
Uncovering Hidden Diversity: Three New Species of the Keratella Genus (Rotifera, Monogononta, Brachionidae) of High Altitude Water Systems from Central Mexico †


Abstract
:1. Introduction
2. Materials and Methods
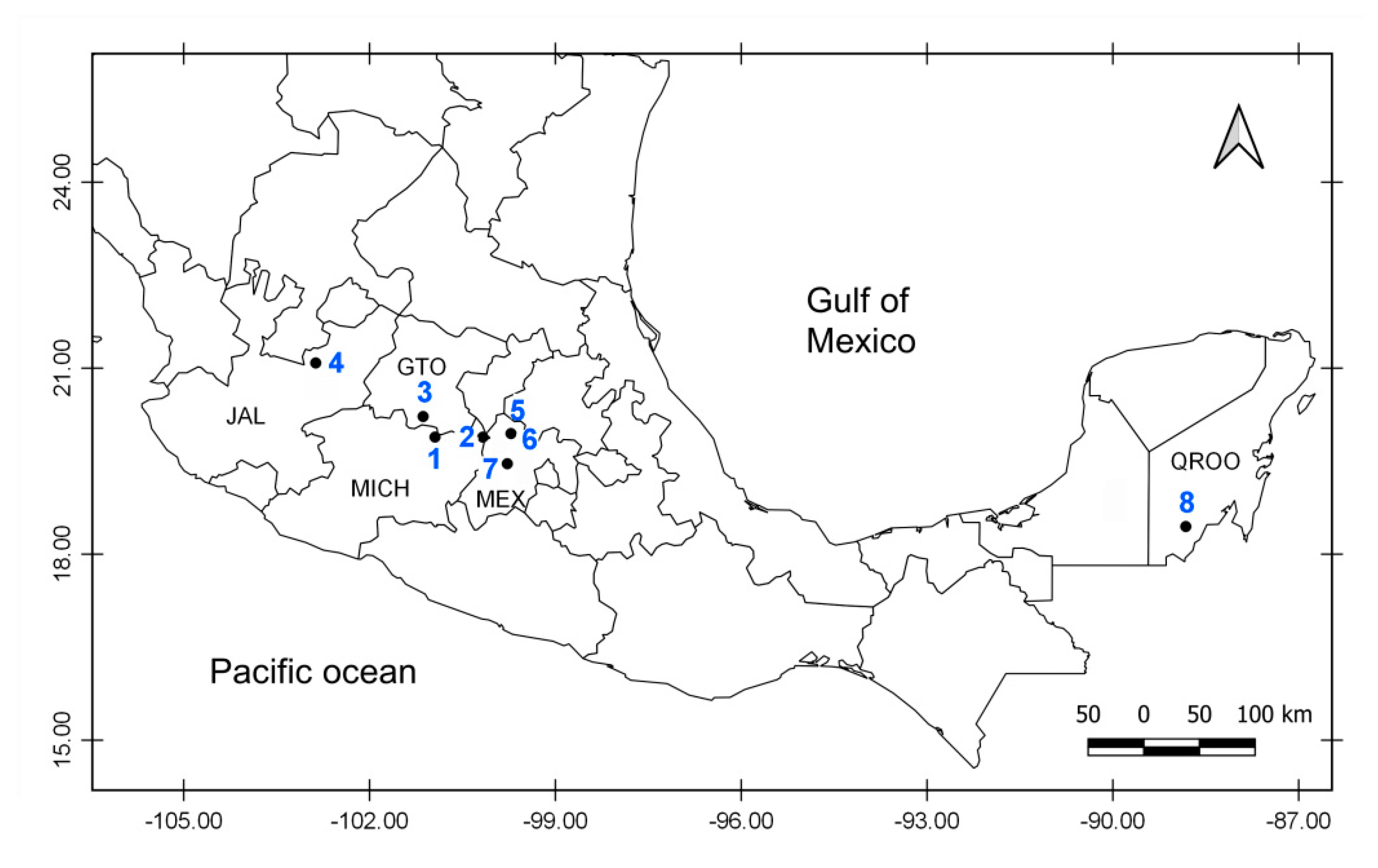
2.1. Sampling
2.2. DNA Extraction and Amplification
2.3. Alignment and Phylogenetic Analyses
2.4. Species Delimitation
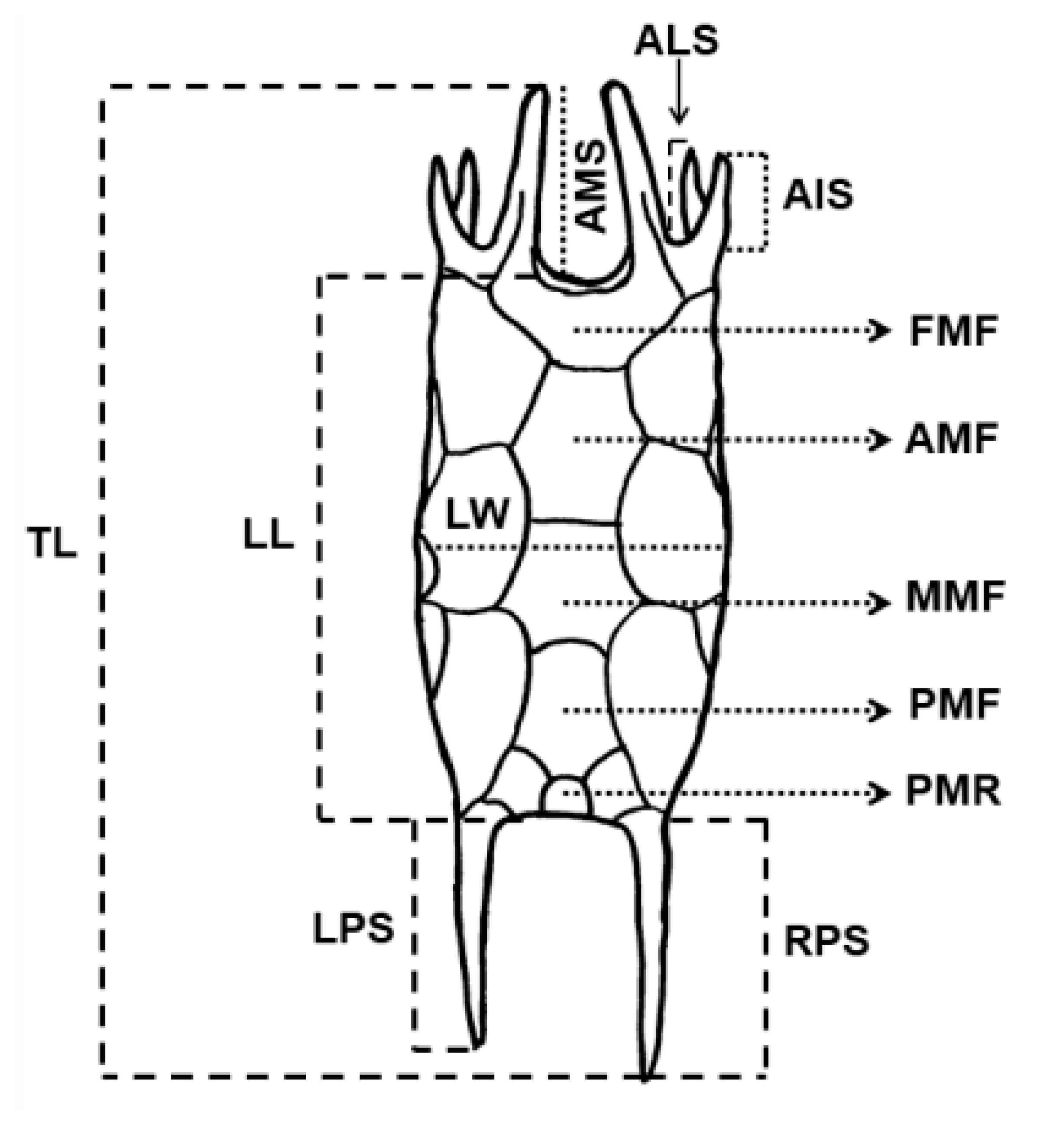
2.5. Measurements of Specimens and Morphological Analyses
2.6. Statistical Analysis of Morphological Measurements
3. Results
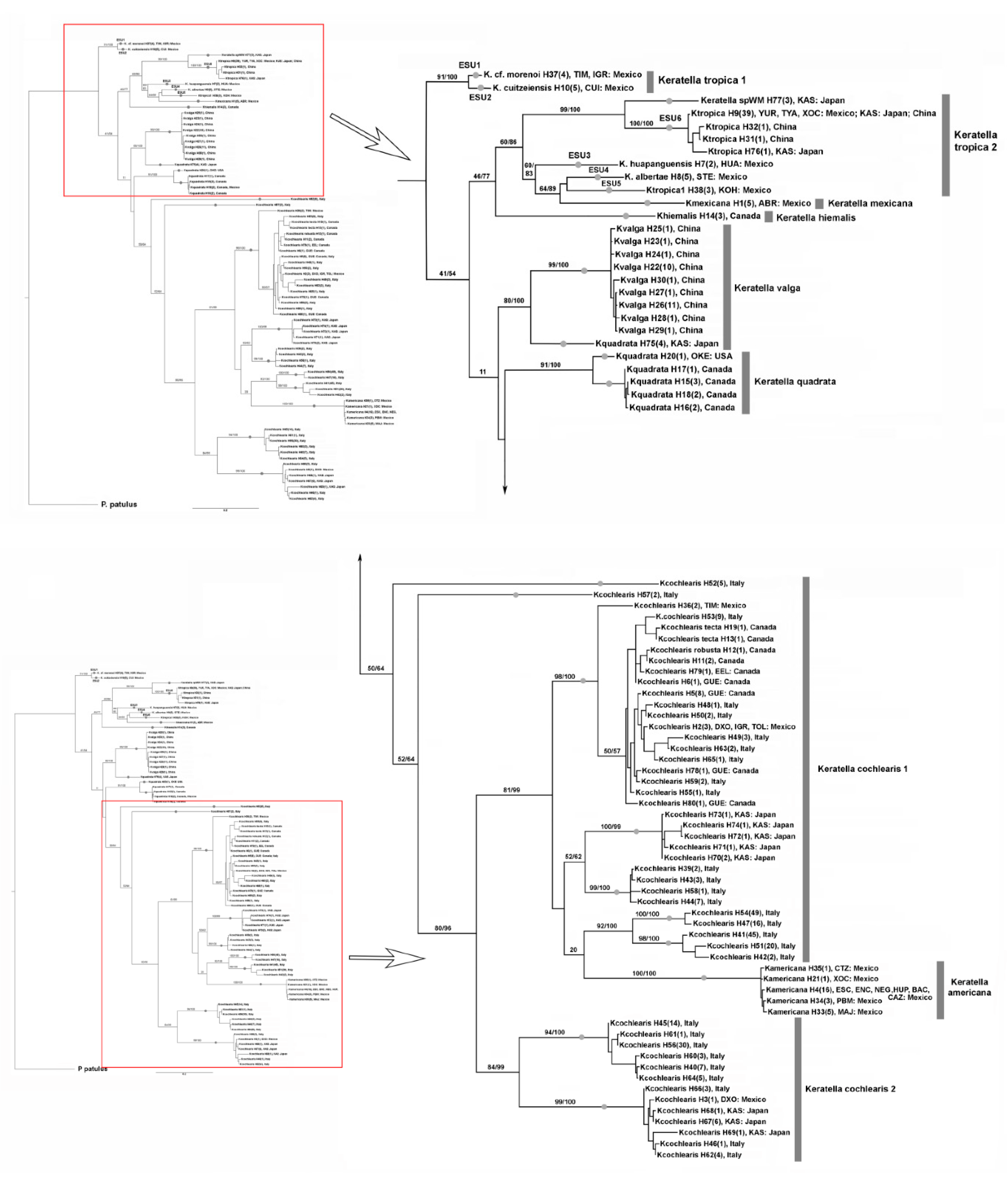
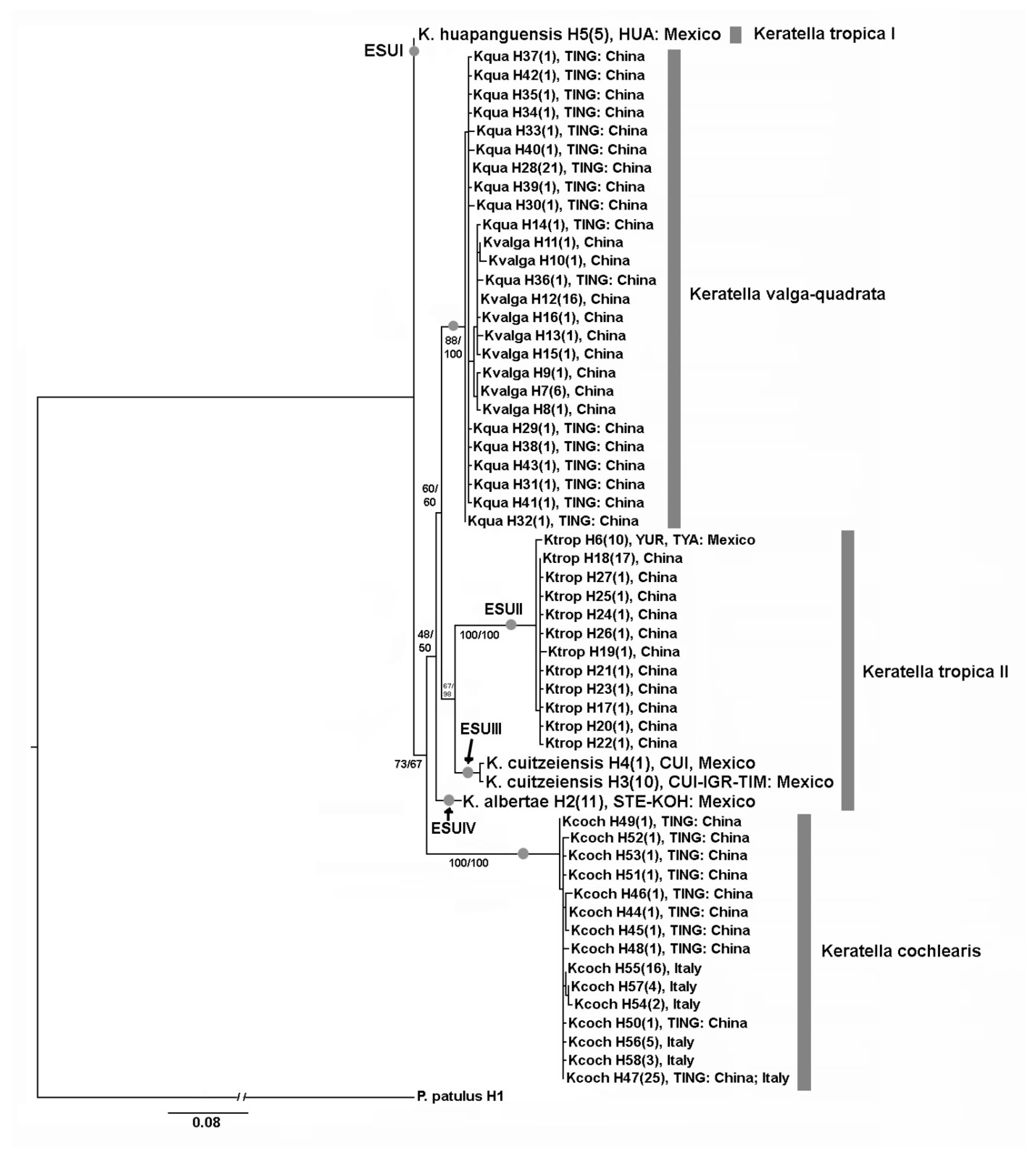
3.1. DNA Taxonomy
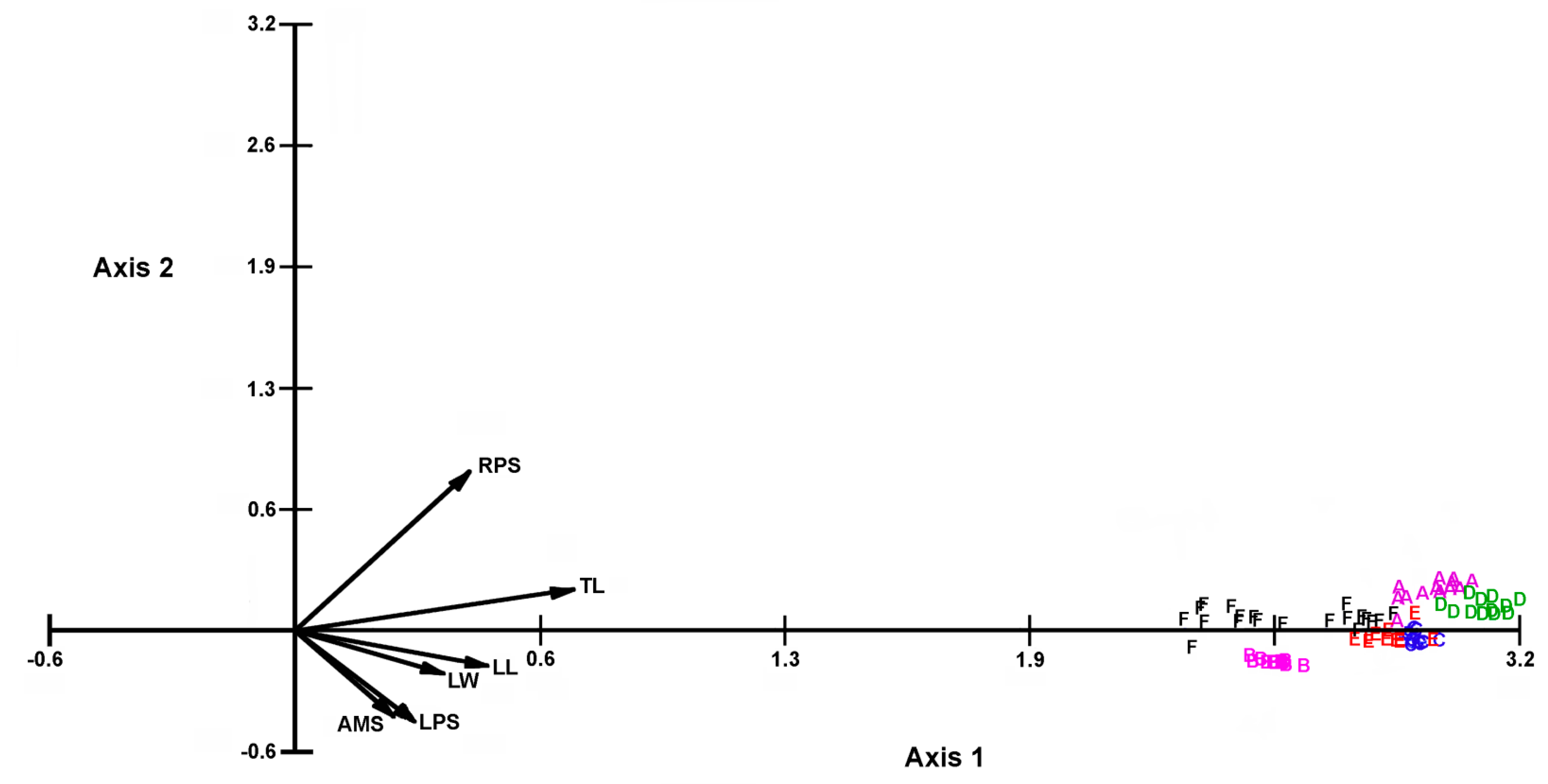
3.2. Statistical Analysis of Morphometric Measurements
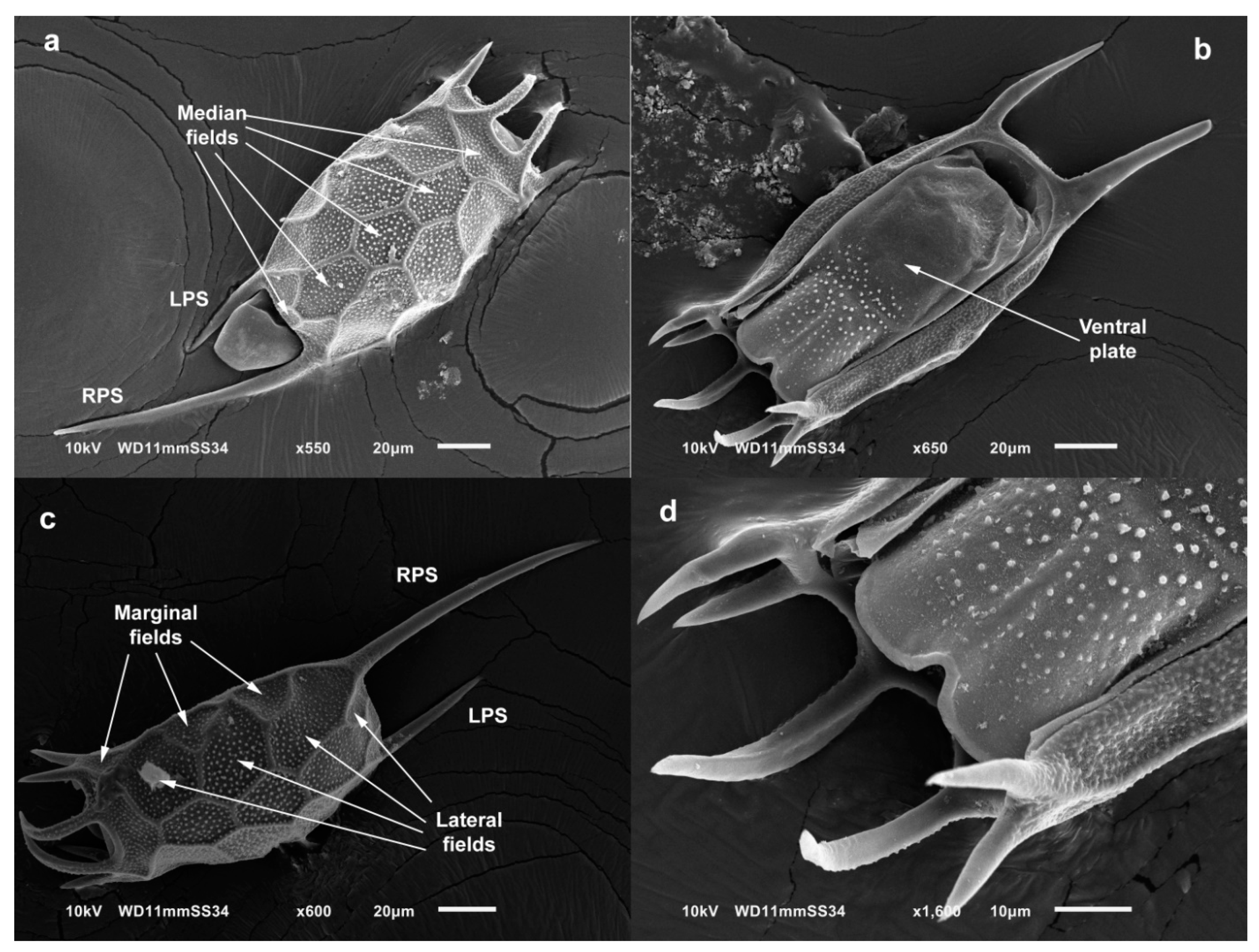
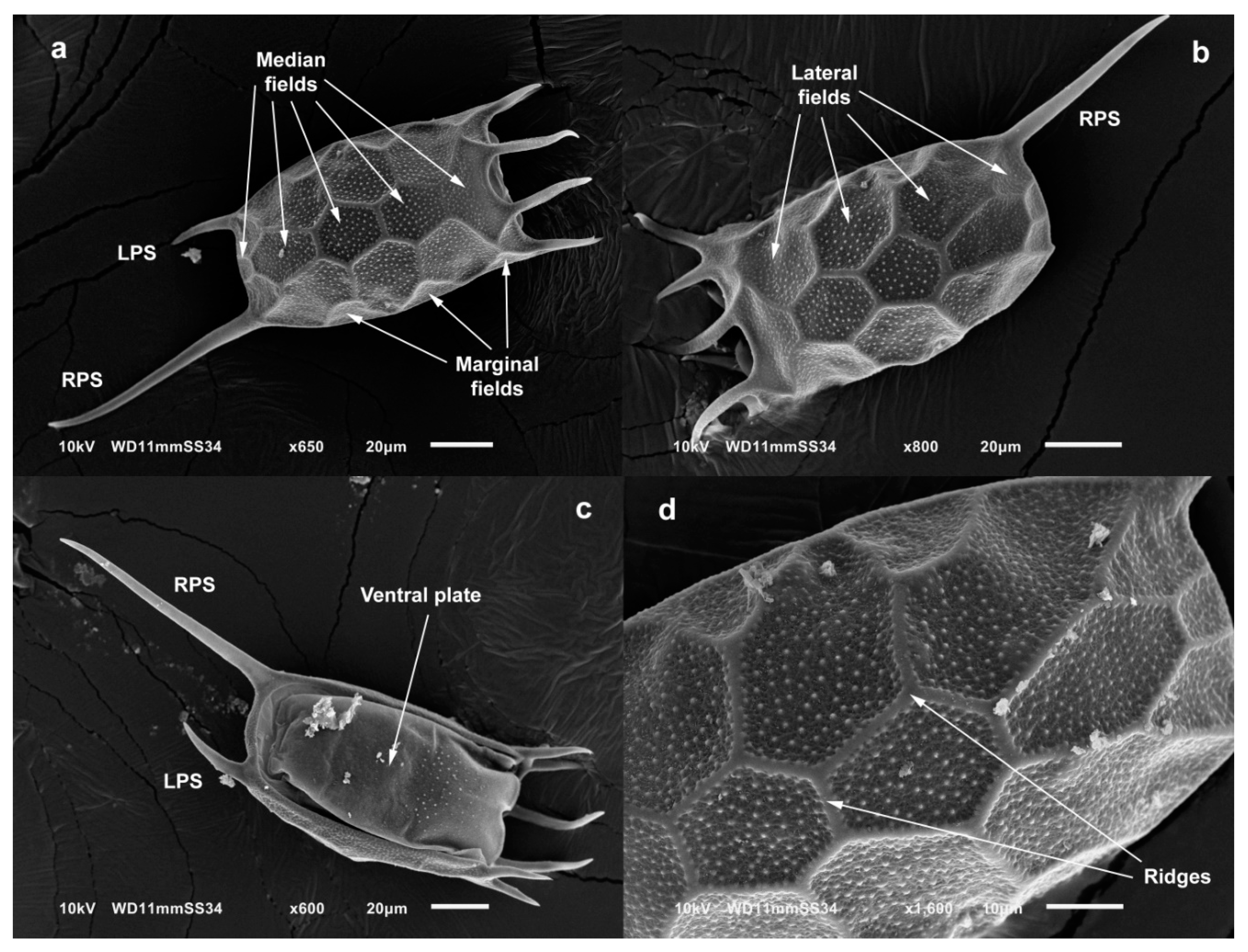
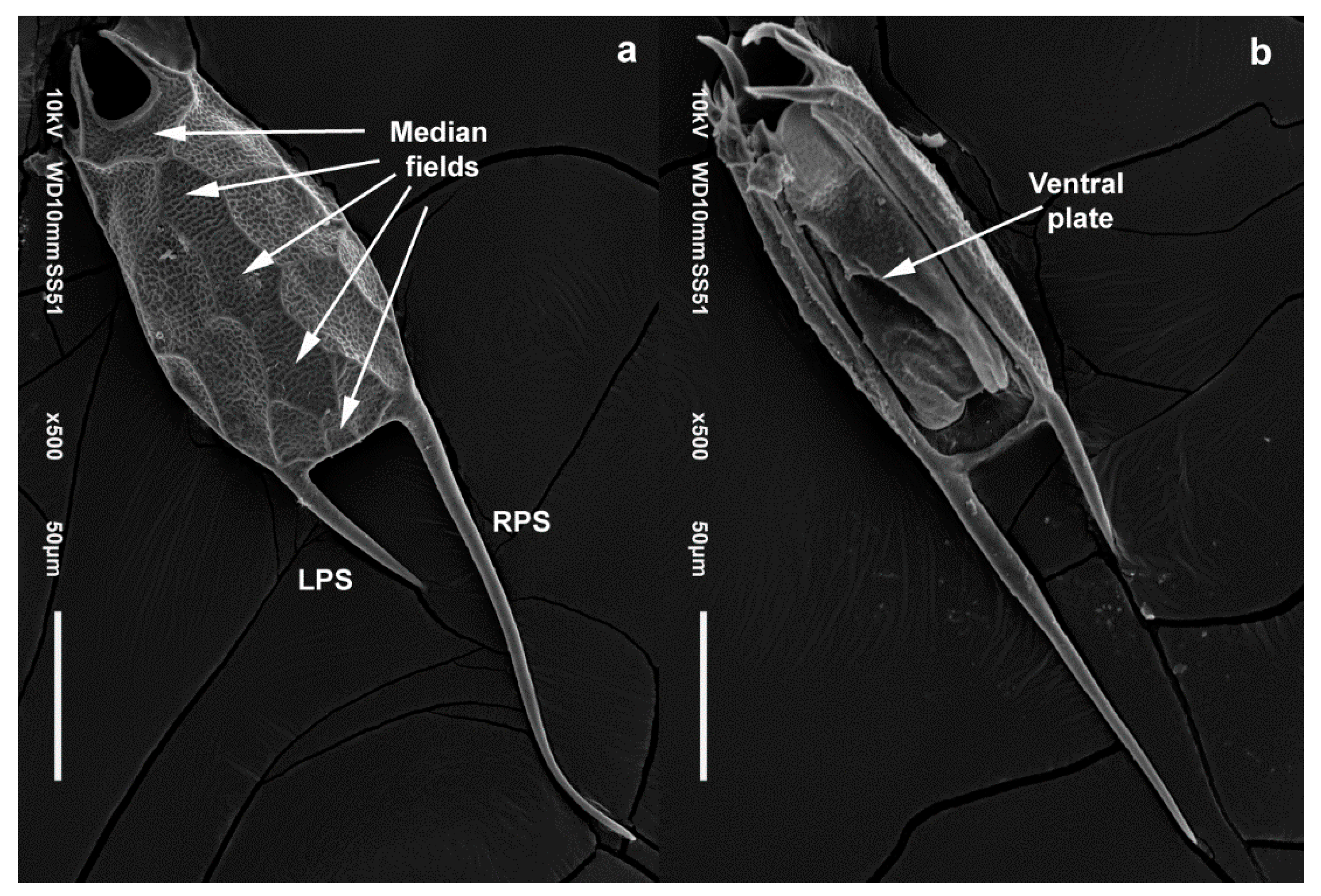
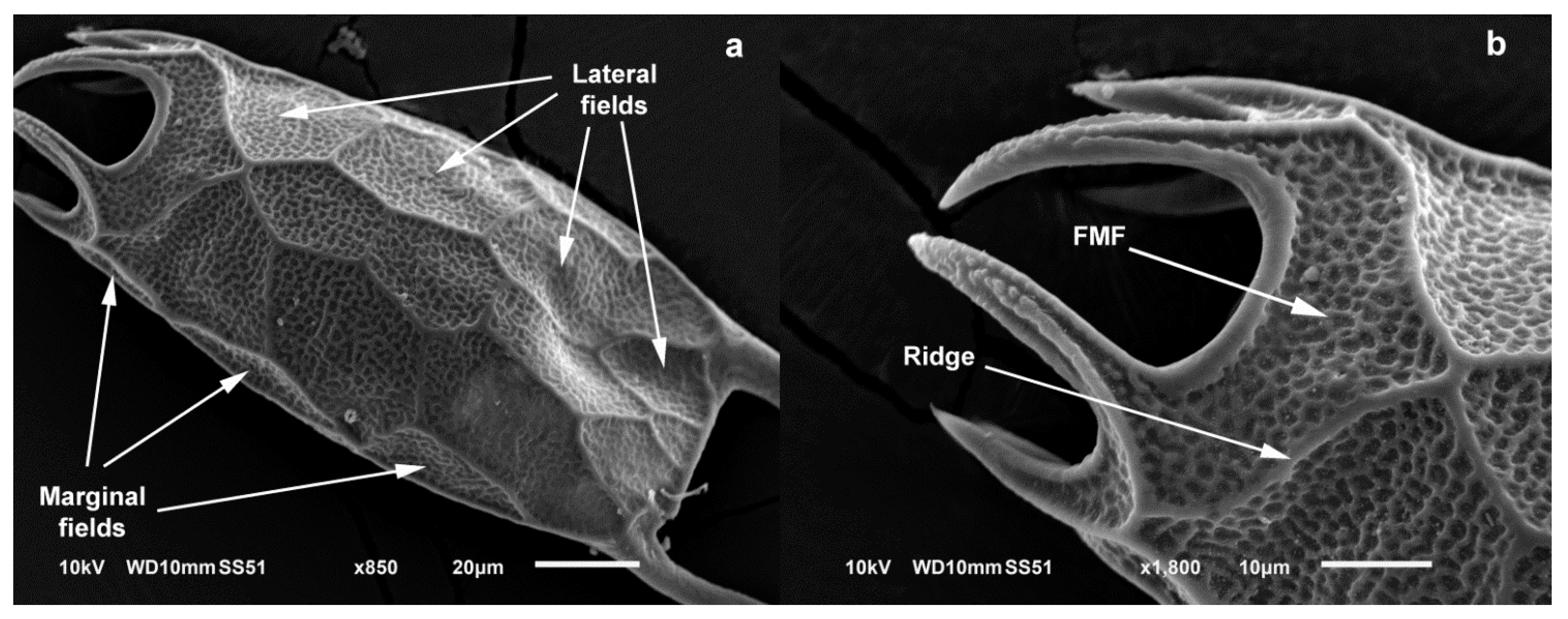
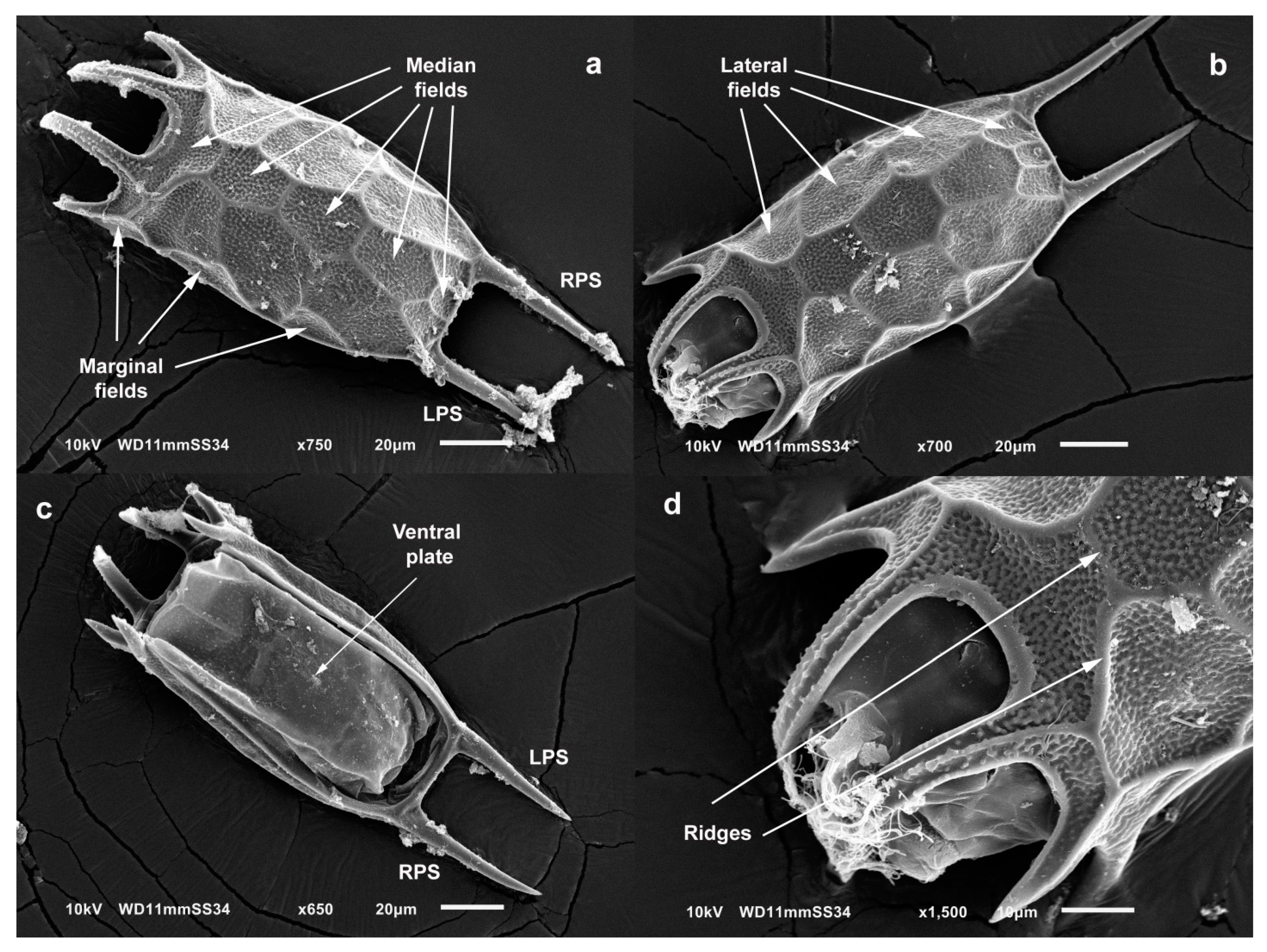
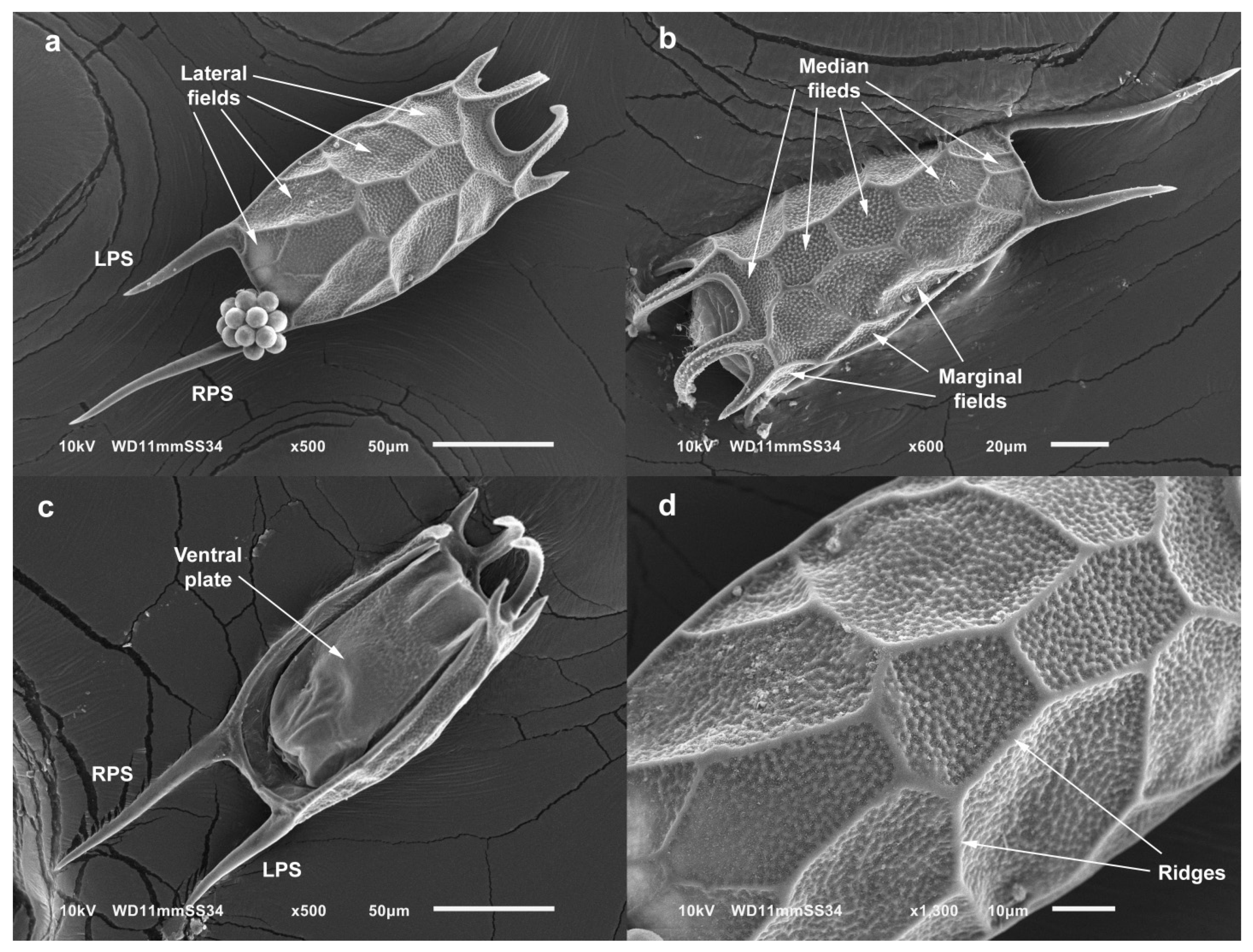
3.3. Taxonomy
4. Discussion
4.1. Morphology
4.2. Distribution and Ecological Comments
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fontaneto, D.; Kaya, M.; Herniou, E.A.; Barraclough, T.G. Extreme levels of hidden diversity in microscopic animals (Rotifera) revealed by DNA taxonomy. Mol. Phylogenet. Evol. 2009, 53, 182–189. [Google Scholar] [CrossRef]
- Mills, S.; Alcántara-Rodríguez, J.A.; Ciros-Pérez, J.; Gómez, A.; Hagiwara, A.; Hinson, K.; Jersabek, C.D.; Malekzadeh-Viayeh, R.; Leasi, F.; Lee, J.S.; et al. Fifteen species in one: Deciphering the Brachionus plicatilis species complex (Rotifera, Monogononta) through DNA taxonomy. Hydrobiologia 2017, 796, 39–58. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.L.; Snell, T.W.; Ricci, C.; Nogrady, T. Rotifera: Biology, Ecology and Systematics. Guides to the Identification of the Microinvertebrates of the Continental Waters of the World, 2nd ed.; Kenobi Productions and Backhuys Publishers: The Hague, The Netherlands, 2006; pp. 1–20. [Google Scholar]
- Brandl, Z. Freshwater copepods and rotifers: Predators and their prey. Hydrobiologia 2005, 546, 475–489. [Google Scholar] [CrossRef]
- Xiang, X.L.; Xi, Y.L.; Zhu, L.Y.; Xu, Q.L. Comparative studies of the population genetic structure of the Brachionus calyciflorus species complex from four inland lakes in Wuhu, China. Biochem. Syst. Ecol. 2017, 71, 69–77. [Google Scholar] [CrossRef]
- García-Morales, A.E.; Elías-Gutiérrez, M. DNA barcoding of freshwater Rotifera in Mexico: Evidence of cryptic speciation in common rotifers. Mol. Ecol. Res. 2013, 13, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- García-Morales, A.E.; Domínguez-Domínguez, O. Cryptic molecular diversity in the morphologically variable rotiferan Brachionus quadridentatus (Rotifera: Monognonta). Rev. Biol. Trop. 2019, 67, 1114–1130. [Google Scholar] [CrossRef]
- Gómez, A.; Serra, M.; Carvalho, G.R.; Lunt, D.H. Speciation in ancient cryptic species complex: Evidence from the molecular phylogeny of Brachionus plicatilis (Rotifera). Evolution 2002, 56, 1431–1444. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-Jiménez, G.; Vannucchi, P.E.; Silva-Briano, M.; Adabache-Ortiz, A.; Rico-Martínez, R.; Roberts, D.; Neilson, R.; Elías-Gutiérrez, M. Brachionus paranguensis sp. nov. (Rotifera, Monogononta), a member of the L group of the Brachionus plicatilis complex. Zookeys 2019, 880, 1–23. [Google Scholar] [CrossRef]
- Schröder, T.; Walsh, E.J. Cryptic speciation in the cosmopolitan Epiphanes senta complex (Monogononta, Rotifera) with the description of new species. Hydrobiologia 2007, 593, 129–140. [Google Scholar] [CrossRef]
- Gilbert, J.J.; Walsh, E.J. Brachionus calyciflorus is a species complex: Mating behavior and genetic differentiation among four geographically isolated strains. Hydrobiologia 2005, 546, 257–265. [Google Scholar] [CrossRef]
- Obertegger, U.; Flaim, G.; Fontaneto, D. Cryptic diversity within the rotifer Polyarthra dolichoptera along an altitudinal gradient. Freshw. Biol. 2014, 59, 2413–2427. [Google Scholar] [CrossRef]
- Cieplinski, A.; Weisse, T.; Obertegger, U. High diversity in Keratella cochlearis (Rotifera, Monogononta): Morphological and genetic evidence. Hydrobiologia 2017, 796, 145–159. [Google Scholar] [CrossRef] [Green Version]
- Derry, A.M.; Hebert, P.D.N.; Prepas, E.E. Evolution of rotifers in saline and subsaline lakes: A molecular phylogenetic approach. Limnol. Oceanogr. 2003, 48, 675–685. [Google Scholar] [CrossRef] [Green Version]
- Kordbacheh, A.; Wallace, R.L.; Walsh, E.J. Evidence supporting cryptic species within two sessile microinvertebrates, Limnias melicerta and L. ceratophylli (Rotifera, Gnesiotrocha). PLoS ONE 2018, 13, e0205203. [Google Scholar] [CrossRef] [PubMed]
- Segers, H. Global diversity of rotifers (Rotifera) in freshwater. Hydrobiologia 2008, 595, 49–59. [Google Scholar] [CrossRef]
- Segers, H.; De Smet, W.H. Diversity and endemism in Rotifera: A review, and Keratella Bory de St Vincent. Biodivers. Conserv. 2008, 17, 303–316. [Google Scholar] [CrossRef]
- Koste, W.; Shiel, R.J. Rotifera from Australian inland waters II Ephiphanidae and Brachionidae (Rotifera: Monogononta). Invertebr. Taxon. 1987, 1, 949–1021. [Google Scholar] [CrossRef]
- Koste, W. Rotatoria. Die Rädertiere Mitteleuropas. Ein Bestimmungswerk, begründet von Max Voigt. Überordnung Monogononta. I. Textband; Gebrüder Borntraeger: Stuttgart, Germany, 1978. [Google Scholar]
- Koste, W. Rotatoria. Die Rädertiere Mitteleuropas. Ein Bestimmungswerk, begründet von Max Voigt. Überordnung Monogononta. II. Tafelband; Gebrüder Borntraeger: Stuttgart, Germany, 1978; pp. 16–25. [Google Scholar]
- Apstein, C. Das plankton im Colombo-See auf Ceylon. Zool. Jahrbücher Abt. Syst. Geogr. Und Biol. Tiere 1907, 25, 201–244. [Google Scholar]
- Segers, H. Annotated checklist of the rotifers (Phylum Rotifera), with notes on nomenclature, taxonomy and distribution. Zootaxa 2007, 1564, 1–104. [Google Scholar] [CrossRef]
- Yermolaeva, N.I.; Kirillov, V.V. First record of Keratella tropica (Apstein, 1907) (Rotifera: Brachionidae) in western Siberia. Russ. J. Biol. Invasions 2018, 9, 38–43. [Google Scholar] [CrossRef]
- Berzins, B. Taxonomie und Verbreitung von Keratella valga und verwandten Formen. Arkiv Zoologi. 1955, 8, 549–559. [Google Scholar]
- Xi, Y.L.; Xu, D.D.; Ma, J.; Ge, Y.L.; Wen, X.L. Differences in life table parameters between Keratella tropica and Keratella valga (Rotatoria) from subtropical shallow lakes. J. Freshw. Ecol. 2013, 28, 539–545. [Google Scholar] [CrossRef] [Green Version]
- Fontaneto, D.; Flot, J.F.; Tang, C.Q. Guidelines from DNA taxonomy with focus on the meiofauna. Mar. Biodivers. 2015, 45, 433–451. [Google Scholar] [CrossRef]
- Walsh, E.J.; Schröder, T.; Wallace, R.L.; Rico-Martinez, R. Cryptic speciation in Lecane bulla (Monogononta: Rotifera) in Chihuahuan Desert waters. Int. Ver. Theor. Angew. Limnol. Verhandlungen 2009, 30, 1046–1050. [Google Scholar]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- Suatoni, E.; Vicario, S.; Rice, S.; Snell, T.; Caccone, A. An analysis of species boundaries and biogeographic patterns in a cryptic species complex: The rotifer Brachionus plicatilis. Mol. Phylogenet. Evol. 2006, 41, 86–98. [Google Scholar] [CrossRef]
- Ferrusquía-Villafranca, I. Ensayo sobre la caracterización y significación biológica. In Biodiversidad de la Faja Volcánica Transmexicana; Luna, I., Morrone, J.J., Espinosa, D., Eds.; Jiménez Editores e Impresores S.A de C.V.: Ciudad de México, México, 2007; pp. 7–23. [Google Scholar]
- Hernández, M.E.; Carrasco, G. Rasgos climáticos más importantes. In Biodiversidad de la Faja Volcánica Transmexicana; Luna, I., Morrone, J.J., Espinosa, D., Eds.; Jiménez Editores e Impresores S.A de C.V.: Ciudad de México, México, 2007; pp. 57–72. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–297. [Google Scholar]
- Palumbi, S.R. The polymerase chain reaction. In Molecular Systematics; Hillis, D.M., Moritz, C., Marble, K., Eds.; Sinauer Associates: Sunderland, MA, USA, 1996; pp. 205–247. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetic analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Asimenos, G.; Toh, H. Multiple alignment of DNA sequences with MAFFT. Methods. Mol. Biol. 2009, 537, 39–64. [Google Scholar] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods. 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. Mrbayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, J.; Barraclough, T.G.; Gómez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A. Sequence based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Heled, J.; Kuehnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.Q.; Obertegger, U.; Fontaneto, D.; Barraclough, T.G. Sexual species are separated by larger genetic gaps than asexual species in rotifers. Evolution 2014, 68, 2901–2916. [Google Scholar] [CrossRef] [Green Version]
- Wilke, T.; Schultheiß, R.; Albrecht, C. As time goes by: A simple fool´s guide to molecular clock approaches in invertebrates. Am. Malacol. Bull. 2009, 27, 25–45. [Google Scholar] [CrossRef]
- DeJong, R.J.; Morgan, J.A.; Lobato, W.; Pointier, J.P.; Amarista, M.; Ayeh-Kumi, P.F.; Babiker, A.; Barbosa, C.S.; Brémond, P.; Canese, A.P.; et al. Evolutionary relationships and biogeography of Biomphalaria (Gastropoda: Planorbidae) with implications regarding its role as host of the human bloodfluke, Schistosoma mansoni. Mol. Biol. Evol. 2001, 18, 2225–2239. [Google Scholar] [CrossRef]
- Schlötterer, C.; Hauser, M.; von Haeseler, A.; Tautz, D. Comparative evolutionary analysis of rDNA ITS regions in Drosophila. Mol. Biol. Evol. 1994, 11, 513–522. [Google Scholar] [PubMed] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Obertegger, U.; Cieplinski, A.; Fontaneto, D.; Papakostas, S. Mitonuclear discordance as a confounding factor in the DNA taxonomy of monogonont rotifers. Zool. Scr. 2018, 47, 122–132. [Google Scholar] [CrossRef]
- Modenutti, B.E.; Diéguez, M.C.; Segers, H. A new Keratella from Patagonia. Hydrobiologia 1998, 389, 1–5. [Google Scholar] [CrossRef]
- Papakostas, S.; Michaloudi, E.; Proios, K.; Brehm, M.; Verhage, L.; Rota, J.; Peña, C.; Stamou, G.; Pritchard, V.L.; Fontaneto, D.; et al. Integrative taxonomy recognizes evolutionary units despite widespread mitonuclear discordance: Evidence from a rotifer cryptic species complex. Syst. Biol. 2016, 65, 508–524. [Google Scholar] [CrossRef] [Green Version]
- Liao, D. Concerted evolution. In Encyclopedia of the Human Genome; Macmillan Publishers Ltd.: New York, NY, USA; Nature Publishing Group: London, UK, 2003; Available online: www.ehgonline.net (accessed on 6 October 2021).
- Elder, J.F.; Turner, B.J. Concerted evolution of repetitive DNA sequences in eukaryotes. Q. Rev. Biol. 1995, 70, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Shi, W.; Yang, M.; Kong, X.Y. Characterization of 18S-ITS1-5.8S rDNA in eleven species in Soleidae: Implications for phylogenetic analysis. Hydrobiologia 2018, 819, 161–175. [Google Scholar] [CrossRef]
- Egborge, A.B.M.; Ogbekene, L. Cyclomorphosis in Keratella tropica (Apstein) of Lake Asejire, Nigeria. Hydrobiologia 1986, 135, 179–191. [Google Scholar] [CrossRef]
- Marinone, M.C.; Zagarese, H.E. A field and laboratory study on factors affecting polymorphism in the rotifer Keratella tropica. Oecologia 1991, 86, 372–377. [Google Scholar] [CrossRef]
- Green, J. Asymmetry and variation in Keratella tropica. Hydrobiologia 1980, 73, 241–248. [Google Scholar] [CrossRef]
- Dieguez, M.C.; Modenutti, B.E. Keratella distribution in North Patagonian lakes (Argentina). Hydrobiologia 1996, 321, 1–6. [Google Scholar] [CrossRef]
- Garza, G.; Silva-Briano, M.; Nandini, S.; Sarma, S.S.S.; Castellanos-Páez, M.E. Morphological and morphometrical variations of selected rotifer species in response to predation: A seasonal study of selected brachionid species from Lake Xochimilco (Mexico). Hydrobiologia 2005, 546, 169–179. [Google Scholar] [CrossRef]
- Tausz, C.; Beaver, J.R.; Renicker, T.R.; Klepach, J.A.; Pollard, A.I.; Mitchell, R.M. Biogeography and co-occurrence of 16 planktonic species of Keratella Bory de St. Vincent, 1822 (Rotifera, Ploima, Brachionidae) in lakes and reservoirs of the United States. Zootaxa 2019, 4624, 337–350. [Google Scholar] [CrossRef]
- Alcocer, J.; Bernal-Brooks, F.W. Limnology in Mexico. Hydrobiologia 2010, 644, 15–68. [Google Scholar] [CrossRef]
- Arriaga, L.; Aguilar, V.; Alcocer, J. Aguas Continentales y Diversidad Biológica de México; Comisión Nacional para el Conocimiento y Uso de la Biodiversidad: Ciudad de México, México, 2000; pp. 153–167. [Google Scholar]
- COMPESCA. Atlas Pesquero y Acuícola de Michoacán; Gobierno del Estado de Michoacán: Morelia, Mexico, 2013; pp. 165–170. [Google Scholar]
- García, M.; Herrera, F. La cuenca hidrosocial presa Huapango, México: Un análisis de la gestión integrada de los recursos hídricos y la gobernanza en cuerpos de agua compartidos. Agua y Territorio 2019, 14, 69–84. [Google Scholar] [CrossRef]
- Sarma, S.S.S.; Nandini, S.; Morales-Ventura, J.; Delgado- Martínez, I.; González-Valverde, L. Effects of NaCl salinity on the population dynamics of freshwater zooplankton (rotifers and cladocerans). Aquatic Ecol. 2006, 40, 349–360. [Google Scholar] [CrossRef]
- Israde, I.; Velázquez-Durán, R.; Lozano, M.S.; Bischoff, J.; Domínguez, G.; Garduño, V.H. Evolución paleolimnológica del lago Cuitzeo, Michoacán durante el Pleistoceno-Holoceno. Bol. Soc. Geol. Mex. 2010, 62, 345–357. [Google Scholar] [CrossRef]
- Chacón, A.; Rosas, C.; Alvarado, J. El lago de Cuitzeo. In Las Aguas Interiores de México: Conceptos y Casos; De la Lanza, G., Hernandez, S., Eds.; AGT Editor S.A: Ciudad de México, México, 2007; pp. 305–338. [Google Scholar]
- Gutiérrez-Aguirre, M.A.; Cervantes-Martínez, A.; Elías-Gutiérrez, M. An example of how barcodes can clarify cryptic species: The case of the calanoid copepod Mastigodiaptomus albuquerquensis (Herrick). PLoS ONE 2014, 9, e85019. [Google Scholar] [CrossRef] [Green Version]
- Campillo, S.; García-Roger, E.M.; Carmona, M.J.; Serra, M. Local adaptation in rotifer populations. Evol. Ecol. 2011, 25, 933–947. [Google Scholar] [CrossRef]
- Ciros-Pérez, J.; Gómez, A.; Serra, M. On the taxonomy of three sympatric sibling species of the Brachionus plicatilis (Rotifera) complex from Spain, with the description of B. ibericus n. sp. J. Plankton. Res. 2001, 23, 1311–1328. [Google Scholar] [CrossRef] [Green Version]
- Montero-Pau, J.; Ramos-Rodríguez, E.; Serra, M.; Gómez, A. Long-term coexistence of rotifer cryptic species. PLoS ONE 2011, 6, e21530. [Google Scholar] [CrossRef] [PubMed]
- Papakostas, S.; Michaloudi, E.; Triantafyllidis, A.; Kappas, I.; Abatzopoulos, T.J. Allochronic divergence and clonal succession: Two microevolutionary processes sculpturing population structure of Brachionus rotifers. Hydrobiologia 2013, 700, 33–45. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TL | LL | LW | RPS | LPS | AMS | AIS | ALS | |
|---|---|---|---|---|---|---|---|---|
| COI-ESU1 | ||||||||
| Mean | 312.9 | 149.6 | 84.8 | 130.4 | 49.3 | 32.8 | 25.9 | 20.4 |
| Median | 314 | 150 | 86 | 131 | 51 | 32 | 25 | 20 |
| Min | 297 | 142 | 77 | 115 | 30 | 25 | 25 | 17 |
| Max | 322 | 157 | 87 | 147 | 57 | 37 | 30 | 25 |
| Population = Ignacio Ramirez (12) | ||||||||
| COI-ESU2 | ||||||||
| Mean | 293.4 | 132.3 | 75.4 | 134.8 | 49.7 | 26.1 | 17.9 | 15.8 |
| Median | 301 | 132 | 75 | 142 | 50 | 26 | 18 | 16 |
| Min | 260 | 124 | 74 | 100 | 42 | 26 | 17 | 15 |
| Max | 314 | 140 | 80 | 152 | 62 | 27 | 18 | 16 |
| Population = Cuitzeo (15) | ||||||||
| COI-ESU3 | ||||||||
| Mean | 201.3 | 113.3 | 65.9 | 56.1 | 45.9 | 31.8 | 19.6 | 17.6 |
| Median | 202 | 114 | 66 | 56 | 45 | 32 | 20 | 18 |
| Min | 194 | 108 | 64 | 52 | 40 | 30 | 19 | 17 |
| Max | 208 | 117 | 72 | 60 | 58 | 32 | 20 | 18 |
| Population = Huapango (15) | ||||||||
| COI-ESU4 | ||||||||
| Mean | 265.6 | 128.4 | 80.6 | 101.4 | 63 | 35.6 | 21.8 | 19.8 |
| Median | 264 | 128 | 80 | 100 | 67 | 36 | 22 | 20 |
| Min | 260 | 126 | 74 | 96 | 48 | 34 | 21 | 19 |
| Max | 274 | 132 | 84 | 106 | 70 | 36 | 22 | 20 |
| Population = Santa Teresa (10) | ||||||||
| COI-ESU5 | ||||||||
| Mean | 257.7 | 109.6 | 74.4 | 104.7 | 58.1 | 43.3 | 22.5 | 20.2 |
| Median | 259 | 107 | 75 | 105 | 57 | 43.5 | 22 | 20 |
| Min | 240 | 105 | 67 | 95 | 52 | 40 | 20 | 17 |
| Max | 275 | 125 | 77 | 122 | 65 | 47 | 25 | 22 |
| Population = Kohunlich (12) | ||||||||
| COI-ESU6 | ||||||||
| Mean | 215.8 | 103.6 | 61.4 | 84.7 | 31.9 | 27.3 | 20.4 | 19.4 |
| Median | 206 | 99 | 63 | 80 | 30 | 26 | 20 | 20 |
| Min | 168 | 90 | 48 | 54 | 8 | 20 | 18 | 18 |
| Max | 264 | 120 | 70 | 118 | 50 | 36 | 24 | 20 |
| Population = Yuriria (10), Tepatitlan (10) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Morales, A.E.; Domínguez-Domínguez, O.; Elías-Gutiérrez, M. Uncovering Hidden Diversity: Three New Species of the Keratella Genus (Rotifera, Monogononta, Brachionidae) of High Altitude Water Systems from Central Mexico. Diversity 2021, 13, 676. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120676
García-Morales AE, Domínguez-Domínguez O, Elías-Gutiérrez M. Uncovering Hidden Diversity: Three New Species of the Keratella Genus (Rotifera, Monogononta, Brachionidae) of High Altitude Water Systems from Central Mexico. Diversity. 2021; 13(12):676. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120676
Chicago/Turabian StyleGarcía-Morales, Alma E., Omar Domínguez-Domínguez, and Manuel Elías-Gutiérrez. 2021. "Uncovering Hidden Diversity: Three New Species of the Keratella Genus (Rotifera, Monogononta, Brachionidae) of High Altitude Water Systems from Central Mexico" Diversity 13, no. 12: 676. https://0-doi-org.brum.beds.ac.uk/10.3390/d13120676