
Diversity of Brazilian Troglobitic Fishes: Models of Colonization and Differentiation in Subterranean Habitats


Institute of Biosciences, University of São Paulo, São Paulo 05508-090, Brazil
†
Current address: Rua Nobre Vieira, 364, São Paulo 05587-180, Brazil.
Diversity 2021, 13(3), 106; https://0-doi-org.brum.beds.ac.uk/10.3390/d13030106
Submission received: 17 February 2021
/
Accepted: 22 February 2021
/
Published: 1 March 2021
(This article belongs to the Special Issue Cave Communities: From the Surface Border to the Deep Darkness)
Abstract
:The Brazilian subterranean ichthyofauna is distinguished worldwide by high taxonomic and phylogenetic diversity, involving at least 30 exclusively subterranean (troglobitic) lineages. This may be explained by high native epigean diversty, allied to opportunities for colonization and genetic isolation in the subterranean biotope, thence originating troglobites. An updated list of Brazilian troglobitic lineages and a review of previous hypotheses on modes of colonization and differentiation in subterranean habitats are presented with fresh data and models. Colonization of and isolation in the subterranean biotope are independent processes, usually separate over time. Access to subterranean habitats varies from the (sub)horizontal through sinkholes and resurgences of base-level streams and vadose tributaries to the vertical by shallow and deep phreatic waters through the hyporheic zone. Phenotypic differentiation of subterranean populations originating troglobites may be achieved by various, non-mutually exclusive modes of genetic isolation, e.g., directly in base-level streams and upper tributaries through the extinction of epigean populations, due to drainage discontinuation in dry paleoclimatic phases (or by other causes), by topographic isolation due to a lowering of the regional base level and karst catchments or by parapatric differentiation. Differentiation may also be a consequence of transition from lotic to lentic waters in flooded caves, and from shallow to deep phreatic habitats.
1. Introduction
Subterranean (subsurface) fishes are excellent models for studies of comparative evolution. Due to the high taxonomic, ecological and morphological diversity, the Brazilian ichthyofauna is particularly suitable for this. So far, more than 30 troglobitic fish lineages (exclusively subterranean source populations—[1]) have been recognized, including 21 valid species described as troglobitic. Mattox et al. [2] presented a list of Brazilian troglobitic and troglophilic fishes (source populations in both subsurface and surface habitats, genetically connected by mutually commuting individuals [1]). This was subsequently updated by Gallão and Bichuette [3] for the troglobites. Nonetheless, the discovery and description of novel subterranean fishes continue. Troglobites usually, but not necessarily, present troglomorphisms, i.e., autapomorphies related to isolation and specialization to the subterranean life. The most frequent are the regression of visual structures and dark cutaneous pigmentation ([1,4], among others).
Brazilian species of subterranean fishes are allocated into 12 genera, seven families and three orders, thus configuring relatively high phylogenetic diversity. In comparison, the 78 species (in 20 genera) reported as troglomorphic for China [5], which has a larger total karst area, are distributed in four families and two orders, and two genera concentrate ca. 70% if this species richness [5]. The troglobitic ichthyofauna from Mexico is less phylogenetically diverse at lower taxonomic levels, but more so at the higher levels: the 11 species recorded by Proudlove [6] are distributed in six genera and families belonging to five orders. The elevated lower and higher phylogenetic levels of Brazilian subterranean ichthyofauna, both troglobites and troglophiles, could originate from a highly diversified and rich epigean (surface) fauna providing a large array of potential colonizers of hypogean habitats, thereby giving rise to troglophilic populations. Furthermore, karst fragmentation and paleoclimatic fluctuations would favor the isolation of these populations, thence originating troglobites. The extensive coastal karst of Mexico, propitious for marine colonizer adaptation and isolation in freshwaters ideal for originating troglobites [7], among others, explains the higher level of phylogenetic diversity. Only small Brazilian karst areas to the north of Bahia State and close enough to the current coastal line are appropriate for this. Epicontinental seas dating to the Early Cretaceous are too ancient [8].
Regular studies of Brazilian subterranean ichthyofauna started in the early 1980s, initially focusing on the blind catfish Pimelodella kronei from the Iporanga, Alto Ribeira karst area ([9,10,11,12], among others), with focus on taxonomy, morphology, ecology and behavior. Representative lineages of several genera have also been studied in detail see [13,14,15,16,17,18]. Hypotheses based on paleoclimatic models (isolation by extinction of epigean relatives during unfavorable phases—[19]) have been proposed to explain the origin of P. kronei [20] and Rhamdiopsis krugi from northeastern Brazil [21]. Topographic models were proposed for P. spelaea [22] and Ituglanis spp. [23] from the São Domingos karst area, Central Brazil.
I hereby present a review of the data from these studies, prior to discussing previous hypotheses on forms of colonization and speciation in the various subterranean habitats generating troglobitic fish species in Brazil. With the aim of explaining the high diversity of subterranean ichthyofauna, former and new data were analyzed, as a means of generating novel proposals. Robust models with high predictive potential are scientifically relevant as possible indicators of areas for further research, as well as being the means of testing the generality of hypotheses. Moreover, species presenting intra-population variability, the case of most Brazilian troglobitic fishes [15], are highly propitious for testing hypotheses on the evolution (regression) of characters/features.
Models emerge from the detection of congruent attributed patterns. I compared data on geographic distribution, degree of troglomorphism (primarily eye and pigmentation regression), habitat type and body shape, including hypotheses on apomorphic states related to specialized habitats and distribution. Data are mostly from both our studies and those of collaborators (sometimes unpublished).
2. Taxonomic Considerations
Herein, I embraced the idea of species as separately evolving metapopulation lineages, which is the common element shared by the contemporary alternative species concepts [24]. As regards species delimitation (a separate issue) among the properties acquired by lineages during the course of divergence, viz., intrinsic reproductive isolation, monophyly and diagnosability, I followed de Queiroz [24], for whom diagnosability, i.e., the presence of an exclusive set of character states (not necessarily autapomorphic), widely used by taxonomists, is the most practical operational criterion when assessing lineage separation. Based on the comparative method, it facilitates recognition of subterranean-restricted species [1].
Due to variations in taxonomic approaches and the application of Schiner–Racovitza classification, comparisons of subterranean taxonomic richness between different countries must proceed with caution. Furthermore, when differences do occur in the degree of regional richness (rich vs. poor) [25], discrimination in total diversity (trogloxenes + troglophiles + troglobites), and specifically among troglobites themselves, should become apparent.
There are several instances of cryptic lineages within species described by traditional morphological taxonomy. Morphological stasis in highly specialized ancient lineages may be a consequence of life in a stable phreatic environment over a long period, viz., the Brazilian troglobitic fishes Rhamdiopsis krugi and Trichomycterus dali. On the other hand, in the case of isolation over relatively shorter periods (“recent troglobites”), with contemporary specialization to subterranean habitats, morphological differentiation may progress faster than molecular marker evolution, due to the accumulation of mutations in structural gene systems, viz., Ancistrus cryptophthalmus [26,27].
Due to the emphasis on Red Lists as a basis for defining protection policies, and the tendency to include therein only nominal species from traditional taxonomy, as diagnosed by morphological characters, the case of Brazil, thence leading to legal failure in protecting genetically distinct lineages without discernible morphological differences (cryptic diversity), the aforementioned issues pose important consequences for conservation.
3. Diversity among Brazilian Subterranean Fishes
3.1. Taxonomic Diversity (Species Richness) and Distribution
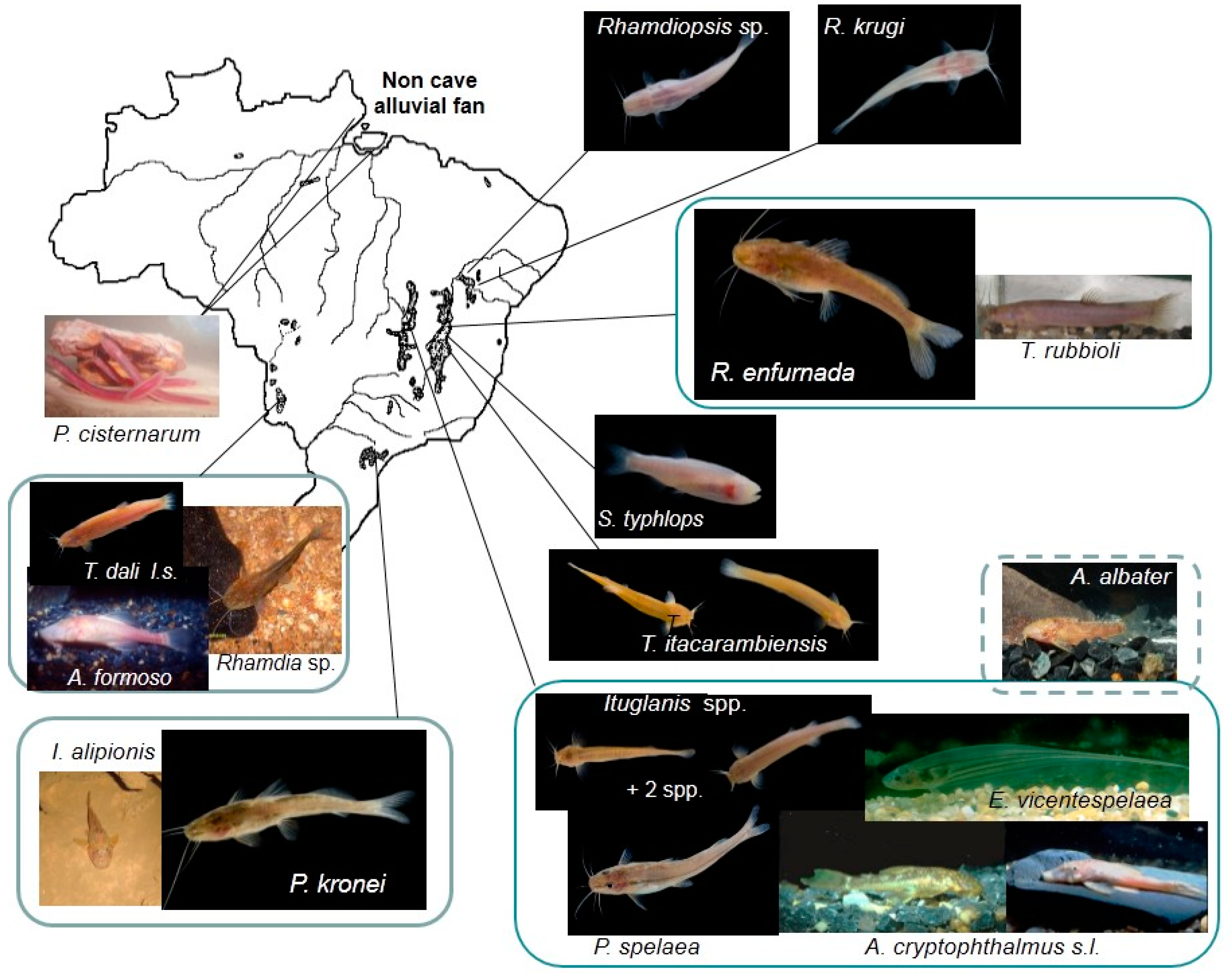
Taxonomic and geographic correlations between Brazilian putative troglobitic fishes (troglomorphic and those evidently restricted to the subterranean realm) and their respective types of habitats (according to the classification by Trajano [14]) in karst areas (Figure 1), as well as their degree of troglomorphism, are listed in Table 1. The following categories of troglomorphism (“Troglom. degree” in the Table), based mainly on the eyes and melanic pigmentation, were distinguished: (1) no detected troglomorphisms (–); (2) low degree—eyes and pigmentation only slightly, but significantly, reduced compared to epigean congeners (+); (3) moderate degree—high intra-population variation, from the epigean state to its complete absence (++); (4) high degree—eyes and pigmentation conspicuously reduced, but still noticeable in at least part of the population (+++); and (5) very high—population homogeneously anophthalmic and depigmented, frequently with other morphological, physiological and/or behavioral troglomorphisms (++++). In addition to these lineages, two troglomorphic Trichomycterus catfishes have been reported by Gallão and Bichuette [28] for Carinhanha Co., Bahia.
Among the fish genera listed in Table 1, the (so far) monotypic Glaphyropoma and Stygichthys are exclusively subterranean.
The São Domingos karst area is distinguished by high fish diversity, both at the species level (more than seven troglobitic species) and above (three genera in three families—two orders). However, when also considering the neighboring Mambaí karst area, there is a jump to 10+ species located in four genera and four families. This represents one third of the total troglobitic fish diversity known so far.
Due to their many peculiarities, the familial allocation of the Amazonian Phreatobius has become a matter of debate. During the course of its taxonomic history, it has been progressively allocated into six familes of siluriforms. Consequently, a new separate family was erected for the genus [29]. P. sanguijuela is the only Brazilian subterranean fish whose distribution encompasses another country: it was originally described based on specimens from wells in the Rio Paraguá drainage system in Bolivia. The known Brazilian and Bolivian localities are ca. 290 km apart [30].
Except for Eigenmannia vicentespelaea, an electric fish (Gymnotiformes), and Stygichthys typhlops, a characin (Characiformes) like the amply studied Mexican blind tetra characins of the genus Astyanax, all Brazilian subterranean fishes are siluriforms (catfishes). When considering described and undescribed species, as well as diverging lineages of fishes putatively isolated in subterranean habitats, taxonomic diversity, as currently known for Brazil, involves more than 30 lineages.
These lineages have been accessed through caves (mostly in carbonate rocks) and artificial openings, such as wells. Caves are generally defined as natural underground openings in rocks that are large enough for human entry [31]. This is the operational definition adopted by the Union International of Speleology (UIS), and since it is based on the size of our own species, it is clearly anthropocentric. Caves are large spaces in networks of interconnected heterogeneous spaces of the subsurface and filled with water and/or air that constitute the subterranean realm ([32], among others). Subterranean habitats include karst and non-karst caves (e.g., lava tubes), as well as smaller spaces and interstitial habitats (such as the Mesovoid Shallow Substratum- MSS and the hyporheic). Therefore, the distribution of subterranean populations may, and usually does, surpass the boundaries of “human caves” [4].
3.2. Habitat, Morphology and Adaptations to a Subterranean Life
Brazilian subterranean fishes live in habitats as diverse as the epikarst (uppermost weathered zone of carbonate rocks, capable of delaying water infiltration, as well as storage), vadose tributaries, seasonal or permanent base-level water bodies in the oscillation zone and the upper and deep phreatic zones. While some lineages show adaptation to life in still, low-oxygen waters, and others to life in fast moving waters in typical lotic environments, there are scarce correlations between taxonomic groups and the environment they dwell in [14,15]. On the other hand, there seems to be geographic correlations with the degree of troglomorphism.
About one half of these species, mostly from base-level streams, are endemic to a single cave or cave system (a karst system is a functional unit, involving organized flow pathways forming a drainage unit with input–output zones and finite configurations—[33]). Fishes occupying the phreatic zone present the highest degree of troglomorphy and the widest distribution. A few inhabit areas hundreds of kilometers in length (e.g., Trichomycterus dali sensu lato, maximum distance between localities ca. 120 km; P. sanguijuela from localities ca. 290 km apart).
For several species, there are no reported epigean relatives living in the same karst area (e.g., Pimelodella spelaea endemic to a vadose tributary inside São Bernardo Cave, Ituglanis spp. from the São Domingos karst area and Trichomycterus dali sensu lato from the Serra da Bodoquena), or in the river basin encompassing the cave system occupied by the troglobitic species (Rhamdia enfurnado endemic to Lapa do Enfurnado, Rhamdiopsis undesc. sp. from Salitre Cave and Eigenmannia vicentespelaea from São Vicente II Cave). Nevertheless, in other cases, epigean congeners are sympatric, or even syntopic, with troglomorphic populations, viz., Pimelodella kronei, Rhamdia sp. from Gruta das Fadas and Ancistrus cryptophthalmus. Epigean Eigenmannia electric fishes have been observed in the São Domingos karst area, but not in the microbasin encompassing the cave inhabited by E. vicentespelaea.
The trichomycterids Ituglanis epikarsticus, I. bambui and I. ramiroi show adaptations to life in confined spaces, viz., their small size when compared with epigean congeners, a reduced number of vertebrae (36–37 in I. bambui vs. 38 in epigean species) and a reduced laterosensory canal system. A reduction in laterosensory canals (which may be fragmented or even absent in some), a common occurrence among troglobitic fishes, is also observed in some cryptobiotic epigean fishes living in confined spaces, such as clefts and crevices, or in interstitial environments, or buried in leaf litter, rocks and sand, the case of Typhlogobius californienesis and glanapterygines [15]. The regression of laterosensory canals (but not free neuromasts, which may be hyperdeveloped) is also observed in North American troglobitic amblyopsids and ictalurids, such as Trogloglanis, Satan and Prietella, as well as in Phreatichthys from Namibia, the Chinese Sinocyclocheilus and the Brazilian Glaphyropoma, Rhamdiopsis spp. (from Bahia State), T. itacarambienses and T. dali [34]. Ituglanis passensis is medium-sized for the genus, reaching a maximum 65 mm standard length in caves (vs. 42–122 mm recorded for six described congeneric species). However, in spite of its large size, it presents a low number of vertebrae (36) [35] and a reduced lateral line in the body. This also occurs in the other subterranean Ituglanis species from São Domingos/GO.
The Phreatobiidae are remarkable for their non-karst habitats in the Amazon basin. Phreatobius cisternarum, the first subterranean troglomorphic fish described for South America, lives in alluvial fans distributed around the Amazon delta and Marajó Island, only accessible through hand-dug wells. Epigean specimens have already been encountered in wet-leaf litter in Amapá State [36]. Reichel [37] undertook an anatomical study of this species, which was by far the most detailed for any Brazilian subterranean fish. P. dracunculus, even more troglomorphic than P. cisternarum, inhabits groundwaters also only accessible through artificial wells. It occurs in Rondônia State, ca. 1900 km in a straight line from Marajó Island. Other still undescribed phreatobiids are to be found in wet-leaf litter on the margins of the Amazonas and Negro rivers [36].
Rhamdiopsis (an undescribed species from Campo Formoso, from now on referred to as Rhamdiopsis sp. CF), Ancistrus formoso, Trichomycterus sp. 2 and T. dali sensu stricto (see below—Section 3.3) present the highest degree of troglomorphism (Table 1), with adaptation to deep phreatic zones. The first three species are homogeneously anophthalmic and devoid of any trace of dark pigmentation in vivo. Furthermore, A. formoso has even lost the ability to metabolize carotenoids, such as those present in commercial food for aquarium fishes (see [38]). Specialization to life in deep phreatic zones includes the following: (1) adaptation to respiration under hypoxic conditions: in its normal habitat, the skin is dark pink, becoming gradually paler under normoxic conditions in the laboratory, hence indicating facultative cutaneous oxygen uptake; (2) density-dependent mechanisms for population control (cannibalism in S. typhlops and Rhamdiopsis sp. CF, not observed in the closely related R. krugi); and (3) large amounts of subcutaneous fatty tissue, either scattered throughout the body or located at the base of fins, such as the pre-dorsal adipose fold retained in adult Trichomycterus dali stricto sensu and Trichomycterus sp. 2 sensu Cordeiro [16].
As already mentioned, the degree of troglomorphism correlates with geographic distribution and type of habitat, but not with taxonomy. For instance, troglobitic fishes from the São Domingos karst area, Central Brazil, are moderately troglomorphic, whereas those from the semiarid central-north Bahia State, Northeast Brazil, are highly troglomorphic (Table 1). Likewise, the most troglomorphic species are phreatobic, and the less troglomorphic are stream dwellers. This indicates the influence of common geographic and ecological factors, such as paleoclimate, geomorphology and hydrogeology, on evolution in isolation in subterranean habitats.
3.3. Hidden Diversity
Data on Brazilian subterranean fishes point to landmark morphometrics, a set of techniques with the highest discrimination potential based on shape [26], as a useful tool for uncovering hidden diversity. As an example, studies of Trichomycterus dali, Rhamdiopsis krugi and Ancistrus cryptophthalmus resulted in splitting these nominal species into two or three lineages [16,21,26].
When geometric morphometric techniques were applied to widely distributed Brazilian troglobitic fishes (T. dali, R. krugi), the differences found among sets of populations pointed to separate clades, usually associated with alternative microbasins or aquifer vicariant events. Except for A. cryptophthalmus, molecular data were consistent with the differences revealed by landmark morphometrics. The importance of multiple and complementary approaches to improve the robustness of species discovery hypotheses and associated descriptions was highlighted by Zimmermamm et al. [39], among others.
Geometric morphometric comparison between two populations of Rhamdiopsis krugi (“Una” and “Irecê”), from (currently) separate sedimentary microbasins, revealed differences in body shape corresponding to the respective site/location, hence indicating divergence in the isolation (or semi-isolation) of the two morphotypes. These results are congruent with molecular data [21].
Extending from north to south, congruent data from geometric morphometric analysis and molecular studies revealed three divergent lineages within the nominal species Trichomycterus dali, viz., Trichomycterus sp. 1, T. dali stricto sensu and Trichomycterus sp. 2 [16]. Trichomycterus dali stricto sensu, known from five sites, including its type locality, the Saracura Cave on the south-left margin of the Rio Salobra, is distributed throughout an area of ca. 400 km2. It differs morphologically from the other two lineages by the unique cranial fontanel extending from the posterior half of the supraoccipital to the posterior region of the two frontal bones, with a conspicuous constriction at the meeting point of the supraoccipital and the two frontal bones. Trichomycterus sp. 2 (sensu Cordeiro [16]), located at a single site on the Rio Miranda basin, South Bodoquena Plateau, is characterized by an elongated fontanel, a larger number of opercular odontoids and the total absence of melanic pigmentation. Trichomycterus sp. 1 (sensu Cordeiro [16]), encountered at five sites across 500 km2 on the north-left margin of the Rio Salobra basin, North Bodoquena Plateau, is the most divergent and differs from both T. dali sensu Cordeiro and Trichomycterus sp. 2, by the reduction in the cranial fontanel to the parieto-supraoccipital bone, the more posterior position of the dorsal fin and the formation of a cilinder-shaped body in the adults. The latter is due to regression of the pre-dorsal adipose fold in adults, as observed in epigean Trichomycteus spp. This fold is retained in the original form in the other two subterranean lineages, probably as a neotenic adaptation to life in the deep phreatic zone. Trichomycteus sp. 1 inhabits a vadose tributary in the Urubu Rei Cave that opens through a resurgence ca. 150 m above the regional base level (probably a nursery), with source populations in the shallow phreatic zone connected to the surface through caves, resurgences and sinkholes. The highly troglomorphic Trichomycterus dali sensu stricto and Trichomycterus sp. 2 occupy shallow to deep phreatic zones accessible through springs and caves [16].
The nominal species Ancistrus cryptophthalmus comprises four partially differentiated populations, viz., one at Passa Três, the type locality, another distributed between two caves of the São Vicente system and one each in the Angélica and Bezerra Caves of the Angélica–Bezerra system, Goiás State, whence the mosaic of morphological and behavioral characteristics originates [26,40,41]. The Passa Três catfish is the most distinct, without any overlap with the other populations as regards body shape [26]. Ancistrus armored catfish represent the only case among troglobitic fishes from Goiás State for which epigean congeners are currently present in the same microbasin, with parapatric distribution in Angélica Cave.
According to molecular studies of the hypervariable Region I of MtDNA, there was no apparent divergence between the cave populations of Ancistrus cryptophthalmus [27]. Mendes and colleagues [42] used Illumina sequencing technology to produce partial genome sequences of A. cryptophthalmus from the Passa Três and Angélica Caves and epigean Ancistrus specimens taken near the Angélica entrance. Disregarding variability in eye development, dark pigmentation and divergence in body shape in progress among the cave populations [27], as well as the clear separation of the Passa Três population, not only as to body shape and eye development (these fishes are born with well-developed eyes, which degenerate with growth and become externally invisible in adults [43]), but also the presence of exclusive haplotypes (H7 and H8—[42]), they proposed conspecific status for A. cryptophalmus and epigean armored catfish from the Angélica microbasin. Similarities between the epigean and Angélica catfish, including shared haplotypes (H1, H4 and H5), may be due to recent divergence in the cave population and/or some hybridization due to secondary contact with surface specimens. It is relevant that the studied epigean specimens were collected near the cave entrance, which is a sinkhole. Strong floods during the rainy season may carry fish into the cave, whence introgression of epigean genes into the resident cave population occurs. Since lack of proof is not evidence of absence, so far, molecular data have provided no relevant contribution to elucidate the question.
Pimelodella kronei has been found at six sites in four distinct systems in the Betari microbasin, São Paulo State, viz., the Areias de Cima and Areias de Baixo Caves (separated by a karst window, where blind catfish have occasionally been observed at night) and Ressurgência da Areias da Água Quente (=Águas Quentes) (Areias system), Ressurgência das Bombas (Bombas system), Córrego Seco Cave (Córrego Fundo system) and the Alambari de Cima and Gurutuva Caves (Alambari system). These systems were defined according to hydrogeological studies using tracing tests [44]. The Alambari de Cima population probably became extinct in the 1970s due to a pollution event caused by mining activities [34]. The Bombas population is only accessible through individuals foraging at the resurgence during the night.
On comparing specimens of the sympatric epigean Pimelodella transitoria with those of P. kronei from Areias de Cima and Bombas, Pavan [45] detected differences in morphology as to head shape, number of serrae on the pectoral spine, length of barbels and dark pigmentation. Except for pigmentation, the highest variation occurred in Areias, and the lowest occurred in Bombas (the most divergent). This was confirmed in studies of gross brain anatomy [11] and behavior [46]. Molecular studies focusing on COI and CytB haplotypes are consistent with hypotheses on at least two divergent lineages within P. kronei, one from Bombas and the other from the remaining populations (V. Slobodian, G.P.S. Figueiredo and J. Muriel-Cunha, pers. comm. 2016).
4. Origin and Evolution of Subterranean Fishes in Brazil
4.1. The Challenge of Colonizing Subterranean Habitats: The Establishment of Troglophilic Populations
In general, Brazilian troglobitic fishes belong to taxa comprised of typical stream dwellers in the epigean realm. Hence, it is reasonable to suppose that the colonizers of subterranean habitats were already adapted to the lotic environment, and that this state was retained by several troglobitic species as plesiomorphic traits (see heptapterid catfishes [47]).
Subterranean colonization occurs in two distinct surroundings: (1) in large spaces that impose no limitations on movement for large-sized individuals; and (2) in small limited spaces involving adaptation through body and locomotor changes, such as miniaturization, elongation and enhanced maneuverability. The first consists of almost horizontal epigean–hypogean colonization from large to large spaces through sinkholes and resurgences, as well as colonization through wide shafts, as proposed for the Mexican blind fish Astyanax [48]. The second would be karst habitat colonization through overlying hyporheic zones, such as interstitial spaces between sediments of the streambed, which could be considered transition zones between the surface and groundwater [33]. Vertical colonization of the subterranean realm, eventually followed by lateral dispersion throughout the aquifer, was proposed by Bichuette and colleagues [21] for Rhamdiopsis krugi.
With few exceptions, colonization leads to the establishment of troglophilic populations (species with source populations from both epigean and subterranean habitats, genetically connected by intercommuting individuals—[1]). If submitted to speciation processes, usually due to vicariant events, genetically isolated troglophilic populations may differentiate into troglobitic species. These processes are widely separated in time. Exceptions include the simultaneous colonization and isolation that occur in stream captures [48].
It is noteworthy that the climatic model is an allopatric model for the origin of troglobites due to isolation from epigean populations, and not for colonization [49].
4.2. Modes and Models of Isolation and Differentiation in Subterranean Habitats: Origin of Troglobites
The main allopatric models proposed to explain the origin of troglobites are: (1) the paleoclimatic model—isolation of epigean populations during unfavorable climatic phases/events, with survival in the hypogean realm, whereby species are protected from extreme climatic fluctuations [19]; and (2) the appearance of geomorphological (topographic) or hydrogeological barriers, due to karst evolution.
The paleoclimatic model was indicated for identifying the origin of Pimelodella kronei, by differentiation among subterranean populations isolated during dry glacial phases [20]. Secondary contact with epigean descendants of a common ancestor resulted in the present-day situation: six P. kronei populations sympatric with the eyed and pigmented P. transitoria comprising troglophilic populations in various caves. Based on differences observed in the external morphology between specimens of P. kronei from populations in Areias de Cima and Bombas, Pavan [45] debated a possible independent origin through the same P. transitoria-like ancestor. Further studies revealed differences in brain anatomy and behavior between the two, consistent with a possibly independent origin [11,46], viz., more attenuated agonistic behavior among Bombas individuals, while those of Areias presented enlarged brains and variable responses to light (vs. consistent photophobic reactions shown in epigean P. transitoria), hence configuring a mosaic distribution of traits. Correlations between the Areias and Bombas populations, either as sister taxa or the result of parallel evolution, are still an open question.
In the Bombas population, the high homogeneity and enhanced divergence are indications of founder effects and a small population size. Past hydrological connections facilitating dispersion of cave-adapted Pimelodella (a proto-P. kronei) may have been disrupted by landscape changes brought about by geological and depositional events. The larger size of the Areias population mirrors the variability in and proximity to P. transitoria. The small number of specimens collected in the Córrego Seco and Gurutuva Caves impedes interpopulation comparison.
Counterwise, the São Domingos karst area, located in the Cerrado core area (Brazilian biome with savannah-like vegetation), is divest of accentuated paleoclimatic fluctuations such as those observed in other karst areas, such as in Northeast Brazil and Alto Ribeira [50]. This could explain the relative paucity of local invertebrate troglobites. On the other hand, the diversity of troglobitic fishes could arise from other geomorphological and hydrological factors, such as rapid alluvial down-cutting and the presence of a well-developed epikarst [23].
As stated, the nominal species Ancistrus cryptophthalmus from the São Domingos karst area comprises four differentiated cave populations from two systems. Streams of the Angélica, Bezerra and São Vicente 2 system are base level and contiguous with epigean reaches through open sinkholes, where epigean Ancistrus catfish form large populations, without any physical barriers separating them from cave dwellers. On the other hand, in the Passa Três system, the stream, a vadose upper tributary of São Vicente 2 Cave, is isolated by topographic barriers, whereby morphological differentiation is consistent with the allopatric model [26]. For base-level stream populations, although parapatric differentiation is a possibility, allopatric differentiation of cave populations followed by secondary dispersion of epigean catfish cannot be ruled out.
Pimelodella spelaea dwells in a vadose stream comprising an upper tributary of the large São Bernardo Cave stream and constantly fed by infiltrated water from the epikarst. This conduit opens onto the surface through the sinkhole of a temporary epigean stream (S.Secutti pers. comm.). Eyed Pimelodella probably colonized the São Bernardo system when the base level was higher and the epigean tributary was permanent. With a lowering of the base level, due to either alluvial down-cutting or karst catchment, the stream became temporary, thereby leading to the extinction of aquatic communities. The only fish that successfully colonized and survived in isolation in the cave, probably due to preadaptation (in the sense of exaptation—[51]) to the subterranean life (nocturnal activity, cryptobiotic habits, predominant chemorientation, as in most siluriforms), were P. spelaea ancestors.
A similar case is that of the cave population of the armored catfish Isbrueckerichthys alipionis from Santana Cave, in the Alto Ribeira karst area. These fishes have been regularly observed for more than three decades (C.F. Lino pers. comm. 1980s; S.Secutti pers. comm. 2015) in an upper vadose stream in the Takeupa gallery, the Ronco stream, which flows into the base-level Roncador stream below. Nowadays, the Takeupa gallery is ca. 70 m below the Furnas streambed. There are several sinks from the centripetal intermittent drainage of polygonal karst basins just over Takeupa [52]. Thus, it is probable that the Rio Ronco is now fed by water infiltrated from the Furnas basin and intermittently by autogenic water from polygonal karst basins. As in the case of P. spelaea, these catfish may have colonized the subterranean environment from a past permanent epigean drainage that was part of what is now the Furnas microbasin. With a lowering of the regional base level and karst catchments, there was a disruption: drainage is now represented by polygonal basins fed intermittently by rainwater. Santana Cave has been intensively surveyed, and no I. alipionis catfish were ever seen in Rio Roncador. The strong current generated by gradient differences between the upper Rio Ronco and the base-level Rio Roncador hampers fish colonization, hence the topographic isolation. Since isolation in a subterranean realm per se, not necessarily together with troglomorphisms, defines the troglobite status [1], the armored catfish population living in Rio Ronco could be considered troglobitic.
4.3. Stream Dwellers: (Sub)Horizontal Colonization through Sinkholes and Resurgences, Eventually Followed by Isolation
Most Brazilian stream-dwelling troglobitic fishes show moderate to low degrees of troglomorphisms, regarding the development of eyes and melanic pigmentation. The plesiomorphic body shape and size indicate primary evolution in lotic habitats, where no specialization to phreatic groundwater would be required. Hence, stream-dwelling troglobites would retain the primitive body shape and so-called “normal” size (average size of the putative ancestor, inferred by examination of the probably closest epigean relative). This is the case with the two Pimelodella and the two Rhamdia species, Rhamdiopsis sp. from Salitre Cave, Ancistrus cryptophthalmus and Ancistrus sp. from the Gruta das Fadas, Trichomycterus itacarambiensis, Ituglanis mambai and I. boticario, Glaphyropoma spinosum and the syntopic Copionodon sp., the Aspidoras albater cave form and Eigenmannia vicentespelaea. My hypothesis is that (sub)horizontal colonization events occurred through cave “entrances”, such as sinkholes and resurgences (spaces much larger than the fish body, thus facilitating maneuverability), as against vertical colonization through the hyporheic zone (see below). These fishes include species with the lowest degree of troglomorphism (eyes and dark pigmentation visible, although reduced when compared to epigean relatives), viz., Pimelodella spelaea, Rhamdiopsis from Salitre Cave, I. mambai and I. boticario, Copionodon sp., A. albater and E. vicentespelaea.
Caves, as defined above, are transient features within karst cycles. According to the commonest speleogenetic process, which originates most karst caves, the initiation phasis starts at the appearance of small fissures in the rock (protoconduits) at the top of the phreatic zone. These progressively become enlarged through dissolution until reaching a critical size (5–15 mm diameter). In the following enlargement phasis, a network of spaces becomes progressively available for colonization by larger and even larger organisms, until reaching the cave size in the human-centered definition (around 1 m). During this phasis, when the water flow changes from laminar to turbulent, with erosion also contributing to conduit enlargement, fast-flowing waters become available for colonization by preadapted populations living in lotic epigean habitats. Eventually, in the cessation phasis, large subterranean spaces disappear due to breakdowns or filling by chemical or clastic sediments [31,53]. This transient characteristic in karst cycles explains why stream-dwelling troglobitic fishes in general are less troglomorphic than the phreatobic ones. Moreover, in the cessation phasis, stream dwellers that have managed to adapt to the lentic conditions of the phreatic zone below may survive over a long geological period until the next cycle.
4.4. Transition from Lotic to Lentic Waters: Adaptation to Phreatic Waters
4.4.1. Stygichthys Typhlops
White piabas, Stygichthys typhlops, are typical troglobites of the phreatic zone in the Jaíba aquifer, with adaptations to deep oxygen- and food-poor waters [54]. Although they are currently accessible only through man-made wells, according to local inhabitants, these fishes were also found at the bottom of a certain small natural cave, which was subsequently filled with sediment to avoid vampire bats. Doubts as to their correct position within the large characine subfamily circumscribe applying the comparative method to further studies.
Unlike the slender, flexible siluriforms, allocation and accommodation to small confined spaces in interstitial habitats would require extreme changes in the characiform body plan. The maximum body size and shape of S. typhlops are within the range normally observed in the group, with no clear evidence of miniaturization. Hence, there is no clear evidence of karst vertical colonization through hyporheic zones. Ancestors of this fish were probably stream dwellers that colonized caves and non-cave karst spaces through connections with the surface, such as resurgences, insurgences, sinkholes and shafts. It is possible that this population had been isolated by climatic changes that disrupted epigean drainage, leading to alterations from lotic to lentic conditions. Adaptation to deep waters includes physiological and behavioral specialization, such as density-dependent cannibalism, highly vascularized skin and the loss of circadian locomotor rhythms [54].
4.4.2. Flooded Caves: Troglobitic Fishes in Serra da Bodoquena Karst Area
The Bodoquena Carbonatic Plateau stands above the plains of the Pantanal wetlands. The Bodoquena karst area is undergoing subsidence, related to the development of the Pantanal. Apart from climatic variations recorded for the Quaternary (variations in the water table level indicate a drier regional period of ca. 5000 years BP, followed by a wetter climate), tectonic subsidence would be a major contributor to the relative uprising of the water table due to a lowering of the regional base level [55]. There is a difference of almost 1000 m between the higher northern Plateau and the Rio Miranda Depression in the south. Consequently, due to an increase in the hydraulic gradient, Bodoquena subsidence progresses from south to north; hence, the relative uprising of the water table flooding caves and other subterranean habitats in the southern plains predates the observed flooding in the northern plateau.
Hydrological and topographic barriers may account for variations in subterranean populations of Trichomycterus. Trichomycterus sp. 1 sensu Cordeiro [16], while occupying the upper phreatic zone, with some individuals reproducing in a vadose cave, has retained the body shape typical of epigean Trichomycterus that live in lotic habitats, as well as the interopercular odontodes [family sinapomorphy], an adaptation for anchoring in fast-running waters, and vestiges of melanic pigmentation. Trichomycterus dali sensu stricto and Trichomycterus sp. 2 sensu Cordeiro, although showing adaptations to the deep phreatic zone, preferably reproduce in the less food-limited upper zone [16]. There is no evidence of miniaturization. Studied specimens of T. dali and Trichomycterus sp. 1 and sp. 2 reached lengths of 83.3, 100.8 and 98.4 mm, respectively (SL) [16,34], which are within the normal range for medium-sized epigean Trichomyterus catfishes.
In spite of intense efforts, no Trichomycterus catfishes have been encountered in the epigean streams of Serra da Bodoquena. The extinct local populations could have preceded isolation in subterranean habitats. With the lack of evidence of adaptation to small spaces, sub-horizontal colonization of cave streams through large connections with the surface (caves, sinkholes, resurgences, shafts) is supposed. Adaptation to lentic environments as subterranean spaces became flooded due to progressive subsidence of the Bodoquena Plateau from south to north explains the higher degree of troglomorphy in the southern lineage (i.e., Trichomycterus sp. 2). Moreover, low genetic and morphological divergence between the latter and T. dali may be an outcome of the simultaneous adaptation of both to deep phreatic conditions.
The same model—direct (sub)horizontal colonization of karst spaces through connections with the surface by stream-dwelling ancestors, and secondary adaptation to phreatic groundwaters due to cave flooding—applies to the subterranean armored catfishes Ancistrus spp. from Bodoquena. As could be expected, and based on the above-described scenario, the highly troglomorphic Ancistrus formoso lives in a flooded cave system in the Formoso microbasin (south plateau) [38], whereas the less troglomorphic and undescribed Ancistrus sp. is from Fadas Cave on the northern plateau [16].
4.4.3. Trichomycterus Rubbioli: The Problem of Sink Populations
Trichomycterus rubbioli was studied in two separate caves of the Água Clara system, Bahia State, in an upper vadose stream ca. 180 m long, fed by water infiltration inside Lapa dos Peixes, and from a side pool in Gruna da Água Clara. It shares several troglomorphisms with T. dali sensu lato, but at a less advanced stage of specialization (comparisons are based on Trichomycterus sp. 2 and T. dali sensu Cordeiro), viz., very reduced eyes, externally visible only as spots (vs. eyes that are absent); depigmented when alive, but still capable of light-induced melanogenesis (vs. color-unresponsive to light); barbels longer than in epigean congeners, especially the nasal one; pre-dorsal adipose fold fully developed in juveniles, but poorly so in adults (vs. developed in adults); circadian locomotor activity rhythms reduced (but not so much as in Trichomycterus sp. 2); frequents mid-waters and the surface. This indicates life in slow-moving waters in relatively large subterranean spaces. On the other hand, the occurrence in discontinuous caves points to dispersion throughout non-cave spaces.
After the first collections in Lapa dos Peixes (12 specimens caught), the number of observed fishes declined sharply for several months, thus indicating this to be a sink population, i.e., that which, if cut off from migrants, will eventually become extinct, contrary to source poulations that have excess production and so continue to grow if isolated (D. Culver—[56]). So far, the Trichomycterus rubbioli source population has not been identified, a factor compromising the formation of a hypothesis on colonization and isolation in the subterranean realm.
Sink populations pose problems for subterranean biology, due to the difficulties in distinguishing the selective regime under which the species population evolved within the chosen habitat. This can lead to ambiguous conclusions on ecology and evolution. The same applies to individuals found in peripheral habitats, such as illuminated cave sectors and artificial wells, the case with Stygichthys typhlops [57]. Furthermore, epigean sink populations obscure recognition of the troglobitic status (exclusively subterranean source populations) of troglomophic organisms.
4.5. Multiple-Step Model of Colonization and Differentiation in Subterranean Habitats: The Way through the Hyporheic Zone and Epikarst
4.5.1. Rhamdiopsis spp. from Semiarid Bahia
Besides presenting adaptations to date, morphology also supplies information about certain past evolutionary steps (phylogenetic signals), which presume evolutionary constraints. Mosaics of incongruent character states provide evidence of miscellaneous adaptive regimes over time [21]. In order to explain the coexistence of adaptations to both small, confined spaces and large ones of the troglobitic heptapterid Rhamdiopsis krugi from northeastern Brazil, Bichuette and colleagues [21] proposed for this species a two-step model of vertical colonization of subterranean habitats through the epikarst, followed by adaptation to the larger spaces underneath (Table 2).
Step 1: Colonization of the hyporheic zone by typically stream-dwelling Rhamdiopsis-like catfishes was achieved through miniaturization (probably by paedomorphosis), as well as by possessing slender, elongated and flexible bodies with a reduced lateral line on the trunk, and other adaptations for energy economy in a food-poor environment, when compared to the surface, for instance, the distribution of fatty cells throughout the body. Reduction in body size may also be an adaptation for the more efficient use of energy in extreme environments [58]. Survival and establishment of self-sustained populations in the hyporheic zone were possible due to: (1) preadaptations of epigean ancestors (nocturnal activity, cryptobiotic habits, generalist carnivory disposition and elongated bodies, typical of most siluriforms); and (2) the presence of potential prey appropriate for small catfish, in the form of a diversified community of microcrustaceans, such as copepods, ostracods and amphipods, together with micro-oligochaetes, nematoids and other minute organisms (e.g., [59,60]).
Step 2: Colonization through the hyporheic zone to the underlying subterranean habitat implied acquiring the ability to navigate in large and heterogeneous spaces. This requires adaptations that contrast with those achieved for the hyporheic zone. These comprise enhancement of sound sensitivity (highly exposed pseudotympanum), as well as a broadening of the head and snout to reduce pitching and rolling during swimming in mid-water, and which present a wider surface for free neuromasts, taste buds and other sensory structures. Isolation in subterranean habitats can occur during the dry phases of paleoclimatic cycles (such as that of the present day in the region), when epigean drainage is disrupted. Based on speleothem dating, the alternation of dry and wet periods in central Bahia over the last 210,000 years has been well documented [61]. Thus, due to the local extinction of epigean relatives, the paleoclimatic model could explain R. krugi speciation by allopatry [21].
In order to explain differences between the populations from two distinct microbasins (Una and Irecê), Bichuette and colleagues [21] proposed geographic isolation (a least partial) by geological (insoluble rocks between the two limestone outcrops), depositional (colmated connecting ducts) and hydrological (running waters) barriers. Since the monophyly of R. krugi is well supported by 24 autapomorphies [62], it is reasonable to assume a past connection through a continuous aquifer in the limestone outcrop, which was subsequently disrupted by karst denudation.
Taxonomic studies based on an extensive and detailed examination of the morphology and anatomy of Rhamdiopsis sp. CF point to a sister group relationship between this catfish and R. krugi. When compared to epigean Rhamdiopsis, these troglobitic species share at least 14 sinapomorphies, of which at least five can be considered troglomorphisms. These consist of the absence of eyes and an atrophied optic foramen, the very short laterosensory canal in the trunk, regression of circadian rhythms and cryptobiotic habits and extended spatial activity involving the mid-water and surface. They also have in common small-sized adults (wild-caught specimens less than 46 mm SL), a short trunk with few segments, usually 38–40 vertebrae, and a widely exposed pseudotympanum, at least in juveniles, as well as several osteological characters, such as anterior and posterior branches of the transverse process of vertebra 4 with broad bases, an undivided or weakly separated anterior portion of the spatulate-shaped posterior branch of the transverse process of vertebra 4, a parhypural fused to the complex plate formed by hypurals 1 and 2 and hypural 5 co-ossified with the complex plate formed by hypurals 3 and 4 [F.A.Bockmann and E.Trajano, unpubl. data].
Adaptations of R. krugi to phreatic waters are also present in the even more troglomorphic Rhamdiopsis sp. CF. The latter probably represents an even further step in phreatic zone colonization, viz., adaptation to a deep phreatic habitat characterized by hypoxia and energy deprivation conditions, as well as high hydrostatic pressure, representing an additional step, Step 3, in the previous model (Table 2). At this stage, the head and trunk have become almost entirely filled by fatty subcutaneous corpuscles, cannibalism may occur probably as a density-dependent mechanism for population control and the skin becomes dark pink, when in the natural habitat, thereby presuming facultative cutaneous oxygen uptake.
The putative sister group relationship of R. krugi and Rhamdiopsis sp. CF implies a subterranean connection between areas. Rhamdiopsis sp. CF data are based on specimens found in the upper zone connected with the surface through Toca do Gonçalo, which developed in the Caatinga limestones, where karstification probably begun only ca. 2.5 mi years ago. On the contrary, the contiguous Una carbonates and the Salitre Formation, present not only in Campo Formoso, but also in Chapada Diamantina, were exposed at the beginning of the Paleocene, ca. 65 mi years ago [63]. The high degree of troglomorphism and relictual status indicate an ancient origin not only for Rhamdiopsis sp. CF, but also for syntopic troglobitic invertebrates, such as the calabozoan Pongycarcinia xiphidiouros and amphipod Spelaeogammarus trajanoae (widespread in Campo Formoso). Their ancestors probably evolved in the Una karst and subsequently dispersed to the vicinity of the Caatinga limestones, becoming available for biological colonization through karstification. Hence, Toca do Gonçalo would be a peripheral habitat.
Aquifer connectivity throughout a large area in north-central Bahia is corroborated by the presence of amphipods of the exclusively subterranean genus Spelaeogammarus, with six nominal species distributed over 1200 km on the N–S axis. These species are found in several karst areas (defined by surface features), including Campo Formoso and Chapada Diamantina [63].
Dispersal throughout the deep phreatic zone of the Una limestones between Chapada Diamantina and Campo Formoso by a common ancestor would explain the adaptations of Rhamdia sp. CF to life in deep groundwater, whereas R. krugi remained at least partially associated with the upper phreatic zone. Although capable of descending to as deep as 45 m, the highest population densities of the latter are observed near the surface. According to this model, the local presence of troglobitic Rhamdiopsis could be expected in small discontinuous limestone outcrops between Chapada Diamantina and Campo Formoso.
4.5.2. Ituglanis Catfishes from the São Domingos Karst Area
The four troglomorphic Ituglanis species found in the São Domingos karst area inhabit caves in distinct microbasins, viz. (from north to south), I. bambui in Angélica Cave, I. passensis in the Passa Três, I. epikarstikus in the São Matheus and I. ramiroi in the São Bernardo (see Table 1). The mosaic distribution of morphological traits and separation of the cave systems indicate the independent origin of the four species from a common epigean ancestor, nowadays extinct in the region [23].
As mentioned, these catfishes show special adaptations to confined spaces, such as miniaturization (except for I. passensis), a reduced number of vertebrae and a shorter lateral line on the trunk. Ituglanis ramiroi, I. bambui and I. epikarsticus occur in habitats associated with the epikarst: the first two in vadose tributaries fed by epikarst waters; the latter in a permanent rimstone pool. The first specimens of I. ramiroi were collected in small side pools by the main river channel. The small number of specimens and the high population turnover, with very low recapture rates [64], indicate that these are sink populations, with source populations living in the epikarst. Additional evidence is provided by the occurrence of I. bambui at the resurgence end of Angélica Cave, ca. 6 km in a straight line from the vadose stream where they were first studied. Dispersion throughout the epikarst is the most parsimonious explanation for such a disjunct distribution inside the cave.
Colonization of the epikarst probably occurred throughout the hyporheic zone of the epigean drainage system formerly crossing the area, when the regional base level was above the present-day caves. With the opening of the caves and rapid alluvial down-cutting, the regional water table dropped to its present-day position, while suspended aquifers became isolated in the epikarst of parallel microbasins [26] (Figure 1). Surviving populations would then differentiate, thereby giving rise to troglobitic Ituglanis species. Hence, these species can be placed in Step 1 of our model of vertical colonization and differentiation in subterranean karst habitats (Table 2).
In contrast, Ituglanis passensis, medium-sized for the genus, lives in the open channel of an upper tributary, a typical lotic habitat. This is an allochthonous stream, carrying large amounts of organic matter through the sinkhole during the rainy season. Nonetheless, these catfish also present traits associated with Step 1 in the other species from São Domingos (36 vertebrae, reduced lateral line on the trunk). Hence, I propose that I. passensis originated from a miniaturized ancestor living in a vadose tributary, as occurred with I. bambui and I. ramiroi. When the ceiling collapsed following erosion, the large opening to the surface became the sinkhole of a small epigean stream, thereby facilitating the input of larger amounts of organic matter. Intraspecific competition in the absence of food limitation could account for the selection of larger sizes, thus leading to a reversal of miniaturization. This would be a third step in a different model (Table 2).
4.5.3. The Amazon Basin: Phreatobius spp.
The uniqueness of Phreatobiidae (sensu Cunha [29]) indicates ancient adaptation to a very specialized habitat (wet-leaf litter at stream margins, interstitial subterranean spaces). The subterranean Phreatobius populations known so far, those of P. cisternarum, on the one hand, and P. dracunculus and P. sanguijuela, on the other, occur at nearly opposite sides of the extensive Brazilian Amazon, at least 1900 km apart [36], thus indicating a widely distributed ancestor. The possible isolation of Phreatobius populations at the periphery of the huge Amazon basin was due to a lowering of the water table, as a result of drainage cut-down, associated with the Andean uplift and cycles of global sea level drops, as recorded in the Middle Miocene (~12 Ma) [65]. The vertical colonization of subterranean habitats through streambeds, followed by speciation in non-karst interstitial habitats, is a reasonable scenario (Table 2).
It is noteworthy that the Phreatobius dracunculus and P. sanguijuela sites in Rondônia State are separated by Serra dos Pacaás Novos and Chapada dos Parecis, important water dividers in the region [30]. Dispersion through a hyporheic zone with continuous drainage is a possible explanation for the wide distribution of the highly troglomorphic P. sanguijuela in Brazil and Bolivia at distances of up to 290 km.
5. Conclusions
The high taxonomc and phylogenetic diversity of Brazilian troglobitic (exclusively subterranean) fishes (30+ lineages in seven families and three orders, mostly Siluriformes) could be attributed to high epigean diversty, allied to opportunities for colonization and genetic isolation in the subterranean biotope.
Colonization of and isolation in the subterranean biotope are two independent processes, usually separated in time. Colonization with the establishment of self-sustained hypogean populations originates troglophiles (source populations in both epigean and subterranean habitats, genetically connected by mutually commuting individuals).
Pathways of fish access to subterranean habitats vary from (sub)horizontal colonization through sinkholes and resurgences of base-level streams (typical lotic habitats) and through vadose tributaries to vertical colonization of shallow and deep phreatic waters (lentic habitats) through the hyporheic zone.
Phenotypic (morphological, physiological, behavioral) differentiation in subterranean populations originating troglobites is achieved by various, unmutually exclusive modes of genetic isolation:
Directly in base-level streams and upper tributaries through the extinction of epigean populations brought about by drainage discontinuation during dry paleoclimatic phases (e.g., Pimelodella kronei, Rhamdia enfurnada, Trichomycterus itacarambiensis) or by other causes (Pimelodella undesc sp. from Fadas Cave).
By topographic isolation due to a lowering of the regional base level and karst catchments (Ancistrus cryptophthlmus from Passa Três Cave, Isbrueckerichthys alipionis).
By parapatric differentiation (possibly the case of Ancistrus cryptophthalmus from the Angélica system).
Adaptation to the deep pheatic zone may be a consequence of active dispersion (Rhamdiopsis sp. CF) or cave flooding due to the relative uprising of the water table (Trichomycterus dali sensu stricto and Trichomycterus sp. 2 sensu Cordeiro).
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
I am greatly indebted to Marcelo Carvalho, for the critical reading of the manuscript, to Ramon A. Clark, for the revision of the English style, to William Sallun Filho, for geological counseling, and to Maria Elina Bichuette, for fruitful discussions and exchange of information along all these years.
Conflicts of Interest
The author declares no conflict of interest.
References
- Trajano, E.; Carvalho, M.R. Towards a biologically meaningful classification of subterranean organisms: A critical analysis of the Schiner-Racovitza system from a historical perspective, difficulties of its application and implications for conservation. Subterr. Biol. 2017, 22, 1–26. [Google Scholar] [CrossRef]
- Mattox, G.M.T.; Bichuette, M.E.; Secutti, S.; Trajano, E. Surface and subterranean ichthyofauna in the Serra do Ramalho karst area, northeastern Brazil, with updated lists of Brazilian troglobitic and troglophilic fishes. Biota Neotrop. 2008, 8, 145–152. [Google Scholar] [CrossRef] [Green Version]
- Gallão, J.E.; Bichuette, M.E. Brazilian obligatory subterranean fauna and threats to the hypogean environment. Zookeys 2018, 746, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Giachino, P.M.; Vailati, D. The Subterranean Environment; WBA Handbooks: Verona, Italy, 2010; Volume 3, pp. 1–132. [Google Scholar]
- Ma, L.; Zhao, Y.; Yang, J. Cavefish of China. In Encyclopedia of Caves, 2nd ed.; White, W., Culver, D.C., Pipan, T., Eds.; Academic Press: Waltham, MA, USA, 2019; pp. 237–254. [Google Scholar]
- Proudlove, G.S. Biodiversity and Distribution of the Subterranean Fishes of the World. In Biology of Subterranean Fishes; Trajano, E., Bichuette, M.E., Kapoor, B.G., Eds.; Science Publ.: Enfield, UK, 2010; Chapter 2; pp. 41–63. [Google Scholar]
- Wilkens, H. Regressive evolution and phylogenetic age: The history of colonization of freshwaters of Yucatan by fish and Crustacea. Assoc. Mex. Cave Stud. Bull. 1982, 8, 237–243. [Google Scholar]
- Arai, M. Aptian/Albian (Early Cretaceous) paleogeography of the South Atlantic: A paleontological perspective. Braz. J. Geol. 2014, 44, 339–350. [Google Scholar] [CrossRef]
- Trajano, E. Populational ecology of Pimelodella kronei, troglobitic catfish from southeastern Brazil (Siluriformes, Pimelodidae). Environ. Biol. Fishes 1991, 30, 407–421. [Google Scholar] [CrossRef]
- Trajano, E. Agonistic behaviour of Pimelodella kronei, a troglobitic catfish from Southeastern Brazil (Siluriformes, Pimelodidae). Behav. Process. 1991, 23, 113–124. [Google Scholar] [CrossRef]
- Trajano, E. Comparative study on the brain and olfactory organ of the troglobitic catfish, Pimelodella kronei (Ribeiro, 1907), and its putative ancestor, P. transitoria (Ribeiro, 1912) (Siluriformes, Pimelodidae). Trop. Zool. 1994, 7, 145–160. [Google Scholar] [CrossRef]
- Trajano, E.; Ueno, J.C.H.; Menna-Barreto, L. Evolution of time-control mechanisms in subterranean organisms: Cave fishes under light-dark cycles (Teleostei: Siluriformes, Characiformes). Biol. Rhythm. Res. 2012, 43, 191–203. [Google Scholar] [CrossRef]
- Trajano, E. Ecology and ethology of subterranean fishes. In Catfishes; Arratia, G., Kapoor, B.C., Chardon, M., Diogo, R., Eds.; Science Publ.: Enfield, UK, 2003; Volume 2, pp. 601–635. [Google Scholar]
- Trajano, E. Ecology of subterranean fishes: An overview. Environ. Biol. Fishes 2001, 62, 133–160. [Google Scholar] [CrossRef]
- Trajano, E.; Bichuette, M.E. Subterranean Fishes of Brazil. In Biology of Subterranean Fishes; Trajano, E., Bichuette, M.E., Kapoor, B.G., Eds.; Science Publ.: Enfield, UK, 2010; Chapter 9; pp. 331–355. [Google Scholar]
- Cordeiro, L.M. Distribuição, Ecologia e Filogeografia dos Bagres Troglóbios do Gênero Trichomycterus na Área Cástica Da Serra da Bodoquena, MS. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2014. [Google Scholar]
- Bichuette, M.E.; Trajano, E. Population density and habitat of an endangered cave ifsh Eigenmannia vicentespelaea Triques, 1996 (Ostariophysi: Gymnotiformes) from a karst area in central Brazil. Neotrop. Ichthyol. 2015, 13, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Bichuette, M.E.; Trajano, E. Biology and behavior of Eigenmannia vicentespelaea, a troglobitic electric fish from Brazil (Teleostei: Gymnotiformes: Sternopygidae): A comparison to the epigean species, E. trilineata, and the consequences of cave life. Trop. Zool. 2017, 30, 68–82. [Google Scholar] [CrossRef]
- Barr, T.C. Cave ecology and the evolution of troglobites. Evol. Biol. 1968, 2, 35–102. [Google Scholar]
- Trajano, E.; Britski, H.A. Pimelodella kronei (Ribeiro,1907) e seu sinônimo Caecorhamdella brasiliensis Borodin, 1927: Morfologia externa, taxonomia e evolução (Teleostomi, Siluriformes). Bol. Zool. 1992, 12, 53–89. [Google Scholar] [CrossRef]
- Bichuette, M.E.; Rantin, B.; Zingst-Zaher, E.; Trajano, E. Geometric morphometrics throws light on evolutionof the subterranean catfish Rhamdiopsis krugi (Teleostei: Siluriformes: Heptapteridae) in eastern Brazil. Biol. J. Linn. Soc. 2015, 114, 136–151. [Google Scholar] [CrossRef] [Green Version]
- Trajano, E.; Reis, R.E.; Bichuette, M.E. Pimelodella spelaea, a new cave catfish from Central Brazil, with data on ecology and evolutionary considerations (Siluriformes: Heptapteridae). Copeia 2004, 315–325. [Google Scholar] [CrossRef]
- Bichuette, M.E.; Trajano, E. Three new subterranean species of Ituglanis from Central Brazil (Siluriformes: Trichomycteridae). Ichthyol. Explor. Freshw. 2004, 15, 243–256. [Google Scholar]
- de Queiroz, K. Species concepts and species delimitation. Syst. Biol. 2007, 56, 879–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trajano, E. The challenge of estimating the age of subterranean lineages: Examples from Brazil. Acta Carsol. 2007, 36, 191–198. [Google Scholar] [CrossRef] [Green Version]
- Reis, R.E.; Trajano, E.; Hingst-Zaher, E. Shape variation in surface and cave populations of the armoured catfish Ancistrus (Siluriformes: Loricariidae) from the São Domingos karst area, Upper Tocantins River, Brazil. J. Fish Biol. 2006, 68, 414–429. [Google Scholar] [CrossRef]
- Moller, D.; Parzefall, J. Single or multiple origin of the subterranean catfish, Ancistrus cryptophthalmus. What we can learn from molecular data. Brazil ISB 2001, 1, 38. [Google Scholar]
- Gallão, J.E.; Bichuette, M.E. A Lista de Fauna Ameaçada de Extinção e os entraves para a inclusão de espécies-o exemplo dos peixes troglóbios brasileiros. Nat. Conserv. 2012, 10, 83–87. [Google Scholar] [CrossRef]
- Cunha, J.M.F.L. Biodiversidade e Sistemática Molecular de Phreatobiidae (Ostariophysi, Siluriformes)—Com Uma Proposta Sobre sua Posição Filogenética em Siluriformes e uma Discussão Sobre a Evolução do Hábito Subterrâneo. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2008. [Google Scholar]
- Ohara, W.M.; Costa, I.D.; Fonseca, M.L. Behaviour, feeding habits and ecology of the blind catfsh Phreatobius sanguijuela (Ostariophysi: Siluriformes). J. Fish Biol. 2016, 89, 1285–1301. [Google Scholar] [CrossRef] [PubMed]
- Ford, D.; Williams, P. Karst Hydrology and Geomorphology; John Wiley & Sons: Chichester, UK, 2007; 562p. [Google Scholar]
- Howarth, F.G. Ecoloy of cave arthropods. Ann. Rev. Entomol. 1983, 28, 365–389. [Google Scholar] [CrossRef]
- Gibert, J.; Stanford, J.A.; Dole-Olivier, M.-J.; Ward, J.V. Basic attributes of groundwater ecosystems and prospects for research. In Groundwater Ecology; Danielopol, D.L., Stanford, J.A., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 7–40. [Google Scholar]
- Rizzato, P.P.; Costa, E.P., Jr.; Trajano, E.; Bichuette, M.E. Trichomycterus dali: A new highly troglomorphic catfish (Silurifomes: Trichomycteridae), from Serra da Bodoquena, Mato Grosso do Sul State, Central Brazil. Neotrop. Ichthyol. 2011, 9, 477–491. [Google Scholar] [CrossRef]
- Fernández, L.; Bichuette, M.E. A new cave dwelling species of Ituglanis from heSão Domingos karst, Cenral Brazil (Siluriformes: Trichomycteridae). Ichhyol. Explor. Feshw. 2002, 13, 273–278. [Google Scholar]
- Shibatta, O.K.; Muriel-Cunha, J.; de Pinna, M.C.C. A new subterranean species of Phreatobius Goeldi, 1905 (Siluriformes, incertae sedis) from the Southwestern Amazon basin. Pap. Avulsos Zool. 2007, 47, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Reichel, M. Étude anatomique de Phreatobius cisternarum Goeldi, Silure aveugle do Brésil. Rev. Suisse Zool. 1927, 34, 28–401+6 plates. [Google Scholar]
- Sabino, J.; Trajano, E. A new species of blind armoured catfish, genus Ancistrus, from caves of Bodoquena region, Mato Grosso do Sul, southwestern Brazil (Siluriformes, Loricariidae, Ancistrinae). Rev. Fr. Aquariol. 1998, 24, 73–77. [Google Scholar]
- Zimmermann, B.L.; Campos-Filho, I.S.; Araujo, P.B. Integrative taxonomy reveals a new genus and new species of Philosciidae (Crustacea: Isopoda: Oniscidea) from the Neotropical region. Can. J. Zool. 2018, 96, 473–485. [Google Scholar] [CrossRef]
- Trajano, E.; Souza, A.M. The behaviour of Ancistrus cryptophthalmus, an armoured blind catfish from caves of Central Brazil, with notes on syntopic Trichomycterus sp. (Siluriformes: Loricariidae, Trichomycteridae). Mém. Biospéol. 1994, 21, 151–159. [Google Scholar]
- Bessa, E.; Trajano, E. Light reation and cryptobiotic habits in armoured catfishes, genus Ancistrus, from caves in central and northeastern Brazil (Siluriformes: Loricariidae). Mém. Biospéol. 2002, 28, 29–37. [Google Scholar]
- Mendes, I.S.; Prosdocimi, F.; Schomaker-Bastos, A.; Furtado, C.; Ferreira, R.L.; Pompeu, P.S.; Carvalho, D.C. On the evolutionary origin of Neotropical cavefish Ancistrus cryptophthalmus (Siluriformes, Loricariidae) based on the mitogenome and genetic structure of cave and surface populations. Hydrobiologia 2019, 842, 157–171. [Google Scholar] [CrossRef]
- Secutti, S.; Trajano, E. Reproductive behavior, development and eye regression in the cave armored catfish, Ancistrus cryptophthalmus Reis, 1987 (Siluriformes: Loricariidae), breed in laboratory. Neotrop. Ichthyol. 2009, 7, 479–490. [Google Scholar] [CrossRef] [Green Version]
- Genthner, C.; Ferrari, J.A.; Karmann, I. Identificação das áreas de recarga de fontes cársticas com o uso do traçador Rodamina FWT (Área Carbonática Lajeado—Bombas, Iporanga-SP). Rev. Inst. Geol. 2003, 24, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Pavan, C. Os peixes cegos das cavernas de Iporanga e a evolução. Bol. Fac. Fil. Ciênc. Let. 1945, 79, 9–104. [Google Scholar]
- Hoenen, S. Comparative field ecology and morphology of two populations of the troglobitic catfish Pimelodellla kronei (Siluriformes: Pimelodidae) from southeastern Brazil. Pap. Avulsos Zool. 1998, 40, 323–335. [Google Scholar]
- Trajano, E.; Bockmann, F.A. Evolution of ecology and behaviour in Brazilian cave Heptapterinae catfishes, based on cladistic analysis (Teleostei: Siluriformes). Mém. Biospéol. 1999, 26, 123–129. [Google Scholar]
- Mitchell, R.W.; Russell, W.H.; Elliott, W.R. Mexican eyeless characin fishes, genus Astyanax: Environment, distribution, and evolution. Spec. Publ. Mus. Tex. Tech. Univ. 1977, 12, 1–89. [Google Scholar]
- Trajano, E. Ecological classification of subterranean organisms. In Encyclopedia of Caves; White, W.B., Culver, D.C., Eds.; Academic Press: Waltham, MA, USA, 2012. [Google Scholar]
- Trajano, E. Evolution of tropical troglobites: Applicability of the model of Quaternary climatic fluctuations. Mém. Biospéol. 1995, 22, 203–209. [Google Scholar]
- Buss, D.M.; Haselton, M.G.; Shackelford, T.K.; Bleske, A.L.; Wakefield, J.C. Adaptations, Exaptations, and Spandrels. Am. Psychol. 1998, 53, 533–548. [Google Scholar] [CrossRef]
- Karmann, I. Evolução e Dinâmica Atual do Sistema Cárstico do Alto Vale do Rio Ribeira de Iguape, Sudeste do Estado de São Paulo. Ph.D. Thesis, Universidade de São Paulo, São Paulo, CA, USA, 1994. [Google Scholar]
- Bosák, P. Karst processes from the beginning to the end: How can they be dated¿. In Evolution of Kars: From Prekarst to Cessation; Gabrovšek, F., Ed.; Inštitut za raziskovanje krasa, ZRC SAZU, Zalošba: Postojna-Ljubljana, Slovenija, 2002. [Google Scholar]
- Moreira, C.R.; Bichuette, M.E.; Oyakawa, O.T.; Pinna, M.C.C.; Trajano, E. Rediscovery and redescription of the unusual subterranean characiform Stygichthys typhlops, with notes on its life history. J. Fish Biol. 2010, 76, 1815–1824. [Google Scholar] [CrossRef] [PubMed]
- Sallun-Filho, W.; Karmann, I.; Wang, X.; Edwards, R.L.; Cheng, H.; Asmeromd, Y.; Polyak, V.J. Neotectônica vs. clima na submersão de cavernas no carste da Serra da Bodoquena (MS). In XLIII Congresso Brasileiro de Geologia; Anais, ST06:AO-462: Aracaju, Brazil, 2006; p. 122. [Google Scholar]
- Fong, D.W. Intermittent pools at headwaters of subterranean drainage basins as sampling sites for epikarst fauna. In Epikarst; Jones, W.K., Culver, D.C., Herman, J.S., Eds.; Karst Waters Institute Special Publication: Chales Town, WV, USA, 2004; Volume 9, pp. 114–188. [Google Scholar]
- Trajano, E.; Moreira, C.S. Stygichthys typhlops Brittan & Böhlke, 1965 (Teleostei:Characiformes), a phreatobic fish from eastern Brazil: Comments on Sampaio et al. (2012). Speleobiol. Notes 2014, 6, 33–41. [Google Scholar]
- Passow, C.N.; Greenway, R.; Arias-Rodriguez, L.; Jeyasingh, P.D.; Tobler, M. Reduction of Energetic Demands through modification of body size and routine metabolic rates in extremophile fish. Physiol. Biochem. Zool. 2015, 88, 371–383. [Google Scholar] [CrossRef] [Green Version]
- Pacioglu, O. Ecology of the hyporheic zone: A review. Cave Karst Sci. 2010, 36, 69–76. [Google Scholar]
- Moldovan, O.T.; Levei, E. Temporal variability of fauna and the importance of sampling frequency in the hyporheic zone. Hydrobiologia 2015, 755. [Google Scholar] [CrossRef]
- Wang, X.; Auler, A.S.; Edwards, R.L.; Cheng, H.; Cristalli, P.S.; Smart, P.L.; Richard, D.A.; Shen, C.-C. Wet periods in northeastern Brazil over the past 210 kyr linked to distant climate anomalies. Nature 2004, 432, 740–743. [Google Scholar] [CrossRef]
- Bockmann, F.A.; Castro, R.M.C. The blind catfish from the caves of Chapada Diamantina, Bahia, Brazil (Siluriformes: Heptapteridae): Description, anatomy, phylogenetic relationships, natural history, and biogeography. Neotrop. Ichthyol. 2010, 8, 673–706. [Google Scholar] [CrossRef] [Green Version]
- Araújo, A.V.; Leal, L.R.B.; Gomes, D.F. Anfípode subterrâneo do gênero Spelaeogammarus como um indicador de conectividade em um aquífero cárstico da bacia do rio Salitre, centro norte do Estado da Bahia. In Proceedings of the Anais do XIX Congresso Brasileiro de Aguas Subterrâneas, Campinas, Brazil, 20–23 September 2016; Associação Brasileira de Águas Subterrâneas: São Paulo, Brazil, 2016. [Google Scholar]
- Bichuette, M.E. Distribuição, Biologia, Ecologia Populacional e Comportamento de Peixes Subterrâneos Gêneros Ituglanis (Siluriformes: Trichomycteridae) e Eigenmannia (Gymnotiformes: Sternopygidae) da Área Cárstica de São Domingos Nordeste de Goiás. Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brazil, 2003. [Google Scholar]
- Hoorn, C.; Wesselingh, F.P.; ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartín, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia Through Time: Andean Uplift, Climate Change, Landscape Evolution, and Biodiversity. Science 2010, 330, 927–931. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Distribution of selected troglobitic fishes in Brazilian karst areas. Photos: D. Fenolio (Rhamdiopsis sp., Rhamdiopsis krugi, Rhamdia enfurnada, Stygichthys typhlops, Ituglanis spp., Trichomycterus itacarambiensis, T. dali sensu lato, Pimelodella kronei), J. Muriel Cunha (Phreatobius cisternarum), Alexandre Camargo (Aspidoras albater cave form), I. Sazima (Ancistrus cryptophthalmus, left), E. Trajano (Ancistrus cryptophthalmus, right) J. Sabino (Ancistrus formoso, Eigenmannia vicentespelaea), L.M. Cordeiro (Rhamdia sp. Serra da Bodoquena), S. Secutti (Isbrueckerichthys alipionis).
Figure 1.
Distribution of selected troglobitic fishes in Brazilian karst areas. Photos: D. Fenolio (Rhamdiopsis sp., Rhamdiopsis krugi, Rhamdia enfurnada, Stygichthys typhlops, Ituglanis spp., Trichomycterus itacarambiensis, T. dali sensu lato, Pimelodella kronei), J. Muriel Cunha (Phreatobius cisternarum), Alexandre Camargo (Aspidoras albater cave form), I. Sazima (Ancistrus cryptophthalmus, left), E. Trajano (Ancistrus cryptophthalmus, right) J. Sabino (Ancistrus formoso, Eigenmannia vicentespelaea), L.M. Cordeiro (Rhamdia sp. Serra da Bodoquena), S. Secutti (Isbrueckerichthys alipionis).


{kind=link}
Table 1.
Brazilian troglobitic fishes (valid species with authors; undescribed species and cryptic lineages with references), indicating the kast areas where they occur (Figure 1) (SP = State of São Paulo; GO = Goiás, BA = Bahia; MS = Mato Grosso do Sul; MG = Minas Gerais), habitat type and degree of troglomorphism (See text for “Troglom. Degree”). Undesc. sp. = undescribed species. Gray lines: endemic to a single cave or cave system. * with adaptations to the deep phreatic zone. Grey lines highlight lneages that are highly endemic.
Table 1.
Brazilian troglobitic fishes (valid species with authors; undescribed species and cryptic lineages with references), indicating the kast areas where they occur (Figure 1) (SP = State of São Paulo; GO = Goiás, BA = Bahia; MS = Mato Grosso do Sul; MG = Minas Gerais), habitat type and degree of troglomorphism (See text for “Troglom. Degree”). Undesc. sp. = undescribed species. Gray lines: endemic to a single cave or cave system. * with adaptations to the deep phreatic zone. Grey lines highlight lneages that are highly endemic.
| Species | Distribution—Karst Area | Habitat | Troglom. Degree |
|---|---|---|---|
| Order Siluriformes | |||
| HEPTAPTERIDAE | |||
| Pimelodella kronei Ribeiro 1907 [at least two divergent lineages—Pavan 1945, among others] | Alto Ribeira/SP | Lotic—Base-level streams | ++ |
| P. spelaea Trajano, Reis and Bichuette 2004 | São Domingos/GO | Lotic—Vadose tributary | + |
| Rhamdia enfurnada Bichuette and Trajano 2005 | Serra do Ramalho/BA | Lotic—Base-level stream | ++ |
| Rhamdia undesc. sp. (Gruta das Fadas) [Borghezan 2013] | Serra da Bodoquena/MS | Lotic—Base-level stream | ++ |
| Rhamdiopsis krugi Bockmann and Castro 2010 [two lineages—Bichuette et al. 2015)] | Chapada Diamantina/BA | Lentic—Upper phreatic zone | +++ |
| Rhamdiopsis undesc. sp. (Toca do Gonçalo) [Trajano and Bichuette 2010] | Campo Formoso/BA | Lentic—Upper phreatic zone * | ++++ |
| Rhamdiopsis undesc. sp. (Gruta do Salitre) [Trajano and Bichuette 2010] | Cordisburgo/MG | Base-level stream | + |
| TRICHOMYCTERIDAE | |||
| Trichomycterus dali Rizzato, Costa-Jr., Trajano and Bichuette 2011 [three lineages—Cordeiro 2014] | S. Bodoquena/MS | Upper and deep phreatic zones; vadose tributary | +++/++++ |
| T. rubbioli Bichuette and Rizzato 2012 | Serra do Ramalho/BA | Vadose tributary; phreatic | +++ |
| T. itacarambiensis Trajano and Pinna 1996 | Peruaçu/MG | Lotic—Base-level streams | ++ |
| Ituglanis bambui Bichuette and Trajano 2004 | São Domingos/GO | Vadose tributary | ++ |
| I. epikarsticus Bichuette and Trajano 2004 | Epikarst | ++ | |
| I. ramiroi Bichuette and Trajano 2004 | Vadose tributary/epikarst | ++ | |
| I. passensis Fernández and Bichuette 2002 | Base-level stream | ++ | |
| I. mambai Bichuette and Trajano 2008 | Mambaí/GO | Base-level streams | + |
| I. boticario Bichuette and Rizzato 2012 | + | ||
| Glaphyropoma spinosum Bichuette, Pinna and Trajano 2008 | Chapada Diamantina/BA—sandstones | Base-level streams | ++ |
| Copionodon sp. Gruna dos Torras [Bichuette et al. 2008] | + | ||
| LORICARIIDAE | |||
| Ancistrus formoso Sabino and Trajano 1997 | Serra da Bodoquena/MS | Flooded cave system | ++++ |
| Ancistrus sp. G. Fadas [Borghezan 2013] | Base-level stream | ++ | |
| A. cryptophthalmus Reis 1987 [2+ differentiated lineages—Reis et al 2006] | São Domingos/GO | Lotic—Base-level streams | ++ |
| Isbrueckerichthys alipionis from Santana Cave | Alto Ribeira/SP | Upper vadose tributary | - |
| CALLYCHTHYIDAE | |||
| Aspidoras albater cave form [Secutti et al. 2011] | Mambaí/GO | Base-level stream | + |
| PHREATOBIIDAE (sensu Cunha 2008) | |||
| Phreatobius cisternarum Goeldi 1904 | Amazonas River delta and Marajó Is. | Non-karst phreatic waters (accessible through artificial wells) | ++ |
| P. dracunculus Shibata, Muriel-Cunha and Pinna 2007 | Rio Branco drainage/RO | +++ | |
| P. sanguijuela Fernandéz, Saucedo, Carvajal-Vallejos and Schaeffer 2007 | Rio Guaporé drainage/RO (also in Bolivia) | ||
| Order Gymnotiformes | |||
| STERNOPYGIDAE | |||
| Eigenmannia vicentespelaea Triques 1996 | São Domingos/GO | Base-level stream | + |
| Order Characiformes | |||
| CHARACIDAE | |||
| Stygichthys typhlops Brittan and Böhlke 1965 | Jaíba/MG | Phreatic | ++++ |
Table 2.
Vertical colonization of subterranean habitats by stream dwellers: schematic sequence of evolutionary steps originating present-day Brazilian troglobitic fishes.
Table 2.
Vertical colonization of subterranean habitats by stream dwellers: schematic sequence of evolutionary steps originating present-day Brazilian troglobitic fishes.
| Species | Step 1 | Step 2 | Step 3 | |
|---|---|---|---|---|
| Rhamdiopsis krugi | Colonization of the hyporheic zone | Colonization of the subterranean habitat beneath the hyporheic zone | Genetic isolation | ------- |
| Rhamdiopsis sp. CF | Transition from upper to deep phreatic zone | |||
| Ituglanis bambui, I. epikarsticus, I. ramaroi | Colonization of epikarsts | Sink populations in vadose streams and the main river channel | ||
| I. passensis | Secondary adaptation to a lotic habitat due to the opening of large entrances | |||
| Phreatobius spp. | Colonization of sedimentary layers beneath the hyporheic zone | -------- |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Trajano, E. Diversity of Brazilian Troglobitic Fishes: Models of Colonization and Differentiation in Subterranean Habitats. Diversity 2021, 13, 106. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030106
AMA Style
Trajano E. Diversity of Brazilian Troglobitic Fishes: Models of Colonization and Differentiation in Subterranean Habitats. Diversity. 2021; 13(3):106. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030106
Chicago/Turabian StyleTrajano, Eleonora. 2021. "Diversity of Brazilian Troglobitic Fishes: Models of Colonization and Differentiation in Subterranean Habitats" Diversity 13, no. 3: 106. https://0-doi-org.brum.beds.ac.uk/10.3390/d13030106
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.