
Diversity and Distribution of Mid- to Late-Stage Phyllosomata of Spiny and Slipper Lobsters (Decapoda: Achelata) in the Mexican Caribbean

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
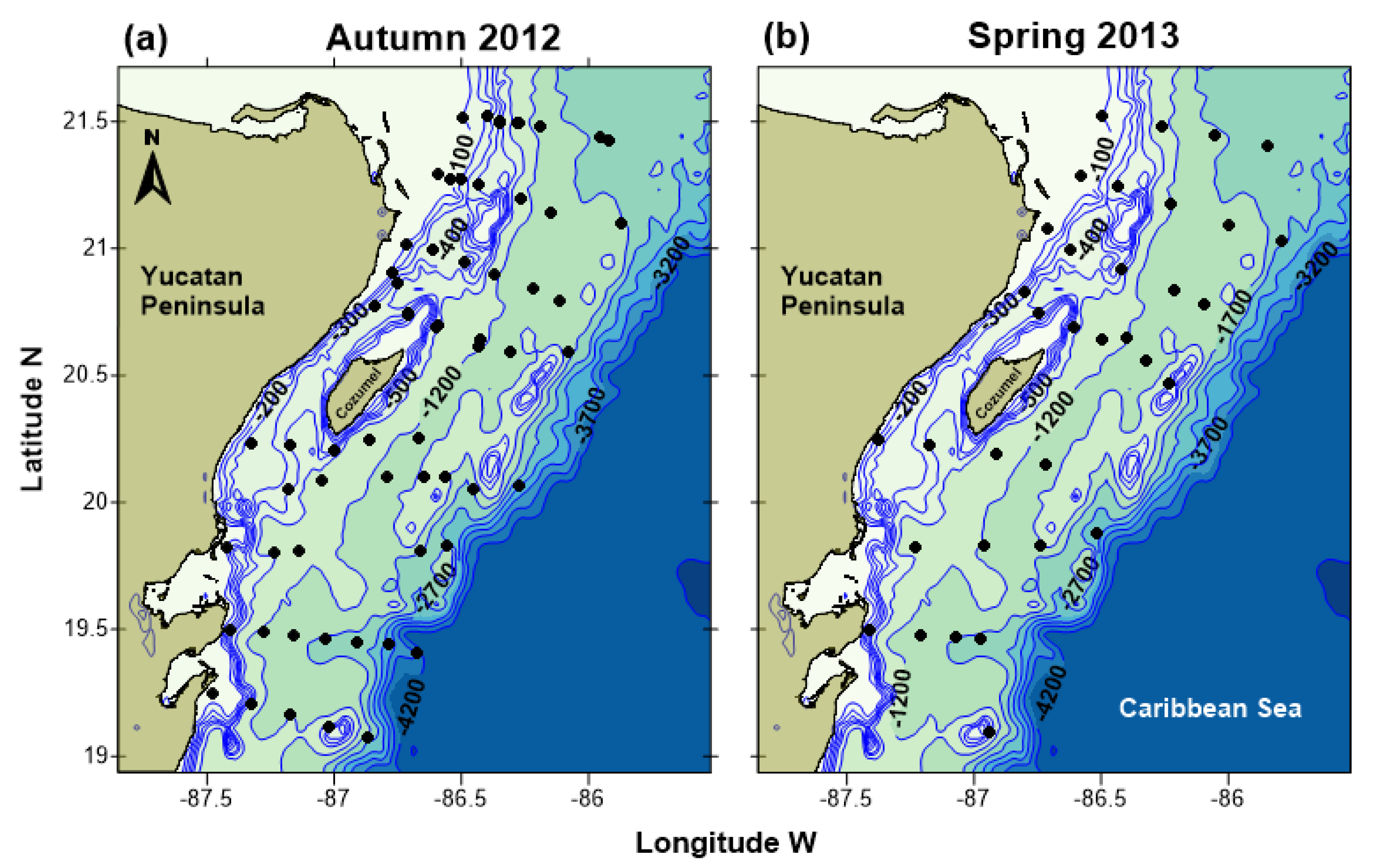
2.1. Study Area
2.2. Larval Sampling and Identification
2.3. Data Analyses
3. Results
3.1. Surface Geostrophic Current Fields
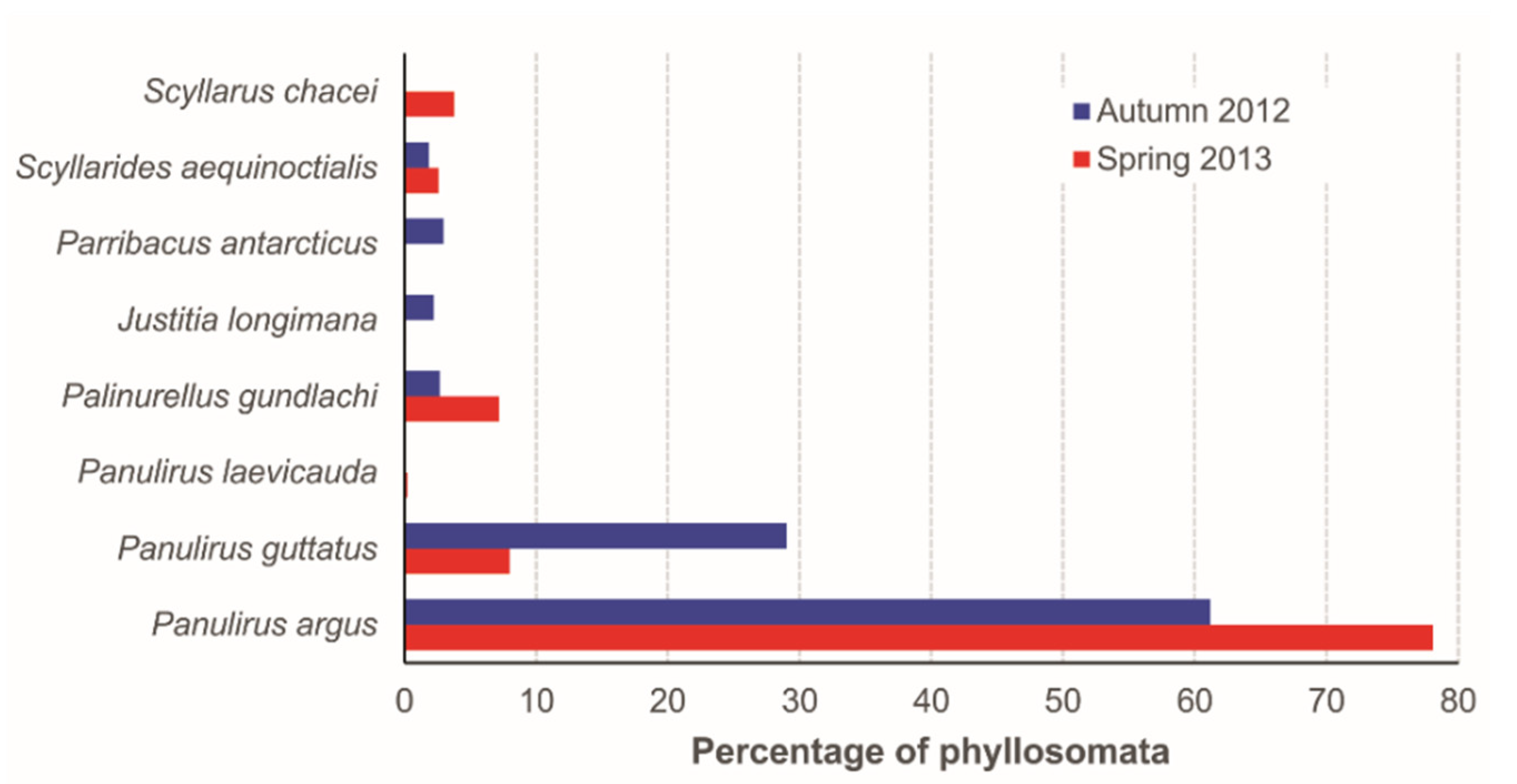
3.2. Species Abundance and Stage Composition
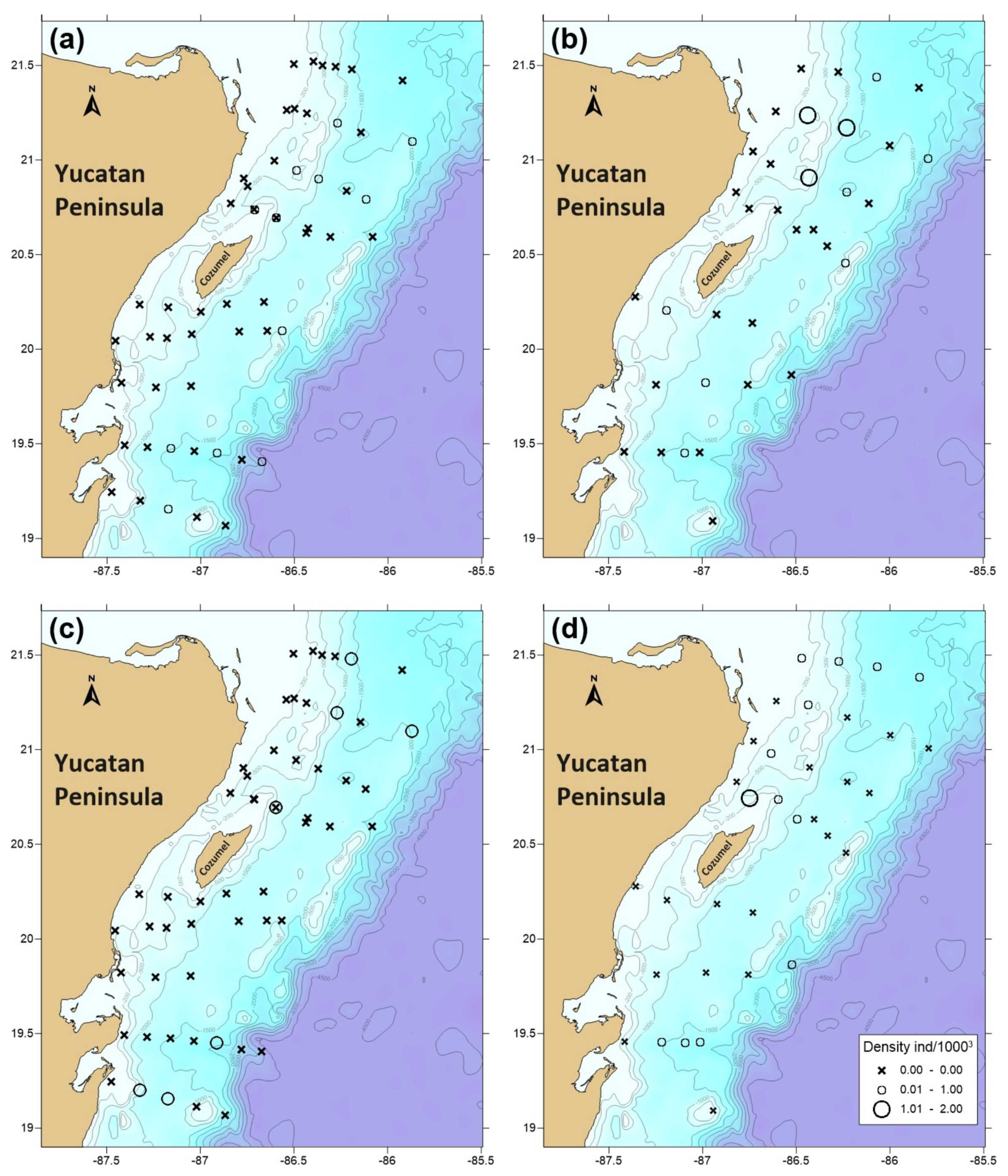
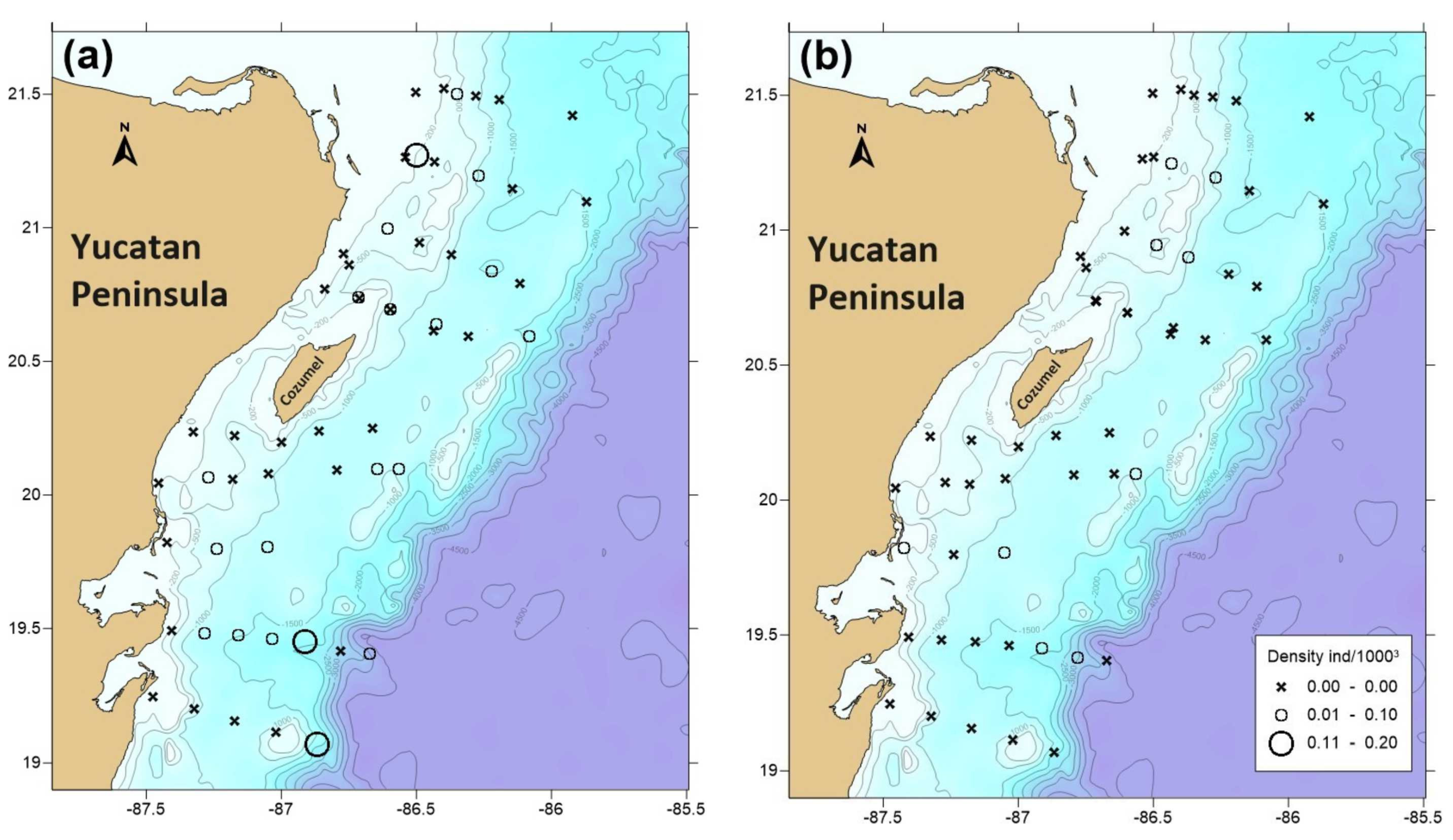
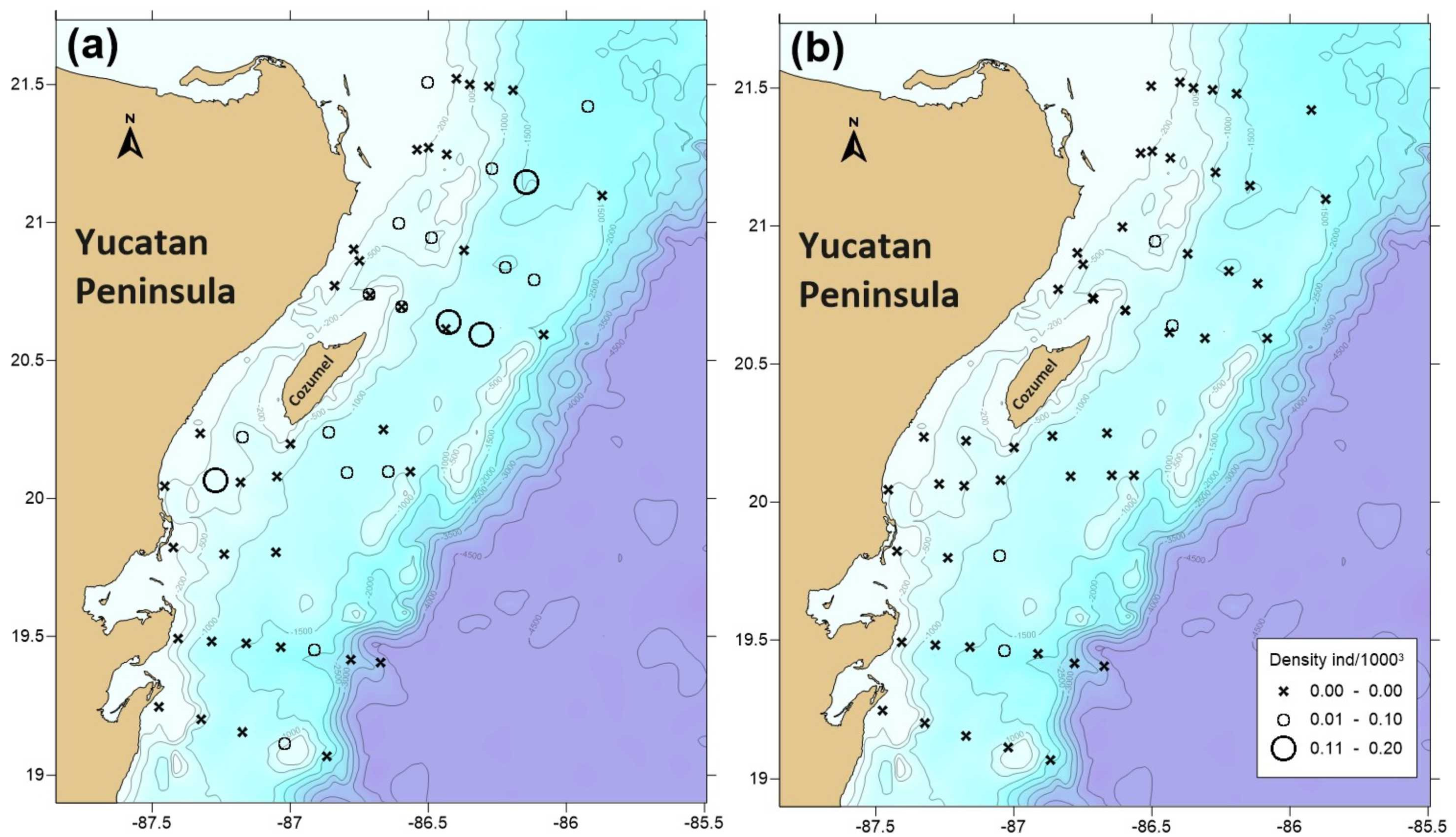
3.3. Phyllosomata Density and Spatial Distribution
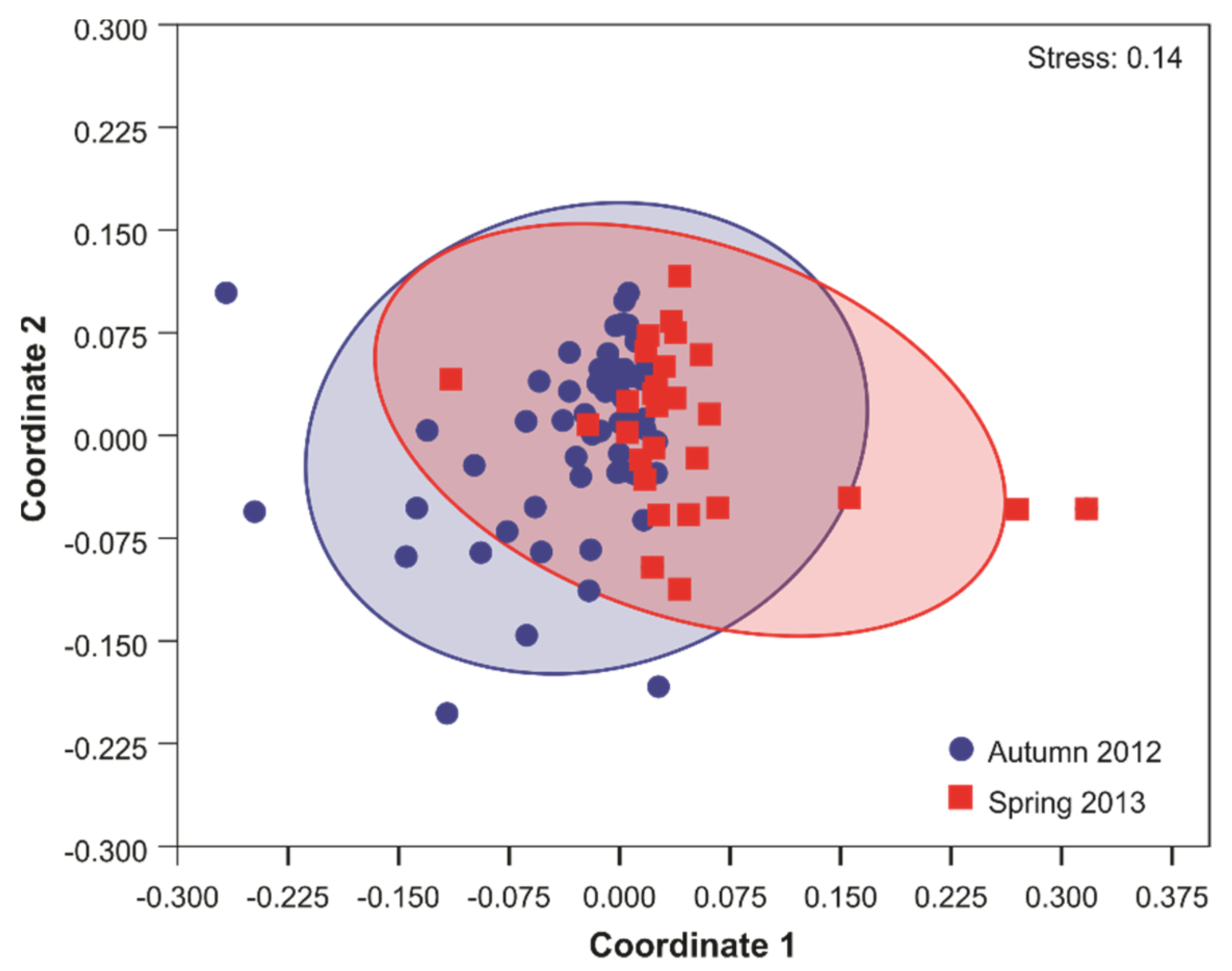
3.4. Phyllosomata Biodiversity and Assemblage Composition
4. Discussion
4.1. Palinurids
4.2. Scyllarids
4.3. Phyllosomata Assemblages and Oceanographic Features
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Autumn Cruise: Average Similarity: 45.94% | ||||||
| Species | Av.Abund | Av.Sim | Sim/SD | Contrib% | Cum.% | |
| Panulirus argus mid-stages | 3.0 | 16.6 | 1.4 | 36.1 | 36.1 | |
| Panulirus guttatus mid-stages | 2.1 | 11.3 | 1.2 | 24.6 | 60.7 | |
| Panulirus argus late stages | 1.8 | 10.0 | 1.3 | 21.7 | 82.5 | |
| P. guttatus late stages | 1.0 | 4.3 | 0.7 | 9.3 | 91.8 | |
| Spring Cruise: Average Similarity: 44.36% | ||||||
| Species | Av.Abund | Av.Sim | Sim/SD | Contrib% | Cum.% | |
| Panulirus argus mid-stages | 3.49 | 22.95 | 1.43 | 51.7 | 51.7 | |
| Panulirus argus late stages | 1.74 | 8.26 | 0.86 | 18.6 | 70.4 | |
| Panulirus guttatus mid-stages | 1.05 | 4.17 | 0.71 | 9.4 | 79.8 | |
| Scyllarus chacei late stages | 0.7 | 3.35 | 0.39 | 7.6 | 87.3 | |
| Palinurellus gundlachi late stages | 0.7 | 2.44 | 0.48 | 5.5 | 92.8 | |
| Autumn and Spring Cruises: Average Dissimilarity = 59.68% | ||||||
| Species | Autumn Av.Abund | Spring Av.Abund | Av.Diss | Diss/SD | Contrib% | Cum.% |
| Panulirus argus mid-stages | 3.00 | 3.49 | 14.5 | 1.2 | 24.3 | 24.3 |
| Panulirus argus late stages | 1.83 | 1.74 | 9.4 | 1.2 | 15.7 | 40.0 |
| Panulirus guttatus mid-stages | 2.12 | 1.05 | 9.3 | 1.2 | 15.6 | 55.6 |
| Panulirus guttatus late stages | 0.98 | 0.35 | 5.2 | 0.9 | 8.6 | 64.3 |
| Scyllarus chacei late stages | 0.02 | 0.70 | 4.6 | 0.6 | 7.8 | 72.0 |
| Palinurellus gundlachi late stages | 0.42 | 0.70 | 4.2 | 0.9 | 7.1 | 79.1 |
| Scyllarides aequinoctialis mid-stages | 0.29 | 0.52 | 3.6 | 0.7 | 6.1 | 85.2 |
| Parribacus antarcticus mid-stages | 0.52 | 0.00 | 2.7 | 0.6 | 4.5 | 89.7 |
| Palinurellus gundlachi mid-stages | 0.11 | 0.34 | 1.9 | 0.6 | 3.2 | 92.9 |
References
- Chan, T.Y. Annotated checklist of the world’s marine lobsters (Crustacea: Decapoda: Astacidea, Glypheidea, Achelata, Polychelida). Raffles Bull. Zool. 2010, 23, 153–181. [Google Scholar]
- Chan, T.Y. Updated checklist of the world’s marine lobsters. In Lobsters: Biology, Fisheries and Aquaculture; Radhakrishnan, E.V., Phillips, B.F., Achamveetil, G., Eds.; Springer: Singapore, 2019; pp. 35–64. [Google Scholar]
- Booth, J.D.; Webber, W.R.; Sekiguchi, H.; Coutures, E. Diverse larval recruitment strategies within the Scyllaridae. N. Z. J. Mar. Freshw. Res. 2005, 39, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, H.; Booth, J.D.; Webber, W.R. Early life histories of slipper lobsters. In The Biology and Fisheries of the Slipper Lobster; Lavalli, K.L., Spanier, E., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 69–86. [Google Scholar]
- McWilliam, P.S.; Phillips, B.F. Metamorphosis of the final phyllosoma and secondary lecithotrophy in the puerulus of Panulirus cygnus George: A review. Mar. Freshw. Res. 1997, 48, 783–790. [Google Scholar] [CrossRef]
- Booth, J.D.; Phillips, B.F. Early life history of spiny lobster. Crustaceana 1994, 66, 271–294. [Google Scholar]
- Phillips, B.F.; McWilliam, P.S. Spiny lobster development: Where does successful metamorphosis to the puerulus occur? A review. Rev. Fish Biol. Fish. 2009, 19, 193–215. [Google Scholar] [CrossRef] [Green Version]
- Lyons, W.G. Scyllarid lobsters (Crustacea, Decapoda). Mem. Hourglass Cruises 1970, 1, 1–74. [Google Scholar]
- Holthuis, L.B. Marine Lobsters of the World; FAO Species Catalogue No. 13; FAO: Rome, Italy, 1991. [Google Scholar]
- Lewis, J.B. The phyllosoma larvae of the spiny lobster Panulirus argus. Bull. Mar. Sci. Gulf Carib. 1951, 1, 89–103. [Google Scholar]
- Goldstein, J.S.; Matsuda, H.; Takenouchi, T.; Butler, M.J., IV. The complete development of larval Caribbean spiny lobster Panulirus argus (Latreille, 1804) in culture. J. Crust. Biol. 2008, 28, 306–327. [Google Scholar] [CrossRef]
- Robertson, P.B. Phyllosoma larvae of a palinurid lobster, Justitia longimana (H. Milne Edwards), from the western Atlantic. Bull. Mar. Sci. 1969, 19, 922–944. [Google Scholar]
- Robertson, P.B. The complete larval development of the sand lobster, Scyllarus americanus (Smith) (Decapoda, Scyllaridae) in the laboratory, with notes on larvae from the plankton. Bull. Mar. Sci. 1968, 18, 294–342. [Google Scholar]
- Robertson, P.B. The larvae and postlarva of the scyllarid lobster Scyllarus depressus (Smith). Bull. Mar. Sci. 1971, 21, 841–865. [Google Scholar]
- Robertson, P.B. The Larval Development of Some Western Atlantic Lobsters of the Family Scyllaridae. Ph.D. Thesis, University of Miami, Coral Gables, FL, USA, 1968. [Google Scholar]
- Robertson, P.B. Larval development of the scyllarid lobster Scyllarus planorbis (Holthuis) reared in the laboratory. Bull. Mar. Sci. 1979, 29, 320–328. [Google Scholar]
- Baisre, J.A.; Alfonso, I. Later stage larvae of Panulirus guttatus (Latreille, 1804) (Decapoda, Palinuridae) with notes on the identification of phyllosomata of Panulirus in the Caribbean Sea. Crustaceana 1994, 66, 32–44. [Google Scholar] [CrossRef]
- Goldstein, J.S.; Matsuda, H.; Matthews, T.R.; Fumihiko, A.; Yamakawa, T. Development in culture of larval spotted spiny lobster Panulirus guttatus (Latreille, 1804) (Decapoda: Achelata: Palinuridae). J. Crust. Biol. 2019, 39, 574–581. [Google Scholar] [CrossRef]
- Baisre, J.A.; Ruiz de Quevedo, M.E. Two phyllosome larvae of Panulirus laevicauda (Latreille, 1817) (Decapoda, Palinuridae) from the Caribbean Sea with a discussion about larval groups within the genus. Crustaceana 1982, 43, 147–153. [Google Scholar] [CrossRef]
- Sims, H.W., Jr. The phyllosoma larvae of the spiny lobster Palinurellus gundlachi von Martens (Decapoda, Palinuridae). Crustaceana 1966, 11, 205–215. [Google Scholar] [CrossRef] [Green Version]
- Robertson, P.B. The early larval development of the scyllarid lobster Scyllarides aequinoctialis (Lund) in the laboratory, with a revision of the larval characters of the genus. Deep Sea Res. 1969, 16, 557–586. [Google Scholar] [CrossRef]
- Sims, H.W., Jr. The phyllosoma larvae of Parribacus (Decapoda, Palinuridae). Quart. J. Florida Acad. Sci. 1965, 28, 142–172. [Google Scholar]
- Johnson, M.W. The phyllosoma larvae of slipper lobsters from the Hawaiian Islands and adjacent areas (Decapoda, Scyllaridae). Crustaceana 1971, 20, 77–103. [Google Scholar] [CrossRef]
- Ehrhardt, N.M.; Puga, R.; Butler, M.J., IV. Implications of the ecosystem approach to fisheries management in large ecosystems: The case of the Caribbean spiny lobster. In Towards Marine Ecosystem-Based Management in the Wider Caribbean; Fanning, L., Mahon, R., McConney, P., Eds.; Amsterdam University Press: Amsterdam, The Netherlands, 2011; pp. 157–175. [Google Scholar]
- Shanks, A.L. Pelagic larval duration and dispersal distance revisited. Biol. Bull. 2009, 216, 373–385. [Google Scholar] [CrossRef] [Green Version]
- Minami, H.; Inoue, N.; Sekiguchi, H. Vertical distribution of phyllosoma larvae of palinurid and scyllarid lobsters in the western North Pacific. J. Oceanogr. 2001, 57, 743–748. [Google Scholar] [CrossRef]
- Bradford, R.W.; Bruce, B.D.; Chiswell, S.M.; Booth, J.D.; Jeffs, A.; Wotherspoon, S. Vertical distribution and diurnal migration patterns of Jasus edwardsii phyllosomas off the east coast of the North Island, New Zealand. N. Z. J. Mar. Freshw. Res. 2005, 39, 593–604. [Google Scholar] [CrossRef]
- Butler, M.J., IV; Paris, C.B.; Goldstein, J.S.; Matsuda, H.; Cowen, R.K. Behavior constrains the dispersal of long-lived spiny lobster larvae. Mar. Ecol. Prog. Ser. 2011, 422, 223–237. [Google Scholar] [CrossRef] [Green Version]
- Mishra, M.; Jeffs, A.; Meyer-Rochow, V.B. Eye structure of the phyllosoma larva of the rock lobster Jasus edwardsii (Hutton, 1875): How does it differ from that of the adult? Invertebr. Reprod. Dev. 2006, 49, 213–222. [Google Scholar] [CrossRef]
- Funes-Rodríguez, R.; Ruíz-Chavarría, J.A.; González-Armas, R.; Durazo, R.; Guzmán-del Proó, S.A. Influence of hydrographic conditions on the distribution of spiny lobster larvae off the west coast of Baja California. Trans. Am. Fish. Soc. 2015, 144, 1192–1205. [Google Scholar] [CrossRef]
- Snyder, E.R.; Paris, C.B.; Vaz, C.A. How much do marine connectivity fluctuations matter? Am. Nat. 2014, 184, 523–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Segura-García, I.; Garavelli, L.; Tringali, M.; Matthews, T.; Chérubin, L.M.; Hunt, J.; Box, S.J. Reconstruction of larval origins based on genetic relatedness and biophysical modeling. Sci. Rep. 2019, 9, 7100. [Google Scholar] [CrossRef] [Green Version]
- Briones-Fourzán, P.; Candela, J.; Lozano-Álvarez, E. Postlarval settlement of the spiny lobster Panulirus argus along the Caribbean coast of Mexico: Patterns, influence of physical factors, and possible sources of origin. Limnol. Oceanogr. 2008, 53, 970–985. [Google Scholar] [CrossRef]
- Kough, A.S.; Paris, C.B.; Butler, M.J., IV. Larval connectivity and the international management of fisheries. PLoS ONE 2013, 8, e64970. [Google Scholar] [CrossRef] [Green Version]
- Lara-Hernández, J.A.; Zavala-Hidalgo, J.; Sanvicente-Añorve, L.; Briones-Fourzán, P. Connectivity and larval dispersal pathways of Panulirus argus in the Gulf of Mexico: A numerical study. J. Sea Res. 2019, 155, 101814. [Google Scholar] [CrossRef]
- Saunders, M.I.; Thompson, P.A.; Jeffs, A.G.; Säwström, C.; Sachlikidis, N.; Beckley, L.E.; Waite, A.M. Fussy feeders: Phyllosoma larvae of the Western Rock lobster (Panulirus cygnus) demonstrate prey preference. PLoS ONE 2012, 7, e36580. [Google Scholar] [CrossRef] [Green Version]
- Olvera-Limas, R.M.; Ordóñez-Alcalá, L. Distribución, abundancia relativa y desarrollo larvario de langostas Panulirus argus y Scyllarus americanus en la ZEE del Golfo de México y mar Caribe. Cienc. Pesq. 1988, 6, 7–31. [Google Scholar]
- Manzanilla-Domínguez, H.; Gasca, R. Distribution and abundance of phyllosoma larvae (Decapoda, Palinuridae) in the southern Gulf of Mexico and the western Caribbean Sea. Crustaceana 2004, 77, 75–94. [Google Scholar]
- Briones-Fourzán, P.; Lozano-Álvarez, E.; Monroy-Velázquez, L.V. Langostas Palinuroideas. In Camarones, Langostas y Cangrejos de la Costa Este de México; Hernández-Aguilera, J.L., Ruiz-Nuño, J.A., Toral-Almazán, R.E., Arenas-Fuentes, V., Eds.; CONABIO: Mexico City, Mexico, 2005; Volume 1, pp. 207–235. [Google Scholar]
- Manzanilla-Domínguez, H.; Gasca, R.; Suárez-Morales, E. Notes on the distribution of phyllosoma larvae in an oceanic atoll-like reef system in the western Caribbean. Crustaceana 2005, 78, 505–512. [Google Scholar]
- Canto-García, A.; Goldstein, J.; Sosa-Cordero, E.; Carrillo, L. Distribution and abundance of Panulirus spp. phyllosomas off the Mexican Caribbean coast. Bull. Mar. Sci. 2016, 92, 207–227. [Google Scholar] [CrossRef]
- Carrillo, L.; Largier, J.L.; Johns, E.; Smith, R.; Lamkin, J. Pathways and upper hydrography in the Mesoamerican Barrier Reef System–Part 1: Circulation. Contin. Shelf Res. 2015, 109, 164–176. [Google Scholar] [CrossRef]
- Badan, A.; Candela, J.; Sheinbaum, J.; Ochoa, J. Upper-layer circulation in the approaches to Yucatan Channel. In Circulation in the Gulf of Mexico: Observations and Models; Geophysical Monograph Series 161; American Geophysical Union: Washington, DC, USA, 2005; pp. 57–69. [Google Scholar]
- Cetina, P.; Candela, J.; Sheinbaum, J.; Ochoa, J.; Badam, A. Circulation along the Mexican Caribbean coast. J. Geophys. Res. 2006, 111, C08021. [Google Scholar] [CrossRef] [Green Version]
- Muhling, B.A.; Smith, R.H.; Vázquez-Yeomans, L.; Lamkin, J.T.; Johns, E.M.; Carrillo, L.; Sosa-Cordero, E.; Malca, E. Larval fish assemblages and mesoscale oceanographic structure along the Mesoamerican Barrier Reef System. Fish. Oceanogr. 2013, 22, 409–428. [Google Scholar] [CrossRef]
- Merino, M. Aspectos de la circulación costera superficial del Caribe mexicano con base en observaciones utilizando tarjetas de deriva. An. Inst. Cienc. del Mar y Limnol. Univ. Nal. Autón. México 1986, 13, 31–46. [Google Scholar]
- Chávez, G.; Candela, J.; Ochoa, J. Subinertial flows and transports in Cozumel Channel. J. Geophys. Res. 2003, 108, 3037. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Candela, J.; Carrillo, L.; Espinosa-Magaña, A.; Negrete-Soto, F.; Barradas-Ortiz, C.; Escalante-Mancera, E.; Muñoz de Cote-Hernández, R.; Martínez-Calderón, R.; Lozano-Álvarez, E. Metamorphosis of spiny lobsters (Panulirus argus and Panulirus guttatus) in the Yucatan current as inferred from the distribution of pueruli and final stage phyllosomata. Limnol. Oceanogr. 2021, 66, 3421–3438. [Google Scholar] [CrossRef]
- Lozano-Álvarez, E.; Briones-Fourzán, P.; Huchin-Mian, J.P.; Segura-García, I.; Ek-Huchim, J.P.; Améndola-Pimenta, M.; Rodríguez-Canul, R. Panulirus argus virus 1 detected in oceanic postlarvae of Caribbean spiny lobster: Implications for disease dispersal. Dis. Aquat. Org. 2015, 117, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Magaña, A.; Briones-Fourzán, P.; Jeffs, A.; Lozano-Álvarez, E. Energy cost of the onshore transport of postlarvae of the Caribbean spiny lobster. Bull. Mar. Sci. 2018, 94, 801–819. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Espinosa-Magaña, A.F.; Lozano-Álvarez, E.; Jeffs, A. Analysis of fatty acids to examine larval and settlement biology of the Caribbean spiny lobster Panulirus argus. Mar. Ecol. Prog. Ser. 2019, 630, 137–148. [Google Scholar] [CrossRef]
- Rimmer, D.W.; Phillips, B.F. Diurnal migration and vertical distribution of phyllosoma larvae of the Western rock lobster Panulirus cygnus. Mar. Biol. 1979, 54, 109–124. [Google Scholar] [CrossRef]
- Alfonso, I.; Frías, M.P.; Baisre, J. Distribución, abundancia y migración vertical de la fase larval de la langosta comercial Panulirus argus en aguas cubanas. Rev. Inv. Mar. 1999, 20, 23–32. [Google Scholar]
- Yeung, C.; McGowan, M.F. Differences in inshore-offshore and vertical distribution of phyllosoma larvae of Panulirus, Scyllarus and Scyllarides in the Florida Keys in May-June, 1989. Bull. Mar. Sci. 1991, 49, 699–714. [Google Scholar]
- Robertson, P.B. A giant phyllosoma larva from the Caribbean Sea, with notes on smaller specimens. Crustaceana 1968, 15 (Suppl. 2), 83–97. [Google Scholar]
- Palero, F.; Guerao, G.; Hall, M.; Chan, T.Y.; Clark, P.F. The ‘giant phyllosoma’ are larval stages of Parribacus antarcticus (Decapoda: Scyllaridae). Invertebr. Syst. 2014, 28, 258–276. [Google Scholar] [CrossRef]
- Phillips, B.F.; Rimmer, D.W.; Reid, D.D. Ecological investigations of the late-stage phyllosoma and puerulus larvae of the western rock lobster Panulirus longipes cygnus. Mar. Biol. 1978, 45, 347–357. [Google Scholar] [CrossRef]
- Phillips, B.F.; Brown, P.A.; Rimmer, D.W.; Braine, S.J. Distribution and dispersal of the phyllosoma larvae of the western rock lobster Panulirus cygnus, in the south-eastern Indian Ocean. Aust. J. Mar. Freshw. Res. 1979, 30, 773–783. [Google Scholar] [CrossRef]
- Mallol, S.; Mateo-Ramírez, A.; Alemany, F.; Álvarez-Berastegui, D.; Díaz, D.; López-Jurado, J.L.; Goñi, R. Abundance and distribution of scyllarid phyllosoma larvae (Decapoda: Scyllaridae) in the Balearic Sea (Western Mediterranean). J. Crust. Biol. 2014, 34, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Morris, E.K.; Caruso, T.; Buscot, F.; Fischer, M.; Hancock, C.; Maier, T.S.; Meiners, T.; Müller, C.; Obermaier, E.; Prati, D.; et al. Choosing and using diversity indices: Insights for ecological applications from the German Biodiversity Exploratories. Ecol. Evol. 2014, 4, 3514–3524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phleger, C.F.; Nelson, M.M.; Mooney, B.D.; Nichols, P.D.; Ritar, A.J.; Smith, G.G.; Hart, P.R.; Jeffs, A.G. Lipids and nutrition of the southern rock lobster, Jasus edwardsii, from hatch to puerulus. Mar. Freshw. Res. 2001, 52, 1475–1486. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; O’Rorke, R.; Waite, A.M.; Beckley, L.E.; Thompson, P.; Jeffs, A.G. Fatty acid profiles of phyllosoma larvae of western rock lobster (Panulirus cygnus) in cyclonic and anticyclonic eddies of the Leeuwin Current off Western Australia. Prog. Oceanogr. 2014, 122, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, A.; Gasca, R.; Sosa-Cordero, E.; Camacho-Cruz, K. Stable carbon and nitrogen isotopes in Panulirus argus phyllosomas in the Mexican Caribbean. Reg. Stud. Mar. Sci. 2021, 42, 101617. [Google Scholar] [CrossRef]
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Anterpretation, 2nd ed.; PRIMER-E: Plymouth, UK, 2001. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. PRIMER v6: User/Manual Tutorial; PRIMER-E: Plymouth, UK, 2006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Phillips, B.F.; Booth, J.D.; Stanley, J.C.; Jeffs, A.G.; McWilliam, P. Larval and postlarval ecology. In Lobsters: Biology, Management, Aquaculture and Fisheries; Phillips, B.F., Ed.; Blackwell: Oxford, UK, 2006; pp. 231–235. [Google Scholar]
- Johnson, M.W. Production and distribution of larvae of the spiny lobster Panulirus interruptus (Randall) with records on P. gracilis (Streets). Bull. Scripps Inst. Oceanogr. 1960, 7, 412–461. [Google Scholar]
- Lazarus, B.I. The occurrence of phyllosomata off the Cape with particular reference to Jasus lalandii. S. Afr. Div. Sea Fish. Invest. Rep. 1967, 63, 1–38. [Google Scholar]
- Lesser, J.H.R. Phyllosoma larvae of Jasus edwardsii (Hutton) (Crustacea: Decapoda: Palinuridae) and their distribution off the east coast of the North Island, New Zealand. N. Z. J. Mar. Freshw. Res. 1978, 12, 357–370. [Google Scholar] [CrossRef]
- Inoue, N.; Minami, H.; Sekiguchi, H. Distribution of phyllosoma larvae (Crustacea: Decapoda: Palinuridae, Scyllaridae and Synaxidae) in the western north Pacific. J. Oceanogr. 2004, 60, 963–976. [Google Scholar] [CrossRef]
- Johnson, M.J. The palinurid and scyllarid lobster larvae of the tropical eastern Pacific and their distribution as related to the prevailing hydrography. Bull. Scripps Inst. Oceanogr. 1971, 19, 1–36. [Google Scholar]
- Sims, H.W., Jr.; Ingle, R.M. Caribbean recruitment of Florida’s spiny lobster population. Quart. J. Florida Acad. Sci. 1966, 29, 207–242. [Google Scholar]
- Yeung, C.; Jones, D.L.; Criales, M.M.; Jackson, T.L.; Richards, W.J. Influence of coastal eddies and counter-currents on the influx of spiny lobster, Panulirus argus, postlarvae into Florida Bay. Mar. Freshw. Res. 2001, 52, 1217–1232. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Lozano-Álvarez, E. Essential habitats for Panulirus spiny lobsters. In Lobsters: Biology, Management, Aquaculture and Fisheries, 2nd ed.; Phillips, B.F., Ed.; Wiley-Blackwell: Oxford, UK, 2013; pp. 186–220. [Google Scholar]
- Bertelsen, R.D.; Matthews, T.R. Fecundity dynamics of female spiny lobster (Panulirus argus) in a south Florida fishery and Dry Tortugas National Park lobster sanctuary. Mar. Freshw. Res. 2001, 52, 1559–1565. [Google Scholar] [CrossRef]
- Fonseca-Larios, M.E.; Briones-Fourzán, P. Fecundity of the spiny lobster Panulirus argus (Latreille, 1804) in the Caribbean coast of Mexico. Bull. Mar. Sci. 1998, 63, 21–32. [Google Scholar]
- Castillo, A.; Lessios, H.A. Lobster fishery by the Kuna Indians in the San Blas Region of Panama (Kuna Yala). Crustaceana 2001, 74, 459–475. [Google Scholar]
- Yallonardo, M.; Posada, J.M.; Schweizer, D.M. Current status of the spiny lobster, Panulirus argus, fishery in the Los Roques Archipelago National Park, Venezuela. Mar. Freshw. Res. 2001, 52, 1615–1622. [Google Scholar] [CrossRef]
- Martínez-Calderón, R.; Lozano-Álvarez, E.; Briones-Fourzán, P. Morphometric relationships and seasonal variation in size, weight, and a condition index of post-settlement stages of the Caribbean spiny lobster. PeerJ 2018, 6, e5297. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, H.; Yamakawa, T. Effects of temperature on growth of the Japanese spiny lobster, Panulirus japonicus phyllosomas under laboratory conditions. Mar. Freshw. Res. 1997, 48, 791–796. [Google Scholar] [CrossRef]
- Marfin, J.P. Biologie et pêche de la langouste Panulirus guttatus en Martinique. Sci. Pêche 1978, 278, 1–10. [Google Scholar]
- Sharp, W.C.; Hunt, J.H.; Lyons, W.G. Life history of the spotted spiny lobster, Panulirus guttatus, an obligate reef-dweller. Mar. Freshw. Res. 1997, 28, 142–172. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Contreras-Ortiz, G. Reproduction of the spiny lobster Panulirus guttatus (Decapoda: Palinuridae) on the Caribbean Coast of Mexico. J. Crust. Biol. 1999, 19, 171–179. [Google Scholar] [CrossRef]
- Losada-Tostesón, V.; Posada, J.M.; Losada, F. Size and reproductive status of fished spotted spiny lobster, Panulirus guttatus, in Morrocoy National Park, Venezuela: A preliminary report. Mar. Freshw. Res. 2001, 52, 1599–1603. [Google Scholar] [CrossRef]
- Wynne, S.; Côté, I. Effects of habitat quality and fishing on Caribbean spotted spiny lobster populations. J. Appl. Ecol. 2007, 44, 488–494. [Google Scholar] [CrossRef]
- Monod, T.; Postel, E. Notes sur une langouste brévicorne peu connue, Justitia longimana (H. Milne Edwards). Crustaceana 1968, 14, 178–184. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Barradas-Ortiz, C.; Negrete-Soto, F.; Segura-García, I.; Lozano-Álvarez, E. Occurrence of Panulirus meripurpuratus and P. laevicauda (Decapoda: Achelata: Palinuridae) in Bahía de la Ascensión, Mexico. Lat. Am. J. Aquat. Res. 2019, 47, 694–698. [Google Scholar]
- Webber, W.R.; Booth, J.D. Taxonomy and evolution. In The Biology and Fisheries of the Slipper Lobster; Lavalli, K.L., Spanier, E., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 25–52. [Google Scholar]
- Sharp, W.C.; Hunt, J.H.; Teehan, W.H. Observations on the ecology of Scyllarides aequinoctialis, Scyllarides nodifer, and Parribacus antarcticus and a description of the Florida scyllarid lobster fishery. In The Biology and Fisheries of the Slipper Lobster; Lavalli, K.L., Spanier, E., Eds.; CRC Press: Boca Raton, FL, USA, 2007; pp. 231–242. [Google Scholar]
- Holthuis, L.B. Biological investigations of the deep sea. 42. A new species of shovel-nose lobster, Scyllarus planorbis, from the southwestern Caribbean and northern South America. Bull. Mar. Sci. 1969, 19, 149–158. [Google Scholar]
- Navas, G.R.; Campos, N.H. Las langostas chinas (Crustacea: Decapoda: Scyllaridae) del Caribe colombiano. Bol. Invest. Mar. Cost. 1998, 27, 51–66. [Google Scholar] [CrossRef]
- Yeung, C.; Lee, T.E. Larval transport and retention of the spiny lobster, Panulirus argus, in the coastal zone of the Florida Keys, USA. Fish. Oceanogr. 2002, 11, 286–309. [Google Scholar] [CrossRef]
- Chiswell, S.M.; Booth, J.D. Rock lobster Jasus edwardsii larval retention by the Wairarapa Eddy off New Zealand. Mar. Ecol. Prog. Ser. 1999, 183, 227–240. [Google Scholar] [CrossRef]
- Chiswell, S.M.; Booth, J.D. Distribution of mid- and late-stage Jasus edwardsii phyllosomas: Implications for larval recruitment processes. N. Z. J. Mar. Freshw. Res. 2005, 39, 1157–1170. [Google Scholar] [CrossRef] [Green Version]
- Yoshimura, T.; Yamakawa, H.; Kozasa, E. Distribution of final stage phyllosoma larvae and free-swimming pueruli of Panulirus japonicus around the Kuroshio Current off southern Kyusyu, Japan. Mar. Biol. 1999, 133, 293–306. [Google Scholar] [CrossRef]
- Inoue, N.; Sekiguchi, H. Distribution of late-stage phyllosoma larvae of Panulirus japonicus in the Kuroshio Subgyre. Mar. Freshw. Res. 2001, 52, 1201–1209. [Google Scholar] [CrossRef]
- Truelove, N.K.; Griffiths, S.; Ley-Cooper, K.; Azueta, J.; Majil, I.; Box, S.J.; Behringer, D.C.; Butler, M.J., IV; Preziosi, R.F. Genetic evidence from the spiny lobster fishery supports international cooperation among Central American marine protected areas. Conserv. Genet. 2015, 16, 347–358. [Google Scholar] [CrossRef]
- Caputi, N.; de Lestang, S.; Frusher, S.; Wahle, R.A. The impact of climate change on exploited lobster stocks. In Lobsters: Biology, Management, Aquaculture and Fisheries, 2nd ed.; Phillips, B.F., Ed.; Wiley-Blackwell: Oxford, UK, 2013; pp. 84–112. [Google Scholar]
- Phillips, B.F.; Pérez-Ramírez, M.; de Lestang, S. Lobsters in a changing climate. In Climate Change Impact on Fisheries and Aquaculture: A Global Analysis; Phillips, B.F., Pérez-Ramírez, M., Eds.; Wiley-Blackwell: Hoboken, NJ, USA, 2018; pp. 815–849. [Google Scholar]
- Woodings, L.N.; Murphy, N.P.; Jeffs, A.; Suthers, I.M.; Liggins, G.W.; Strugnell, J.M. Distribution of Palinuridae and Scyllaridae phyllosoma larvae within the East Australian Current: A climate change hot spot. Mar. Freshw. Res. 2019, 70, 1020–1033. [Google Scholar] [CrossRef]











| Species | Early Stages | Mid Stages | Late Stages |
|---|---|---|---|
| Panulirus argus | I–III | IV–VIII | IX–X |
| Panulirus guttatus | I–III | IV–VIII | IX–XI |
| Panulirus laevicauda | I–III | IV–VIII | IX–XI |
| Palinurellus gundlachi | I–V | VI–VIII | IX–XII |
| Justitia longimana | I–IV | V–VIII | IX–X |
| Parribacus antarcticus | I–V | VI–VIII | IX–XI |
| Scyllarides aequinoctialis | I–V | VI–IX | X–XI |
| Scyllarus chacei | I–III | IV–V | VI–VII |
| Species | Autumn 2012 | Spring 2013 | ||||||
|---|---|---|---|---|---|---|---|---|
| N | Early | Mid | Late | N | Early | Mid | Late | |
| Panulirus argus | 1049 | 0 | 763 | 286 | 662 | 0 | 476 | 153 |
| Panulirus guttatus | 497 | 0 | 392 | 105 | 68 | 0 | 56 | 11 |
| Panulirus laevicauda | 1 | 0 | 0 | 1 | 2 | 0 | 0 | 2 |
| Palinurellus gundlachi | 46 | 1 | 9 | 36 | 61 | 14 | 12 | 35 |
| Justitia longimana | 38 | 0 | 25 | 13 | 1 | 0 | 1 | 0 |
| Parribacus antarcticus | 51 | 1 | 46 | 4 | 0 | 0 | 0 | 0 |
| Scyllarides aequinoctialis | 32 | 1 | 23 | 8 | 22 | 1 | 20 | 1 |
| Scyllarus chacei | 1 | 0 | 0 | 1 | 32 | 0 | 0 | 32 |
| Species and Stage | N | BL (Range) | BL (Mean ± SD) | CL (Mean ± SD) | CW (Mean ± SD) |
|---|---|---|---|---|---|
| Panulirus argus | |||||
| Stage V | 54 | 4.3–8.1 | 6.0 ± 0.9 | 4.3 ± 0.8 | 2.3 ± 0.4 |
| Stage VI | 403 | 5.7–12.8 | 9.3 ± 1.4 | 6.8 ± 1.1 | 3.7 ± 0.6 |
| Stage VII | 424 | 9.2–17.0 | 13.3 ± 1.4 | 9.6 ± 1.1 | 5.4 ± 0.6 |
| Stage VIII | 319 | 13.4–22.6 | 18.4 ± 1.8 | 12.4 ± 1.2 | 7.2 ± 0.7 |
| Stage IX | 209 | 18.0–23.0 | 20.1 ± 0.9 | 13.3 ± 1.1 | 7.8 ± 0.6 |
| Stage X | 64 | 22.0–30.0 | 25.0 ± 1.3 | 15.1 ± 1.1 | 8.5 ± 0.7 |
| Panulirus guttatus | |||||
| Stage V | 28 | 6.9–11.0 | 9.2 ± 1.1 | 6.7 ± 0.8 | 3.6 ± 0.6 |
| Stage VI | 234 | 10.0–16.4 | 12.6 ± 1.4 | 9.4 ± 1.4 | 5.3 ± 0.8 |
| Stage VII | 123 | 11.5–21.0 | 16.7 ± 1.9 | 12.5 ± 1.8 | 7.3 ± 1.4 |
| Stage VIII | 64 | 14.5–28.5 | 21.7 ± 2.1 | 16.0 ± 1.5 | 9.6 ± 1.2 |
| Stage IX | 43 | 19.5–32.1 | 26.9 ± 2.7 | 19.1 ± 1.9 | 11.6 ± 1.6 |
| Stage X | 10 | 28.0–33.0 | 30.6 ± 1.4 | 21.0 ± 1.0 | 13.0 ± 0.5 |
| Stage XI | 4 | 37.5–42.5 | 39.8 ± 2.1 | 24.8 ± 0.9 | 14.9 ± 0.8 |
| Palinurellus gundlachi | |||||
| Stage V | 11 | 5.0–6.5 | 5.9 ± 0.6 | 3.7 ± 0.9 | 3.8 ± 1.0 |
| Stage VI | 5 | 6.3–7.0 | 6.7 ± 0.3 | 5.0 ± 0.4 | 4.4 ± 0.4 |
| Stage VII | 3 | 7.4–8.5 | 8.0 ± 0.6 | 5.3 ± 0.6 | 5.8 ± 0.2 |
| Stage VIII | 9 | 8.5–11.2 | 9.6 ± 0.8 | 7.0 ± 0.7 | 6.3 ± 1.2 |
| Stage IX | 8 | 9.0–10.7 | 10.0 ± 0.6 | 6.6 ± 1.2 | 6.8 ± 0.4 |
| Stage X | 18 | 11.0–13.5 | 11.7 ± 0.7 | 8.0 ± 0.7 | 8.1 ± 1.2 |
| Stage XI | 14 | 11.3–16.0 | 13.0 ± 1.6 | 8.9 ± 1.1 | 8.3 ± 0.8 |
| Stage XII | 33 | 12.2–22.0 | 15.7 ± 1.1 | 10.4 ± 1.2 | 9.4 ± 0.7 |
| Justitia longimana | |||||
| Stage V | 4 | 8.0–10.0 | 8.8 ± 0.9 | 6.8 ± 0.6 | 4.1 ± 0.6 |
| Stage VI | 8 | 11.3–16.1 | 13.5 ± 1.6 | 10.6 ± 1.2 | 7.0 ± 1.2 |
| Stage VII | 5 | 17.0–20.0 | 18.1 ± 1.2 | 13.9 ± 1.0 | 10.1 ± 1.4 |
| Stage VIII | 6 | 20.0–23.8 | 21.0 ± 1.4 | 14.8 ± 2.3 | 11.8 ± 0.7 |
| Stage IX | 12 | 24.0–28.0 | 26.0 ± 1.3 | 18.3 ± 1.0 | 14.4 ± 0.8 |
| Stage X | 1 | 35.5 | 22.6 | 16.6 | |
| Panulirus laevicauda | |||||
| Stage IX | 1 | 14.8 | 11.5 | 7.0 | |
| Stage X | 1 | 16.5 | 12.5 | 8.0 | |
| Stage XI | 1 | 22.5 | 17.0 | 10.5 |
| Species and Stage | N | BL (Range) | BL (mean ± SD) | CL (mean ± SD) | CW (mean ± SD) |
|---|---|---|---|---|---|
| Parribacus antarcticus | |||||
| Stage V | 1 | 6.0 | 4.8 | 2.7 | |
| Stage VI | 24 | 8.5–15.0 | 11.0 ± 1.7 | 8.9 ± 1.6 | 5.8 ± 1.1 |
| Stage VII | 12 | 13.0–17.5 | 15.3 ± 1.3 | 12.5 ± 1.0 | 9.3 ± 3.2 |
| Stage VIII | 9 | 18.8–28.3 | 21.3 ± 2.8 | 17.4 ± 2.1 | 13.0 ± 3.0 |
| Stage IX | 2 | 22.9–25.1 | 24.0 ± 1.6 | 20.0 ± 1.3 | 14.9 ± 1.2 |
| Stage X | 2 | 33.0–36.1 | 34.6 ± 2.2 | 28.3 ± 1.8 | 22.3 ± 1.5 |
| Scyllarides aequinoctialis | |||||
| Stage IV | 2 | 4.0–4.4 | 4.2 ± 0.3 | 3.4 ± 0.6 | 1.9 ± 0.2 |
| Stage VI | 7 | 6.3–9.8 | 7.8 ± 1.1 | 6.0 ± 1.1 | 3.8 ± 0.7 |
| Stage VII | 9 | 7.3–10.3 | 9.3 ± 0.8 | 7.7 ± 1.8 | 4.5 ± 0.6 |
| Stage VIII | 13 | 10.3–18.4 | 14.4 ± 2.5 | 11.1 ± 2.0 | 7.6 ± 1.6 |
| Stage IX | 13 | 18.0–26.0 | 23.3 ± 2.7 | 18.6 ± 2.0 | 14.0 ± 2.1 |
| Stage X | 7 | 27.0–33.0 | 29.8 ± 2.3 | 22.2 ± 1.5 | 17.1 ± 1.3 |
| Stage XI | 2 | 38.6–41.2 | 39.9 ± 1.8 | 27.4 ± 1.5 | 20.1 ± 0.8 |
| Stage XII | 1 | 42.0 | 28.0 | 22.0 | |
| Scyllarus chacei | |||||
| Stage VI | 2 | 5.0–7.3 | 6.2 ± 1.6 | 4.2 ± 1.1 | 4.7 ± 0.7 |
| Stage VII | 31 | 9.2–11.4 | 10.5 ± 0.5 | 5.9 ± 0.5 | 7.1 ± 1.3 |
| Ecological Index | Autumn 2012 | Spring 2013 | df | t | p |
|---|---|---|---|---|---|
| S | 4.9 ± 0.29 | 4.3 ± 0.38 | 85 | 1.276 | 0.2056 |
| N | 29.0 ± 3.28 | 28.5 ± 4.97 | 85 | 0.082 | 0.9348 |
| H′ | 1.2396 ± 0.0425 | 1.0023 ± 0.0625 | 76 | 3.15 | 0.0050 |
| J′ | 0.7380 ± 0.0159 | 0.6989 ± 0.0258 | 76 | 2.888 | 0.0023 |
| λ | 0.3203 ± 0.0163 | 0.4472 ± 0.0256 | 76 | 4.303 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz de Cote-Hernández, R.; Briones-Fourzán, P.; Barradas-Ortiz, C.; Negrete-Soto, F.; Lozano-Álvarez, E. Diversity and Distribution of Mid- to Late-Stage Phyllosomata of Spiny and Slipper Lobsters (Decapoda: Achelata) in the Mexican Caribbean. Diversity 2021, 13, 485. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100485
Muñoz de Cote-Hernández R, Briones-Fourzán P, Barradas-Ortiz C, Negrete-Soto F, Lozano-Álvarez E. Diversity and Distribution of Mid- to Late-Stage Phyllosomata of Spiny and Slipper Lobsters (Decapoda: Achelata) in the Mexican Caribbean. Diversity. 2021; 13(10):485. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100485
Chicago/Turabian StyleMuñoz de Cote-Hernández, Rubén, Patricia Briones-Fourzán, Cecilia Barradas-Ortiz, Fernando Negrete-Soto, and Enrique Lozano-Álvarez. 2021. "Diversity and Distribution of Mid- to Late-Stage Phyllosomata of Spiny and Slipper Lobsters (Decapoda: Achelata) in the Mexican Caribbean" Diversity 13, no. 10: 485. https://0-doi-org.brum.beds.ac.uk/10.3390/d13100485