessible [3] and show an extreme diversity of dispersal strategies although they are sessile superorganisms [4].
Local ant species richness is strongly correlated with temperature, which is the most important factor determining the structure of ant communities [1]. Ants are highly diverse in hot and dry habitats [5] and in low-elevation, low-latitude forests [6]. The latitudinal diversity gradient [6] with an asymmetric northern hemispheric pattern has been detected for ants, which is impossible to explain by only contemporary biotic and abiotic variables [5]. None of the current factors sufficiently correlate to explain current ant diversity [2,5,7]. Moreover, modern ant assemblages suggest a climate-driven past reorganization of the Palaearctic ant fauna [7]. Many ant lineages which previously lived in a warmer Europe, were not able to survive through long-term cooling (Pliocene) and glacial cycles (Pleistocene) but some of them persisted in Indomalayan and Australian regions [7]. Thus, the high extinction pattern in the northern hemisphere appears to have conditioned its marked asymmetric latitudinal diversity gradient [5,7].
Knowledge of the ants found in the Iberian Peninsula (the IP including Andorra, Gibraltar, Portugal and Spain) is still fragmentary [8]. Moreover, there are very few publications treating data regarding their biogeography, and they are limited to some genera, species or zones [8,9,10,11,12,13]. The biogeography or philogeography of other Iberian arthropods have been addressed for example for the coleopteran genera Berberomeloe [14], Pimelia [15], Blaps [16], Cephalota [17], Hydraena [18] and some others cited throughout this article.
The IP forms part of the Mediterranean biodiversity hotspot, unique in the Palaearctic region. It is a threatened area with elevated species richness and a high level of endemism in all the taxa [19,20,21]. Hewitt [22] established that the IP has an intermediate position as far as species richness and the number of paleoendemic species are concerned, compared to the other two Mediterranean peninsulas (Italy and the Balkans). This high level of diversity within the IP was attributed mainly to its environmental heterogeneity conditioned by its complex orography. The IP mountain ranges are at different latitudes and altitudes with an east-western orientation. These ranges are sometimes permeable, but others act as geographic barriers producing isolated valleys, plateaus and plains. The east-western orientation also permits the establishment of particular and markedly different microhabitats on the southern and northern slopes, which vary depending on the altitude and latitude.
On the other hand, changing paleogeography and paleoclimate in the region favoured the convergence of different lineages, sharing evolutionary history for a long period [23]. Thus, in the early Paleogene (60 mya), the small Iberian plate, fragmented during the Mesozoic (100 mya) and united with the European plate, had a semitropical climate until the end of the Miocene (23-5.3 mya) [24,25] defining similar biomes to the current African savannah. Moreover, in the southern territories a succession of separations and unifications with the African plate continued until the end of the Miocene, closing the IP with Africa during the evaporitic Messiniense [26]. These semi-tropical communities from the Miocene were composed of species of Asiatic and African origin [27]. Later, successive climatic and orographic changes in the Pliocene and Pleistocene produced migrations towards the IP [25,28,29,30] which led to the formation of a mosaic of fragmented populations that persisted in small refugia throughout the entire Iberian refugium (the “refugia-within-refugium” scenario) [22,31,32,33] which is very important for the understanding of the diversity and phylogeography of some Iberian taxa [31,32,34]. After the glacial periods the IP played an important role when the Palaearctic peninsulas acted as providers of re-colonizing species to the northern territories [35,36,37]. These relatively recent biogeographical processes have greatly conditioned the current faunistic composition of the IP.
Thus, to the best of our knowledge, this is the first biogeographic global study on the richness and distribution of the Iberian formicids. We have undertaken this challenge well aware that the knowledge about ant distribution and endemic species in the IP is still incomplete. Our goals have been (1) to analyse the number of ant genera and species in the IP but also to compare them with other Mediterranean countries, especially France (linking the IP with the European Palaearctic) and Morocco (with the African Palaearctic), (2) to take into account the putative refugia within the Iberian refugium, and we propose a subdivision of the IP in refugium areas and attribute a chorological category to each species, and (3) we have defined four different groups of species depending on the different dispersal patterns (centre of origin and dispersal routes) based on biological traits, molecular phylogenies, fossil deposits and geological processes, when available.
2. Materials and Methods
2.1. Species Included in the Study
In this study we have included known species in the IP (Andorra, Gibraltar, Portugal and Spain) and compared them with the species found in nearby countries such as France and Morocco. We have tried to use the most updated published lists in the four countries closely involved adding new citations such as some general reviews [38,39,40]. Only described and named species are included in our study (those species appearing as “sp.”, species 1, 2, species under description, etc. are not included). Moreover, we have only considered mainland species, avoiding the species which only appear on the islands belonging to the four countries. In general, we have not undertaken taxonomic problems such as synonyms or misidentifications of species included in the different faunistic lists, assuming as correct the most recent reference. For the nomenclature of the social parasite species of Temnothorax and Tetramorium we have adopted the criterion of Seifert et al. [41]. With this method of managing citations our final results might show slight differences with some published lists, but this has not affected our main goals.
For the Portuguese species we focused on Salgueiro [42], taking into account previous publications of Collingwood & Prince [43] and later ones such as Boeiro et al. [44] and Gonçalves et al. [45]. We have excluded from our lists Temnothorax caparica Henin, Paiva and Collingwood, 2001, which resulted to be a misidentification of Cardiocondyla mauritanica Forel, 1890 [46].
For the Spanish species we focused on the list included in Sánchez-García et al. [47] and added some recently described species [48] and some new findings [49].
In the case of Andorra we have used the list of species of Bernadou et al. [50]. For Gibraltar there is no up-to-date catalogue and citations appear in general articles for the Iberian ants, except for some recent records that have been taken into account [51,52,53].
For the French species we used Casevitz-Weulersse & Galkowski [54] adding some of the variations included in Monnin et al. [55], and Seifert [56] to clarify the status and current distribution of the European Lasius species.
The faunistic list of the Moroccan ant species was constructed from the results of Cagniant [57] and also included the additions provided in different publications [53,58,59,60,61,62,63,64,65,66].
We have also used the available data on the myrmecofauna of most of the other Mediterranean countries (Table 1) permitting us to put the IP ant species into a Mediterranean context.
2.2. Biogeographical Analysis
We undertook three different biogeographical aspects of the Iberian biogeography: 1. Analysis of the chorology of all the species included in the Iberian list; 2. Assessment of the distribution range of these species in the IP, emphasizing the endemic, rare and relict species; 3. To analyse the colonization history of the IP and propose four different ant groups, taking into account their most probable centres of origin and dispersal routes.
2.2.1. Chorology
According to Ribera [78] we established similar chorological categories (categories one to four) but adding three more (five to seven). Specifically, the seven chorological categories we have used are: 1. southern species (S) present in northern Africa and in some areas of the IP but never extending their distribution beyond the north of the Pyrenees; 2. northern species (N), mostly distributed in the north of Europe, northern Pyrenees and some other northern areas in the IP; 3. Iberian endemic species present only in the IP (X) or extending their distribution to the northern slopes of the Pyrenees or some areas in southeastern France (XS); 4. trans-Iberian species (T) present in the northern Pyrenees, the IP and northern Africa; 5. species living only in the northern Mediterranean basin (MN); 6. species distributed over all the Mediterranean basin (M); 7. introduced species (I).
2.2.2. Distribution Range
We have also aimed to portray the distribution of all the listed species for the IP in as much detail as possible, although the ant distribution and endemic species is still incomplete and the methods and effort sampling were heterogenous (pitfall traps, visual inspection, etc.) and sometimes unknown to us, such as those taken from bibliography which only report the species occurrence. Therefore, we have defined biogeographical subdivisions following the criteria established in other studies. Arnan et al. [79] established only two subdivisions in the IP, the Atlantic and Mediterranean, but we have subdivided the two biogeographic subregions into eight refugium areas (Figure 1), following Gómez and Lunt’s criteria [32] based on general biota occurrence (plants, vertebrate and invertebrates animals). We have limited the refugium areas to mountain ranges which act as borders because ants prefer low altitudes [6]. We considered an Atlantic and a Mediterranean coast refugia, which have been defined as biogeographical areas for butterflies [80]) and birds [81]. Some of these areas are supported as refugia such as the southern plateau, Atlantic and south-eastern Mediterranean for Pimelia species [15] or the entire Mediterranean during the Pleistocene for Polyommatus species [37]. Thus, we have finally defined the following refugium areas (Figure 1): 1. the Cantabric including the Cantabric and Basque ranges and a part of the eastern Galaic mountains; 2. the Pyrenean including all the southern slopes of the Pyrenean range from Navarra to Catalonia; 3. the Mediterranean, including the eastern and south-eastern coasts; 4. the Atlantic including the western Iberian coasts; 5. the northern plateau, bordered to the south by the northern slopes of the Central and Iberic ranges, and to the north by the Cantabric range; 6. the southern plateau, bordered to the north by the southern slopes of the Central and Iberic ranges and to the south by the Sierra Morena range; 7. the Guadalquivir Valley; and 8. the Ebro Valley. The geographical position of the citations has been obtained from the AntMaps.org web page [82], and confirmed with our own data and recent publications and reviews (see Material and Methods 1.1). Moreover, we have analysed the occurrence and the number of species found in every refugium area, as well as the number of refugium areas occupied by each species. We also analysed the endemic species distribution highlighting the rare species and the refugium areas which they inhabit. Finally, we tested the similarity of each of the refugium areas by mean of hierarchical clustering analysis based on Jaccard’s index as the association estimate and the paired group algorithm UPGMA (Unweighted Pair Grouping method with Arithmetic Means) procedure [80,83,84] as the agglomeration criterion using PAST program V. 4.04 [85] and excluding ubiquitous species. The significant differences in similarity (Jaccard’s Index) have been calculated among all the branches of the cluster [80,86,87].
2.2.3. Origin and Dispersal Routes
We collected biogeographical evidence from the bibliography about biological traits, dated molecular phylogenies (with different estimation methods) and fossil deposits of the different ant subfamilies, genera and species when available. We inferred from these data the origin and the most plausible dispersal routes up to the current distribution range, taking into account the most probable geology and climate distribution admitted for each epoch in the bibliography and the comparison results obtained from other taxa.
3. Results
3.1. Taxonomic Richness
The Iberian Peninsula (IP) has 299 ant species, the highest number compared to the French (211) and the Moroccan (233) ant faunistic lists (see Table S1 in Supplementary Material). The Iberian ant richness is only comparable with the 259 ant species of Greece and the 286 of Turkey. To the north the ant richness followed the latitudinal diversity gradient (Belgium 85 species [88], Norway 57 species [89]). Nevertheless, the ratio species richness/surface in the three countries revealed a similar ratio for the IP and Morocco (5.1 × 10−4 and 5.2 × 10−4 species/km2 respectively) and much lower for France (3.2 × 10−4 species/km2). Even in a wide view of the Mediterranean context, the ant fauna of the IP is the second in species richness, with only a few species less than the Balkan peninsula but not when the ratio of species/surface is taken into account, where the IP has a higher ratio than France, similar to Morocco, but surpassed by Greece and Israel. The Iberian ant species appear grouped in seven subfamilies (Amblyoponinae, Ponerinae, Proceratiinae, Leptanillinae, Dolichoderinae, Formicinae and Myrmicinae), but Morocco has two more subfamilies (Cerapachyinae and Dorylinae) and France one subfamily less than the IP (Amblyoponinae). The subfamilies Amblyoponinae, Cerapachyinae and Dorylinae, which are widely considered as tropical [90], are absent from France and only the subfamily Amblyoponinae is present in the IP.
The subfamily Myrmicinae had the highest species richness for the three studied areas, 114 species (54% of the total ants) in France, 171 (57%) in the IP and 147 (63%) in Morocco. The subfamily Formicinae has a lower number of species than Myrmicinae in all the three areas: 77 in France (36.5%), 95 species (33%) in the IP and 61 (26%) in Morocco. The subfamily Dolichoderinae appears far below, followed by Dorylinae and Cerapachyinae (see Supplementary material Table S1).
The most diverse genus in the IP is Temnothorax (47 species and 16% of the total species richness), followed by Lasius (25 species, 8%), Formica (23 sp., 8%) and Camponotus (20 species, 7%). Temnothorax is also highly diverse in France (28 sp., 13%) and Morocco (39 sp., 17%) but represents a slightly higher percentage of the total species in the latter country. The other most abundant genera in France are Formica (13%), Lasius (12%) and Myrmica (10%) and in Morocco Messor (12%), Aphaenogaster (11%) and Cataglyphis (9%).
Although the total number of endemic species in the IP (72 sp., 24 %) is the highest in the Mediterranean countries (21 endemic ant species in Greece [74]; 25 in Israel [76]; and 56 in Morocco [57]), but again the ratio of endemic species/surface is similar to Morocco (1.25 × 10−4 endemic sp./km2) and surpassed by Greece (1.6 × 10−4 endemic sp./km2) and Israel (12.0 × 10−4 endemic sp./km2). In the IP, the highest number of endemic ants belong to Myrmicinae (41 species) followed by the subfamily Formicinae (25 species, Figure 2). Amongst the Myrmicinae, the genus Temnothorax includes the most endemic species (16 species, 22% of the total endemic species and 30% of the full Temnothorax species). Nevertheless, the genus Cataglyphis is the most proportionally rich in endemic species, all of its 10 Iberian species are endemic. Although a positive correlation exists between the species richness and the number of endemic species within a genus (R = 0.73; F(1,28) = 31.81; p < 0.0001), some genera are especially above the expected number of endemic species, such as Cataglyphis, Temnothorax or Goniomma, and other genera below, such as Lasius, Tetramorium and Formica (Figure 3).
Moreover, the presence of an endemic genus in the IP, Iberoformica, with only one living species in the world, I. subrufa, is very remarkable. This species extends its distribution to some southern French locations. The exceptional appearance of the genus Rossomyrmex in the IP is also noteworthy. This genus has three different species in Asia (the Anatolian plateau, the plains of the Caspian Sea and other regions of Central Asia), which are different to the one in the IP, R. minuchae [30,58], the only species of this genus which exists in the full western Palaearctic.
Another interesting group of species because of their natural history are the social parasites of which 43 species appear in the IP, including temporal and permanent parasitism (22 and 21 species). Together they account for 14.4% of the total Iberian species. The number and percentage of parasite species decreased in France (20 temporal + 13 permanent which represent 15.7%), and many more in Morocco (5 temporal + 7 permanent which constitutes 5.2%).
Finally, comparing the shared species belonging to every subfamily amongst the IP, France and Morocco, (Table 2; Table S1) we obtained the highest percentage of overlapping species between the French and the Iberian ant lists (84%). Nevertheless, the IP only shared 59% of its species with the French list. Morocco shared the lowest number of species with France 21%. and the highest with the IP 37% (Table 2). A cluster based on the similarity (Jaccard’s Index, Cophenetic correlation index = 0.98) amongst the three areas, grouped France and the IP together (Figure 4) without significant differences between them (Jaccard’s Index = 0.52, p > 0.05) and showed significant differences between Morocco with France and the IP (Jaccard’s Index = 0.12 and 0.19 respectively, p < 0.01). Moreover, the replacement of species is more frequent between Morocco and the IP, but the overlapping is mainly between France and the IP (Table S1). For instance, France has three Bothriomyrmex species shared with the IP, but only one of the six Bothriomyrmex from Morocco appear amongst the four species in the IP. Even more striking is the case of the genus Cataglyphis, none of the 22 species in Morocco appear amongst the 10 species in the IP, which moreover are Iberian endemic species. Something similar occurred in other genera, such as Aphaenogaster, Oxyopomyrmex and Temnothorax, although to a lesser extent.
3.2. Biogeographical Analysis
3.2.1. Chorology
The most frequent chorological category in the IP was the northern species (N, 30.8%), which together with the trans-Iberian species (T, 11.7%) are 42.5% (Figure 5). On the other hand, the Mediterranean species, including the northern Mediterranean (MN, 6.7%), all the Mediterranean basin (M, 6.7%), the southern Mediterranean (S, 12%), and the Iberian endemic species (X + XS, 24.1%) represented 49.5% of the total species. The lacking percentage corresponded to the introduced species (I, 8%) (Figure 5). Amongst the ant subfamilies, the northern species (N) are clearly more abundant in Formicinae and Myrmicinae (Figure 6). In the latter subfamily the “endemic species” is the next most abundant chorological category (Figure 6). Nevertheless, the percentage of endemic species is similar for Formicinae and Myrmicinae (26% and 25%). The next most abundant category in Myrmicinae is the southern species (S), contrarily under-represented amongst the Formicinae ants (Figure 6).
3.2.2. Distribution Range
We have evaluated the importance of every refugium area harbouring ant species richness (see Supplementary material Table S2). We found that all the aprioristic defined refugium areas presented significant differences in similarity (Jaccard’s index, p < 0.01) with respect to the others (Table 3). Those refugia with the high similarity index were the northern plateau, the Guadalquivir Valley and the southern plateau (Jaccard’s index = 0.4; Table 3, Figure 7).
The refugium area with the highest number of ant species is the Mediterranean (144 species), followed by the Guadalquivir Valley (118 species) (Figure 8). These two refugium areas also contain the highest number of endemic species, followed by the northern plateau and the Pyrenees. The Atlantic and the southern plateau refugia contain an intermediate number of species, and the Cantabric together with the Ebro Valley present a similarly low number. The number of ubiquitous species, occurring in all the eight refugium areas was relatively low with only 46 species.
Many of the Iberian ant species appeared exclusively in one of the Iberian refugium areas (86 species, 29%) and may be considered as rare species, although they are not always endemic. Progressively a lower number of species occupied more refugium areas, with only a final increase for ubiquitous species (Table 4). The 21 endemic species which appear in only one refugium area are shown in Table 5.
Analysing the most diversified or peculiar ant genera (see Annex S2, Supplementary Material), we found the genus Lasius (25 species) to be the most widely distributed across all the IP and, when local, generally appear in the northern half of the IP and frequently in the Pyrenees, the Ebro Valley and the Mediterranean refugia. There are only two endemic species in this genus that are widely distributed (L. cinereus and L. piliferus). The genus Formica is composed of 23 species, the majority appearing in the Pyrenees (14 species) or the Mediterranean (9 species) refugia. Twelve Formica species of the IP belong to the rufa group; again, the majority of these latter species (7 of them) were present in the Pyrenean refugium and only 3 in the Guadalquivir Valley. The genus Camponotus is composed of 20 species, of which 16 are widely distributed in the IP. Nevertheless, the only two strictly endemic species (X chorological category) belonging to this genus (C. haroi and C. amaurus) presented a narrow distribution linked to the Mediterranean coast and the northern plateau. In the case of the genus Cataglyphis, 10 species are represented in the Iberian myrmecofauna, all of which are endemic (nine strictly X and one shared with France XS). Species from this genus have never been found in the most northern zones (Cantabric or Pyrenees) and principally were present in the southern areas (southern plateau, the Guadalquivir Valley and on the Mediterranean coast). Four of the endemic Cataglyphis species (C. floricola, C. gadeai, C. humeya and C. tartessica) are exclusively distributed in either the Guadalquivir Valley or the Mediterranean refugium areas. The genus Themnothorax, includes 47 species in the IP, 16 of which are endemic. Twenty-five percent of the Temnothorax species are distributed in only one refugium area, and the areas with the most Temnothorax species are the northern plateau, the Mediterranean coast and the Guadalquivir Valley. The refugium area with the lowest number of Temnothorax species is the Cantabric. Finally, the genus Stigmatomma is composed of only three species, with a narrow distribution, in only one or two refugium areas, thus in the Guadalquivir Valley we found the three species and one of them also on the Mediterranean and Atlantic coasts. In fact, the Guadalquivir Valley is the refugium containing the highest number of ant species linked with tropical environments, such as Stigmatomma emeryii, S. impressifrons, and the Ponerinae Anochetus ghilianii and Chryptopone ochracea.
4. Discussion
4.1. Taxonomic Richness
Ants showed a higher number of species in the IP (299 species), compared with the adjacent Mediterranean countries, with only a few species less than the Balkan Peninsula. A similar number of species is found in Greece and Turkey [74,75] (Table 1). Following the expected latitudinal diversity gradient there is a decrease in species richness to the north [88,89]. Although the IP has a higher ant species richness than Morocco [5,91,92], the last has two more subfamilies than the IP and three more than France, which follows the latitudinal diversity gradient. These results varied when the area surface is taken into account, the ratio is similar between the IP and Morocco, being higher than in France and much lower than other Mediterranean countries such as Israel.
With respect to the richness of the endemic species the same pattern occurred. The IP shows the highest number (72 endemic species), compared with other Mediterranean countries such as Greece, (21 species [74]); Israel (25 species [76]) or Morocco (56 species [57]), but when the surface areas are considered, a similar ratio between the IP and Morocco again occurred, but this was much higher in the eastern Mediterranean.
Nevertheless, not only the number of species and endemic taxa in the IP are remarkable but also the singularity of the Iberian myrmecofauna including one endemic genus (Iberoformica), with only one endemic species I. subrufa and the fossil of its probable ancestor I. horrida [93]. Moreover, the IP is the only place in Europe where the genus Rossomyrmex is located outside Asia. The presence of these two exclusive genera highlights the importance of the IP as a refugium. At least for Rossomyrmex, there is evidence that have been several episodes of extinction of this genus between the nearest geographical point (Turkey) and the IP. This genus has survived in the IP and not in other nearby countries, as have other emblematic animals such as the Lynx or the Azure-winged magpie [94].
The presence of these two ant genera as well as the low number of Iberian ant species shared with Morocco (29%) and with France (59%), while France shares with the IP 84%, and Morocco 37% with the IP, indicates that the IP harbours a unique and fairly exclusive ant fauna and functions as a "cul de sac" of ant species [94].
Several factors are responsible for the high ant biodiversity in the IP, as mentioned in the introduction, derived from the highly complex Iberian orography and convulse geological and climatic history. Another factor affecting the Iberian ant diversity is the peninsular effect, which explains the increase in diversity near the isthmus [95,96], assessed in the IP for birds [97] and butterflies [98]. Moreover, the role of the Palaearctic peninsulas during the Quaternary glaciations, as refugia for biota and centres of speciation [36], together with the Iberian complex orography producing “refugia-within-refugium” effect that has been assessed for different animal groups, contributed to increase the Iberian biodiversity [14,99].
Analysing the contribution in the IP ant diversity of the most relevant worldwide genera according to Wilson (1976 in Pie & Feitosa, [100]), only Camponotus make an important contribution to Iberian biodiversity, whilst Pheidole and Crematogaster had few species represented. This phenomenon is common to all Europe [69]. In the IP, the genus that we can consider “to have conquered the world” following Wilson’s criteria must be Temnothorax, the most diverse Iberian genus (47 Iberian species) with the highest number of Iberian endemic species (16). This is an abundant genus in all the IP habitats, except in the high mountains above 2,700 m a.s.l., reflecting a high plasticity and adaptation ability. Probably its small size and its mode of mate location behaviour, by means of female pheromones, act as a speciation driver in this genus. Nevertheless, other diverse genera such as Lasius, Formica or Camponotus have a much lower number of endemic species than expected. Again, the mating behaviour by mass nuptial flights, which facilitates the populations mixing at a landscape level, might influence the lower rates of speciation in these genera. Moreover, other genera, such as Cataglyphis or Goniomma, showed a higher number of endemic species than expected. Other factors, probably linked with the biogeography, biotic traits and plasticity of the genera, may be responsible for this high level of endemicity.
All these data contribute to highlight the singularity of the Iberian myrmecofauna and its relevance in the Mediterranean region, placing the IP in a similar position to the Balkan Peninsula as a centre of ant diversification.
4.2. Biogeographical Analysis
4.2.1. Chorology
The chorological analysis showed two groups of ants, one widely distributed in the northern half of Iberia, sharing species with the western Palaearctic region but sometimes extending up to northern Africa, (N + T chorological categories), and the other group, mainly including species belonging to the Mediterranean basin and northern Africa (S + MN + M + X + XS chorological categories). This result is congruent with those obtained for aquatic coleopterans [78] and showed the IP acts as a transitional element between Europe (north Palaearctic) and north Africa (south Palaearctic).
This north-south distribution pattern is shown at different taxonomic levels (ant genera and species), thus we can find genera preferentially distributed in the north of the IP, such as Formica, Lasius, Myrmica and Leptothorax and others in the southern half, such as Cataglyphis, Proformica, Stigmatomma, Anochetus, Messor, etc. At species level, we can find the same pattern, with some typically northern species such as Lasius fuliginosus, most of the Formica rufa group, Camponotus herculeanus, C. ligniperda, Tapinoma pygmaeum, the three Leptothorax species, and more examples included in the Supplementary Material (Annex S2). In the same way, a similar number of species could be considered as southern, such as some of those included in the genus Cataglyphis (C. humeya, C. floricola, C. tartessica), Goniomma (G. collingwoodi, G. compressiquama), Messor timidus, Aphaenogaster striativentris, the three species of the genus Stigmatomma, or Anochetus ghilianii, amongst others (see Table S2).
4.2.2. Distribution Ranges
The complex orography of the IP produced not only a north-south division of species distribution, but even in the eight proposed refugium areas, we can find a significantly different ant fauna. This result supports the fact that similar processes affected all the biota, with common patterns emerging, although some slight differences in shape probably depend on biotic traits of the studied species such as dispersal ability [32,80,81,84,101]. Most of the proposed biogeographical subdivisions of the IP established between five (Mollusca: Pulmonata [101]) to eleven zones (birds [81]). Our proposal is closer to that of vascular plants and vertebrates [32,81,84]. The social character of ants does not appear to alter the biogeographical pattern followed for the general biota, except for their presence in the high mountains. Amongst the IP refugia the most similar for occurrence of ants (high Jaccard’s diversity Index) are the northern plateau, the Guadalquivir Valley and the southern plateau. From the analysis of the species occurrence within each refugium areas, we have proved that the Mediterranean and Guadalquivir Valley refugia show the highest species richness and number of endemic species, highlighting their importance for ant conservation. In fact, Andalusia is considered the Iberian region with the highest general biodiversity [21] where both refugium areas are represented. In the Guadalquivir Valley refugium, we can find some special habitats such as the prequaternary forest with relict plant species located near the Straits of Gibraltar [102]. In this habitat we can also find the very few semitropical ant species appearing in the IP, which belong to the subfamilies Amblyoponinae together with some Ponerinae such as Anochetus ghilianii.
The Pyrenean and the northern plateau are the following refugium areas in biodiversity importance. The Pyrenean range is in the isthmus, concentrating the peninsular effect, which together with the high heterogeneity derived from the altitudinal variation and its west-east position, produced an important climatic variability, responsible for the species richness in this refugium. Nevertheless, the number of endemic species is not very high (10 species), pointing to the possibility of gene flow between the Pyrenean and the European populations, diluting speciation possibilities. The northern plateau is the oldest region of the IP, including the Hercynian plate; its age may be the cause of the increment in biodiversity in this refugium.
Most of the species are distributed in more than one refugium area, but about a third is found in only one IP refugium (86 species). On the other hand, only 46 species are ubiquitous in all the IP.
Similarly, from analysing the distribution of endemic species in the different refugium areas, we have been unable to find a clear pattern, finding 21 endemic species limited to only one refugium and only five endemic species being considered ubiquitous. These results are congruent with those found by different authors [78,99] pointing to different causes for the absence of clear patterns of endemicity in the IP, including sampling artifacts.
On the other hand, although mountain ranges promoted species richness and endemicity [103], this pattern is not detectable for the Iberian and Palaearctic formicids. Only one ant species is exclusive to high mountains: Proformica longiseta (between 1800 and 3000 m a.s.l. in the Sierra Nevada mountains [104] and the Baetic range [13]). According to these findings, the majority of the Iberian ant endemic species are related with semiarid habitats with scarce vegetation at low altitudes, such as some of the species pertaining to the genus Temnothorax, as T. ansei, T. blascoi, T. caesari, T. crepuscularis [105]. Another important number of endemic species are found in medium altitude mountain ranges, such as Teleutomyrmex kutteri (1700–2250 m a.s.l. in different Baetic ranges [106,107]), Temnothorax gredosi or T. conatensis (900–1500 m a.s.l. [108,109]).
Whereas the rare species, i.e., those found in only one or two refugium areas, include for instance the social parasite ant species, where their way of life makes them difficult to find. These are known in very few and sometimes very distant areas, such as Rossomyrmex minuchae [12,110], Myrmoxenus bernardi [111] or Anergates atratulus [112]. Another example of a rare species is Aphaenogaster cardenai, an Iberian endemic species of relatively wide distribution [113], but living in scarcely sampled habitats (the mesovoid shallow substratum, shallow caves and mines [114]).
4.2.3. Origin and Dispersal Routes
Other different causes responsible for the high ant biodiversity in the IP are related with the centre of origin and dispersal routes of the ancestors of the current ant fauna. In agreement with Blaimer et al. [115] the origin of formicids probably occurred during late Cretaceous (104–117 mya), but the rise of the modern ant fauna probably occurred during the early Cenozoic and continued until the present day. There are signs that all the genus-level taxa appeared from the first 50–60 mya are now extinct in the western Palaearctic [7]. Blaimer et al. [115] gave the estimated time of divergence for several genera from the subfamily Formicinae, thus, the clade giving place to the tribe Formicini, appeared 60 mya, and the most probable ancestral range was the Palaearctic region. These data are congruent with the estimated age of Cataglyphoides constrictus, a probable ancestor of the genus Cataglyphis (but see [116]), or Formica horrida, probably a species belonging to the current genus Iberoformica [93]. Both fossils were found in the Baltic amber (middle to late Eocene 37–42 mya) [117]. According to Blaimer et al. [115] the Formica/Iberoformica clade appearance is dated to 50 mya, the clade of Rossomyrmex, Cataglyphis/Bajcaridris, Proformica diverged in 45 mya, Bajcaridris/Proformica in 20 mya and finally Rossomyrmex/Cataglyphis in 15 mya, which are in agreement with the results of Guenard et al. [7].
The genus Temnothorax, which belongs to the subfamily Myrmicinae, showed its centre of origin in the Palaearctic region during the Eocene-Oligocene transition (38–33 mya) [118]. The genus Myrmica appeared to diversify following drastic climatic cooling in the same epoch (34 mya), and its most probable centre of origin is central or south-eastern Asia [119]. The genus Stenamma shared its time of origin with the previous genera but the centre of origin is estimated in the Nearctic region, and two dispersal waves to the Palaearctic were detected, the first in early Oligocene (30 mya), leaving in the Iberian myrmecofauna the species S. striatulum, and the second at late Pliocene (about 3 mya) with S. debile [120]. Undoubtedly posterior dispersals have occurred, as has been demonstrated for other insects such as the butterfly Parnassius apollo with a recent (late Pleistocene) colonization of most of their range [121].
With this retrospective view, we know that most of the current extant ant genera inhabited the Palaearctic region 15 mya (Middle Miocene), but the question is whether they inhabited the IP at the same time.
From the Cenozoic (60 mya), the Iberian plate was fused with the European plate [95]. Unfortunately, the ant fossil data from this time are scarce and, in some cases, too ancient to resolve the question [122,123]. We only have some scarce data from recent periods, thus, the existence of the genera Camponotus, Dolichoderus, Iridomyrmex, Lasius or Formica, Liometopum and Messor, in the Middle and Late Miocene of the IP has been reported (13 to 7 mya) [124,125]. The paleoclimate and paleoenvironments of the Miocene (23 mya) have been recreated more accurately through the coral reef and mammal fossils [24,29,30,126]. During this period the IP climate was semitropical and the biota was equivalent to the current African savannah [127], including important Asian and African components [27]. Nevertheless, from the Miocene until the Pliocene (5.33 mya) the biogeography of the IP was quite convulsed, especially on the southern extreme, frequently joined and separated with the African plate [26]. Moreover, the climate was cooling which finally provoked the substitution of the semitropical fauna for others better adapted to temperate or cool climates [7].
In this situation, some ant genera may have inhabited the Iberian savannah from the Miocene and later the steppes from the Pliocene and Pleistocene [7,115]. This dynamic process of sequenced colonization, extinction and diversification did not stop. Thus, during the transition Pliocene-Pleistocene, the effect of Quaternary glaciations on these Asian and African fauna dispersed to the IP, giving way to the current myrmecofauna. This view is reinforced with data on radiation of the Formica genus and the Formica rufa group species, dated in the Middle Pliocene and Pleistocene [128,129], and this was similar for other insects, as has been proved by means of molecular analyses in different coleoptera [14,15,16,17,18]) and in plants [130,131], pointing to a general process for all the biota of the IP.
In our analysis of the origin and dispersal routes which shaped the Iberian ant myrmecofauna, we can differentiate four groups of dispersal patterns:
Relict Species
From the middle to late Eocene (34–42 mya) [117] the climate was semi-tropical and the ant fauna known from the Baltic amber was very diverse including extant genera or their ancestors [132,133]. Some of them, such as Formica or Aphaenogaster, which nowadays have reached a high diversity, must have previously adapted to semi-tropical warm environments that were predominant when they were trapped by the amber. Later they were able to adapt to the cooling produced from the end of the Miocene and have survived until now. Nevertheless, both genera do so in different ways, one adapting to temperate or cool habitats (Formica, now more diversified to the north of the IP) and the other to temperate or warm habitats (Aphaenogaster, currently more diversified in the south of the IP). Nevertheless, some other genera were unable to adapt to the changing conditions and disappeared from the western Palaearctic [7], such as the fossils of the genera Pachycondyla from Denmark, preceding the Paleocene-Eocene transition (55 mya) [134]. In some rare cases, semitropical ant species such as Anochetus and Stigmatomma, found in Santo Domingo amber (16–19 mya) [117,135] and the Baltic amber [133] respectively, and currently well distributed in tropical and semitropical regions, were able to remain in some restricted locations of the IP surrounding the Straits of Gibraltar [136,137], an exceptional zone also inhabited by paleoendemic plant species [102,138]. Although Jowers et al. [9] concluded that Anochetus ghilianii is a recent invader of the IP, we think that the coincidence in this area of botanical paleoendemic species together with different semitropical ants (Anochetus and Stigmatomma species), it must be the result of a similar biogeographical process acting on them. Anyway, A. ghilianii should be considered as a relict Moroccan species and its presence in the IP is noteworthy.
Asian-IP Disjunct Species
In the Pliocene (5.33 mya) the Mediterranean Sea reopened through the Straits of Gibraltar and the climate was cooling, favouring the substitution of the existing semitropical fauna for one better adapted to temperate climates. This temperate fauna proceeded principally from Central Asia, reaching the IP as confirmed by fossil deposits [25,28,29,30]. The Asian colonizing waves have occurred repeatedly during the transition Pliocene-Pleistocene (2.6, 2.5 and 1.7 mya) [25] even during the late Pleistocene [139] and later. The migrant mega-fauna must have been dispersing together with invertebrates, among them obviously ants, sharing the same origin and similar ecological requirements. These Asiatic and African faunas from the later Pliocene, evolved under glacial and interglacial periods during the Pleistocene, and many of these species became extinct or remained isolated in some refugia throughout the entire Iberian refugium, setting up the current myrmecological fauna [22,31,32,33]. Some of these species continued isolated and now have a narrow distribution range, as is the case of the slave-making ant R. minuchae in Spain, and the other three species of the genus, which are dispersed, although with a wider distribution range in Asia. The genus Rossomyrmex can be considered as a relict in the IP. On the other hand, the species of the genus Proformica, the host of Rossomyrmex, with a probable centre of origin in Central Asia [11], were able to recolonize wide distribution ranges both before and after glacial periods, occupying all the southern Palaearctic from Central Asia to the IP. This genus, more diversified in the western Mediterranean, specifically in the IP, is not distributed in Morocco, where it has been replaced by the close genus Bajcaridris [11,115,140]. This fact suggests the hypothesis of a frequent dispersal route on the two sides of the Mediterranean (Figure 9) with speciation concluding in different genera (Proformica and Bajcaridris). An opposite direction of the dispersal route (from western Mediterranean to Asia) has been suggested for Rossomyrmex [11] as occurred, for example, with the plant genus Odontites [130].
Another genus offering interesting disjunct distributions is Cataglyphis, specifically the species group altisquamis shows two different and discontinuous distribution ranges: one group of five species distributed from Central Asia to the western Mediterranean and the other nine species distributed in the western Mediterranean (IP and Morocco) [141] (Figure 10). This second group of species is absent from Turkey to France, by the northern route, and from Egypt and Libya, by the southern route, due to extinction processes in the intermediate points. The age of the entire genus and its high diversity in Asia point to a Central Asian origin. Three different dispersal routes are possible from Asia to the western Mediterranean: 1. the simultaneous dispersal route via the two sides of the Mediterranean, 2. the dispersal by the northern Mediterranean side across to the Balkan region, and 3. the dispersal only by the southern Mediterranean side (Figure 9). The three are possible but until now only the southern route (3) has been suggested for the Cataglyphis albicans group [10]. Nevertheless, the three dispersal routes from Asia have also been proposed for the genus Cephalota (Coleoptera: Cicindelidae) [17] and may be more frequent dispersal patterns than had generally been considered. An extension of the phylogenetic study on Cataglyphis including middle-west distributed species should produce a wide and accurate view for the biogeography and dispersal patterns of these species.
The genera Goniomma and Oxyopomyrmex, both endemic of the Mediterranean basin, also show disjunction between the eastern and western Mediterranean distributions [142,143] (Figure 11). Their centre of origin is unknown, although they probably appeared in the Miocene (12 mya) [90]. A similar distribution pattern is found in the coleopteran species of the genus Pimelia [15], Berberomeloe [14] and the plant genus Odontites [130]. Another example of current Iberian fauna showing Asian-IP disjunct distribution is the Iberian azure-winged magpie [144] or amongst the insects, the lepidopteran Pseudochazara wiliamsi or the coleopteran of the subgenus Parentius (see [145]), and together with these, we find many other species adapted to arid or semi-arid environments such as Monegros (Zaragoza) or Guadix-Baza (Granada) [146,147] or to mountain ranges [148]. Thus, all these examples reinforce the importance of Asia as a centre of origin for the Iberian fauna.
Baetic-Rifan Species
This dispersal pattern includes the shared species between the south of the IP and the north of Africa. Within this group, the ants with apterous or brachypterous females, such as Monomorium algiricum, A. ghilianii, Stigmatomma species or Aphaenogaster senilis group, gain biogeographical relevance, due to their handicap for flight dispersal across the Mediterranean Sea. One plausible explanation for this kind of distribution comes from the geological history of this territory. South-western Iberia and the north of Morocco share stratigraphic deposits, forming the Baetic-Rifan territories [150] derived from the Miocenic island of Alborán, emerged in the western Mediterranean 16 mya [126]. This island was united with Morocco until 8 mya when it again became disconnected and isolated until 5.9 mya. After this, it was again partially united with the IP until 5.3 mya. At this time, the Straits of Gibraltar reopened, dividing the territories of the Miocenic island of Alborán between the IP and Morocco. The existence of this island may explain the relatively high presence of apterous species in this zone, because apterism is one of the known island syndromes. This effect is known in the Baetic-Rifan territories not only for ants [151] but also for Pimelia and Blaps Tenebrionid [15,17] and Meloid (Berberomeloe group) coleoptera [14], both of them apterous. The existence of this island is more extended in geological time than the habitually invoked desiccation of the Mediterranean during the Messiniense (late Miocene) to explain the shared presence of species between the IP and northern Africa.
Alpine Species
Another important and more recent dispersal route is related with the Pleistocene glaciations, the last occurring 10,000 years ago. Many northern species survived and took refuge in the southern Palaearctic peninsulas, and during interglacial periods returned to the north and/or the populations climbed the slopes of the mountains [37]. One good example of this groupis the pair of Iberian endemic species Formica frontalis/F. dusmeti, very close phylogenetically to F. truncorum, distributed in northern Europe ([129], unpublished data). Probably the presence of T. kutterii in the south of the IP and T. schneideri in the northern IP and in the Alps is likely to have had this similar origin. Some species of Myrmica, Temnothorax, Lasius, etc, may belong to this group of alpine species, but this should be confirmed by phylogenetic molecular studies on the Mediterranean species.
5. Conclusions
The taxonomic diversity and distribution patterns we have presented in this study highlight the importance of Iberian ants for a better understanding of the complex evolutionary history and biogeography of the IP. Moreover, we have tried to show the importance of biogeography driving the structure of the current ant communities, ecological interactions, such as hierarchic competence and biotic factors, only explain a part of their structure [152]. Ours results on ant biogeography support the hypothesis that the centre of origin of the species and their dynamic processes (dispersal, vicariance, speciation and extinction) are the missing link to fully explain and understand the evolution and the structure of these ant communities [2,5,7]. We hope the hypothesis and proposals put forward in this study will promote new biogeographical, phylogenetic and evolutionary studies about the ant fauna of the Mediterranean Basin and produce a wide and accurate view of the centre of origin and the dispersal patterns of Iberian ants.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/1424-2818/13/2/88/s1, Table S1. Ant species list of France, Morocco and the IP. Table S2. List of Iberian ant species and their occurrence in the refugium areas.
Author Contributions
Conceptualization, A.T. and F.R.; methodology, A.T. and F.R.; formal analysis, A.T. and F.R.; investigation, A.T. and F.R; writing—original draft preparation, A.T. and F.R.; writing—review and editing, A.T. and F.R.; project administration, F.R.; funding acquisition, F.R. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the project RTA2015-00012-C02-02 (Ministry of Science and Innovation, INIA and FEDER funds).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in supplementary material.
Acknowledgments
We thank José M. Martín, Alfonso Arribas and Elvira Martín who provided some references and exchange of ideas and Pedro Sandoval for his help with the figures. Angela Tate reviewed the English edition. We are also grateful to the three anonymous reviewers who made a careful and significant improvement to the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Jenkins, C.N.; Sanders, N.J.; Andersen, A.N.; Arnan, X.; Brühl, C.A.; Cerda, X.; Ellison, A.M.; Fisher, B.L.; Fitzpatrick, M.C.; Gotelli, N.J.; et al. Global diversity in light of climate change: The case of ants. Divers. Distrib. 2011, 17, 652–662. [Google Scholar] [CrossRef] [Green Version]
- Andersen, A.N.A.N. Ant megadiversity and its origins in arid Australia. Austral Entomol. 2016, 55, 132–137. [Google Scholar] [CrossRef]
- Economo, E.P.; Narula, N.; Friedman, N.R.; Weiser, M.D.; Guénard, B. Macroecology and macroevolution of the latitudinal diversity gradient in ants. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hakala, S.M.; Seppä, P.; Helanterä, H. Evolution of dispersal in ants (Hymenoptera: Formicidae): A review on the dispersal strategies of sessile superorganisms. Myrmecol. News 2019, 29, 35–55. [Google Scholar]
- Dunn, R.R.; Agosti, D.; Andersen, A.N.; Arnan, X.; Bruhl, C.A.; Cerdá, X.; Ellison, A.M.; Fisher, B.L.; Fitzpatrick, M.C.; Gibb, H.; et al. Climatic drivers of hemispheric asymmetry in global patterns of ant species richness. Ecol. Lett. 2009, 12, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.L. Biogeography. In Ant Ecology; Lach, L., Parr, C.L., Abbott, K.L., Eds.; Oxford University Press: Oxford, UK, 2010; pp. 18–37. [Google Scholar]
- Guénard, B.; Perrichot, V.; Economo, E.P. Integration of global fossil and modern biodiversity data reveals dynamism and stasis in ant macroecological patterns. J. Biogeogr. 2015, 42, 2302–2312. [Google Scholar] [CrossRef]
- Catarineu, C.; Barberá, G.G.; Reyes-López, J.L. Zoogeography of the ants (Hymenoptera: Formicidae) of the Segura River Basin. Sociobiology 2018, 65, 383–396. [Google Scholar] [CrossRef] [Green Version]
- Jowers, M.J.; Taheri, A.; Reyes-López, J. The ant Anochetus ghilianii (Hymenoptera, Formicidae), not a Tertiary relict, but an Iberian introduction from North Africa: Evidence from mtDNA analyses. Syst. Biodivers. 2015, 13, 865–874. [Google Scholar] [CrossRef]
- Villalta, I.; Amor, F.; Galarza, J.A.; Dupont, S.; Ortega, P.; Hefetz, A.; Dahbi, A.; Cerdá, X.; Boulay, R. Origin and distribution of desert ants across the Gibraltar Straits. Mol. Phylogenet. Evol. 2018, 118, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Sanllorente, O.; Lorite, P.; Ruano, F.; Palomeque, T.; Tinaut, A. Phylogenetic relationships between the slave-making ants Rossomyrmex and their Proformica hosts in relation to other genera of the ant tribe Formicini (Hymenoptera: Formicidae). J. Zool. Syst. Evol. Res. 2018, 56, 48–60. [Google Scholar] [CrossRef]
- Ruano, F.; Devers, S.; Sanllorente, O.; Errard, C.; Tinaut, A.; Lenoir, A. A geographical mosaic of coevolution in a slave-making host-parasite system. J. Evol. Biol. 2011, 24, 1071–1079. [Google Scholar] [CrossRef] [PubMed]
- Sanllorente, O.; Ruano, F.; Tinaut, A. Large-scale population genetics of the mountain ant Proformica longiseta (Hymenoptera: Formicidae). Popul. Ecol. 2015, 57, 637–648. [Google Scholar] [CrossRef]
- Sánchez-Vialas, A.; García-París, M.; Ruiz, J.L.; Recuero, E. Patterns of morphological diversification in giant Berberomeloe blister beetles (Coleoptera: Meloidae) reveal an unexpected taxonomic diversity concordant with mtDNA phylogenetic structure. Zool. J. Linn. Soc. 2020, 189, 1249–1312. [Google Scholar] [CrossRef]
- Mas-Peinado, P.; Buckley, D.; Ruiz, J.L.; GarcÍa-ParÍs, M. Recurrent diversification patterns and taxonomic complexity in morphologically conservative ancient lineages of Pimelia (Coleoptera: Tenebrionidae). Syst. Entomol. 2018, 125, 331–348. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F.A.H.; Wahlberg, N.; Rasplus, J.Y.; Kergoat, G.J. What causes latitudinal gradients in species diversity? Evolutionary processes and ecological constraints on swallowtail biodiversity. Ecol. Lett. 2012, 15, 267–277. [Google Scholar] [CrossRef]
- Herrera-Russert, J.; López-López, A.; Serrano, J.; Matalin, A.; Galián, J. Influence of the Mediterranean basin history on the origin and evolution of the halophile tiger beetle genus Cephalota (Coleoptera: Cicindelidae). Ann. Soc. Entomol. Fr. 2020, 56, 447–454. [Google Scholar] [CrossRef]
- Ribera, I.; Castro, A.; Díaz, J.A.; Garrido, J.; Izquierdo, A.; Jäch, M.A.; Valladares, L.F. The geography of speciation in narrow-range endemics of the “Haenydra” lineage (Coleoptera, Hydraenidae, Hydraena). J. Biogeogr. 2011, 38, 502–516. [Google Scholar] [CrossRef] [Green Version]
- Myers, N.; Mittermeler, R.A.; Mittermeler, C.G.; Da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Araújo, M.B.; Lobo, J.M.; Moreno, J.C. The effectiveness of Iberian protected areas in conserving terrestrial biodiversity. Conserv. Biol. 2007, 21, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Hewitt, G.M. Mediterranean Peninsulas: The Evolution of Hotspots. In Biodiversity Hotspots; Springer: Berlin/Heidelberg, Germany, 2011; pp. 123–147. [Google Scholar]
- Martín-Piera, F.; Sanmartín, I. Biogeografía de áreas y biogeografía de artrópodos holárticos y mediterráneos. Bol. Soc. Entomol. Aragon. 1999, 26, 535–560. [Google Scholar]
- Braga, J.C.; Martín, J.M. Los arrecifes del Mioceno de Almería. Investig. Gest. 1997, 1, 5–19. [Google Scholar]
- Arribas, A.; Garrido, G.; Viseras, C.; Soria, J.M.; Pla, S.; Solano, J.G.; Garcés, M.; Beamud, E.; Carrión, J.S. A mammalian lost world in Southwest Europe during the late pliocene. PLoS ONE 2009, 4, e7127. [Google Scholar] [CrossRef] [Green Version]
- Martín Martín, J.M.; Braga Alarcón, J.C.; Gómez Pugnaire, M.T. Itinerarios Geológicos por Sierra Nevada; de Andalucía, J., Ed.; Consejería de Medio Ambiente: Seville, Spain, 2008. [Google Scholar]
- Masseti, M.; Mazza, P.P.A. Western European Quaternary lions: New working hypotheses. Biol. J. Linn. Soc. 2013, 109, 66–77. [Google Scholar] [CrossRef] [Green Version]
- Arribas, A.; Riquelme, J.A.; Palmqvist, P.; Garrido, G.; Hernández, R.; Laplana, C.; Soria, J.M.; Viseras, C.; Durán, J.J.; Gumiel, P.; et al. Un nuevo yacimiento de grandes mamíferos villafranquienses en la Cuenca de Guadix-Baza (Granada): Fonelas P-1, primer registro de una fauna próxima al límite Plio-Pleistoceno en la Península Ibérica. Bol. Geol. Min. 2001, 112, 3–34. [Google Scholar]
- Arribas, A.; Baeza, E.; Bermúdez, D.; Blanco, S.; Durán, J.J.; Garrido, G.; Gumiel, J.C.; Hernández, R.; Soria, J.M.; Viseras, C. Nuevos registros paleontológicos de grandes mamíferos en la Cuenca de Guadix-Baza (Granada): Aportaciones del Proyecto Fonelas al conocimiento sobre las faunas continentales del Plioceno-Pleistoceno europeo. Bol. Geol. Min. 2004, 115, 567–582. [Google Scholar]
- Garrido, G. Generalidades Sobre Los Carnívoros Del Villafranquiense Superior En Relación Con El Registro Fósil De Fonelas P-1 Reflections on the Carnivores of the Upper Villafranchian Represented in the Fossil Record of the Fonelas P-1 Site. In Vertebrados del Plioceno Superior Terminal en el Suroeste de Europa: Fonelas P-1 y el Proyecto Fonelas; Arribas, A., Ed.; IGME: Madrid, Spain, 2008; pp. 85–146. ISBN 9788478407644. [Google Scholar]
- Centeno-Cuadros, A.; Delibes, M.; Godoy, J.A. Phylogeography of Southern Water Vole (Arvicola sapidus): Evidence for refugia within the Iberian glacial refugium? Mol. Ecol. 2009, 18, 3652–3667. [Google Scholar] [CrossRef] [PubMed]
- Gómez, A.; Lunt, D.H. Refugia within Refugia: Patterns of Phylogeographic Concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia: Evolutionary Perspectives on the Origins and Conservation of European Biodiversity; Springer: Dordrecht, The Netherlands, 2007; pp. 155–188. ISBN 9781402049040. [Google Scholar]
- Lobo, J.M.; Moreno, J.C. Spatial and environmental determinants of vascular plant species richness distribution in the Iberian Peninsula and Balearic Islands. Biol. J. Linn. Soc. 2001, 73, 233–253. [Google Scholar] [CrossRef]
- Miraldo, A.; Hewitt, G.M.; Paulo, O.S.; Emerson, B.C. Phylogeography and demographic history of Lacerta lepida in the Iberian Peninsula: Multiple refugia, range expansions and secondary contact zones. BMC Evol. Biol. 2011, 11, 170. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the Quaternary. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galkowski, C. Quelques fourmis nouvelles ou intéressantes pour la faune de France (Hymenoptera, Formicidae). Bull. Soc. Linn. Bordx. Tome 2008, 143, 423–433. [Google Scholar]
- Galkowski, C. Une liste des fourmis (Hymenoptera; Formicidae) récoltées dans la région de Grasse, avec la mention d’une nouvelle espèce de la faune de France. Bull. Soc. linn. Provence 2011, 62, 1–4. [Google Scholar]
- Steiner, F.M.; Csősz, S.; Markó, B.; Gamisch, A.; Rinnhofer, L.; Folterbauer, C.; Hammerle, S.; Stauffer, C.; Arthofer, W.; Schlick-Steiner, B.C. Turning one into five: Integrative taxonomy uncovers complex evolution of cryptic species in the harvester ant Messor “structor”. Mol. Phylogenet. Evol. 2018, 127, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Seifert, B.; Buschinger, A.; Aldawood, A.; Antonova, V.; Bharti, H.; Borowiec, L.; Dekoninck, W.; Dubovikoff, D.; Espadaler, X.; Flegr, J.; et al. Banning paraphylies and executing Linnaean taxonomy is discordant and reduces the evolutionary and semantic information content of biological nomenclature. Insectes Soc. 2016, 63, 237–242. [Google Scholar] [CrossRef]
- Salgueiro, J. Catálogo dos formicídeos de Portugal continental e ilhas. Bol. Soc. Entomol. Aragon. 2002, 31, 145–171. [Google Scholar]
- Collingwood, C.A.; Prince, A. A guide to ants of continental Portugal. Bol. Soc. Port. Entomol. 1998, Suplement. [Google Scholar]
- Boeiro, M.; Espadaler, X.; Azedo, A.R.; Collingwood, C.; Serrano, A.R.M. One genus and three ants species new to Portugal (Hymenoptera, Formicidae). Bol. Soc. Entomol. Aragon. 2009, 45, 515–517. [Google Scholar]
- Gonçalves, C.; Espadaler, X.; Pereira, J.A.; Santos, S.; Patanita, M.I. Primeiros registos das espécies Strongylognathus caeciliae Forel, 1897 e Temnothorax tyndalei (Forel, 1909)(Hymenoptera, Formicidae) em Portugal Continental. Bol. Soc. Entomol. Aragon. 2014, 54, 402. [Google Scholar]
- Henin, J.M.; Collingwood, C.; Paiva, M.R. Synonymy between Leptothorax caparica Henin, Paiva & Collingwood, 2001 and Cardiocondyla mauritanica. Bol. Soc. Port. Entomol. 2003, 211, 377–378. [Google Scholar]
- Sánchez-García, D.; Cuesta-Segura, D.A.; Trigos-Peral, G.; Arcos, J.; Catarineu, C.; García-García, F.; Herraiz, J.A.; Espadaler, X.; Gómez, K.; Tinaut, A. Checklist of the Iberian Myrmecofauna, In preparation.
- Tinaut, A.; Reyes-lópez, J. Descripción de una nueva especie para la península ibérica: Temnothorax alfacarensis n. sp. (Hymenoptera, Formicidae). Boln. Asoc. Esp. Ent 2020, 44, 359–378. [Google Scholar]
- Casiraghi, A.; Espadaler, X.; Hidalgo, N.P.; Gómez, K. Two additions to the Iberian myrmecofauna: Crematogaster inermis Mayr, 1862, a newly established, tree-nesting species, and Trichomyrmex mayri (Forel, 1902), an emerging exotic species temporarily nesting in Spain (Hymenoptera, Formicidae). J. Hymenopt. Res. 2020, 78, 57–68. [Google Scholar] [CrossRef]
- Bernadou, A.; Fourcassié, V.; Espadaler, X. A preliminary checklist of the ants Hymenoptera, Formicidae) of Andorra. Zookeys 2013, 277, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Guillem, R.; Bensusan, K. Technomyrmex vexatus (Santschi, 1919) from Gibraltar (Hymenoptera: Formicidae): A new ant species for Europe and genus for Iberia. Myrmecol. News 2008, 11, 21–23. [Google Scholar]
- Guillem, R.; Bensusan, K. Tetramorium parvioculum sp. n. (Formicidae: Myrmicinae) a new species of the T. simillimum group from Gibraltar. Bol. Soc. Entomol. Aragon. 2009, 45, 157–161. [Google Scholar]
- Guillem, R.; Bensusan, K. Two new species of ants (Hymenoptera: Formicidae) for Europe from southern Iberia. Rev. Soc. Gaditana Hist. Nat. 2019, 13, 5–10. [Google Scholar]
- Casevitz-Weulersse, J.; Galkowski, C. Liste actualisée des Fourmis de France (Hymenoptera: Formicidae). Bull. Soc. Entomol. Fr. 2009, 114, 475–510. [Google Scholar]
- Monnin, T.; Espadaler, X.; Lenoir, A.; Peeters, C. Guide des Fourmis de France; Belin: Paris, France, 2013. [Google Scholar]
- Seifert, B. A taxonomic revision of the Palaearctic members of the subgenus Lasius s.str. Soil Org. 2020, 92, 15–86. [Google Scholar]
- Cagniant, H. Liste actualisee des fourmis du Maroc (Hymenoptera: Formicidae ). Myrmecol. Nachr. 2006, 8, 193–200. [Google Scholar]
- Galkowski, C.; Cagniant, H. Contribution à la connaissance des fourmis du groupe angustulus dans le genre Temnothorax (Hymenoptera, Formicidae). Rev. Assoc. Roussill. Entomol. 2017, 26, 180–191. [Google Scholar]
- Taheri, A.; Reyes-López, J. Exotic Ants (Hymenoptera: Formicidae) in Morocco: Checklist, Comments and New Faunistic Data. Trans. Am. Entomol. Soc. 2018, 144, 99–107. [Google Scholar] [CrossRef]
- Taheri, A.; Reyes-López, J.L. Five new records of ants (Hymenoptera: Formicidae) from Morocco. J. Insect Sci. 2015, 15, 37. [Google Scholar] [CrossRef] [Green Version]
- Delabie, J.H.C. Présence de Pheidole teneriffana Forel, 1893, au Maroc (Hym., Formicidae, Myrmicinae). Bull. Soc. Entomol. Fr. 2007, 112, 288. [Google Scholar]
- Cagniant, H. Le Genre Cataglyphis Foerster, 1850 au Maroc (Hyménoptères Formicidae). Orsis 2009, 24, 41–71. [Google Scholar]
- Taheri, A.; Reyes-López, J.; Espadaler, X. Citas nuevas o interesantes de hormigas (Hymenoptera, Formicidae) para Marruecos. Bol. Soc. Entomol. Aragon. 2010, 47, 299–300. [Google Scholar]
- Taheri, A.; Reyes-López, J. Primera cita de Pyramica membranifera (Emery, 1869) (Hymenoptera, Formicidae) y listado actualizado de hormigas alóctonas para Marruecos (Norte de África). Bol. Soc. Entomol. Aragon. 2011, 49, 363. [Google Scholar]
- Guillem, R.; Bensusan, K.; Taheri, A. First record of the ant subfamily Cerapachyinae Forel, 1893 (Hymenoptera: Formicidae) from Morocco. Bull. l’Inst. Sci. 2012, 34, 121–123. [Google Scholar]
- Guillem, R.; Bensusan, K.; Taheri, A. New data on genus Dorylus Fabricius, 1793 (Formicidae, Dorylinae) in Morocco. Bull. l’Inst. Sci. 2015, 37, 47–51. [Google Scholar]
- Borowiec, L. Catalogue of ants of Europe, the Mediterranean Basin and adjacent regions (Hymenoptera: Formicidae); Polish Taxonomical Society: Wroclaw, Poland, 2014; Volume 25, ISBN 9788361764496. [Google Scholar]
- Antwiki. Available online: http://antwiki.org/ (accessed on 20 November 2020).
- Agosti, D.; Collingwood, C.A. A provisional list of the Balkan ants (Hym., Formicidae) and a key to the worker caste. I. Synonymic List. Bull. Soc. Entomol. Fr. 1987, 60, 51–62. [Google Scholar]
- Wagner, H.C.; Seifert, B.; Borovsky, R.; Paill, W. Insight into the ant diversity of the Vjosa valley, Albania (Hymenoptera: Formicidae). Acta ZooBot Austria 2018, 155, 315–321. [Google Scholar]
- Petrov, I.Z. A list of currently known ant species (Formicidae, Hymenoptera) of Serbia. Arch. Biol. Sci. 2004, 56, 121–125. [Google Scholar] [CrossRef]
- Bracko, G. Review of the ant fauna (Hymenoptera: Formicidae) of Croatia. Acta Entomol. Slov. 2006, 14, 131–156. [Google Scholar]
- Bracko, G.; Wagner, H.C.; Schulz, A.; Gioahin, E.; Maticic, J.; Tratnik, A. New investigation and a cheklist of the ants (Hymenoptera: Formicidae) of the republic of Macedonia. North West. J. Zool. 2014, 10, 10–24. [Google Scholar]
- Borowiec, L.; Salata, S. Ants of Greece-additions and corrections (Hymenoptera: Formicidae). Genus 2012, 24, 335–401. [Google Scholar]
- Kiran, K.; Karaman, C. First annotated cheklist of the ant fauna of Turkey (Hymenoptera. Formicidae). Zootaxa 2012, 3548, 1–38. [Google Scholar] [CrossRef]
- Vonshak, M.; Ionescu-Hirsch, A. A checklist of the ants of Israel (Hymenoptera: Formicidae). Isr. J. Entomol. 2009, 39, 33–55. [Google Scholar]
- Mohamed, S.; Zalat, S.; Fadi, H.; Gadalla, S.; Sharaf, M. Taxonomy of ant species (Hymenoptera: Formicidae) collected by pitfall traps from Sinai and Delta region, Egypt. Egypt. J. Nat. Hist. 2001, 3, 40–61. [Google Scholar] [CrossRef]
- Ribera, I. Biogeography and conservation of Iberian water beetles. Biol. Conserv. 2000, 92, 131–150. [Google Scholar] [CrossRef]
- Arnan, X.; Cerdá, X.; Retana, J. Relationships among taxonomic, functional, and phylogenetic ant diversity across the biogeographic regions of Europe. Ecography 2017, 40, 448–457. [Google Scholar] [CrossRef]
- Romo, H.; García-Barros, E. Biogeographic regions of the Iberian Peninsula: Butterflies as biogeographical indicators. J. Zool. 2010, 282, 180–190. [Google Scholar] [CrossRef]
- Carrascal, L.M.; Lobo, J.M. Respuestas a viejas preguntas con nuevos datos: Estudio de los patrones de distribución de la avifauna española y consecuencias para su conservación. In Atlas de las Aves Reproductoras de España; Martí, R., Del Moral, J.C., Eds.; Dirección General de la Conservación de la Naturaleza y Sociedad española de Ornitología: Madrid, Spain, 2003; pp. 651–668. [Google Scholar]
- antmaps.org. Available online: https://antmaps.org/? (accessed on 2 December 2020).
- Sokal, R.R.; Rohlf, F.J. Biometry, 3rd ed.; Freeman and Company: New York, NY, USA, 1995; ISBN 0-7167-2411-1. [Google Scholar]
- Moreno Saiz, J.C.; Donato, M.; Katinas, L.; Crisci, J.V.; Posadas, P. New insights into the biogeography of south-western Europe: Spatial patterns from vascular plants using cluster analysis and parsimony. J. Biogeogr. 2013, 40, 90–104. [Google Scholar] [CrossRef]
- Hammer, D.A.T.; Ryan, P.D.; Hammer, Ø.; Harper, D.A.T. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 178. [Google Scholar]
- Real, R. Tables of significant values of Jaccard’s index of similarity. Miscel-lania Zool. 1999, 22, 29–40. [Google Scholar]
- Baroni-Urbani, C.; Buser, M.W. Similarity of binary data. Syst. Zool. 1976, 25, 251–259. [Google Scholar] [CrossRef]
- Brosens, D.; Vankerhoven, F.; Ignace, D.; Wegnez, P.; Noé, N.; Heughebaert, A.; Borteis, J.; Dekoninck, W. FORMIDABEL: The Belgian Ants Database. Zookeys 2013, 306, 59–70. [Google Scholar]
- Ødegaard, F. New and little known ants (Hymenoptera, Formicidae) in Norway. Nor. J. Entomol. 2013, 60, 172–175. [Google Scholar]
- Moreau, C.S.; Bell, C.D. Testing the museum versus cradle tropical biological diversity hypothesis: Phylogeny, diversification, and ancestral biogeographical range evolution of the ants. Evolution 2013, 67, 2240–2257. [Google Scholar] [CrossRef]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A.; et al. Evolution and the latitudinal diversity gradient: Speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef]
- Willig, M.R.; Kaufman, D.M.; Stevens, R.D. Latitudinal Gradients of Biodiversity: Pattern, Process, Scale, and Synthesis. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 273–309. [Google Scholar] [CrossRef]
- Gómez, K.; Lorite, P.; García, F.; Tinaut, A.; Espadaler, X.; Palomeque, T.; Sanllorente, O.; Trager, J. Differentiating Iberoformica and Formica (Serviformica) with description of the sexual castes of Formica (Serviformica) gerardi Bondroit, 1917 stat. rev. Sociobiology 2018, 65, 463–470. [Google Scholar] [CrossRef] [Green Version]
- O’Regan, H.J. The Iberian Peninsula—Corridor or cul-de-sac? Mammalian faunal change and possible routes of dispersal in the last 2 million years. Quat. Sci. Rev. 2008, 27, 2136–2144. [Google Scholar] [CrossRef]
- Lomolino, M.V.; Riddle, B.R.; Whittaker, R.J. Biogeography: Biological Diversity across Space and Time; Sinauer Associates: Sun-derland, MA, USA, 2017; ISBN 9781605354729. [Google Scholar]
- Jenkins, D.G.; Rinne, D. Red herring or low illumination? The peninsula effect revisited. J. Biogeogr. 2008, 35, 2128–2137. [Google Scholar] [CrossRef]
- González-Taboada, F.; Nores, C.; Álvarez, M.Á. Breeding bird species richness in Spain: Assessing diversity hypothesis at various scales. Ecography 2007, 30, 241–250. [Google Scholar] [CrossRef]
- Martin, J.; Gurrea, P. The Peninsular Effect in Iberian Butterflies (Lepidoptera: Papilionoidea and Hesperioidea). J. Biogeogr. 1990, 85–96. [Google Scholar] [CrossRef]
- Abellán, P.; Svenning, J.-C. Refugia within refugia-patterns in endemism and genetic divergence are linked to Late Quaternary climate stability in the Iberian Peninsula. Biol. J. Linn. Soc. 2014, 113, 13–28. [Google Scholar] [CrossRef] [Green Version]
- Pie, M.R.; Feitosa, R.M. Relictual ant lineages (Hymenoptera: Formicidae) and their evolutionary implications. Myrmecol. News 2016, 22, 55–58. [Google Scholar]
- Puente, A.I.; Altonaga, K.; Prieto, C.E.; Rallo, A. Delimitation of biogeographical areas in the Iberian Peninsula on the basis of Helicoidea species (Pulmonata: Stylommatophora). Glob. Ecol. Biogeogr. Lett. 1998, 7, 97–113. [Google Scholar] [CrossRef]
- Molina-Venegas, R.; Aparicio, A.; Lavergne, S.; Arroyo, J. Climatic and topographical correlates of plant palaeo- and neoendemism in a Mediterranean biodiversity hotspot. Ann. Bot. 2017, 119, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Rahbek, C.; Borregaard, M.K.; Antonelli, A.; Colwell, R.K.; Holt, B.G.; Nogues-Bravo, D.; Rasmussen, C.M.Ø.; Richardson, K.; Rosing, M.T.; Whittaker, R.J.; et al. Building mountain biodiversity: Geological and evolutionary processes. Science 2019, 365, 1114–1119. [Google Scholar] [CrossRef]
- Fernández Escudero, I.; Tinaut, A.; Ruano, F. Ovarian Maturation under Cold Winter Conditions in a High-Mountain Ant (Hymenoptera: Formicidae). Environ. Entomol. 1997, 26, 1373–1377. [Google Scholar] [CrossRef]
- Catarineu, C.; Barberá, G.G.; Reyes-López, J.L. A new ant species, Temnothorax ansei sp.n. (Hymenoptera: Formicidae) from the arid environments of South-Eastern Spain. Sociobiology 2017, 64, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Tinaut, A. Teleutomyrmex kutteri, spec. nov. A new species from Sierra Nevada (Granada, Spain). Spixiana 1990, 13, 201–208. [Google Scholar]
- Reyes Lopez, J.; Benavente Martínez, A. Nueva cita de Teleutomyrmex kutteri Tinaut, 1990 (Hym. Formicidae) para la Península Ibérica. Bol. Soc. Entomol. Aragon. 2011, 49, 206. [Google Scholar]
- Lebas, C.; Galkowski, C.; Wegnez, P.; Espadaler, X.; Blatrix, R. The exceptional diversity of ants on mount Coronat (Pyrénées-Orientales), and Temnothorax gredosi (Hymenoptera, Formicidae) new to France. Rev. Assoc. Roussill. Entomol. 2015, 24, 24–33. [Google Scholar]
- García García, F.; Espadaler Gelabert, X.; Cuesta-Segura, A.D.; Sánchez-García, D. Primera cita ibérica para Temnothorax conatensis Galkowski & Lebas, 2016, y actualización de la distribución para Temnothorax grouvellei (bondroit, 1918) (Hymenoptera: Formicidae). Iberomyrmex 2018, 10, 22–27. [Google Scholar]
- Azcárate, F.M.; Rota, C.; Hevia, V.; Silvestre, M.; Tinaut, A.; Ruano, F.; Seoane, J.; Martín Azcárate, F.; Rota, C.; Hevia, V.; et al. Primera cita de Rossomyrmex minuchae Tinaut, 1981 (Hymenoptera, Formicidae) en el Sistema Central (España). Bol. Asoc. Esp. Entomol. 2016, 40, 535–537. [Google Scholar]
- Reyes-López, J.L.; Obregón Romero, R.; López Tirado, J. Nuevo registro de Myrmoxenus bernardi (Espadaler, 1982) (Hymenoptera: Formicidae) para la península ibérica. Bol. Assoc. Esp. Entomol. 2012, 36, 427–432. [Google Scholar]
- Tinaut, A.; Ruano, F.; Martínez, M.D. Biology, distribution and taxonomic status of the parasitic ants of the Iberian Peninsula (Hymenoptera: Formicidae, Myrmicinae). Sociobiology 2005, 46, 1–42. [Google Scholar]
- Ortuño, V.M.; Gilgado, J.D.; Tinaut, A. Subterranean ants: The case of Aphaenogaster cardenai (Hymenoptera: Formicidae). J. Insect Sci. 2014, 14, 212. [Google Scholar] [CrossRef] [Green Version]
- Tinaut, A.; Barranco, P.; Fernández, D. Nuevas localidades para Aphaenogaster cardenai Espadaler, 1981 (Hymenoptera, Formicidae). Bol. Asoc. Esp. Entomol. 2015, 39, 421–424. [Google Scholar]
- Blaimer, B.B.; Brady, S.G.; Schultz, T.R.; Lloyd, M.W.; Fisher, B.L.; Ward, P.S. Phylogenomic methods outperform traditional multi-locus approaches in resolving deep evolutionary history: A case study of formicine ants. BMC Evol. Biol. 2015, 15, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Radchenko, A.G.; Khomych, M.R. Ants of the extinct genus Cataglyphoides Dlussky, 2008 (Hymenoptera: Formicidae: Formicinae) from the late Eocene European ambers. Invertebr. Zool. 2020, 17, 154–161. [Google Scholar] [CrossRef]
- La Polla, J.S.; Dlussky, G.M.; Perrichot, V. Ants and the fossil record. Annu. Rev. Entomol. 2013, 58, 609–630. [Google Scholar] [CrossRef] [Green Version]
- Prebus, M. Insights into the evolution, biogeography and natural history of the acorn ants, genus Temnothorax Mayr (hymenoptera: Formicidae). BMC Evol. Biol. 2017, 17, 250. [Google Scholar] [CrossRef] [Green Version]
- Jansen, G.; Savolainen, R.; Vepsäläinen, K. Phylogeny, divergence-time estimation, biogeography and social parasite-host relationships of the Holarctic ant genus Myrmica (Hymenoptera: Formicidae). Mol. Phylogenet. Evol. 2010, 56, 294–304. [Google Scholar] [CrossRef] [PubMed]
- Branstetter, M.G. Origin and diversification of the cryptic ant genus Stenamma Westwood (Hymenoptera: Formicidae), inferred from multilocus molecular data, biogeography and natural history. Syst. Entomol. 2012, 37, 478–496. [Google Scholar] [CrossRef]
- Todisco, V.; Gratton, P.; Cesaroni, D.; Sbordoni, V. Phylogeography of Parnassius apollo: Hints on taxonomy and conservation of a vulnerable glacial butterfly invader. Biol. J. Linn. Soc. 2010, 101, 169–183. [Google Scholar] [CrossRef] [Green Version]
- Alonso, J. El yacimiento de ámbar cretácico de Peñacerrada (Álava). Una extraordinaria puerta de acceso al Aptiense (112-114 m.a.). Rev. PH 1999, 29, 142–147. [Google Scholar] [CrossRef]
- Peñalver, E.; Martínez Delclòs, X. Importancia Patrimonial del Arroyo de la Pascueta, un yacimiento de ámbar Cretácico con insectos fósiles en Rubielos de Mora. In El patrimonio paleontológico de Teruel; Meléndez Hevia, G., Mollá Peñalver, E., Eds.; Instituto de Estudios Turolenses: Teruel, Spain, 2002; pp. 201–208. [Google Scholar]
- Peñalver, E.; Martínez-Delclòs, X.; Arillo, A. Yacimientos con insectos fósiles en España. Rev. Esp. Paleontol. 1999, 14, 231–245. [Google Scholar]
- Peñalver, E.; Martínez Delclòs, X. Insectos del Mioceno inferior de Ribesalbes (Castellón, España). Hymenoptera. Treb. Mus. Geol. Barc. 2000, 9, 97–153. [Google Scholar]
- Braga, J.C.; Martín, J.M.; Quesada, C. Patterns and average rates of late Neogene- Recent uplift of the Betic Cordillera, SE Spain. Geomorphology 2003, 50, 3–26. [Google Scholar] [CrossRef]
- Agustí, J.; Antón, M. Memoria de la tierra: Vertebrados fósiles de la Península Ibérica; Ediciones del Serbal: Barcelona, Spain, 1997; ISBN 978-84-7628-195-6. [Google Scholar]
- Goropashnaya, A.V.; Fedorov, V.B.; Seifert, B.; Pamilo, P. Phylogenetic relationships of Palaearctic Formica species (hymenoptera, Formicidae) based on mitochondrial cytochrome b sequences. PLoS ONE 2012, 7, e41697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goropashnaya, A.V.; Fedorov, V.B.; Pamilo, P. Recent speciation in the Formica rufa group ants (Hymenoptera, Formicidae): Inference from mitochondrial DNA phylogeny. Mol. Phylogenet. Evol. 2004, 32, 198–206. [Google Scholar] [CrossRef]
- Gaudeul, M.; Véla, E.; Rouhan, G. Eastward colonization of the Mediterranean Basin by two geographically structured clades: The case of Odontites Ludw. (Orobanchaceae). Mol. Phylogenet. Evol. 2016, 96, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Inda, L.A.; Sanmart, I.; Buerki, S.; Catal, P. Mediterranean origin and Miocene–Holocene Old World diversification of meadow fescues and ryegrasses (Festuca subgenus Schedonorus and Lolium). J. Biogeogr. 2014, 41, 600–614. [Google Scholar] [CrossRef]
- Wheeler, W.M. The Ants of the Baltic Amber; Teubner, B.G., Ed.; BG Teubner: Berlin, Germany, 1915. [Google Scholar]
- Dlussky, G.M. Genera of ants (Hymenoptera: Formicidae) from Baltic amber (in russian). Paleontol. J. 1997, 6, 50–62. [Google Scholar]
- Rust, J.; Andersen, N.M. Giant ants from the Paleogene of Denmark with a discussion of the fossil history and early evolution of ants (Hymenoptera: Formicidae). Zool. J. Linn. Soc. 1999, 125, 331–348. [Google Scholar] [CrossRef]
- de Andrade, M.L. Fossil Odontomachiti Ants from the Dominican republic (Amber Collection Stuttgart: Hymenoptera, Formicidae. VII: Odontomachiti); Staatliches Museum für Naturkunde: Berlin, Germany, 1994. [Google Scholar]
- Tinaut, A. El género Amblyopone Erichson en la península ibérica (Hymenoptera, Formicidae). Miscel.lania Zool. 1988, 12, 189–193. [Google Scholar]
- Tinaut, A.; Bensusan, K. Second record of Amblyopone impressifrons (Emery, 1869) (Formicidae, Amblyoponinae) for Iberia, with some coomments on the genus. Bol. Assoc. Esp. Entomol. 2011, 35, 509–514. [Google Scholar]
- Rodríguez-Sánchez, F.; Pérez-Barrales, R.; Ojeda, F.; Vargas, P.; Arroyo, J. The Strait of Gibraltar as a melting pot for plant biodiversity. Quat. Sci. Rev. 2008, 27, 2100–2117. [Google Scholar] [CrossRef]
- Ros-Montoya, S. El Padul (Granada): Presencia de Mamut Lanudo (Mammuthus primigenius Blumenbach) en el Sur de España. Misc. Paleontológica 2005, 6, 275–292. [Google Scholar]
- Ward, P.S.; Brady, S.G.; Fisher, B.L.; Schultz, T.R. Phylogeny and biogeography of dolichoderine ants: Effects of data partitioning and relict taxa on historical inference. Syst. Biol. 2010, 59, 342–362. [Google Scholar] [CrossRef] [Green Version]
- Tinaut, A. Situación taxonómica del género Cataglyphis Förster, 1850 en la Península Ibérica III. El grupo de C. velox Santschi, 1929 y descripción de Cataglyphis humeya, sp. n. (Hymenoptera, Formicidae). Eos 1991, 66, 215–227. [Google Scholar]
- Espadaler, X. Les formigues granivores de la mediterránia Occidental. Treb. Inst. Catalana Hist. Nat. 1981, 9, 39–44. [Google Scholar]
- Salata, S.; Borowiec, L. A taxonomic revision of the genus Oxyopomyrmex André, 1881 (Hymenoptera: Formicidae). Zootaxa 2015, 4025, 1–66. [Google Scholar] [CrossRef] [PubMed]
- Kryukov, A.; Iwasa, M.A.; Kakizawa, R.; Suzuki, H.; Pinsker, W.; Haring, E. Synchronic east-west divergence in azure-winged magpies (Cyanopica cyanus) and magpies (Pica pica). J. Zool. Syst. Evol. Res. 2004, 42, 342–351. [Google Scholar] [CrossRef]
- Tinaut, A. A new species of the genus Rossomyrmex Arnoldi, 1928 from Turkey (Hymenoptera. Formicidae). Graellsia 2007, 63, 135–142. [Google Scholar] [CrossRef] [Green Version]
- Ribera, I.; Blasco-Zumeta, J. Biogeographical links between steppe insects in the Monegros region (Aragon, NE Spain), the eastern Mediterranean, and central Asia. J. Biogeogr. 1998, 25, 969–986. [Google Scholar] [CrossRef]
- Sánchez Piñero, F.; Tinaut, A.; Aguirre-Segura, A.; Miñano, J.; Lencina, J.L.; Ortiz-Sánchez, F.J.; Pérez-López, F.J. Terrestrial arthropod fauna of arid areas of SE Spain: Diversity, biogeography, and conservation | Elsevier Enhanced Reader. J. Arid Environ. 2011, 75, 1321–1332. [Google Scholar] [CrossRef]
- Olivares, F.J.; Barea-Azcón, J.M.; Pérez-López, F.J.; Tinaut, A.; Henares, I. Las mariposas diurnas de Sierra Nevada; de Andalucía, J., Ed.; Consejería de Medio Ambiente: Seville, Spain, 2011. [Google Scholar]
- Tinaut, A. Taxonomic situation of the genus Cataglyphis Förster, 1850 in the Iberian Peninsula II. New position for C. viatica (Fabricius, 1787) and redescription of C. velox Santschi, 1929 stat. n.(Hymenoptera, Formicidae). Eos 1990, 66, 49–59. [Google Scholar]
- Martín, J.M.; Puga-Bernabéu, Á.; Aguirre, J.; Braga, J.C. Miocene Atlantic-Mediterranean seaways in the Betic cordillera (Southern Spain). Rev. Soc. Geol. Esp. 2014, 27, 175–186. [Google Scholar]
- Tinaut, A.; Heinze, J. Wing reduction in ant queens from arid habitats. Naturwissenschaften 1992, 79, 84–85. [Google Scholar] [CrossRef]
- Cerdá, X.; Arnan, X.; Retana, J. Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology? Myrmecol. News 2013, 18, 131–147. [Google Scholar]
Figure 1.
Aprioristic refugium areas considered in the Iberian Peninsula. 1. Cantabric; 2. Pyrenean; 3. Mediterranean coast; 4. Atlantic; 5. Northern plateau; 6. Southern plateau; 7. Guadalquivir Valley; 8. Ebro Valley.
Figure 1.
Aprioristic refugium areas considered in the Iberian Peninsula. 1. Cantabric; 2. Pyrenean; 3. Mediterranean coast; 4. Atlantic; 5. Northern plateau; 6. Southern plateau; 7. Guadalquivir Valley; 8. Ebro Valley.

Figure 2.
Number of species (orange) and endemic species (blue) belonging to each ant subfamily in the Iberian Peninsula (IP).
Figure 2.
Number of species (orange) and endemic species (blue) belonging to each ant subfamily in the Iberian Peninsula (IP).

Figure 3.
Positive correlation between species richness and the number of endemic species in each genera, including only native species (R = 0.73; p < 0.0001; F(1,28) = 33.32).
Figure 3.
Positive correlation between species richness and the number of endemic species in each genera, including only native species (R = 0.73; p < 0.0001; F(1,28) = 33.32).

Figure 4.
Cluster analysis based on the similarity (Jaccard’s Index) of the ant species found in the three adjacent areas (IP, France and Morocco).
Figure 4.
Cluster analysis based on the similarity (Jaccard’s Index) of the ant species found in the three adjacent areas (IP, France and Morocco).

Figure 5.
Percentage of species pertaining to the different chorological categories (S: Southern; N: Northern; X + XS: Endemic; T: Trans-Iberian; MN: Northern Mediterranean; M: Mediterranean; I: Introduced.
Figure 5.
Percentage of species pertaining to the different chorological categories (S: Southern; N: Northern; X + XS: Endemic; T: Trans-Iberian; MN: Northern Mediterranean; M: Mediterranean; I: Introduced.

Figure 6.
Chorological composition of the three more abundant subfamilies. (S: southern species, N: northern species, X + XS: endemic species, T: Trans-Iberian species, MN: north Mediterranean species, M: Mediterranean and I: introduced species).
Figure 6.
Chorological composition of the three more abundant subfamilies. (S: southern species, N: northern species, X + XS: endemic species, T: Trans-Iberian species, MN: north Mediterranean species, M: Mediterranean and I: introduced species).

Figure 7.
Cluster analysis based on the similarity of ant species occurrence (Jaccard’s Index) in the eight refugium areas of the IP, all of them showing significant differences (p ≤ 0.01; Refugium areas 1. Cantabric; 2. Pyrenean; 3. Mediterranean coast; 4. Atlantic; 5. Northern plateau; 6. Southern plateau; 7. Guadalquivir Valley; 8. Ebro Valley).
Figure 7.
Cluster analysis based on the similarity of ant species occurrence (Jaccard’s Index) in the eight refugium areas of the IP, all of them showing significant differences (p ≤ 0.01; Refugium areas 1. Cantabric; 2. Pyrenean; 3. Mediterranean coast; 4. Atlantic; 5. Northern plateau; 6. Southern plateau; 7. Guadalquivir Valley; 8. Ebro Valley).

Figure 8.
Number of species (orange) and endemism (blue) in the different refugium areas of the IP.

Figure 9.
Suggested dispersal routes affecting the myrmecofauna of the IP. (Base map obtained from https://laboratoriorediam.cica.es/VisorRediam/ (Accessed on 2 December 2020)). Junta de Andalucía. Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible (ámbito de Desarrollo Sostenible).
Figure 9.
Suggested dispersal routes affecting the myrmecofauna of the IP. (Base map obtained from https://laboratoriorediam.cica.es/VisorRediam/ (Accessed on 2 December 2020)). Junta de Andalucía. Consejería de Agricultura, Ganadería, Pesca y Desarrollo Sostenible (ámbito de Desarrollo Sostenible).

Figure 10.
Disjunct distribution of the species belonging to the Cataglyphis altisquamis group (modified from [149]; base map as in Figure 9).

Figure 11.
Disjunct distribution range of the species belonging to genus Goniomma. Note the presence of one species in Israel (citations from [71] are included; base map as in Figure 9).


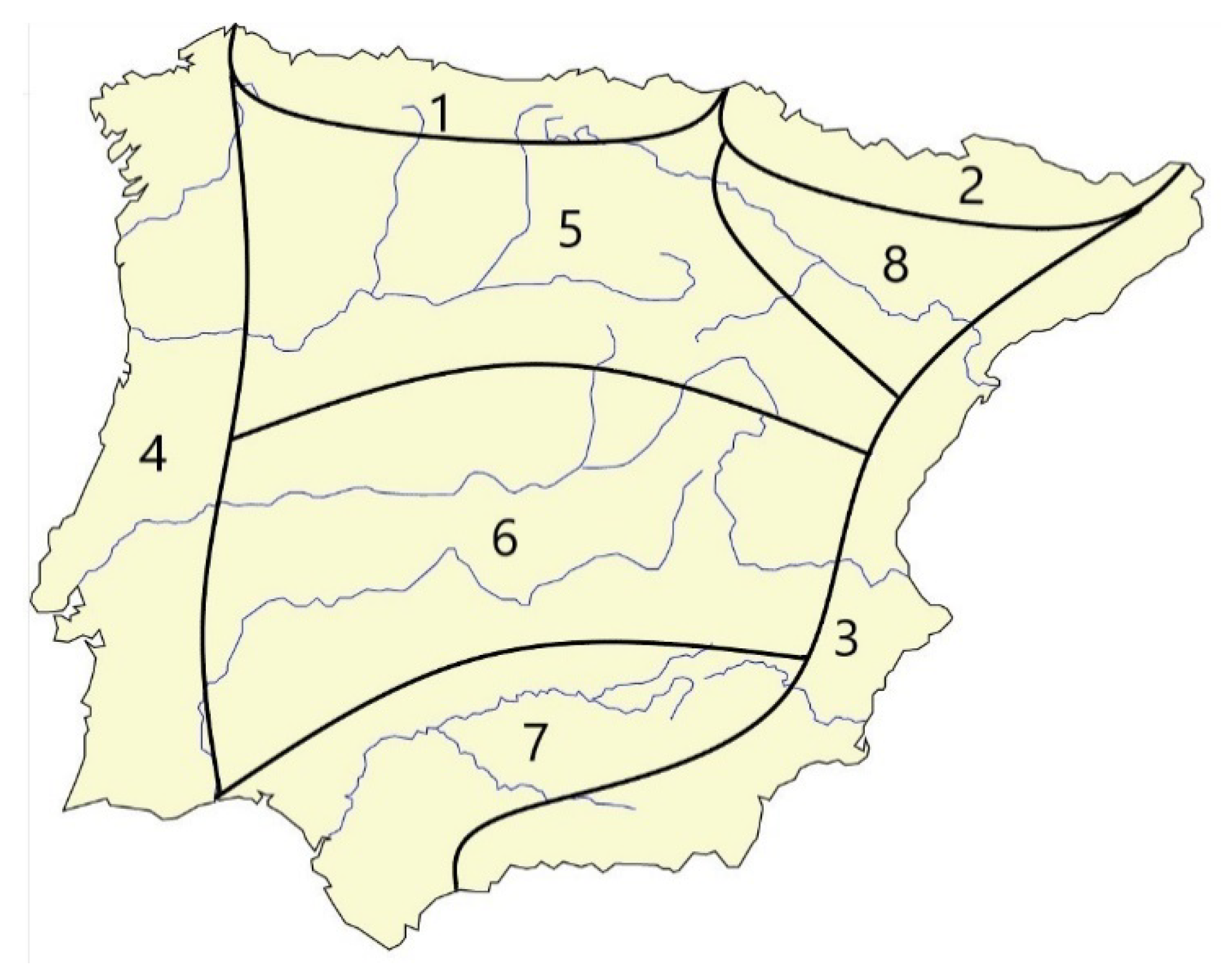{kind=link}
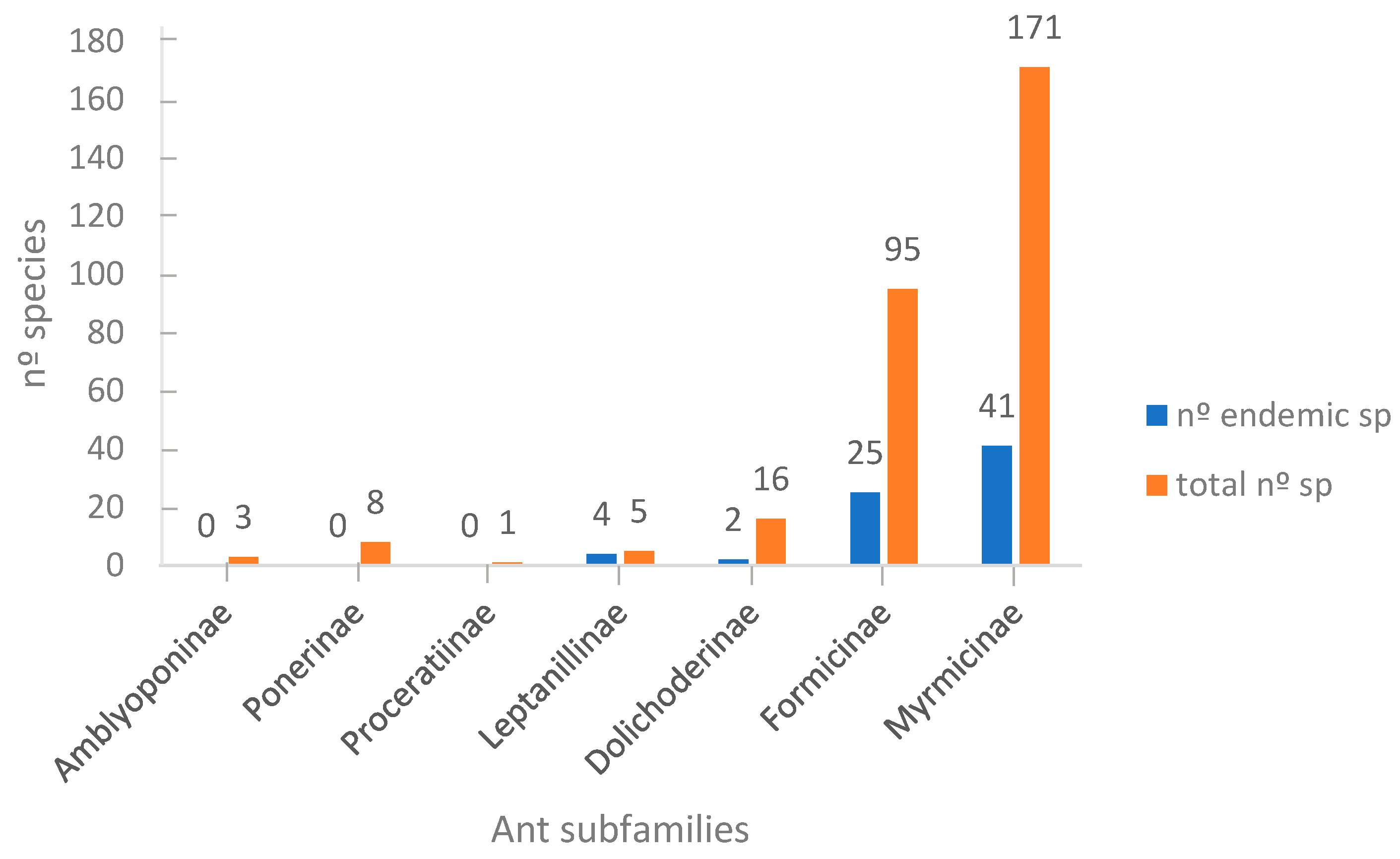{kind=link}
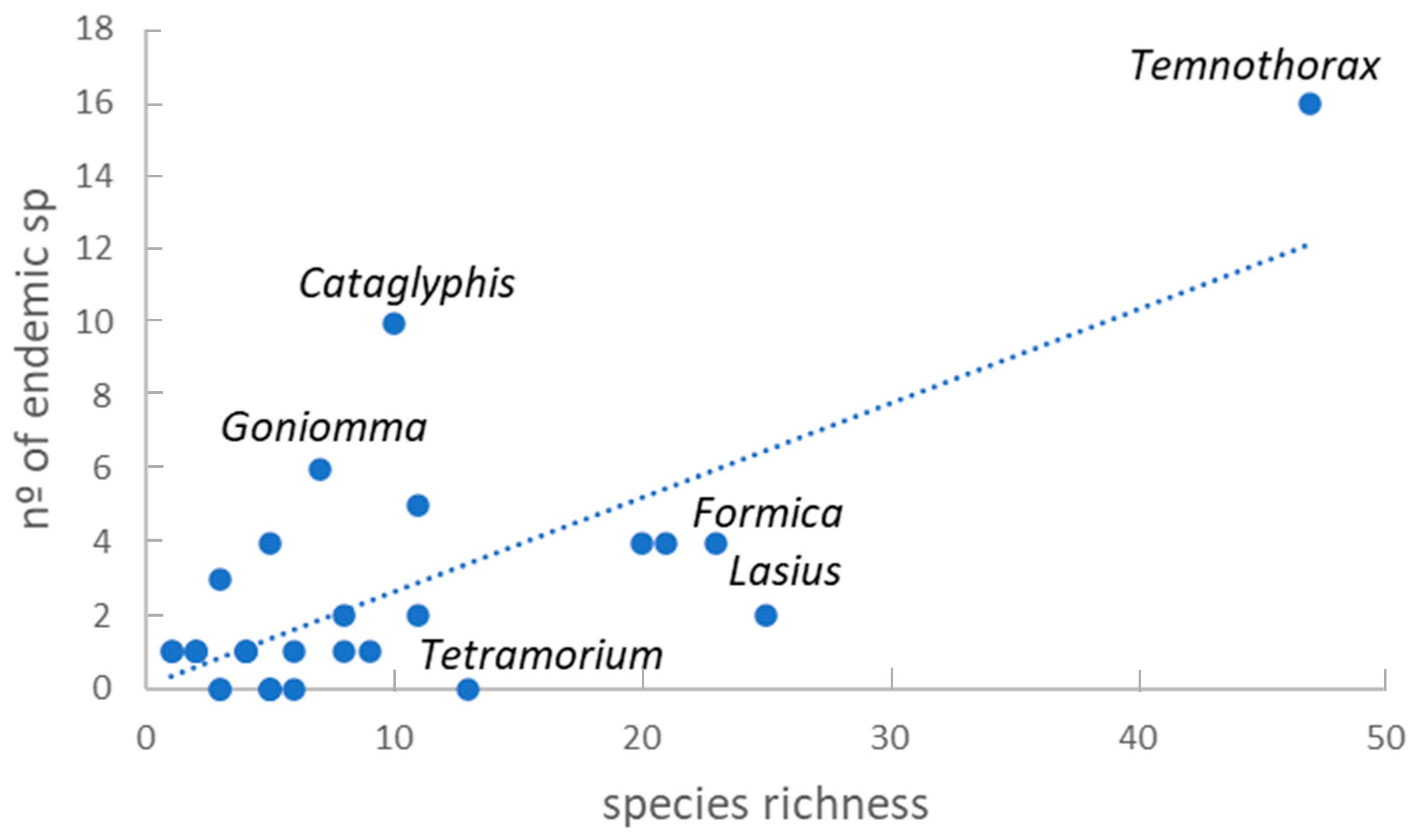{kind=link}
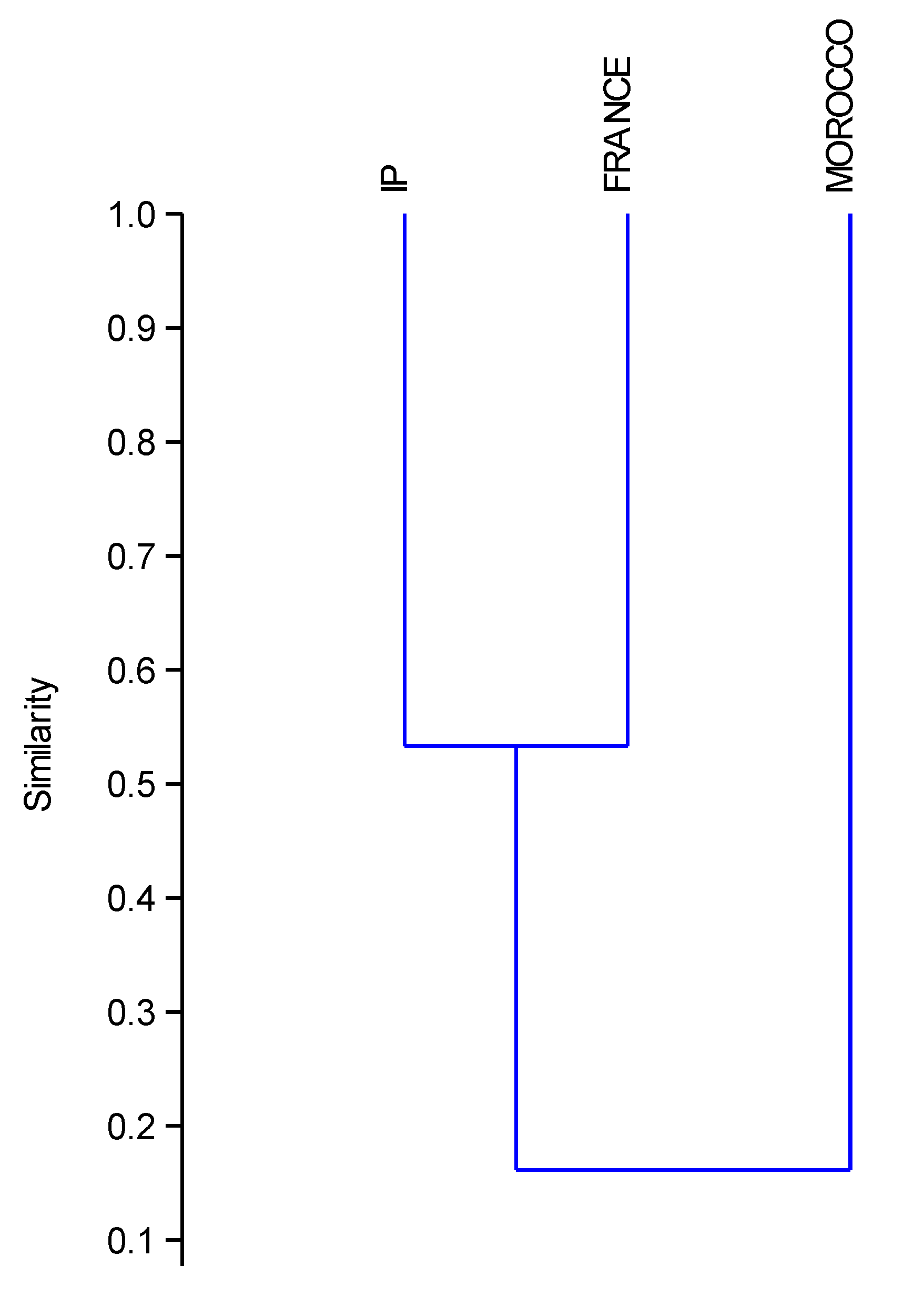{kind=link}
{kind=link}
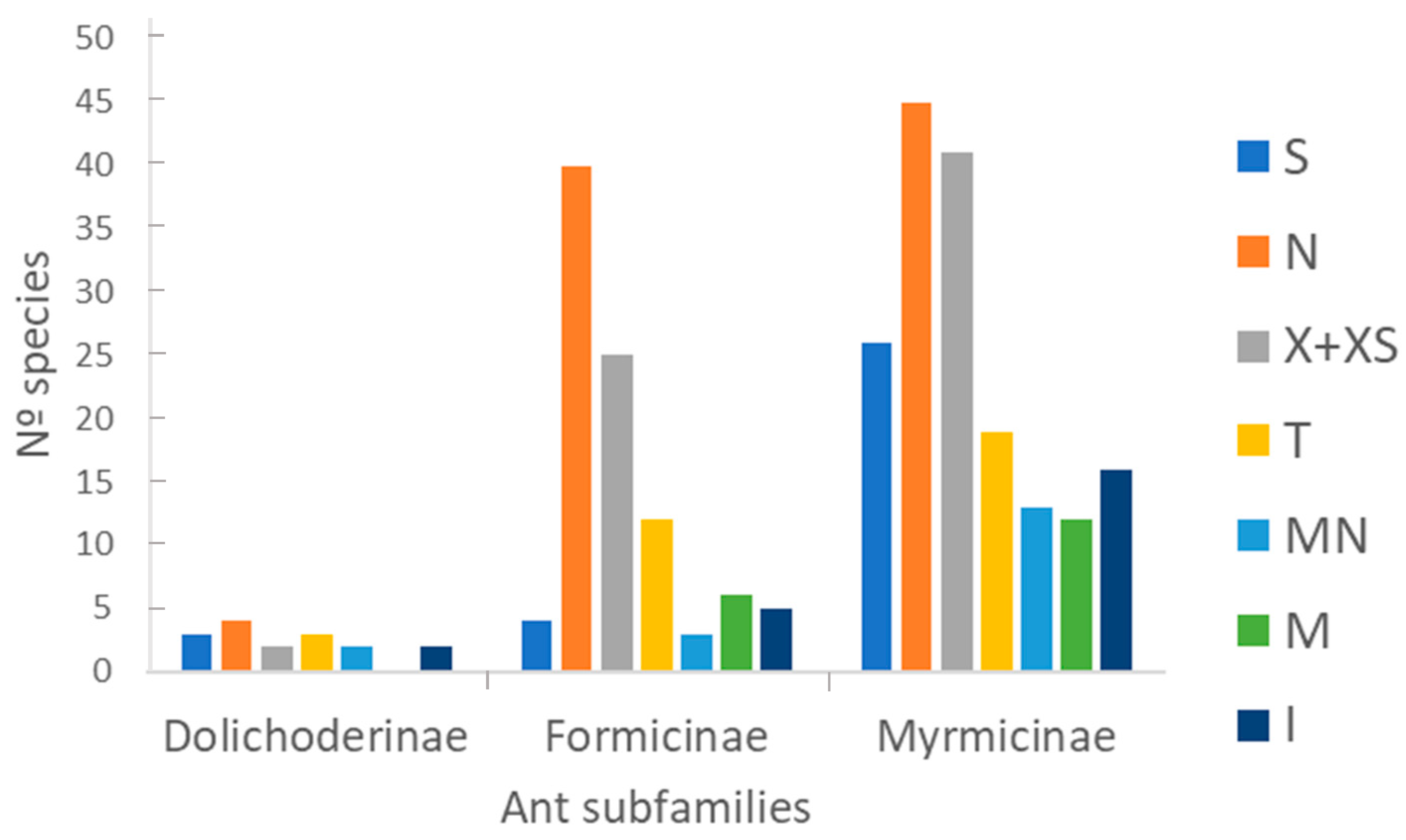{kind=link}
{kind=link}
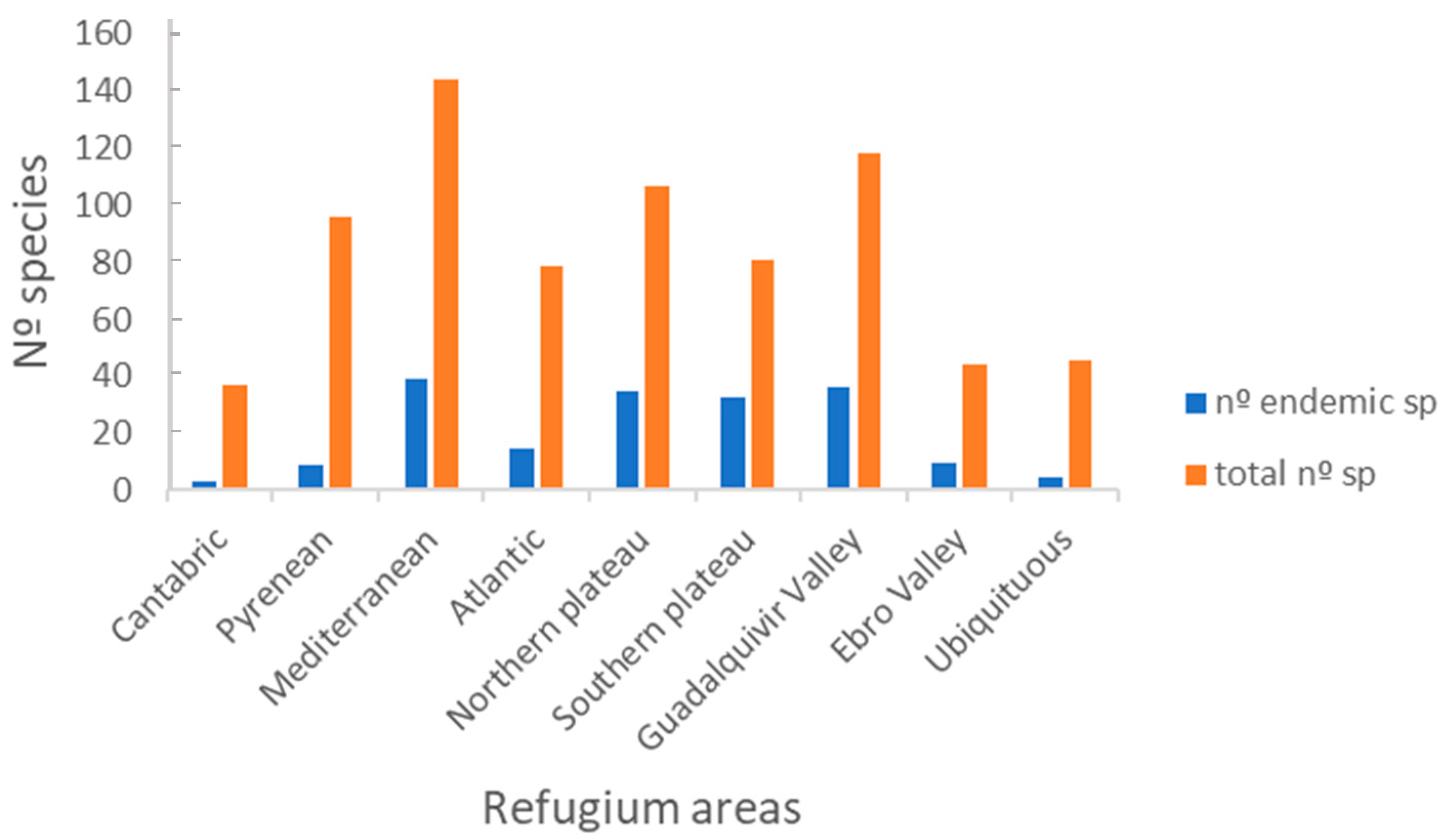{kind=link}
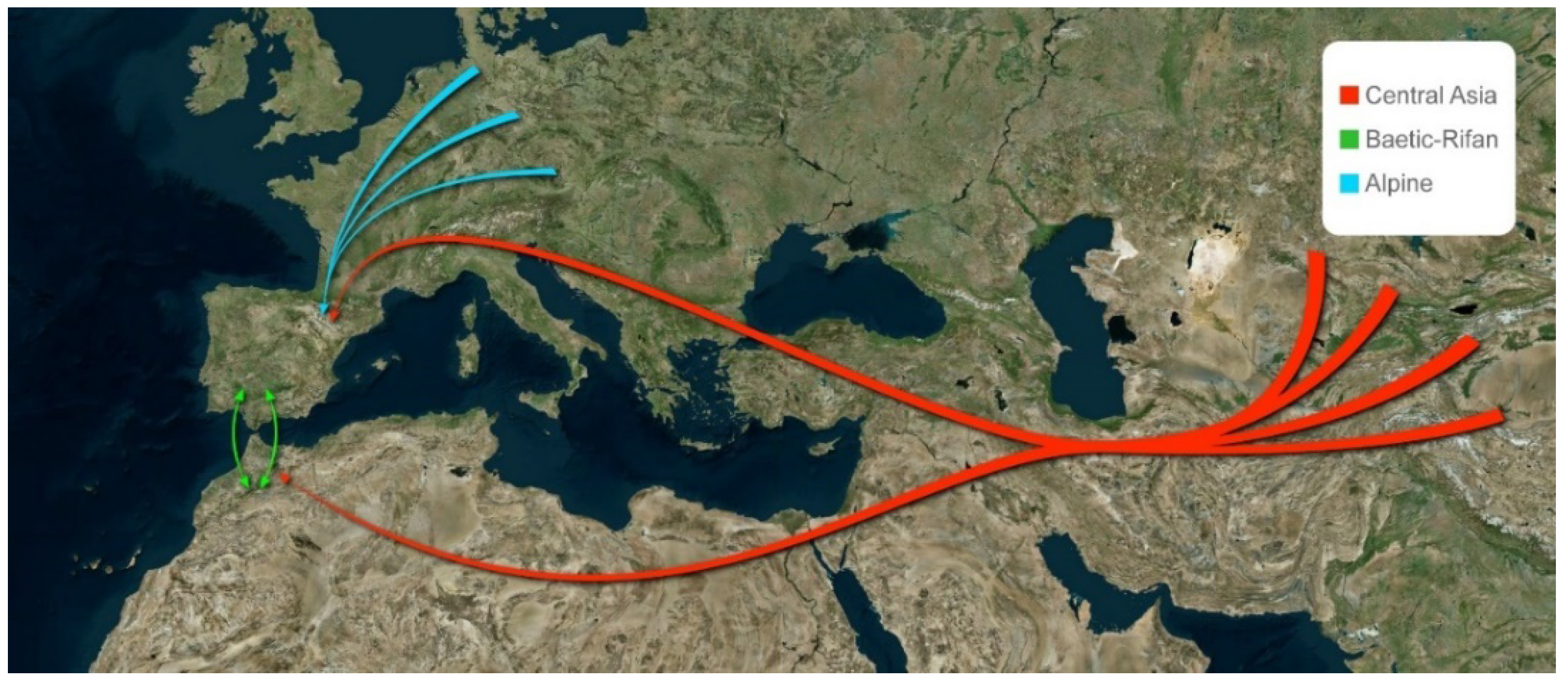{kind=link}
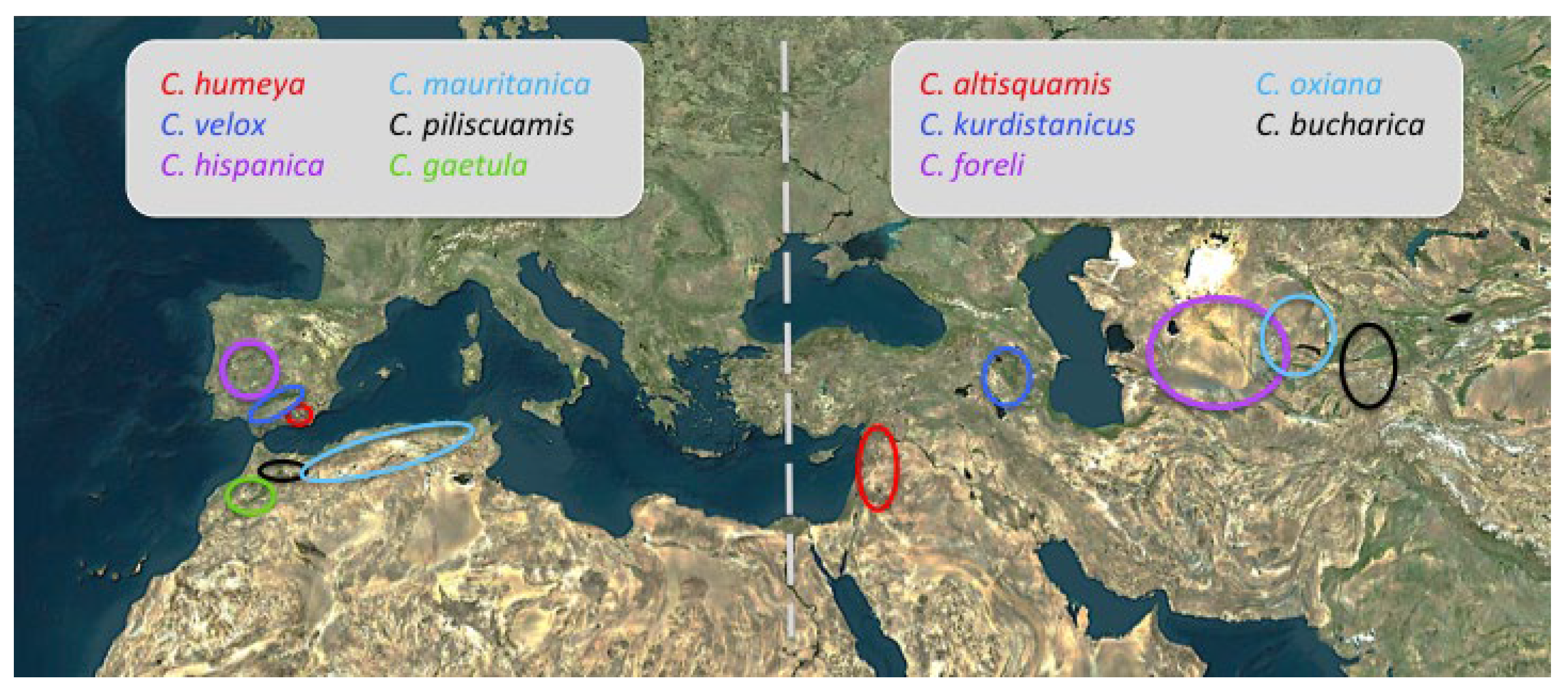{kind=link}
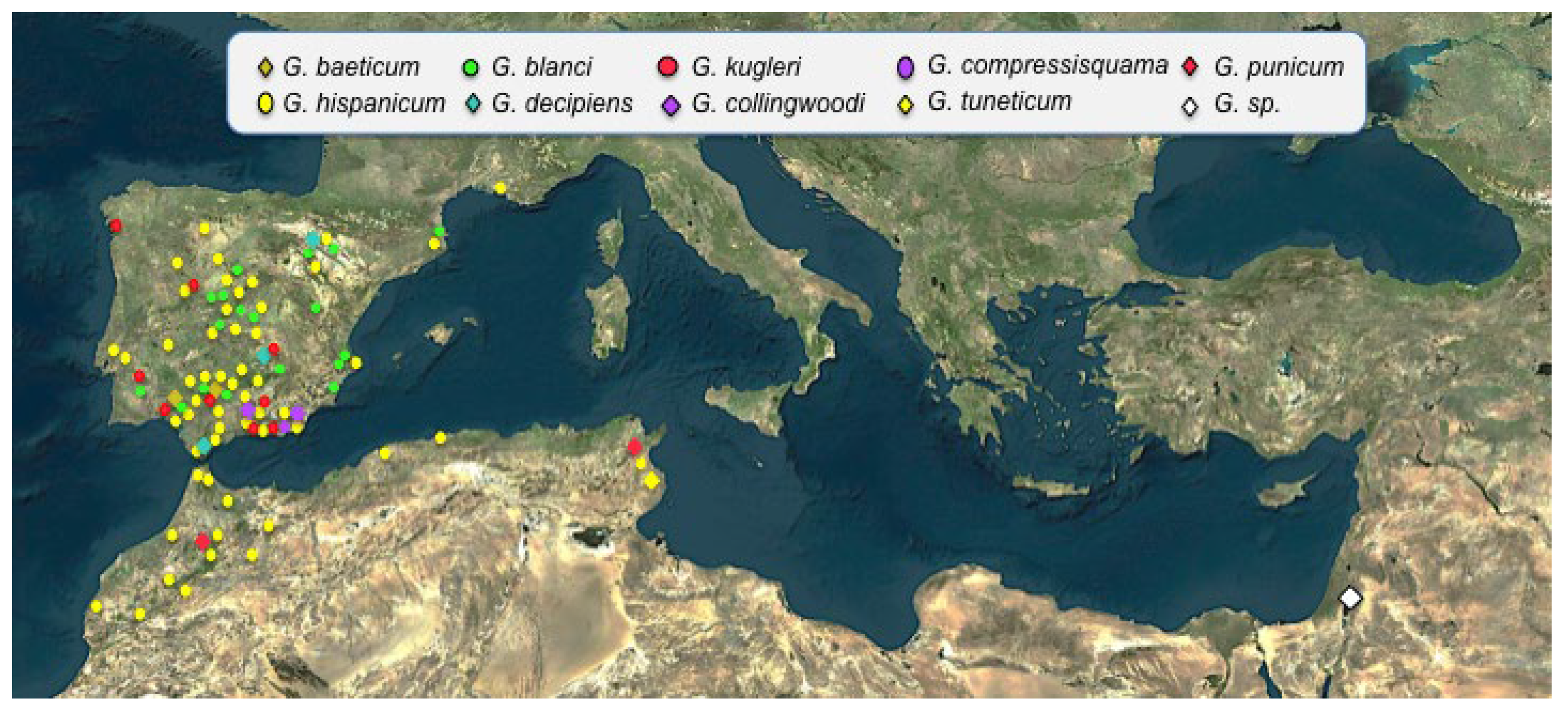{kind=link}
Table 1.
List of Mediterranean countries and number of cited species.
| Country/Region | Nº Species | Reference |
|---|---|---|
| EUROPE | 622 | [67] |
| IP | 299 | present paper |
| FRANCE | 211 | present paper |
| ITALY | 187 | [68] |
| BALKANS | 319 | [69] |
| ALBANIA | 79 | [70] |
| SERBIA | 116 | [71] |
| CROATIA | 140 | [72] |
| YUGOSLAVIA (former) | 171 | [69] |
| MACEDONIA | 80 | [73] |
| GREECE | 259 | [74] |
| TURKEY | 286 | [75] |
| ISRAEL | 203 | [76] |
| EGYPT | 97 | [77] |
| TUNISIA & ALGERIA | 180 | [57] |
| MOROCCO | 233 | present paper |
Table 2.
Total shared and unshared species and percentage amongst France (Fr), Iberian Peninsula (IP) and Morocco (Mo).
Table 2.
Total shared and unshared species and percentage amongst France (Fr), Iberian Peninsula (IP) and Morocco (Mo).
| Subfamily | Total Shared sp/% Fr-IP | Total Shared sp/% Fr-Mo | Total Shared sp/%IP-Fr | Total Shared sp/% IP-Mo | Total Shared sp/% Mo-Fr | Total Shared sp/% Mo/IP |
|---|---|---|---|---|---|---|
| Amblyoponinae | 0/0% | 0/0% | 0/0% | 2/67% | 0/0% | 2/67% |
| Cerapachyinae | 0/0% | 0/0% | 0/0% | 0/0% | 0/0% | 0/0% |
| Dorylinae | 0/0% | 0/0% | 0/0% | 0/0% | 0/0% | 0/0% |
| Ponerinae | 7/87.5% | 4/50% | 7/87.5% | 5/63% | 4/80% | 5/100% |
| Proceratiinae | 1/100% | 0/0% | 1/100% | 0/0% | 0/0% | 0/0% |
| Leptanillinae | 1/100% | 1/100% | 1/20% | 1/20% | 1/33% | 1/33% |
| Dolichoderinae | 10/100% | 3/30% | 10/62% | 4/25% | 3/37% | 3/37% |
| Formicinae | 69/90% | 13/17% | 68/72% | 19/20% | 13/21% | 19/31% |
| Myrmicinae | 89/79% | 28/25% | 89/52% | 57/33% | 28/19% | 45/38% |
| Total shared species | 178/84% | 49/23% | 176/59% | 88/29% | 49/21% | 87/37% |
| Total unshared species | 33/16% | 162/77% | 123/41% | 211/17% | 184/79% | 146/63% |
Table 3.
Jaccard’s Index (black numbers) and probability associated to significant differences (blue numbers) amongst the occurrence of species in paired refugium areas of the IP.
Table 3.
Jaccard’s Index (black numbers) and probability associated to significant differences (blue numbers) amongst the occurrence of species in paired refugium areas of the IP.
| Cantabric | Pyrenees | Mediterranean | Atlantic | Northern Plateau | Southern Plateau | Guadalquivir Valley | Ebro Valley | |
|---|---|---|---|---|---|---|---|---|
| Cantabric | 1 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Pyrenees | 0.28 | 1 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Mediterranean | 0.11 | 0.26 | 1 | <0.01 | <0.01 | <0.01 | <0.01 | <0.01 |
| Atlantic | 0.12 | 0.25 | 0.33 | 1 | <0.01 | <0.01 | <0.01 | <0.01 |
| Northern plateau | 0.23 | 0.32 | 0.35 | 0.36 | 1 | 0.01 | <0.01 | <0.01 |
| Southern plateau | 0.1 | 0.22 | 0.3 | 0.37 | 0.4 | 1 | <0.01 | <0.01 |
| Guadalquivir Valley | 0.1 | 0.21 | 0.4 | 0.35 | 0.4 | 0.4 | 1 | <0.01 |
| Ebro Valley | 0.1 | 0.25 | 0.09 | 0.17 | 0.21 | 0.2 | 0.15 | 1 |
Table 4.
Number of ant species occupying an increasing number of refugium areas.
| Nº Occupied Refugium Areas | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 |
|---|---|---|---|---|---|---|---|---|
| Nº species | 86 | 46 | 35 | 32 | 31 | 16 | 6 | 46 |
| Nº endemic sp | 24 | 12 | 10 | 10 | 10 | 2 | 0 | 5 |
Table 5.
List of 21 endemic species appearing in only one refugium area in the IP.
| Pyrenean Area | |
| Myrmicinae | - |
| Myrmica lemasnei | Bernard, 1967 |
| Mediterranean Area | |
| Leptanillinae | - |
| Leptanilla theryi | Forel, 1903 |
| Formicinae | - |
| Camponotus amaurus | Espadaler, 1997 |
| Cataglyphis gadeai | De Haro & Collingwood, 2003 |
| Cataglyphis humeya | Tinaut, 1991 |
| Myrmicinae | - |
| Goniomma collingwoodi | Espadaler, 1997 |
| Goniomma compressisquama | Tinaut, 1994 |
| Messor timidus | Espadaler, 1997 |
| Teleutomyrmex kutteri | Tinaut, 1990 |
| Temnothorax ansei | Catarineu, Barberá & Reyes-López, 2017 |
| Temnothorax crepuscularis | (Tinaut, 1994) |
| Northern Plateau Area | |
| Leptanillinae | - |
| Leptanilla charonea | López, Martínez & Barandica, 1994 |
| Myrmicinae | - |
| Aphaenogaster ulibeli | Gómez & Espadaler, 2018 |
| Myrmoxenus bernardi | (Espadaler, 1982) |
| Southern Plateau Area | |
| Leptanillinae | - |
| Leptanilla plutonia | López, Martínez y Barandica, 1994 |
| Myrmicinae | - |
| Crematogaster fuentei | Menozzi, 1922 |
| Oxyopomyrmex magnus | Salata & Borowiec, 2015 |
| Guadalquivir Valley Area | |
| Formicinae | - |
| Cataglyphis floricola | Tinaut, 1993 |
| Cataglyphis tartessica | Amor & Ortega, 2014 |
| Myrmicinae | - |
| Goniomma baeticum | Reyes & Rodríguez, 1987 |
| Ebro Valley Area | |
| Myrmicinae | - |
| Temnothorax caesari | (Espadaler, 1997) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tinaut, A.; Ruano, F. Biogeography of Iberian Ants (Hymenoptera: Formicidae). Diversity 2021, 13, 88. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020088
AMA Style
Tinaut A, Ruano F. Biogeography of Iberian Ants (Hymenoptera: Formicidae). Diversity. 2021; 13(2):88. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020088
Chicago/Turabian StyleTinaut, Alberto, and Francisca Ruano. 2021. "Biogeography of Iberian Ants (Hymenoptera: Formicidae)" Diversity 13, no. 2: 88. https://0-doi-org.brum.beds.ac.uk/10.3390/d13020088
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.