1. Introduction
The global climate is changing rapidly in recent years. Industrialization, sea level rise, and urbanization often cause a variety of negative impacts on the aquatic ecosystems. Global warming and climate change have been described as the key factors contributing to rising sea levels. The increase in sea level rise has one of the most severe consequences. Consequently, coastal areas such as saltwater intrusion, coastal flooding, and coastal erosion can affect the inland ecosystem. They can also cause changes in the severity and frequency of severe events due to the combined effects of high spring tides, storm surges, surface waves, and river flooding. Zooplankton populations address environmental fluctuation conditions ranging from seasonal and predictable to unusual and unpredictable occurrences [
1]. Salinity typically fluctuates temporarily and is a vital influence on zooplankton’s composition and dynamics in inland coastal systems, as even marginally increased salinity adversely affects species. Salinity rises particularly as a result of marine intrusions in these aquatic systems. In contrast, salinity decreases after heavy rain and the main river inflow [
2,
3].
Furthermore, salt concentrations in freshwater habitats have increased because of industrial activities and urbanization, with resultant impacts on freshwater animals [
4]. Agricultural operations, urbanization, and other coastal growth projects are rapidly taking place in Southeast Asia that can drastically alter the geomorphological structures [
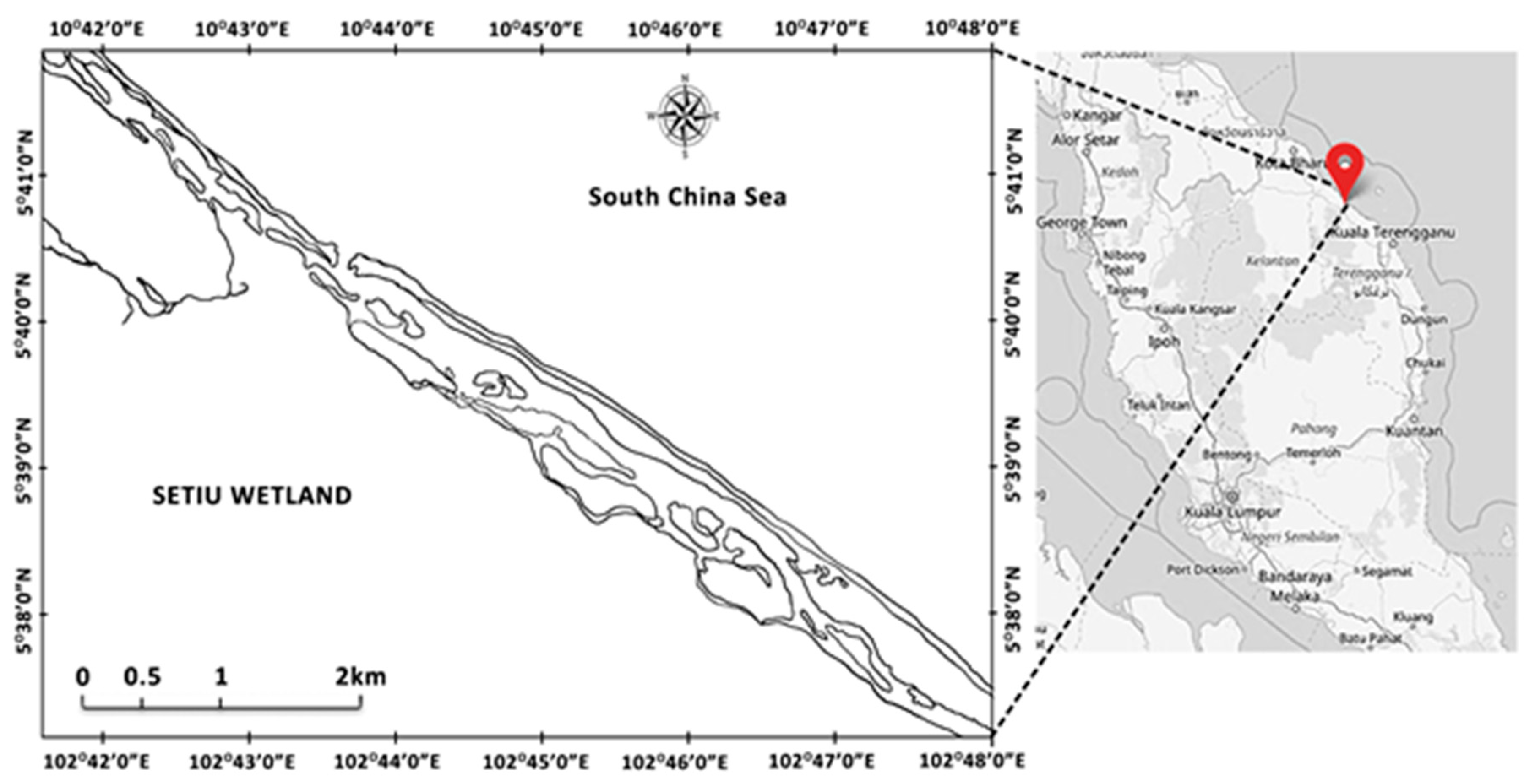
5]. The Setiu Wetland lagoon is considered a major location for brackish water mariculture activities, including finfish cage, pond culture, and shellfish farming [
5]. This anthropogenic interference has both directly and indirectly degraded the natural environment of Setiu Wetland lagoon. Increased seawater levels and coastal erosion have significantly led to increases in salinity [
4], affecting aquatic ecosystems and biodiversity [
5]. Aquatic organisms are sensitive to changes in water quality. Small fish are more opportunistic in nature in that they can adapt and survive in stressful conditions found in shallow lagoons [
6]. Zooplankton is an important community in the aquatic ecosystem for energy transfer from primary producers to fishes [
6,
7]. Factors such as zooplankton and ecosystem species type heavily influence the feeding of small fish [
8]. The ecosystem’s condition affects both foraging efficiency and predation risk and may beneficially increase the former and decrease the latter in fish [
9].
Zooplankton species distribution and abundance are influenced by environmental factors such as water transparency, climate change, and nutritional food content [
10,
11,
12]. The abundance and diversity [
13] of zooplankton and phytoplankton [
4] are affected by salinity and available diets in the environment. Therefore, water salinity shifts also can alter the original taxa composition and ecological processes, such as primary productivity, decomposition, nutrient cycles, and food web function [
14]. Salinity increase in freshwater can reduce zooplankton richness, especially in the cladoceran community, and thus, change the adaptation of the species to a more salt-tolerant species [
5]. Research on cladocerans has been widely conducted due to their ecological importance, sensitivity towards environmental changes, and ease of handling [
15]. Cladocerans are an important group in the zooplankton community with most of the species living in freshwater environments with salinities of <1 [
4]. This is due to osmoregulation adaptations that only allow them to tolerate lower salinity levels [
16]. This represents rapid changes in osmotic hemolymph concentration during salinity acclimation. Rapid osmotic adjustment and the relatively wide tolerance range of cladocera can encourage the colonization of contrasting environments whenever other ecological constraints are less important [
17]. A few cladoceran species have shown an ability to adapt to salinity changes. However, this adaptation can negatively impact the growth and reproduction of the cladoceran species compared to their original habitat. Cladocerans are reported to be abundant in freshwater ecosystems as compared to brackish water environments [
18]. However, a few cladoceran species can live in a saline environment, with brackish water species, tolerating beyond salinity 13 [
16]. The dynamics and abundance of cladocerans are influenced by increased salt levels in freshwater environments [
19]. A previous study by Ismail [
20] showed that the highest salinity at the study area was 17.76 while the normal in-situ parameter was supposed to be in the range of 28–29 °C, 3.8–4.5 mg/L 7.5pH and salinity 2–6, respectively.
Moina sp., commonly referred to as water fleas, are crustaceans within the family of Moinidae, which inhibit freshwater and thrives in both brackish and marine environment [
21]. The reproductive cycle of
Moina sp. has both a sexual and asexual phase. Normally, the population consists of all females that reproduce asexually. Under optimum conditions,
Moina sp. reproduce at only 4 to 7 days of age, with a brood size of 4 to 22 per female [
22]. Broods are produced every 1.5 to 2.0 days [
22], with most females producing 2 to 6 broods during their lifetime [
22]. Under adverse environmental conditions, males are produced, and sexual reproduction occurs, resulting in resting egg (ephippia) production. This case is similar to brine shrimp eggs production. The stimuli for the switch from asexual to sexual reproduction in populations of
Moina sp. are an abrupt reduction in the food supply and significant change in environmental conditions. The density of
Moina sp. cultures routinely reaches densities of 5000 individuals per Litre (19,000/gallon) and therefore, they are well adapted to an intensive culture [
22].
M. macrocopa are rich in protein and nutrients. They are excellent live foods for fish and prawn larvae when compared to other live feeds such as rotifer and
Artemia [
9] However, the fatty acid content of
Moina sp. and other live food species such as copepods varies when they are cultivated with different culture media and different levels of environmental stress [
23,
24].
Studies have shown that lowering ambient temperature tends to increase C20–22 polyunsaturated fatty acid production in planktonic crustaceans [
25]. However, the study on the impact of other environmental parameters on freshwater cladocerans, such as salinity change, is still limited and some quantitative data are only available on the fatty acid profiles of zooplankton at different temperatures but salinity-induced effects on the fatty acid composition of
M. macrocopa have not been evaluated [
26]. Lipids and fatty acids are structural components of cell membranes that play an essential role in growth, survival, and reproduction of aquatic organisms, especially in the early life stages of larvae [
26]. Salinity changes in the freshwater habitat can impact the distribution or, community structure of water fleas, and can affect feeding behavior and also have an indirect impact on nutritional composition. The current study investigated the tolerance of
M. macrocopa to salinity changes. This study’s objectives were to evaluate the ability of
M. macrocopa to adapt to a saline environment up to salinity 15 [
27] and, to investigate the potential of
M. macrocopa to saline tolerance [
16]. Specifically, this study aimed to assess the impact of salinity on fatty acid composition, growth, survival, and reproduction of
M. macrocopa when exposed to different salinity levels.
M. macrocopa is also considered a useful indicating animal to evaluate the impacts of increasing salinity on the aquatic environment.
4. Discussion
Freshwater ecosystems are becoming increasingly threatened, partly due to the rise in the salinity of groundwater (i.e., from seawater intrusion) and water regime modifications, which reduce the frequency of high flow flushing events [
25,
35,
36] These changes will significantly impact both the survival rate [
37] and reproduction [
37,
38] of
M. macrocopa. This study demonstrated how the increase of salinity, especially sodium chloride (NaCl) affects survival and life history traits of a freshwater cladoceran species through an examination of effects on M.
macrocopa, which may be representative the of impact to other members of zooplankton communities. The present study results indicated that the increase in salinity influenced the survival, growth, and reproduction of M.
macrocopa.
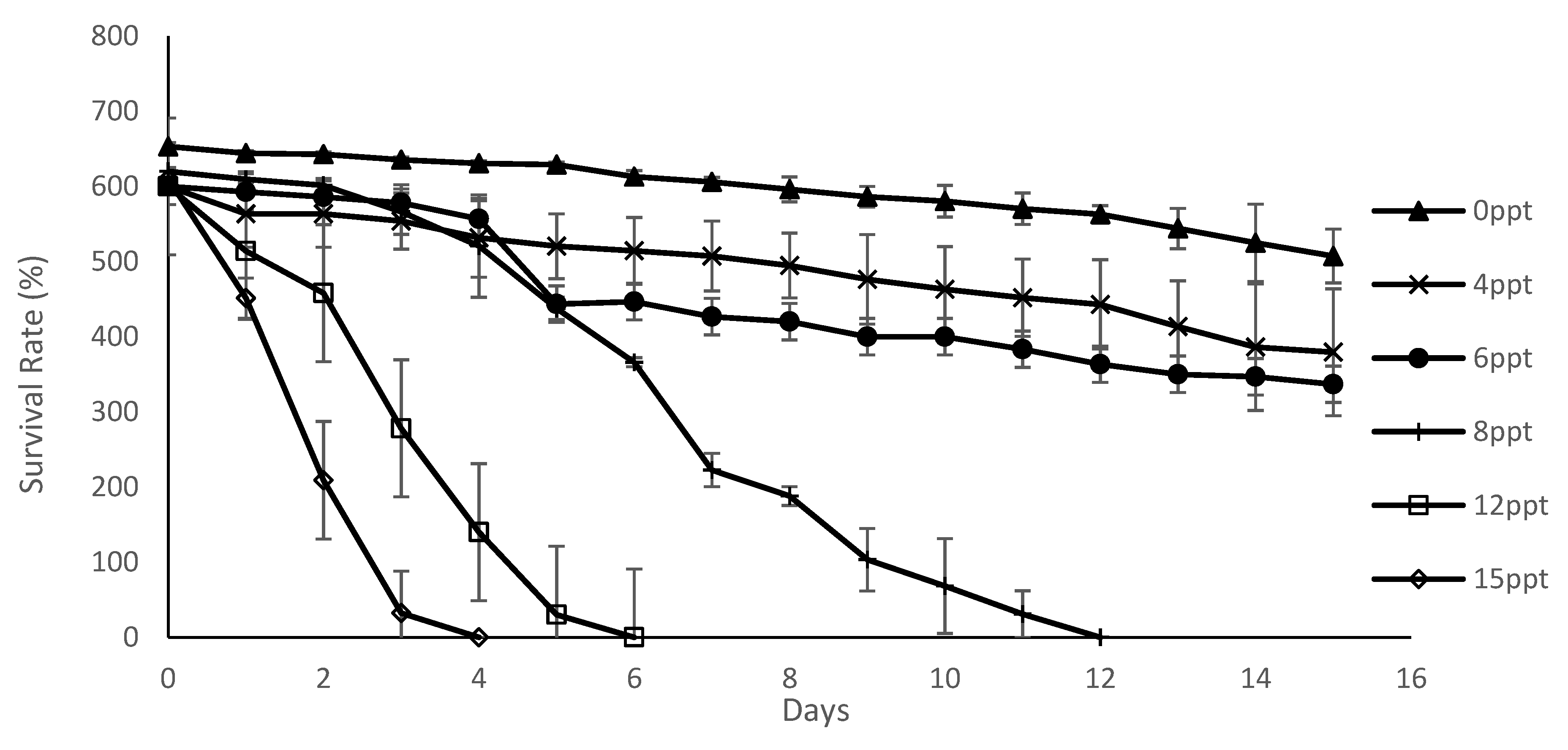
M. macrocopa achieved the highest recorded survival rate at treatment salinity of 0, compared to other salinity range treatments. As indicated by [
39] for another species of cladoceran,
Latonopsis australis, L. australis had the highest survival rate recorded for an environmental salinity of 0, which also decreased, with the increase of the salinity level in the natural environment. Moreover, according to the study of [
40], the survival rate of
Moina eugenie decreased with increased salinity and [
4], further supported this where the survival rate of
Daphnia magna was shown to decrease with increasing salt concentration. It is concluded that the survival rate of
M. macrocopa is undoubtedly, significantly affected by the increase of salinity levels.
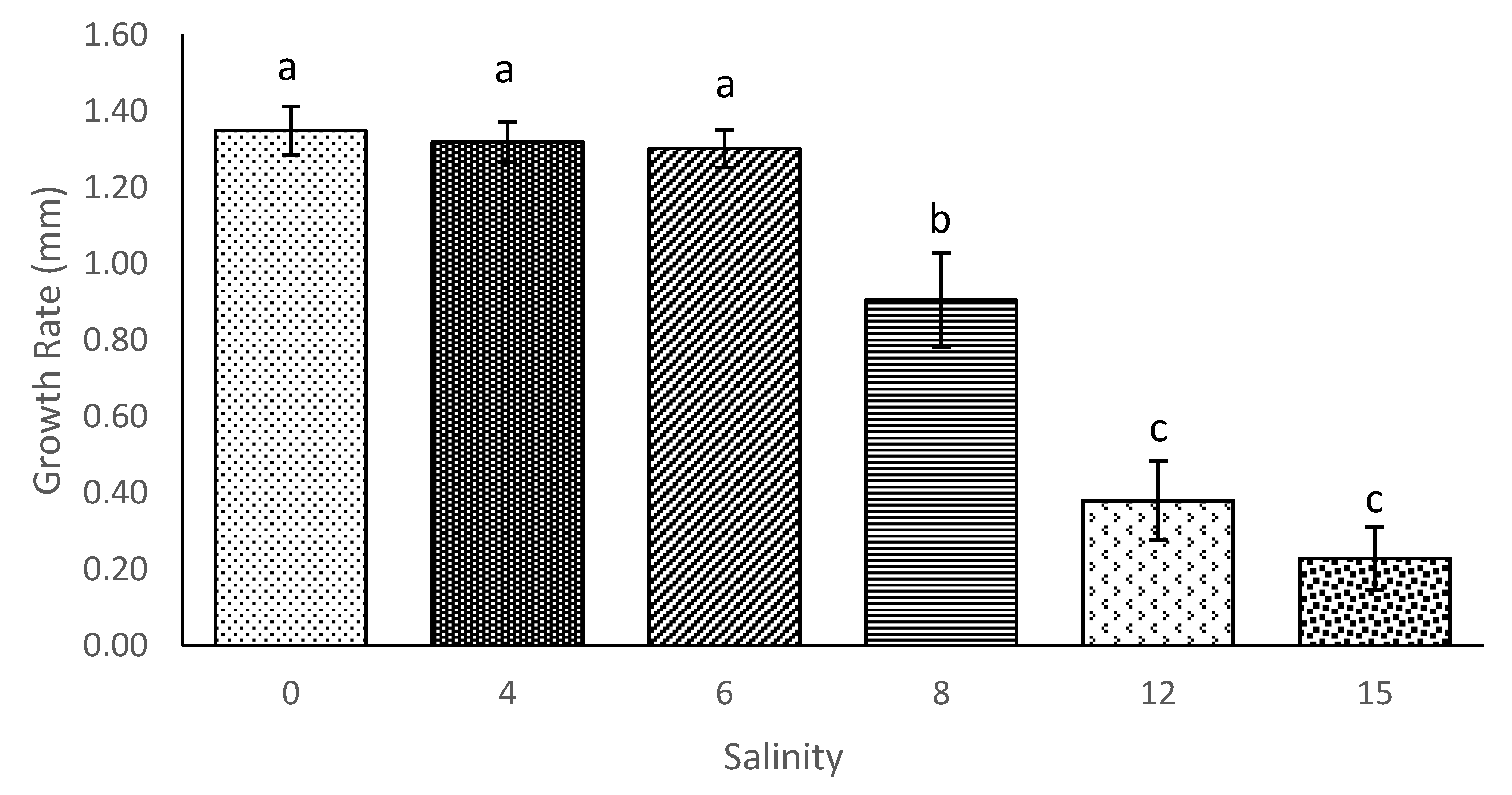
The current study showed that the length of
M. macrocopa does not experience a significant reduction in environments with high salinities, up to a point. The results showed the positive growth of
M. macrocopa in salinity levels from 0 until 6. However, when the salinity was increased to 8, 12, and 15, the growth of
M. macrocopa became slower. The slowest growth was observed at salinity 15, which might have resulted from some regulatory stress that occurred in the culture [
41]. Increased salinity stress can also add additional energy costs for metabolism and osmoregulation to redirect energy from somatic growth. In a stressful environment, animals which spend more energy in reproduction are an adaptation to increase mortality [
12]. This conclusion is also aligned with the findings of [
27] investigating the growth rate of exotic cladoceran,
Daphnia exilis which also did not show any changes under a salinity regime of 6 but started to decline in growth when salinity reached 8 and above.
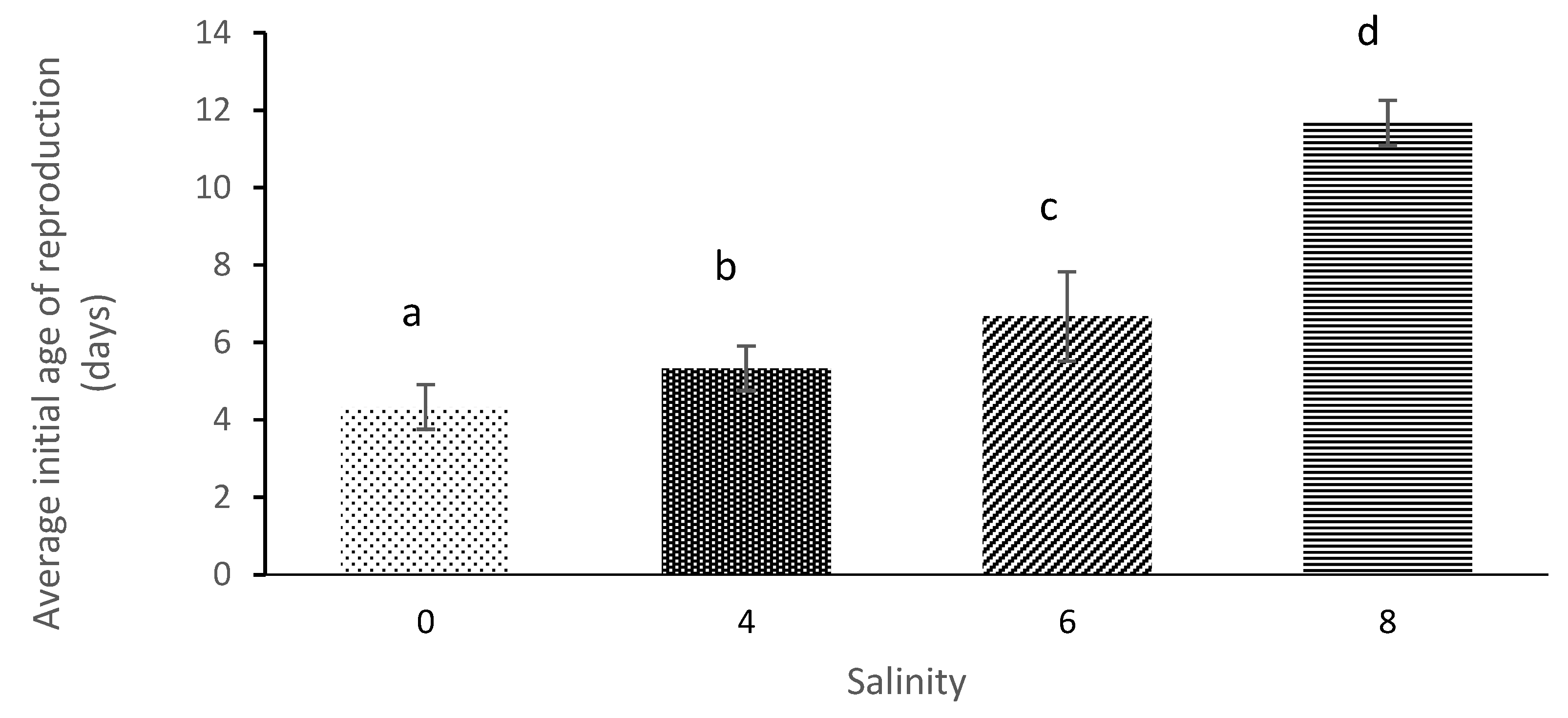
Female reproduction was affected by salinity and the maturation of
M. macrocopa was delayed at the highest salinity. These results are comparable to those obtained by [
42], who noted that the sexual maturation of
M. macrocopa was delayed by 1 day at a salinity level of 5.5 when compared to 0. Additionally, [
4], noted that the reproduction age of
D. magna was delayed from 7 days until 9 days at salinity levels of 0 and 2.66, respectively.
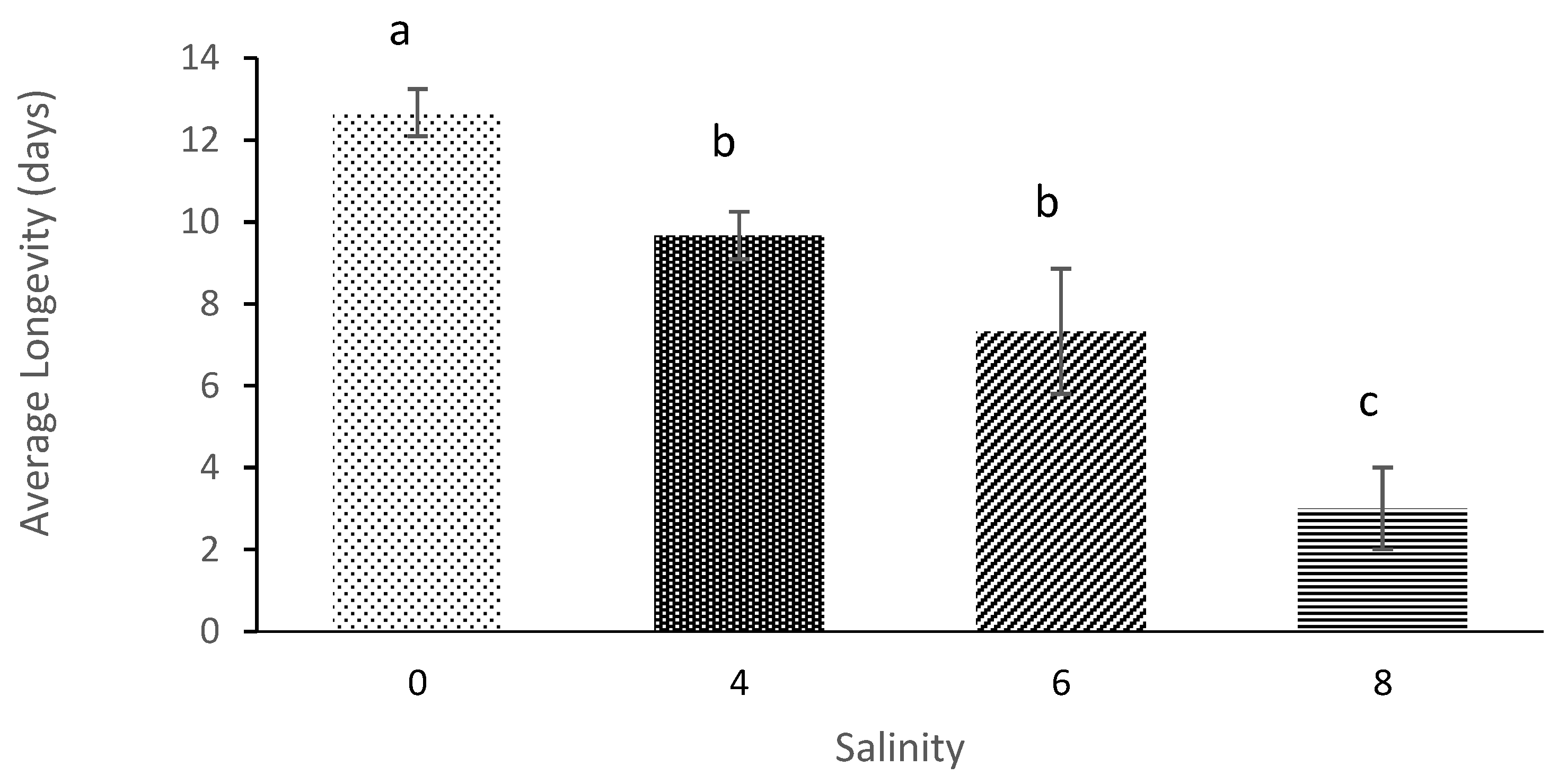
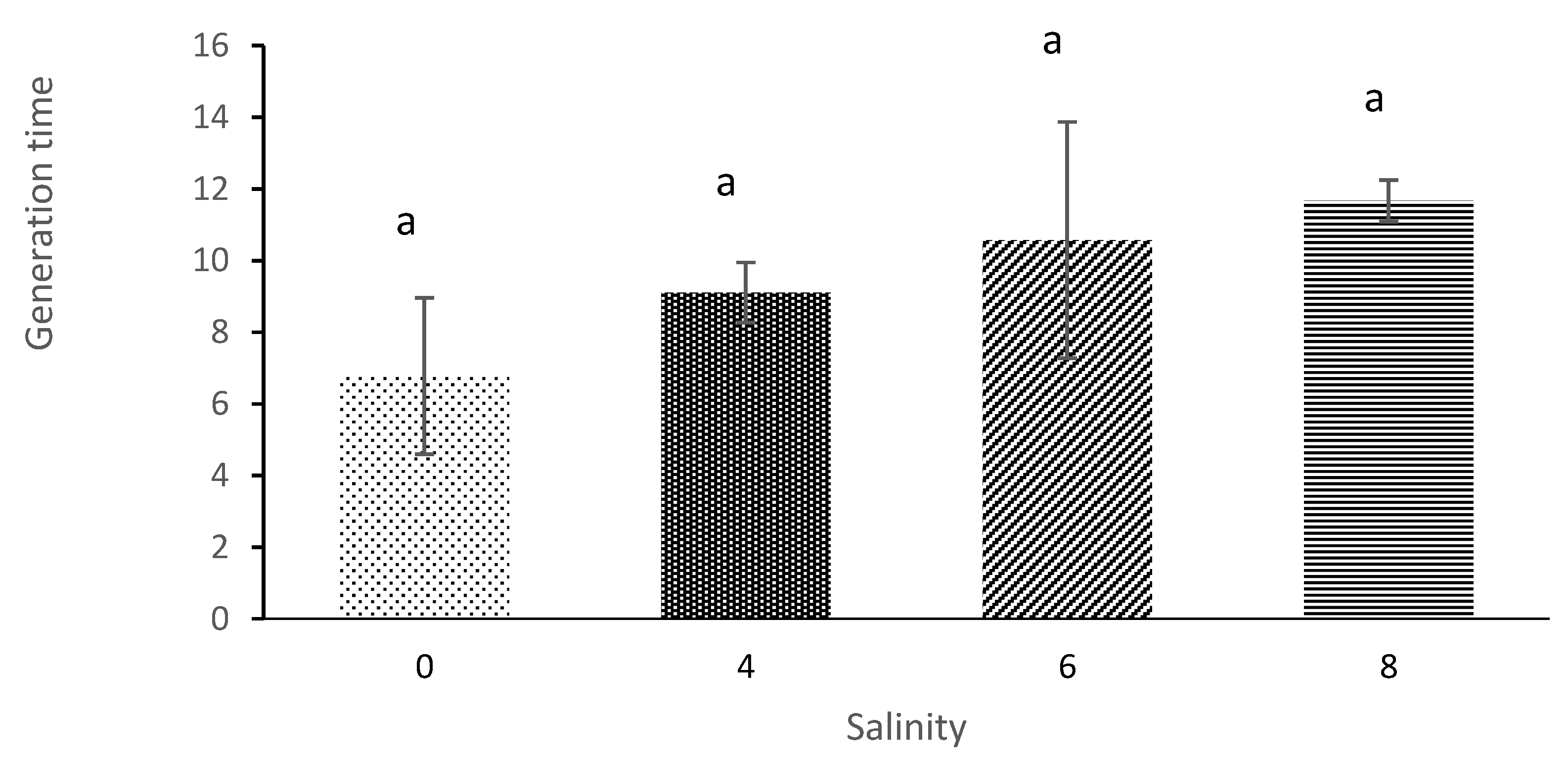
The current study results showed that the average lifespan of
M. macrocopa was 12.3 days of salinity 0 and, 9.6 days at salinity 6. These are similar results to a study by [
43], where the average lifespan of
D. magna was also significantly affected by salt concentration. The results showed that maximum average lifespan (57.7 days) occurred at the salinity of 0. In contrast, the shortest lifespan (25.2 days) occurred at salinity 7. Furthermore, in studies of
Simocephalus vetulus, the results also showed that this cladoceran’s lifespan decreased drastically when the salinity level was increased apart from food impacts that positively influence the lifespan of copepods [
10,
11,
44].
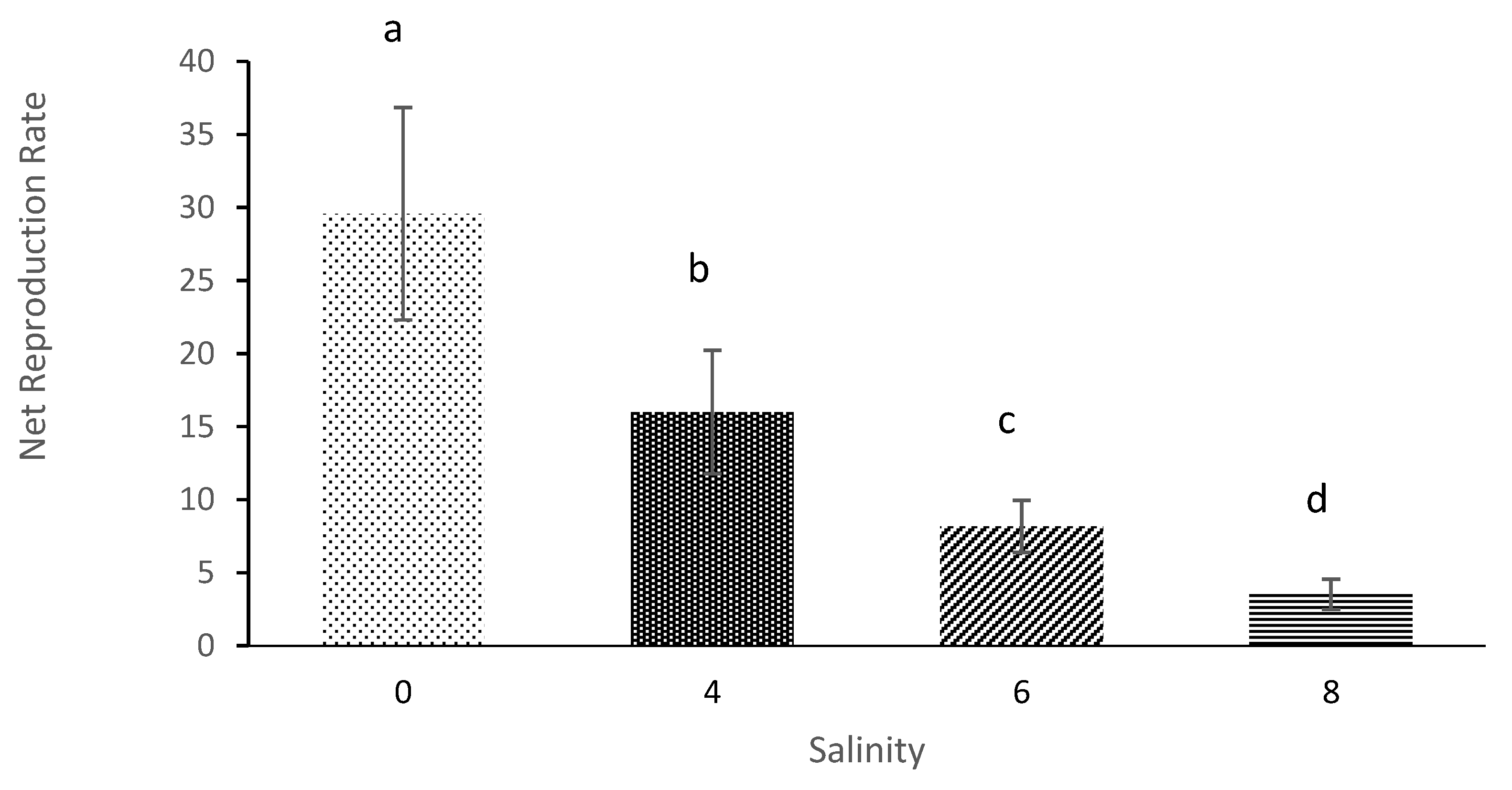
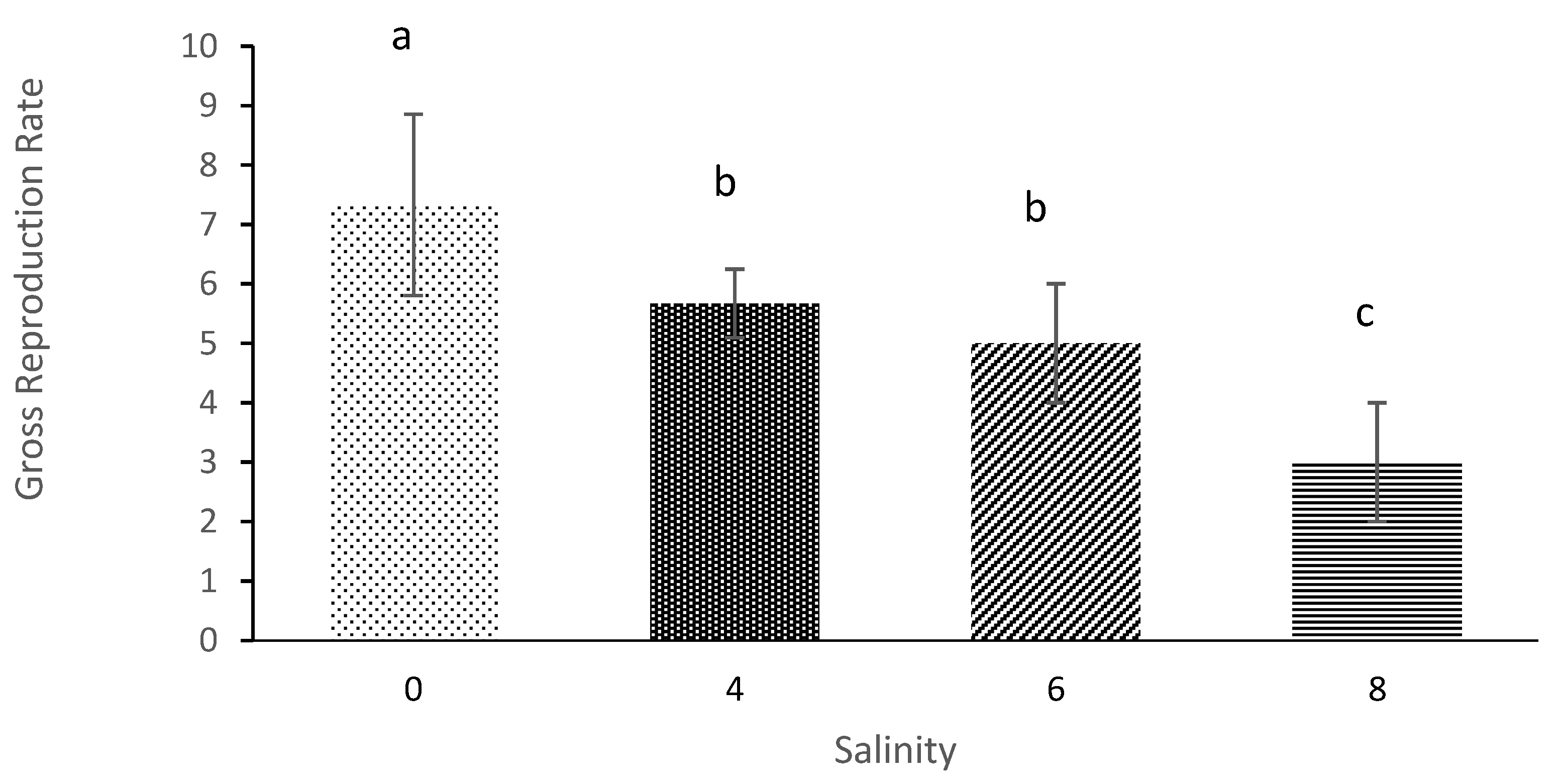
The offspring net and gross reproduction rates of
M. macrocopa were also affected when salinity increased. The previous study also showed that the number of offspring produce was the most affected parameters on the life table as caused by an increase of salinity for
D. magna and
D. longispina [
45]. Furthermore, in an observation-based study by [
39], the findings also showed that the offspring of
Latonopsis australis were significantly influenced by high salinity. It is concluded that salinity clearly impacts reproduction of offspring of this cladoceran.
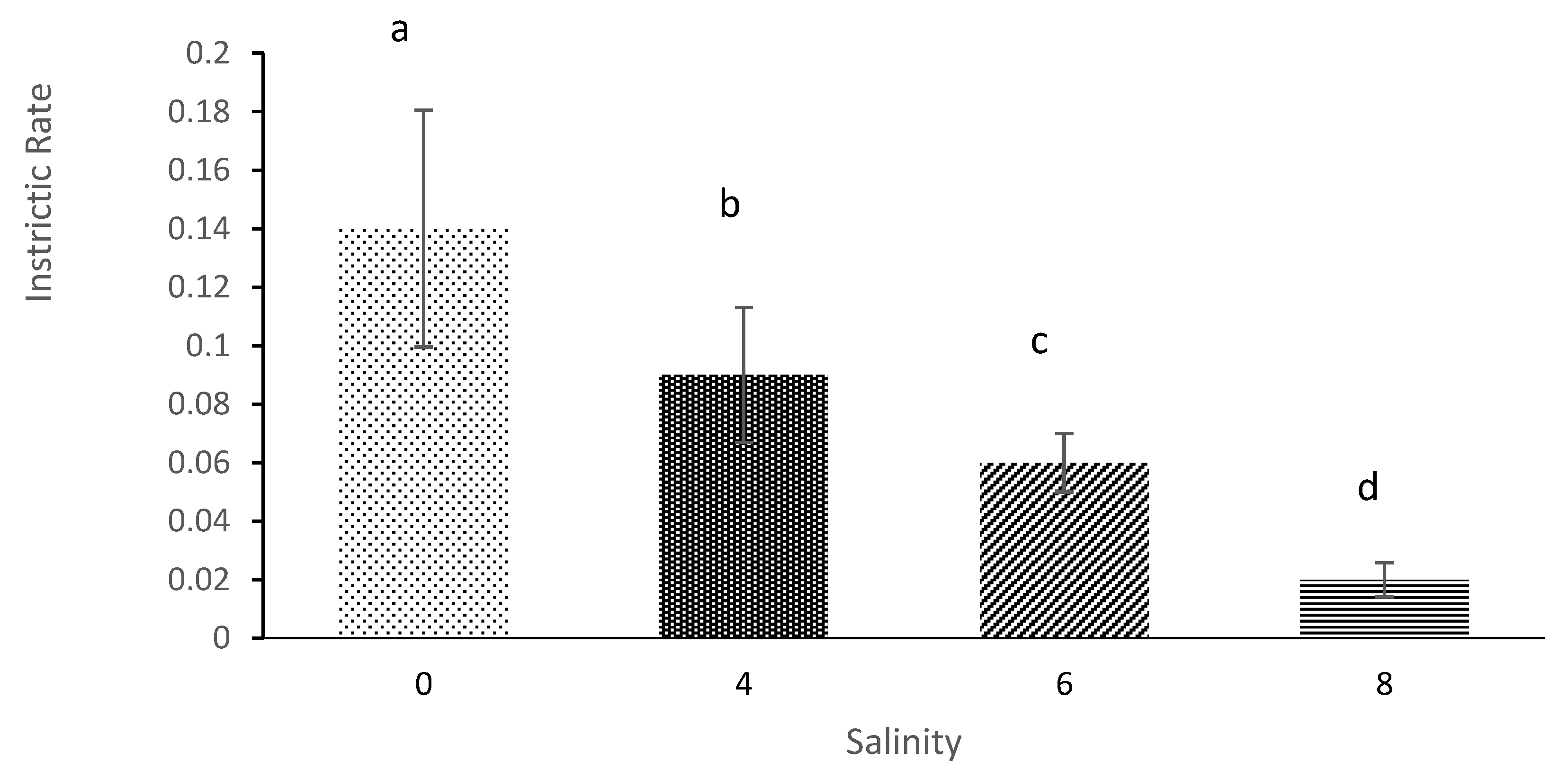
Salinity levels impact the intrinsic rate (births minus deaths) for
M. macrocopa population. This claim was supported in similar research done by [
39] where the intrinsic rate of
Latonopsis australis decreased when the salinity levels of treatments increased. In the current study, life table parameters at salinity more than 8 also cannot be reliably acquired because salinity higher than 8 is beyond the tolerance of
M. macrocopa. [
13]
D. longispina reproduced and lived well in salinity below salinity 5 but then started to decline when salinity levels increased. Thus, the present study concludes that
M. macrocopa can achieve the optimum growth population when salinity is below 6. Zooplankton is sensitive to change in the aquatic environment that can significantly change ambient conditions within the aquatic ecosystem [
46]. The increase of salt concentration can cause the disappearance of species that cannot tolerate highly saline conditions. Meanwhile, salinity is a strong mechanism that can change aquatic communities [
47].
Furthermore, no life table data can be recorded for salinities of 12 and 15 since the
M. macrocopa are dead before being able to reproduce. Therefore, various freshwater invertebrate species (including cladocerans) are more sensitive to NaCl salinity than to the effect produced by the array of chemical compounds present in sea salt [
41]. Higher content of salinity level might be fatal to freshwater cladocerans especially
M. macrocopa. The highest salinity at which life table could be recorded was only at salinity 8. It is strongly supported by [
4], the highest salinity will cause cladoceran stress and mortality. Apart from that,
M. macrocopa that spending most of their life stages under good conditions (food and environment) may adapt their salinity tolerance only with considerable time-lags.
The observations in the current study showed that fatty acid expression is higher at the optimum culture conditions (salinity 0 and 4) compared to other salinity level treatments. This effect is probably due to the minimal stress undergone by the cultured
M. macrocopa at these optimum conditions, which is finally reflected in obtaining a rich fatty acid profile. Similar observations were also reported [
48,
49,
50] where live feed underwent minimal stress during the culture period produced superior survival, growth and reproduction. High polyunsaturated fatty acids achieved by
M. macrocopa might be due to
Nannochloropsis sp. as food sources, which were conducted to imitate feed in the natural environment. The amount of EPA and DHA in algae varies greatly among different algal species and environmental conditions. EPA content in
Nannochloropsis sp. is 23.4 mg g
−1 [
24]. Microalgae such as
Nannochloropsis sp. have received increasing interest as a target live feed for aquatic animals because of the high contents of EPA [
7].
5. Conclusions
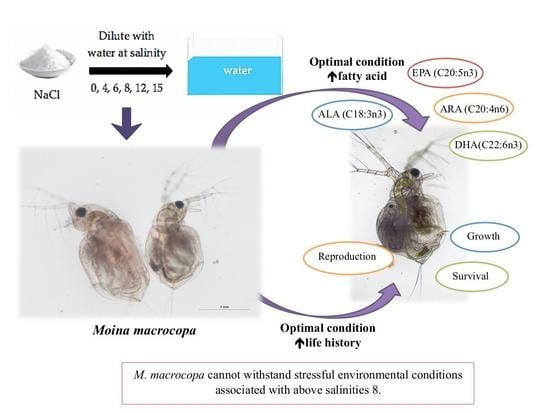
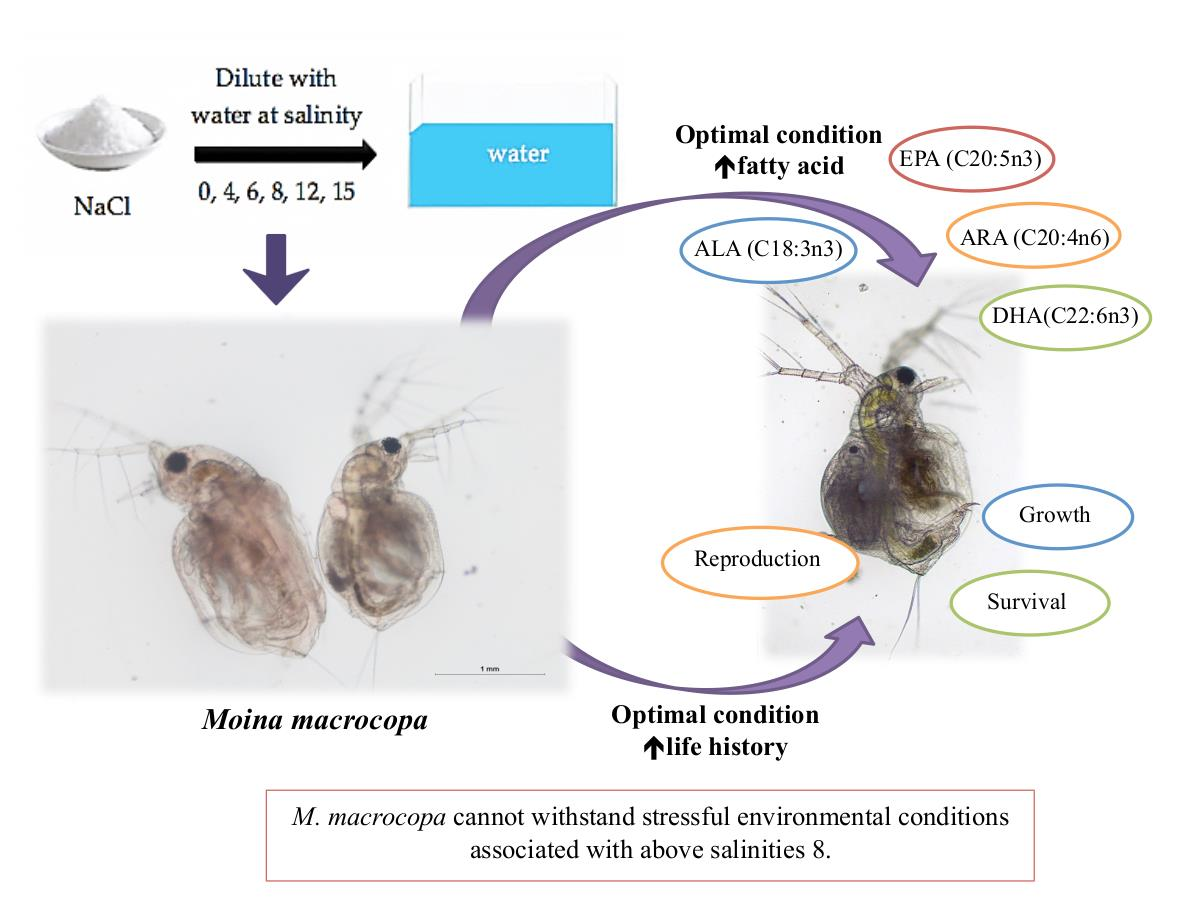
In conclusion, the survival, growth, reproduction rate, and fatty acid profile of M. macrocopa were affected by different salinity treatment ranges.
M. macrocopa produced better growth rate, survival rate, and productivity at the salinity of 0 which is a normal freshwater environment. Salinity at 4 also provided relatively good growth for
M. macrocopa. Most of the present study results showed no significant difference between salinity treatments of 0 and 4. The salinity above 8 was intolerable for
M. macrocopa and to be above the critical salinity maximum (CSMax) or acute phase for cladoceran. Apart from that, fatty acid content also significantly abated with increased salinity levels. The highest content of fatty acid produced was with
M. macrocopa being cultured in optimal salinity levels of 0 and 4, which are considered to be normal in their natural habitat. Some cladocera species’ ability to withstand stressful physiological environments, such as increased salinity, leads to a shelter from severe predation and competition typically found in more complex populations. It also can be expected that cladocera species capable of surviving under these conditions are those competitively disadvantaged or more susceptible to predation. That might be the case for
Moina sp., which can grow at Setiu Wetland during times of significantly increased salinity [
51]. Furthermore, salinity can directly and indirectly affect aquatic organism population dynamics. Small changes in salinity tend to be beneficial under natural conditions, where salinity can directly and indirectly mediate multiple biotic and abiotic processes [
52]. Overall, the result of direct and indirect effects of minor salinity changes under natural conditions, advantaging cladocera’s population growth.
Comparing results in the whole NaCl concentration range tested, we can conclude that the impairment effects on fatty acid content, growth, reproduction, and survival under the current treatments were provoked by salinity stress. Although the result of the present study shows that the survival rate, life history and fatty acids of M. macrocopa were significantly affected by the increase in salinity, future detailed studies are required to understand how freshwater species, including other species of zooplankton which can adapt to a higher saline stressed environment. The present results shown for the responses of neonates towards different salinity can further indicated the similar physiological reactions on the adult’s stages of Moina macrocopa. Thus, future researchers can use these findings for further study and research on the development of species characterization and on the factors affecting the abundance and zooplankton composition within the freshwater community, thus providing more ecologically relevant information on primary productivity in the ecosystem.
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}