
Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China
by
 , and
, and
Zhanen Yang
1,2 ,
,
Bo Zhang
3,
Zhi Qu
2,
Zikun Song
2,3,
Xiaoyan Pan
2,
Changlin Zhao
1,* and
Haixia Ma
2,4,5,* 1
College of Biodiversity Conservation, Southwest Forestry University, Kunming 650224, China
2
Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China
3
College of Plant Protection, Jilin Agricultural University, Changchun 130000, China
4
Hainan Key Laboratory of Tropical Microbe Resources, Haikou 571101, China
5
Hainan Institute for Tropical Agricultural Resources, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China
*
Authors to whom correspondence should be addressed.
Diversity 2022, 14(2), 149; https://0-doi-org.brum.beds.ac.uk/10.3390/d14020149
Submission received: 22 January 2022
/
Revised: 8 February 2022
/
Accepted: 14 February 2022
/
Published: 18 February 2022
(This article belongs to the Special Issue Diversity and Evolution of Fungi)
Abstract
:Two new species of Diatrype collected in northeast China are described and illustrated based on morphological and molecular evidence. Diatrype larissae from Heilongjiang Province is characterised by having 3–6 perithecia in a stroma, asci polysporous, ascospores allantoid, aseptate, slightly or moderately curved, subhyaline. Diatrype betulaceicola from Inner Mongolia has large stroma with 5–14 perithecia, perithecium immersed, asci polysporous, long-stalked, ascospores allantoid, aseptate, slightly curved, subhyaline. The phylogenies inferred from the data set of nrDNA ITS1-5.8S-ITS2 (ITS) and beta-tubulin (β-tubulin) supported the two new species both as members in the genus Diatrype and distinct species. The morphological similarities and dissimilarities of the new species with phylogenetically close relatives are discussed. A dichotomous identification key to the Diatrype spp. known from China is proposed.
1. Introduction
Diatrypaceae Nitschke, an important family of Xylariales, was introduced by Nitschke (1869) with Diatrype Fr. as the type genus [1,2,3]. Members of Diatrypaceae are world-wide in distribution, and, some species parasitize plants and cause plant diseases, which are plant pathogens [4,5,6,7,8,9,10,11].
The genus Diatrype Fr. was established by Fries (1849) with Diatrype disciformis (Hoffm.) Fr. as the type, which has often been identified as saprobe on rotting wood [3]. The known distribution of Diatrype covers Asia, Europe, North America, Oceania and South Africa [3,5,6,11,12,13,14,15,16,17,18]. Traditionally, the genus is characterised by stromata widely effuse or verrucose, flat or slightly convex, with discoid or sulcate ostioles at the surface, eight-spored and long-stalked asci and hyaline or brownish, allantoid ascospores [3,19,20,21].
The molecular phylogenetic analysis of Diatrypaceae based on ITS including five genera, viz. Cryptosphaeria Ces. and De Not., Diatrype, Diatrypella (Ces. and De Not.) De Not., Eutypa Tul. and C. Tul., and Eutypella (Nitschke) Sacc., were performed, and suggesting a polyphyletic origin for the five genera [4]. More recently, many diatrypaceous taxa were described and illustrated based on morphological characters and multi-gene phylogenetic analyses [8,9,10,11,18,22,23,24]. The taxonomy and molecular phylogeny of Diatrype have not been so well studied in China. A new species of Diatrype subundulata Lar. N. Vassiljeva and H.X. Ma is described, mainly based on morphological data from northeastern China [17]. Recently, two new species of Diatrype were studied based on anamorph and molecular data, and recognised Diatrype quercicola H.Y. Zhu and X.L. Fan as a new species with polysporous asci by combining morphological and phylogenetic analyses [24]. In addition, a new species Diatrype lancangensis S.H. Long and Q.R. Li was introduced from Yunnan province of China by studying morphological characteristics and multi-gene phylogenetic analyses.
During investigations on the diversity of diatrypaceous fungi in northeast China, two undescribed species of Diatrype were identified based on morphological criteria and molecular genetic analyses. The primary purposes of the present paper were to study these specimens by using morphological and multi-gene DNA datasets and discuss the phylogeny of the family Diatrypaceae, based on expanded sampling.
2. Materials and Methods
2.1. Morphological Examination
The specimens studied are deposited at the Fungarium of Institute of Tropical Bioscience and Biotechnology, Chinese Academy of Tropical Agricultural Sciences (FCATAS). Microscopical structures of the sexual morph were measured from fresh material mounted in distilled water, 10% KOH and Melzer’s iodine reagent, respectively [25]. The photographs of stromata were taken using a Canon G16 camera. Each specimen measured over 30 ascus, 30 ascospores, 15 stromata, 15 perithecia. The photographs of stromatal surface were taken with a VHX-600E microscope of the Keyence Corporation. Microscopic observations and measurements were taken by an Olympus IX73 inverted fluorescence microscope at magnifications up to 1000×. The following abbreviations are used in the text: n = the quantity of a morphological feature measured from a specified number of specimens, = the arithmetic mean of the measured values.
2.2. DNA Amplification and Sequencing
A CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd., Beijing, China) was used to obtain genomic DNA from dried specimens, according to the manufacturer’s instructions. The internal transcribed spacer (ITS) region and β-tubulin (TUB2) was amplified with primer pairs ITS5/ITS4 and T1/T22, respectively [26,27]. Polymerase chain reaction (PCR) was carried out in a volume of 40 μL, which contained 16 μL of ddH2O, 20 μL of 2× PCR Master Mix (2× HSTM Mix), 1 μL of DNA template and 1 μL of forward and reverse primers in each reaction. The PCR procedure for ITS was as follows: initial denaturation at 95 °C for 3 min, followed by 35 cycles at 94 °C for 40 s, 58 °C for 45 s and 72 °C for 1 min, and a final extension of 72 °C for 10 min. With regard to TUB2, the amplification reaction followed: 3 min at 95 °C, followed by 35 cycles of 1 min at 94 °C, 1 min at 52° C, and 1.5 min at 72 °C, with a final extension of 72 °C for 10 min. DNA sequencing was performed at BGI tech, Guangzhou, China. All newly generated sequences were deposited at GenBank (http://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/genbank (accessed on 13 January 2022); Table 1).
2.3. Phylogenetic Analyses
Sequencher 4.6 (GeneCodes, Ann Arbor, MI, USA) was used to edit the DNA sequence. Sequences were aligned in MAFFT 7 (http://mafft.cbrc.jp/alignment/server/) (accessed on 6 December 2021) using the “G-INS-i” strategy and manually adjusted in BioEdit [28]. The sequence alignment was deposited in TreeBase (submission ID 29299). Xylaria hypoxylon (L.) Grev. and Kretzschmaria deusta (Hoffm.) P.M.D. Martin obtained from GenBank were used as outgroups.
Sequences were analysed using Maximum Likelihood (ML) through the raxmlGUI 2.0 and Cipres Science Gateway (www.phylo.org) (accessed on 6 December 2021) [29]. Branch support (BS) for ML analysis was determined by 1000 bootstrap replicates.
MrModeltest 2.3 [30] was used to determine the best-fit evolution model for each data set for Bayesian inference (BI). Bayesian inference was calculated with MrBayes 3.1.2 with a general time reversible GTR+I+G model of DNA substitution and a gamma distribution rate variation across sites [31]. Four Markov chains were run for 2 runs from random starting trees for 1.2 million generations, and trees were sampled every 100 generations. The first 1/4 generations were discarded as burn-in. A majority rule consensus tree of all remaining trees was calculated. Branches were considered as significantly supported if they received maximum likelihood bootstrap (BS) > 70%, Bayesian posterior probabilities (BPP) > 0.95.
3. Results
3.1. Molecular Phylogeny
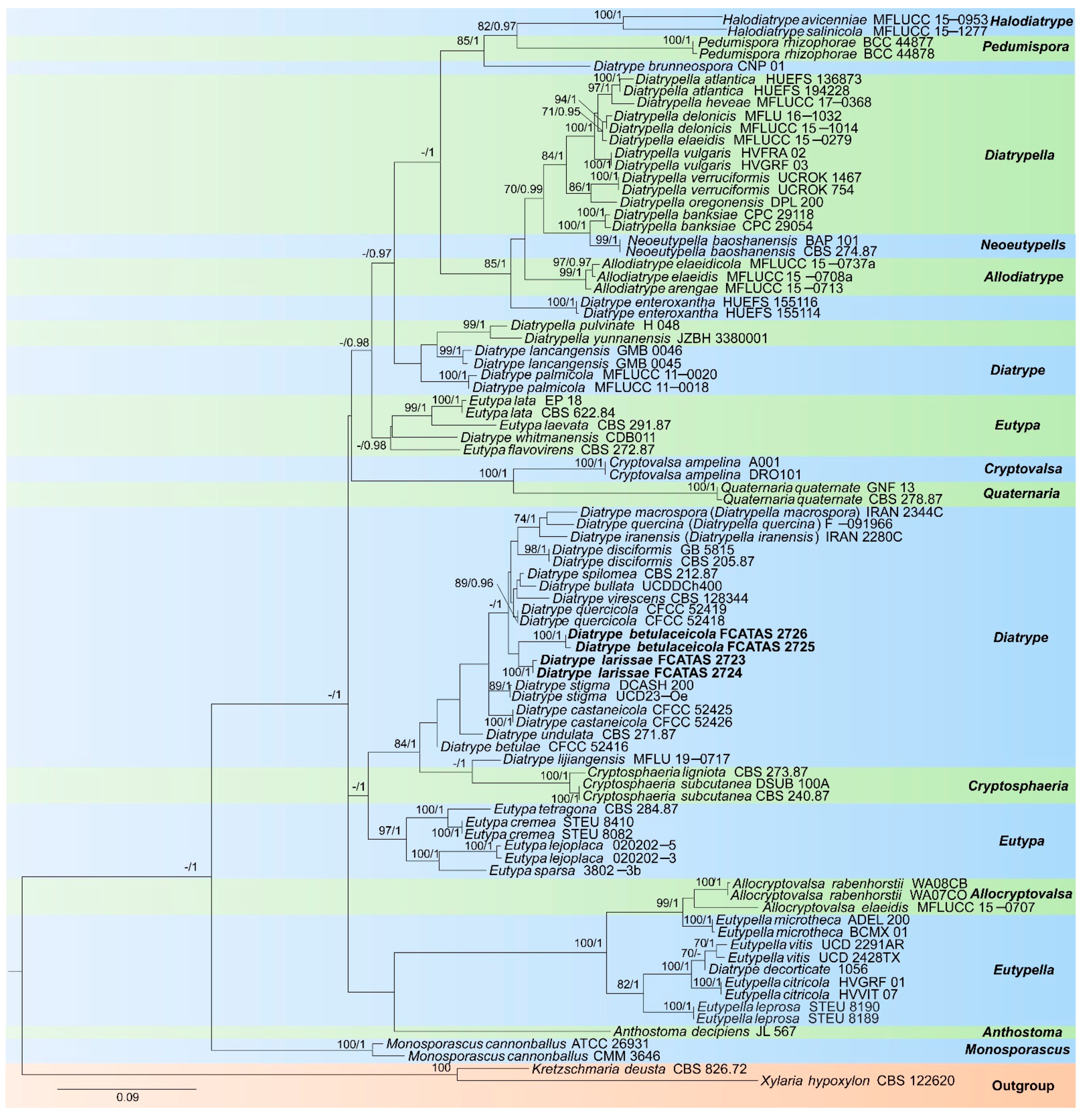
The contribution of the molecular phylogenetic tree based on 151 sequences of two DNA loci (87 ITS and 64 β-tubulin sequences) was composed of 59 strains including two newly described species, two species of Allocryptovalsa, three species of Allodiatrype, one species of Anthostoma, two species of Cryptosphaeria, one species of Cryptovalsa, 20 species of Diatrype, nine species of Diatrypella, seven species of Eutypa, four species of Eutypella, two species of Halodiatrype, one species of Monosporascus, one species of Neoeutypella, one species of Pedumispora, one species of Quaternaria and the X. hypoxylon and K. deusta as outgroups. The dataset had an aligned length of 881 characters, of which 419 characters are constant, 54 are variable and parsimony-uninformative, and 408 are parsimony-informative. Bayesian analysis resulted in a similar topology with an average standard deviation of split frequencies = 0.007642 (BI).
The phylogeny (Figure 1) inferred from ITS and β-tubulin sequences demonstrated that two new taxa were nested in Diatrype and are clearly separated from other sampled species of Diatrype. Diatrype larissae was sister to Diatrype betulaceicola with a weakly supported lineage in the phylogenetic tree.
3.2. Taxonomy
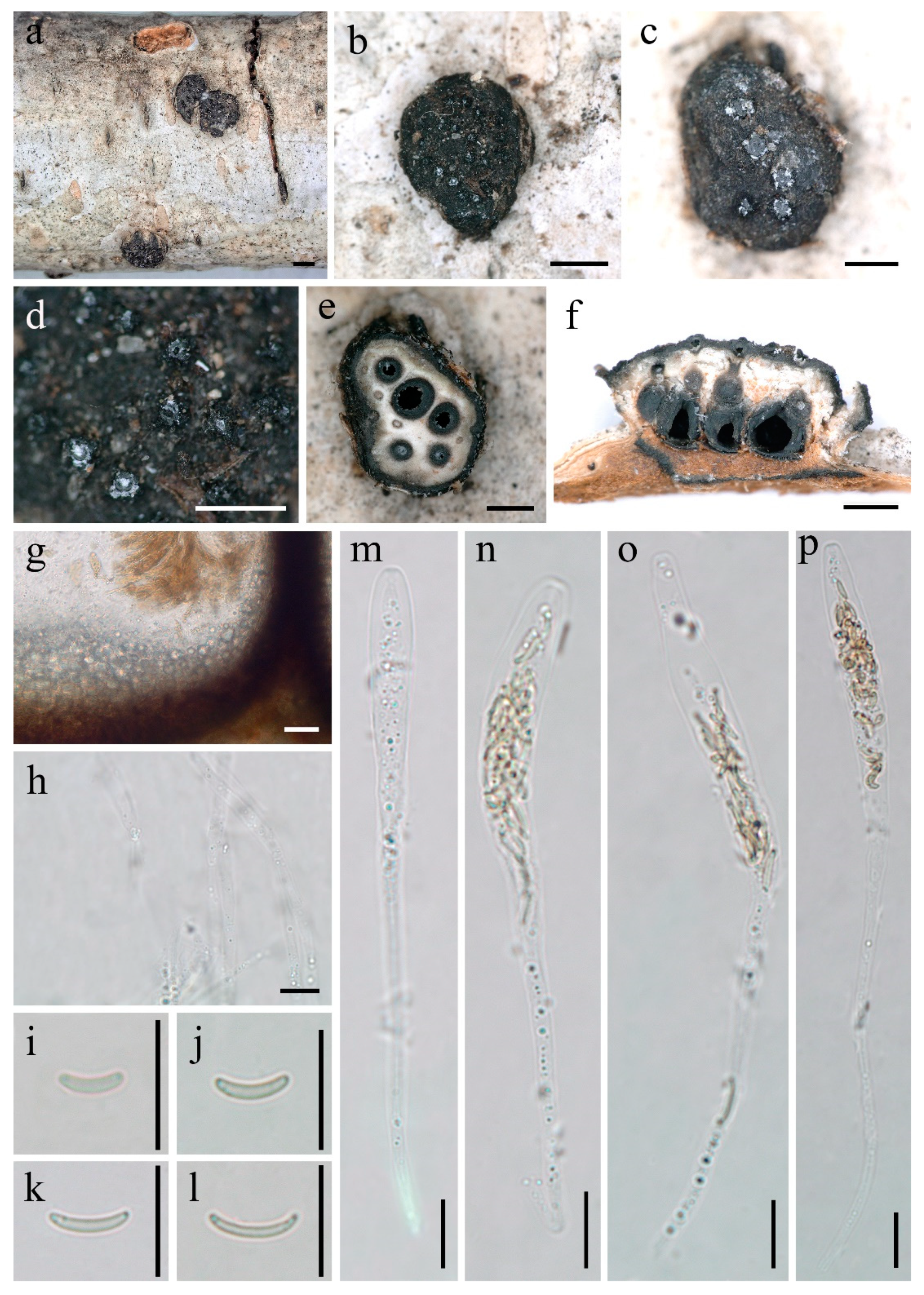
Diatrype betulaceicola Z.E. Yang and Hai X. Ma, sp. nov. Figure 2.
MycoBank: MB 842591.
Holotype—CHINA. Inner Mongolia Autonomous Region, Genhe City, forest maintenance station, saprobic on decaying branches of Betula L., 3 September 2021, Zhang Bo, FCATAS 2725.
Etymology—betulaceicola lat.: Refers to the host family Betulaceae which the fungus inhabits.
Description—Sexual morph: Stromata scattered on the host, erumpent through bark, pustulate, solitary, semi-immersed, 1.9–2.6 mm long × 1.2–2.4 mm broad ( = 2.3 × 1.7 mm, n = 15), ca. 1.1 mm thick, circular to oblong, upper surface nearly flat; surface black; with 5–14 perithecia immersed in per stroma. Endostroma consists of an outer dark brown layer, dense, thin parenchymatous cell layer and an inner white, thick, parenchymatous cell layer, with residual light orange host tissue. Perithecia immersed in stroma, globose to subglobose, 395–588 μm high × 340–516 μm diam ( = 499 × 420 μm, n = 15), individual ostiole with a long neck. Peridium composed of an outer dark brown to black, thin-walled cells, inner layer of hyaline thin-walled cells. Ostiole papillate or apapillate, separately, dark brown to black, 110–191 μm ( = 142 μm, n = 10) in diam. Paraphyses elongate, hyaline, long, filiform, unbranched, septate, guttulate. Asci 88–153 × 7–10 μm ( = 110.4 × 8 μm, n = 30), polysporous, unitunicate, clavate, long-stalked (50–81 μm), apically rounded, with a J-apical ring. Ascospores 4.8–8.6 × 0.9–1.4 μm ( = 6 × 1.2 μm, n = 30), polyseriate, allantoid, aseptate, slightly curved, subhyaline, smooth-walled.
Asexual morph: Undetermined.
Additional specimen examined—CHINA. Inner Mongolia Autonomous Region, Genhe City, forest maintenance station, on fallen branch of Betula L., 3 September 2021, Zhang Bo, FCATAS 2726.
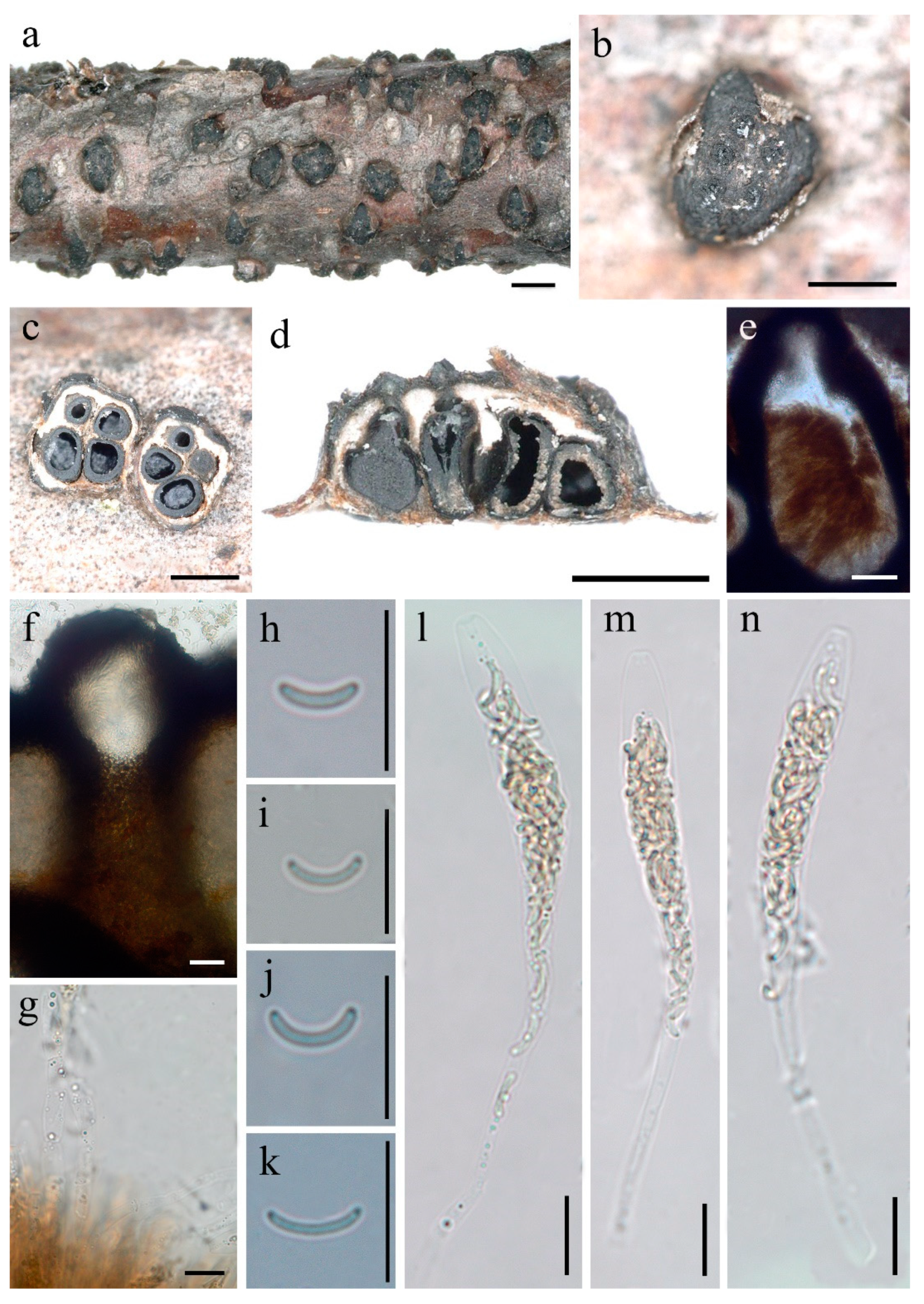
Diatrype larissae Z.E. Yang and Hai X. Ma, sp. nov. Figure 3.
MycoBank: MB 842592.
Holotype—CHINA. Heilongjiang Province, Wudalianchi City, Wudalianchi National Natural Reserve, saprobic on decaying branches of an unidentified plant, 10 September 2021, Zhang Bo, FCATAS 2723.
Etymology—larissae lat.: Refers to honour Russian mycologist Prof. Dr. Larissa N. Vasilyeva, who greatly contributed to studies of pyrenomycetous fungi.
Description—Sexual morph: Stromata scattered on the host, erumpent through bark, pustulate, semi-immersed, solitary, 0.7–1.4 mm long × 0.5–1.1 mm broad, ( = 1.1 × 0.8 mm, n = 30), ca. 0.5 mm thick, circular to oval or rather angular, upper surface nearly flat to slightly convex; surface dark-brown to black, usually smooth, with 3–6 perithecia immersed in per stroma. Endostroma consists of an outer dark brown layer, small, dense, thin parenchymatous cell layer and an inner white, thick, parenchymatous cell layer. Perithecia immersed in stroma, globose to subglobose, 254–401 μm high × 211–362 μm diam ( = 320 × 282 μm, n = 15), individual ostiole with a neck, cylindrical. Peridium composed of outer layer of dark brown, thin-walled cells, inner layer of hyaline thin-walled cells. Ostiole papillate or apapillate, separately, black. Paraphyses elongate, hyaline, filiform, unbranched, septate, guttulate. Asci 83–123 × 7–12 μm ( = 101 × 10 μm, n = 30), polysporous, unitunicate, clavate, long-stalked (26–67 μm), apically truncate, with a J-apical ring. Ascospores 5.6–8.4 × 0.8–1.4 μm ( = 7 × 1.1 μm, n = 30), polyseriate, allantoid, aseptate, slightly or moderately curved, subhyaline, smooth-walled.
Asexual morph: Undetermined.
Additional specimen examined—CHINA. Heilongjiang Province, Wudalianchi City, Wudalianchi National Natural Reserve, on fallen branch of angiosperm, 10 September 2021, Zhang Bo, FCATAS 2724.
4. Discussion
In the present study, two Diatrype species with polysporous asci from China, D. larissae and D. betulaceicola, were described and illustrated as new species based on phylogenetic analyses and morphological characteristics.
Diatrype betulaceicola is characterized by larger stoma with 5–14 perithecia, pustulate, circular to oblong, perithecium immersed, ostiole with a long neck, asci polysporous, ascospores allantoid, aseptate, slightly curved, subhyaline (Figure 2). The new species were found on branch of Betula sp., and, while five Diatrype taxa, D. albopruinosa (Schwein.) Cooke, D. betulae H.Y. Zhu and X.L. Fan, D. oregonensis (Wehm.) Rappaz, D. stigma (Hoffm.) Fr. and D. undulata (Pers.) Fr., were also reported from Betula sp. [6,15,17,24,32]. However, D. betulaceicola can be easily distinguished from the other five taxa by it polysporous asci and phylogenetic analyses. According to our phylogenetic tree based on a combined ITS-TUB2 dataset, and as shown in previous studies [4,11,23,24,33,34,35,36,37], the genus Diatrype as currently circumscribed is of polyphyletic origin within the family Diatrypaceae. Diatrype betulaceicola and D. larissae clustered together with a weakly supported sister branch, which were clearly separated from other sampled species of Diatrype. Morphologically, D. larissae is distinguished by its smaller stroma with 3–6 perithecia, pustulate, circular to oval, rather angular, asci polysporous, ascospores allantoid, slightly or moderately curved, subhyaline (Figure 3). The two new Diatrype are somewhat similar in morphology and size of asci and ascospores, but D. betulaceicola has larger stromata (1.9–2.6 × 1.2–2.4 vs. 0.7–1.4 × 0.5–1.1 mm) and perithecia (395–588 × 340–516 vs. 254–401 × 211–362 μm).
Diatrypequercicola H.Y. Zhu and X.L. Fan also has multisporate asci, but it differs in having larger asci and ascospres (172–183 × 20–43, 17–27 × 4–6 μm) [24], and is distinct from D. betulaceicola and D. larissae phylogenetic trait (Figure 1).
Traditionally, the number of ascospores per ascus (eight-spores vs. more than eight-spores) was morphological criterion for separating the two genera of Diatrype and Diatrypella [19]. Based on phylogenetic analysis of ITS and β-tubulin presented here, we agree with the previous concepts that the polysporous ascus feature cannot be used to distinguish Diatrypella from Diatrype [4,11,33,38,39] (Figure 1). The current phylogenetic analyses show the generic divisions are confusing, and cannot reflect the natural relationships of genera and species within the Diatrypaceae. Therefore, more collections, epitypification, and multi-gene sequences as well as the application of integrative (or polyphasic) taxonomic approach of Diatrypaceae are needed in the future.
| Key to the speices of Diatrype known from China |
| 1. Sexual morph not known ............................................................................................................. 2 |
| 1. Sexual morph present ................................................................................................................... 3 |
| 2. Conidia 10–13 × 1–2 μm ................................................................................................. D. betulae |
| 2. Conidia 4–6 × 1–1.5 μm .......................................................................................... D. castaneicola |
| 3. Stromata solitary, pustulate, circular or elliptic ........................................................................ 4 |
| 3. Stromata widely and indefinitely effuse .................................................................................. 13 |
| 4. Ascus with more than 8 spores ................................................................................................... 5 |
| 4. Ascus with 8 spores ...................................................................................................................... 7 |
| 5. Ascospores 17–27 × 4–6 μm ...................................................................................... D. quercicola |
| 5. Ascospores length less than 10 μm ............................................................................................. 6 |
| 6. Perithecia less than 6 ...................................................................................................... D. larissae |
| 6. Perithecia more than 6 ........................................................................................... D. betulaceicola |
| 7. Ascus length more than 30 μm .................................................................................................... 8 |
| 7. Ascus length less than 30 μm ..................................................................................................... 11 |
| 8. Ascospores 6–8 µm long, size of asci more than 65 μm ....................................... D. lijiangensis |
| 8. Ascospores 8–15 µm long, size of asci less than 65 μm ............................................................ 9 |
| 9. Ascospores brownish ............................................................................................ D. albopruinosa |
| 9. Ascospores subhyaline to light yellow, or light yellow ......................................................... 10 |
| 10. Stromata pulvinate, hemispherical or forming linear stripes .......................... D. oregonensis |
| 10. Stromata circular-polygonal or irregular ......................................................... D. macowaniana |
| 11. Ascospores 4–6 µm long ........................................................................................... D. macounii |
| 11. Ascospores 7–9 µm long ........................................................................................................... 12 |
| 12. Stromata 1–2 × 1–1.5 mm ............................................................................................ D. acericola |
| 12. Stromata 2–7 mm diam ................................................................................................. D. bullata |
| 13. Ascospores 11–19 µm long ................................................................................. D. lancangensis |
| 13. Ascospores 4–9 µm long ........................................................................................................... 14 |
| 14. Ascospores yellowish .......................................................................................... D. subundulata |
| 14. Ascospores hyaline ................................................................................................................... 15 |
| 15. Asci 20–25 μm long, Stromata chocolate-brown ............................................. D. hypoxyloides |
| 15. Asci 25–30 μm long, Stromata different shades of brownish tinges or black .................... 16 |
| 16. Stromata black, perithecia more than 200 µm diam ............................................... D. spilomea |
| 16. Stromata various shades of brownish tinges, perithecia less than 200 µm diam.....D. stigma |
Author Contributions
Conceptualization, C.Z. and H.M.; Data curation, Z.Y.; Formal analysis, Z.Y. and H.M.; Funding acquisition, H.M.; Investigation, Z.Y., B.Z., Z.Q., Z.S. and X.P.; Methodology, Z.Y., C.Z. and H.M.; Project administration, Z.Q. and H.M.; Resources, B.Z. and H.M.; Software, Z.Y.; Supervision, H.M.; Validation, Z.Y. and H.M.; Visualization, Z.Y. and H.M.; Writing—original draft, Z.Y.; Writing—review and editing, Z.Y., C.Z. and H.M. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (no. 31770023, 31972848, U1803232). We are also grateful to the Key Research and Development Program of Hainan (ZDYF2020062) and Hainan Basic and applied research project for cultivating high-level talents (2019RC305).
Institutional Review Board Statement
Not applicable for studies involving humans or animals.
Informed Consent Statement
Not applicable for studies involving humans.
Data Availability Statement
Publicly available datasets were analyzed in this study. This data can be found here: [https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/; https://www.mycobank.org/; http://www.treebase.org, submission ID 29299; accessed on 20 January 2022].
Acknowledgments
We express our gratitude to Zhang Bo (Jilin Agricultural University, Changchun, China) for help during field collections.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nitschke, T.R.J. Grundlage eines Systems der Pyrenomyceten. In Verhandlungen des Naturhistorischen Vereins der Preussischen Rheinlande, Westfalens und des Regierungsbezirks Osnabrück; 1905; Volume 62, pp. 70–77. Available online: https://www.forgottenbooks.com/en/books/VerhandlungendesNaturhistorischenVereinsderPreussischenRheinlandeWestfalensundde_11028671 (accessed on 20 January 2022).
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C. Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Bhat, J.D.; Jones, E.B.G.; McKenzie, E.H.C.; Dai, D.Q.; Daranagama, D.A.; Dayarathne, M.C.; Goonasekara, I.D.; et al. Towards unraveling relationships in Xylariomycetidae (Sordariomycetes). Fungal Divers. 2015, 73, 73–144. [Google Scholar] [CrossRef]
- Acero, F.J.; González, V.; Ballesteros, J.S.; Rubio, V.; Checa, J.; Bills, G.F.; Salazar, O.; Platas, G.; Peláez, F. Molecular phylogenetic studies on the Diatrypaceae based on rDNA-ITS sequences. Mycologia 2004, 96, 249–259. [Google Scholar] [CrossRef]
- Trouillas, F.P.; Sosnowski, M.R.; Gubler, W.D. Two new species of Diatrypaceae from coastal wattle in Coorong National Park, South Australia. Mycosphere 2010, 1, 183–188. [Google Scholar]
- Trouillas, F.P.; Urbez-Torres, J.R.; Gubler, W.D. Diversity of diatrypaceous fungi associated with grapevine canker diseases in California. Mycologia 2010, 102, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Moyo, P.; Damm, U.; Mostert, L.; Halleen, F. Eutypa, Eutypella, and Cryptovalsa Species (Diatrypaceae) associated with Prunus species in South Africa. Plant Disease 2018, 102, 1402–1409. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, M.; Asgari, B.; Hemmati, R. Two new species of Eutypella and a new combination in the genus Peroneutypa (Diatrypaceae). Mycol. Progress 2019, 18, 1057–1069. [Google Scholar] [CrossRef]
- Dayarathne, M.C.; Wanasinghe, D.N.; Devadatha, B.; Abeywickrama, P.; Jones, E.B.G.; Chomnunti, P.; Sarma, V.V.; Hyde, K.D.; Lumyong, S.; McKenzie, E.H.C. Modern taxonomic approaches to identifying diatrypaceous fungi from marine habitats, with a Novel Genus Halocryptovalsa Dayarathne & K.D.Hyde. Gen. Nov. Cryptogam. Mycol. 2020, 41, 21–67. [Google Scholar] [CrossRef]
- Konta, S.; Maharachchikumbura, S.S.N.; Senanayake, I.C.; McKenzie, E.H.C.; Stadler, M.; Boonmee, S.; Phookamsak, R.; Jayawardena, R.S.; Senwanna, C.; Hyde, K.D.; et al. A new genus Allodiatrype, fve new species and a new host record of diatrypaceous fungi from palms (Arecaceae). Mycosphere 2020, 11, 239–268. [Google Scholar] [CrossRef]
- Long, S.H.; Liu, L.L.; Pi, Y.H.; Wu, Y.P.; Lin, Y.; Zhang, X.; Long, Q.D.; Kang, Y.Q.; Kang, J.C.; Wijayawardene, N.N.; et al. New contributions to Diatrypaceae from karst areas in China. Mycokeys 2021, 83, 1–37. [Google Scholar] [CrossRef]
- Doidge, E.M. The South African fungi and lichens to the end of 1945. Bothalia 1950, 5, 1–1094. [Google Scholar]
- Munk, A. Danish pyrenomycetes. A preliminary flora. Dan. Bot. Arkiv 1957, 17, 1–491. [Google Scholar]
- Conners, I.L. An Annotated Index of Plant Diseases in Canada and Fungi Tecorded on Plants in Alaska; Canadian Department of Agriculture: Ottawa, ON, Canada, 1967.
- Rappaz, F. Taxonomie et nomenclature des Diatrypaceae à asques octospores. Mycol. Helv. 1987, 2, 285–648. [Google Scholar]
- Mulenko, W.; Majewski, T.; Ruszkiewicz-Michalska, M. A Preliminary Checklist of Micromycetes in Poland. Kraków: W. Szafer Institute of Botany; Polish Academy of Sciences: Warsaw, Poland, 2008. [Google Scholar]
- Vasilyeva, L.N.; Ma, H.X. Diatrypaceous fungi in north-eastern China. 1. Cryptosphaeria and Diatrype. Phytotaxa 2014, 186, 261–270. [Google Scholar] [CrossRef]
- Peng, M.K.; Zhang, B.; Qu, Z.; Li, Y.; Ma, H.X. New record genus and a new species of Allodiatrype from China based on morphological and molecular characters. Phytotaxa 2021, 500, 275–284. [Google Scholar] [CrossRef]
- Glawe, D.A.; Rogers, J.D. Diatrypaceae in the Pacific Northwest. Mycotaxon 1984, 20, 401–460. [Google Scholar]
- Vasilyeva, L.N.; Stephenson, S.L. Pyrenomycetes of the Great Smoky Mountains National Park. I. Diatrype Fr. (Diatrypaceae). Fungal Divers. 2004, 17, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Vasilyeva, L.N.; Stephenson, S.L. The genus Diatrype (ascomycota, diatrypaceae) in arkansas and texas (USA). Mycotaxon 2009, 107, 307–313. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.J.; Bao, D.F.; Boonmee, S.; Calabon, M.S.; et al. Refined families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Dissanayake, L.S.; Wijayawardene, N.N.; Dayarathne, M.C.; Samarakoon, M.C.; Dai, D.Q.; Hyde, K.D.; Kang, J.C. Paraeutypella guizhouensis gen. et sp. nov. and Diatrypella longiasca sp. nov. (Diatrypaceae) from China. Biodivers. Data J. 2021, 9, e63864. [Google Scholar] [CrossRef]
- Zhu, H.; Pan, M.; Wijayawardene, N.N.; Jiang, N.; Ma, R.; Dai, D.; Tian, C.; Fan, X. The Hidden Diversity of Diatrypaceous Fungi in China. Front. Microbiol. 2021, 12, 646262. [Google Scholar] [CrossRef] [PubMed]
- Kuhnert, E.; Fournier, J.; Per, D.; Luangsaard, J.J.D.; Stadler, M. New Hypoxylon species from Martinique and new evidence on the molecular phylogeny of Hypoxylon based on ITS rDNA and β-tubulin data. Fungal Divers. 2014, 64, 181–203. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium Are Nonorthologous. Mol. Phylogenet. Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop 2010 (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Nylander, J.A.A. MrModeltest v2. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Tiffany, L.H.; Gilman, J.C. Iowa ascomycetes 4: Diatrypaceae. Iowa State J. Sci. 1965, 40, 121–161. [Google Scholar]
- Trouillas, F.P.; Wayne, M.P.; Sosnowski, M.R.; Huang, R.; Peduto, F.; Loschiavo, A.; Savocchia, S.; Scott, E.S.; Gubler, W.D. Taxonomy and DNA phylogeny of Diatrypaceae associated with Vitis vinifera and other woody plants in Australia. Fungal Divers. 2011, 49, 203–223. [Google Scholar] [CrossRef] [Green Version]
- Mehrabi, M.; Hemmati, R.; Vasilyeva, L.N.; Trouillas, F.P. A new species and a new record of Diatrypaceae from Iran. Mycosphere 2015, 6, 60–68. [Google Scholar] [CrossRef]
- Mehrabi, M.; Hemmati, R.; Vasilyeva, L.N.; Trouillas, F.P. Diatrypella macrospora sp. nov. and new records of diatrypaceous fungi from Iran. Phytotaxa 2016, 252, 43–55. [Google Scholar] [CrossRef]
- De Almeida, D.A.C.; Gusmão, L.F.P.; Miller, A.N. Taxonomy and molecular phylogeny of Diatrypaceae (Ascomycota, Xylariales) species from the Brazilian semi-arid region, including four new species. Mycol. Prog. 2016, 15, 1–27. [Google Scholar] [CrossRef]
- Shang, Q.J.; Hyde, K.D.; Phookamsak, R.; Doilom, M.; Bhat, D.J.; Maharachchikumbura, S.S.; Promputtha, I. Diatrypella tectonae and Peroneutypa mackenziei spp. nov. (Diatrypaceae) from northern Thailand. Mycol. Prog. 2017, 16, 463–476. [Google Scholar] [CrossRef]
- Vasilyeva, L.N.; Stephenson, S.L. Pyrenomycetes of the Great Smoky Mountains National Park. II. Cryptovalsa Ces. et De Not. And Diatrypaceae (Ces. et De Not.) Nitschke (Diatrypaceae). Fungal Divers. 2005, 19, 189–200. [Google Scholar]
- Liu, J.K.; Hyde, K.D.; Gareth, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.; Mckenzie, E.H.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
Figure 1.
Phylogram generated from Maximum Likelihood (ML) analyses, based on ITS-β-tubulin matrix. Branches are labelled with maximum likelihood bootstrap ≥ 70%, Bayesian posterior probabilities ≥ 0.95. New species are labelled in bold.
Figure 1.
Phylogram generated from Maximum Likelihood (ML) analyses, based on ITS-β-tubulin matrix. Branches are labelled with maximum likelihood bootstrap ≥ 70%, Bayesian posterior probabilities ≥ 0.95. New species are labelled in bold.

Figure 2.
Diatrype betulaceicola (FCATAS 2725, Holotype). (a–d) Stromata on substrate; (e) cross section of a stroma; (f,g) vertical section through stroma showing ostiole and perithecia; (h) paraphyses in water; (i–l) ascospore in water; (m–p) ascus in water; scale bars ((a,b) = 1 mm; (c–f) = 500 µm; (g–p) = 10 µm).
Figure 2.
Diatrype betulaceicola (FCATAS 2725, Holotype). (a–d) Stromata on substrate; (e) cross section of a stroma; (f,g) vertical section through stroma showing ostiole and perithecia; (h) paraphyses in water; (i–l) ascospore in water; (m–p) ascus in water; scale bars ((a,b) = 1 mm; (c–f) = 500 µm; (g–p) = 10 µm).

Figure 3.
Diatrype larissae (FCATAS 2723, Holotype). (a,b) Stromata on substrate; (c) cross section of a stroma; (d,e) vertical section through stroma showing ostioles and perithecia; (f) ostiolar canal; (g) paraphyses in water; (h–k) ascospore in water; (l–n) ascus in water; scale bars ((a) = 1 mm; (b–d) = 500 µm; (e) = 20 µm; (f–n) = 10 µm).
Figure 3.
Diatrype larissae (FCATAS 2723, Holotype). (a,b) Stromata on substrate; (c) cross section of a stroma; (d,e) vertical section through stroma showing ostioles and perithecia; (f) ostiolar canal; (g) paraphyses in water; (h–k) ascospore in water; (l–n) ascus in water; scale bars ((a) = 1 mm; (b–d) = 500 µm; (e) = 20 µm; (f–n) = 10 µm).


{kind=link}
{kind=link}
{kind=link}
Table 1.
List of species, specimens, and GenBank accession numbers of sequences used in this study.
| Species | Strain | Host/Substrate | Origin | GenBank Accession Numbers | |
|---|---|---|---|---|---|
| ITS | TUB2 | ||||
| Allocryptovalsa elaeidis | MFLUCC 15-0707 | Elaeis guineensis | Thailand | MN308410 | MN340296 |
| A. rabenhorstii | WA07CO | Vitis vinifera | Australia | HQ692620 | HQ692522 |
| A. rabenhorstii | WA08CB | Vitis vinifera | Australia | HQ692619 | HQ692523 |
| Allodiatrype arengaeTS | MFLUCC 15-0713 | Arenga pinnata | Thailand | MN308411 | MN340297 |
| A. elaeidicola | MFLUCC 15-0737a | Elaeis guineensis | Thailand | MN308415 | MN340299 |
| A. elaeidis | MFLUCC 15-0708a | Elaeis guineensis | Thailand | MN308412 | MN340298 |
| Anthostoma decipiensTS | JL567 | Vitis vinifera | Spain | JN975370 | JN975407 |
| Cryptosphaeria ligniota | CBS 273.87 | Populus tremula | Switzerland | KT425233 | KT425168 |
| C. subcutanea | CBS 240.87 | NA | Norway | KT425232 | KT425167 |
| C. subcutanea | DSUB100A | NA | Norway | KT425189 | KT425124 |
| Cryptovalsa ampelina | A001 | NA | Australia | GQ293901 | GQ293972 |
| C. ampelina | DRO101 | NA | America | GQ293902 | GQ293982 |
| Diatrype betulae | CFCC 52416 | Betula davurica | China | MW632943 | NA |
| D. betulaceicola TS | FCATAS 2725 | Betula sp. | China | OM040386 | OM240966 |
| D. betulaceicola TS | FCATAS 2726 | Betula sp. | China | OM040387 | OM240967 |
| D. decorticata | 1056 | NA | NA | KU320621 | NA |
| D. brunneospora | CNP01 | Acacia longifolia subsp. sophorae | Australia | HM581946 | HQ692478 |
| D. bullata | UCDDCh400 | NA | America | DQ006946 | DQ007002 |
| D. castaneicola | CFCC 52425 | Castanea mollissima | China | MW632941 | NA |
| D. castaneicola | CFCC 52426 | Castanea mollissima | China | MW632942 | NA |
| D. disciformisTS | CBS 205.87 | Fagus sylvatica | Switzerland | AJ302437 | NA |
| D. disciformisTS | GB 5815 | Fagus grandifolia | America | KR605644.1 | KY352434.1 |
| D. enteroxantha | HUEFS155114 | NA | Brazil | KM396617 | KT003700 |
| D. enteroxantha | HUEFS155116 | NA | Brazil | KM396618 | KT022236 |
| D. iranensis (Diatrypellairanensis) | IRAN 2280C | Quercus brantii | Iran | KM245033 | NA |
| D. lancangensis | GMB0045 | NA | China | MW797113 | MW814885 |
| D. lancangensis | GMB0046 | NA | China | MW797114 | MW814886 |
| D. larissae TS | FCATAS 2723 | NA | China | OM040384 | OM240964 |
| D. larissae TS | FCATAS 2724 | NA | China | OM040385 | OM240965 |
| D. lijiangensis | MFLU 19-0717 | NA | China | MK852582 | MK852583 |
| D. macrospora (Diatrypellamacrospora) | IRAN 2344C | Quercus brantii | Iran | KR605648 | NA |
| D. oregonensis | DPL200 | Quercus kelloggii | America | GQ293940 | GQ293999 |
| D. palmicola | MFLUCC 11-0018 | Caryota urens | Thailand | KP744438 | NA |
| D. palmicola | MFLUCC 11-0020 | Caryota urens | Thailand | KP744439 | NA |
| D. quercicola | CFCC 52418 | Quercus mongolica | China | MW632938 | NA |
| D. quercicola | CFCC 52419 | Quercus mongolica | China | MW632939 | NA |
| D. quercina (Diatrypellaquercina) | F-091966 | Quercus faginea | Spain | AJ302444 | NA |
| D. spilomea | CBS 212.87 | Acer campestre | Switzerland | AJ302433 | NA |
| D. stigma | DCASH200 | Quercus sp. | America | GQ293947 | GQ294003 |
| D. stigma | UCD23-Oe | NA | NA | JX515704 | JX515670 |
| D. undulata | CBS 271.87 | Betula sp. | Switzerland | AJ302436 | NA |
| D. virescens | CBS 128344 | NA | NA | MH864890 | NA |
| D. whitmanensis | CDB011 | Vitis vinifera | America | GQ293954 | GQ294010 |
| Diatrypella atlantica | HUEFS 136873 | unidentified plant | Brazil | KM396614 | KR259647 |
| D. atlantica | HUEFS 194228 | unidentified plant | Brazil | KM396615 | KR363998 |
| D. banksiae | CPC 29054 | Banksia coccinia | Australia | KY173401 | NA |
| D. banksiae | CPC 29118 | Banksia formosa | Australia | KY173402 | NA |
| D. delonicis | MFLU 16-1032 | Delonix regia | Thailand | MH812995 | MH847791 |
| D. delonicis | MFLUCC 15-1014 | Delonix regia | Thailand | MH812994 | MH847790 |
| D. elaeidis | MFLUCC 15-0279 | Elaeis guineensis | Thailand | MN308417 | MN340300 |
| D. heveae | MFLUCC 17-0368 | Hevea brasiliensis | Thailand | MF959501 | MG334557 |
| D. pulvinata | H048 | Salix alba | Czech Republic | FR715523 | FR715495 |
| D. verruciformisTSQ | UCROK1467 | Quercus agrifolia | America | JX144793 | JX174093 |
| D. verruciformisTSQ | UCROK754 | Quercus agrifolia | America | JX144783 | JX174083 |
| D. vulgaris | HVFRA02 | Fraxinus angustifolia | Australia | HQ692591 | HQ692503 |
| D. vulgaris | HVGRF03 | Citrus paradisi | Australia | HQ692590 | HQ692502 |
| D. yunnanensis | JZBH3380001 | unidentified plant | China | MN653008 | MN887112 |
| Eutypa cremea | STEU 8082 | Vitis vinifera | South Africa | KY111656 | KY111598 |
| E. cremea | STEU 8410 | Prunus armeniaca | South Africa | KY752765 | KY752789 |
| E. flavovirens | CBS 272.87 | Quercus ilex | France | AJ302457 | DQ006959 |
| E. laevata | CBS 291.87 | Salix sp. | Switzerland | HM164737 | HM164771 |
| E. lataTS | EP18 | Vitis vinifera | NSW, Australia | HQ692611 | HQ692501 |
| E. lataTS (Eutypa armeniacae) | CBS 622.84 | Vitis vinifera | Italy | AJ302446 | DQ006964 |
| E. lejoplaca | 020202-3 | Acer pseudoplatanus | Switzerland | AY684238 | AY684197 |
| E. lejoplaca | 020202-5 | Acer pseudoplatanus | Switzerland | AY684221 | AY684196 |
| E. sparsa | 3802-3b | Populus sp. | Switzerland | AY684220 | AY684201 |
| E. tetragona | CBS 284.87 | Sarothamnus scoparius | France | DQ006923 | DQ006960 |
| Eutypella citricola | HVGRF01 | Citrus sinensis | Australia | HQ692589 | HQ692521 |
| E. citricola | HVVIT07 | Vitis vinifera | Australia | HQ692579 | HQ692512 |
| E. leprosa | STEU 8189 | NA | South Africa | MF359637 | MF359672 |
| E. leprosa | STEU 8190 | NA | South Africa | MF359638 | MF359673 |
| E. microtheca | ADEL200 | Ulmus procera | Australia | HQ692559 | HQ692527 |
| E. microtheca | BCMX01 | Vitis vinifera | Mexico | KC405563 | KC405560 |
| E. vitis | UCD2291AR | Vitis vinifera | America | HQ288224 | HQ288303 |
| E. vitis | UCD2428TX | Vitis vinifera | TX, America | FJ790851 | GU294726 |
| Halodiatrype avicenniae | MFLUCC 15-0953 | Avicennia sp. | Thailand | KX573916 | KX573931 |
| H. salinicolaTS | MFLUCC 15-1277 | submerged marine wood | Thailand | KX573915 | KX573932 |
| Kretzschmaria deusta | CBS 826.72 | NA | NA | KU683767 | KU684190 |
| Monosporascus cannonballusTS | ATCC 26931 | NA | NA | FJ430598 | NA |
| M. cannonballusTS | CMM 3646 | Boerhavia sp. | Brazil | JX971617 | NA |
| Neoeutypella baoshanensisTS | BAP101 | Pinus armandii | China | MH822887 | MH822888 |
| N. baoshanensisTS | CBS 274.87 | Ficus carica | France | AJ302460 | NA |
| Pedumispora rhizophoraeTS | BCC44877 | Rhizophora apiculata | Thailand | KJ888853 | NA |
| P. rhizophoraeTS | BCC44878 | Rhizophora apiculata | Thailand | KJ888854 | NA |
| Quaternaria quaternata | GNF13 | Fagus sp. | Iran | KR605645 | KY352464 |
| Q. quaternata | CBS 278.87 | Fagus sulvatica | Switzerland | AJ302469 | NA |
| Xylaria hypoxylon | CBS 122620 | NA | Sweden | AM993141 | KX271279 |
NA: not applicable. Type species are denoted with the superscript “TS” and the disputable type species are denoted with the superscript “TSQ”.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Yang, Z.; Zhang, B.; Qu, Z.; Song, Z.; Pan, X.; Zhao, C.; Ma, H. Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China. Diversity 2022, 14, 149. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020149
AMA Style
Yang Z, Zhang B, Qu Z, Song Z, Pan X, Zhao C, Ma H. Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China. Diversity. 2022; 14(2):149. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020149
Chicago/Turabian StyleYang, Zhanen, Bo Zhang, Zhi Qu, Zikun Song, Xiaoyan Pan, Changlin Zhao, and Haixia Ma. 2022. "Two New Species of Diatrype (Xylariales, Ascomycota) with Polysporous Asci from China" Diversity 14, no. 2: 149. https://0-doi-org.brum.beds.ac.uk/10.3390/d14020149
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.