
Structure and Composition of Rhodolith Beds from the Sergipe-Alagoas Basin (NE Brazil, Southwestern Atlantic)

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
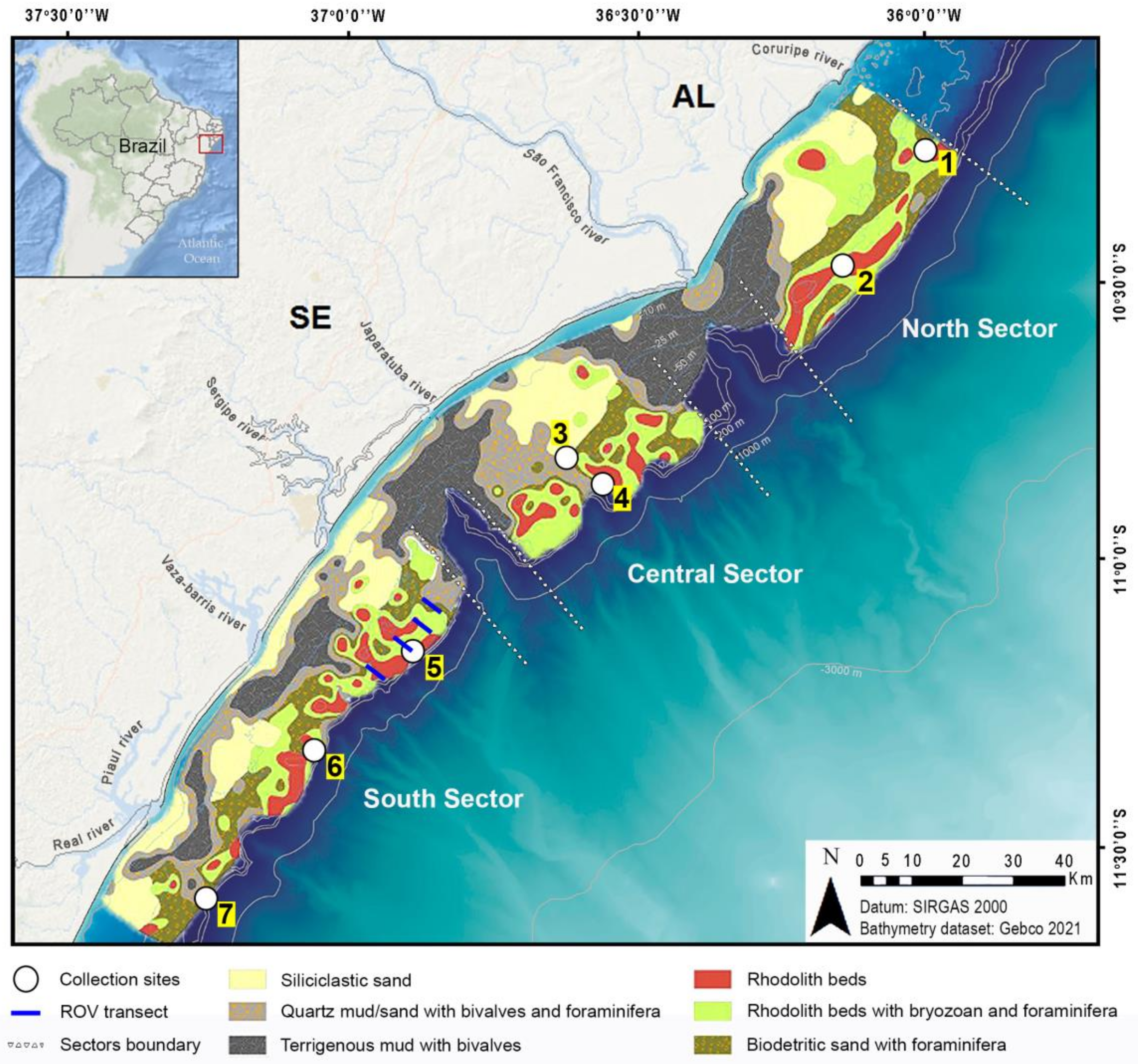
2. Study Area
3. Materials and Methods
3.1. Sampling
3.2. Sample Characterization
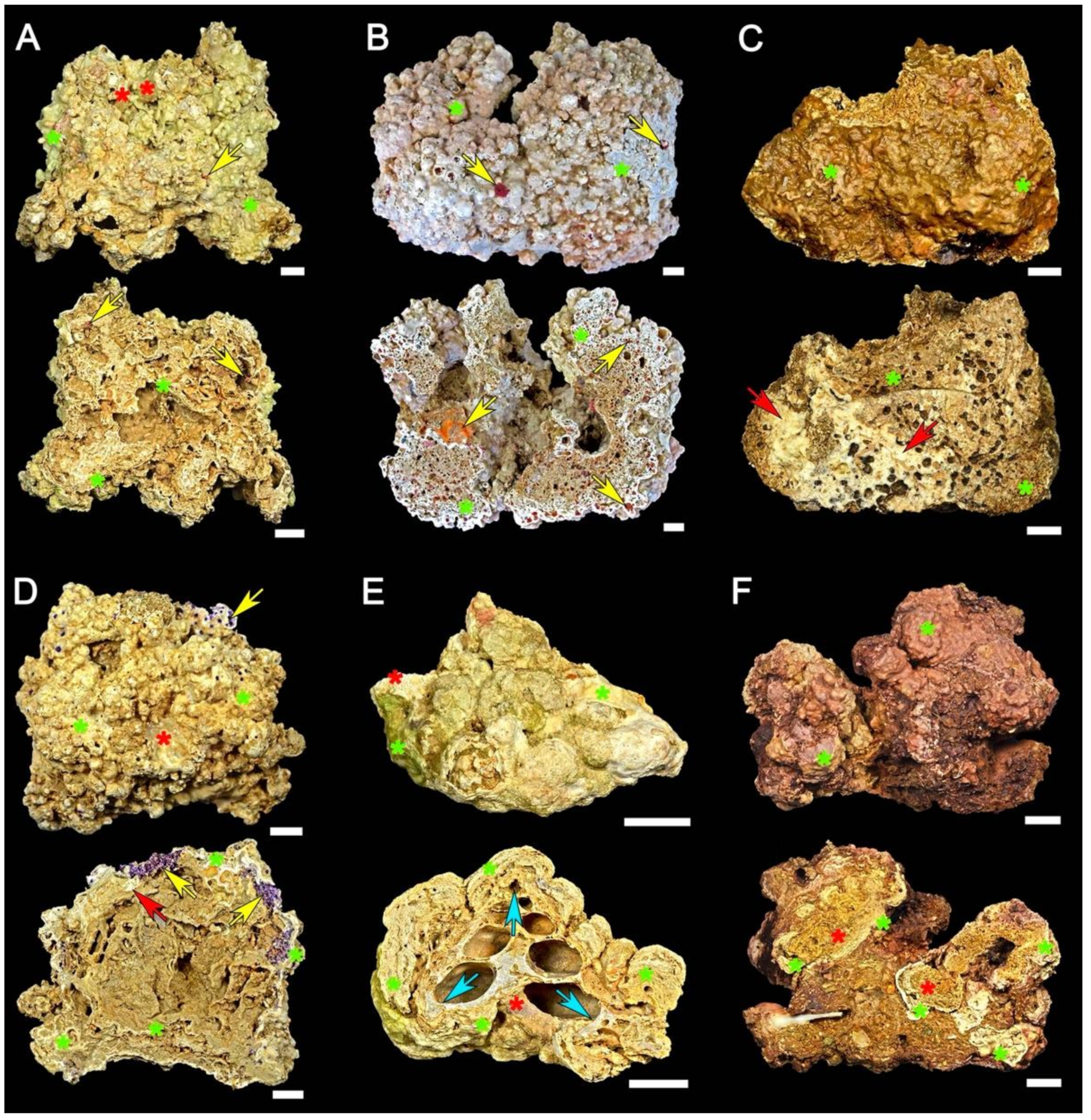
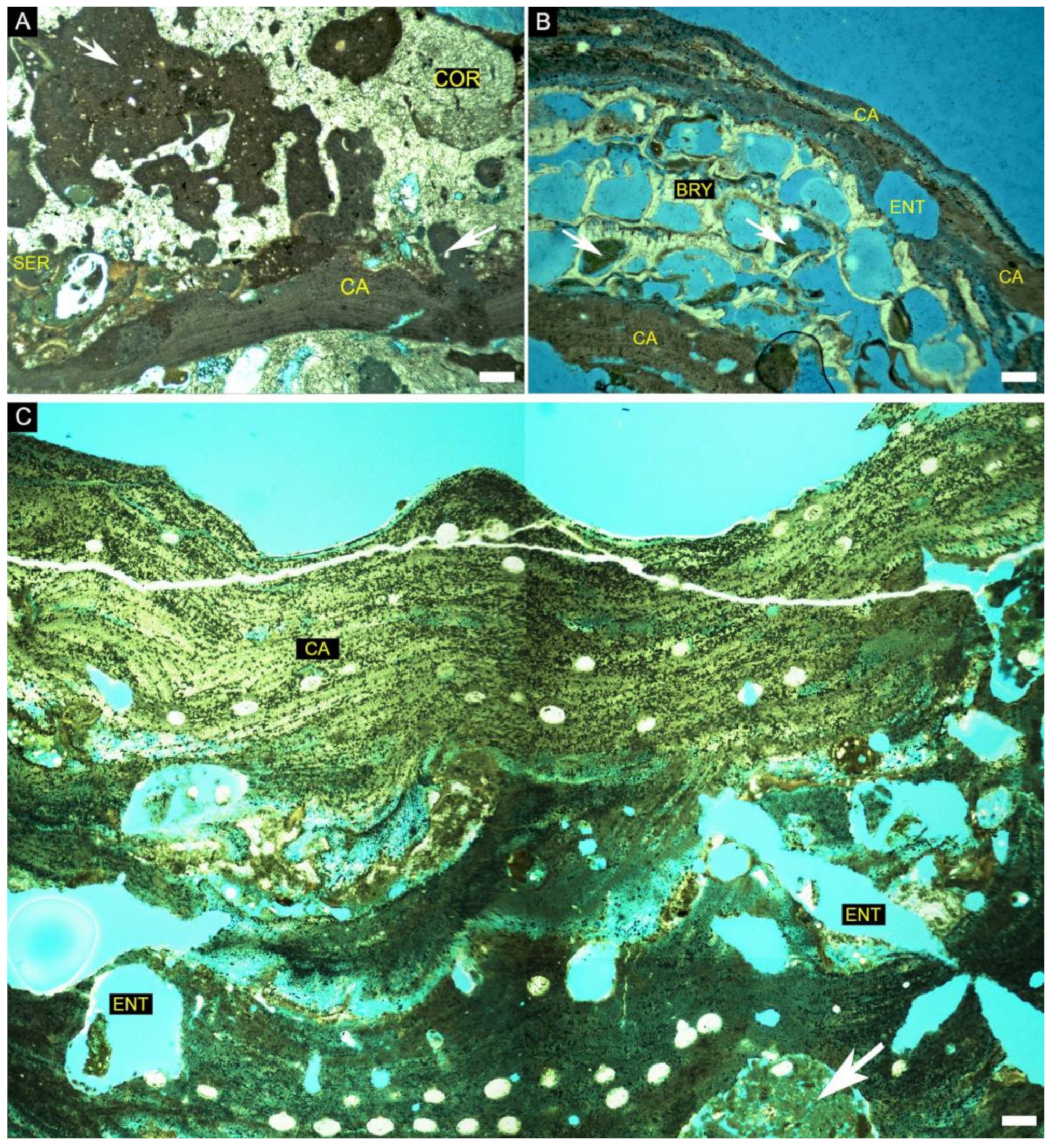
4. Results
4.1. Seabed Characterization in the Examined ROV Transect Lines
4.2. North Sector
4.3. Central Sector
4.4. South Sector
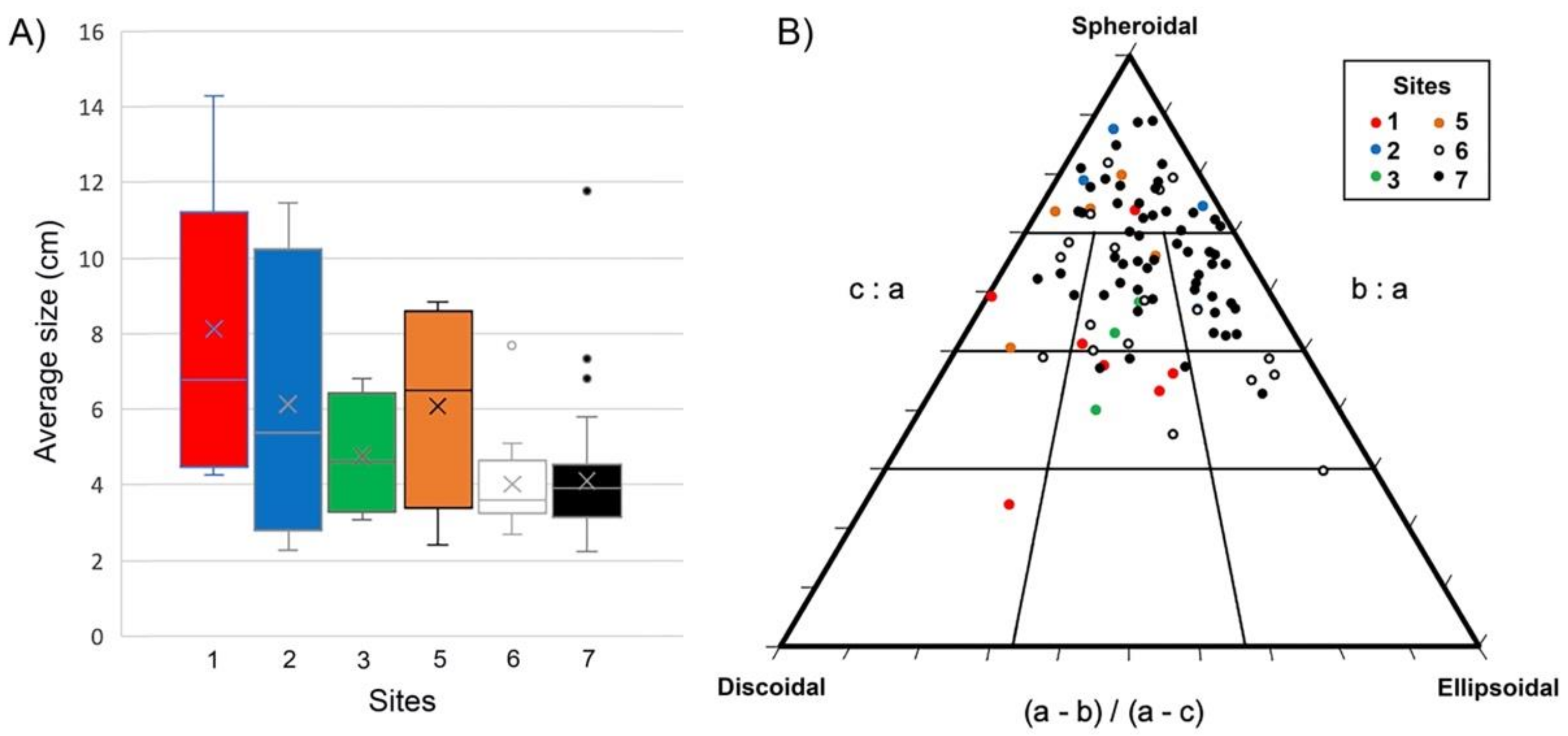
4.5. Comparison of Rhodolith Distribution and Characteristics among Sectors
5. Discussion
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aguirre, J.; Braga, J.C.; Bassi, D. Rhodoliths and rhodolith beds in the rock record. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Geerbestrasse, Switzerland, 2017; Volume 15, pp. 105–138. [Google Scholar]
- Basso, D.; Babbini, L.; Kaleb, S.; Bracchi, V.; Falace, A. Monitoring deep Mediterranean rhodolith beds. Aquat. Conserv. 2016, 26, 549–561. [Google Scholar] [CrossRef] [Green Version]
- Foster, M.S. Rhodoliths: Between rocks and soft places—Minireview. J. Phycol. 2001, 37, 659–667. [Google Scholar] [CrossRef]
- Kamenos, N.A.; Heidi, L.B.; Darrenougue, N. Coralline algae as recorders of past climatic and environmental conditions. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Geerbestrasse, Switzerland, 2017; Volume 15, pp. 27–53. [Google Scholar]
- Foster, M.S.; Amado-Filho, G.M.; Kamenos, N.A.; Riosmena-Rodriguez, R.; Steller, D.S. Rhodolith and rhodolith beds. In Contribution of SCUBA Diving to Research and Discovery in Marine Environments; Lang, M.O., Ed.; Smithsonian Institution Scholarly Press: Washington, DC, USA, 2013; pp. 143–156. [Google Scholar]
- Harvey, A.S.; Woelkerling, W.J. A guide to non-geniculate coralline red algal (Corallinales, Rhodophyta) rhodolith identification. Cienc. Mar. 2007, 33, 411–426. [Google Scholar] [CrossRef] [Green Version]
- Hily, C.; Potin, P.; Floch, J.Y. Structure of subtidal algal assemblages on soft bottom sediments: Fauna/flora interactions and role of disturbances in the Bay of Brest, France. Mar. Ecol. Prog. Ser. 1992, 85, 115–130. [Google Scholar] [CrossRef]
- Birkett, D.A.; Maggs, C.; Dring, M.J. Maërl: An overview of dynamic and sensitivity characteristics for conservation management of marine SACs. Scott. Assoc. Mar. Sci. 1998, 5, 1–90. [Google Scholar]
- Nelson, W.A.; Neill, K.; Farr, T.; Barr, N.; D’Archino, R.; Miller, S.; Stewart, R. Rhodolith Beds in Northern New Zealand: Characterization of associated biodiversity and vulnerability to environmental stressors. N. Z. Aquat. Environ. Biodivers. Rep. 2009, 99, 102. [Google Scholar]
- Harvey, A.S.; Bird, F.L. Community structure of a rhodolith bed from cold temperate waters (southern Australia). Aust. J. Bot. 2008, 56, 437–450. [Google Scholar] [CrossRef]
- Konar, B.; Riosmena-Rodriguez, R.; Iken, K. Rhodolith bed: A newly discovered habitat in the North Pacific Ocean. Bot. Mar. 2006, 49, 355–359. [Google Scholar] [CrossRef]
- Steller, D.L.; Riosmena-Rodriguez, R.; Foster, M.S.; Roberts, C. Rhodolith bed diversity in the Gulf of California: The importance of rhodolith structure and consequences of anthropogenic disturbances. Aquat. Conserv. Mar. Freshw. Ecosyst. 2003, 13, S5–S20. [Google Scholar] [CrossRef]
- Hinojosa-Arango, G.; Riosmena-Rodriguez, R. Influence of rhodolith-forming species and growth-form on associated fauna of rhodolith beds in the central-west Gulf of California, México. Mar. Ecol. 2004, 25, 109–127. [Google Scholar] [CrossRef]
- Sañé, E.; Chiocci, F.L.; Basso, D.; Martorelli, E. Environmental factors controlling the distribution of rhodoliths: An integrated study based on seafloor sampling, ROV and side scan sonar data, offshore the W-Pontine Archipelago. Cont. Shelf Res. 2016, 129, 10–22. [Google Scholar] [CrossRef]
- Concentino, A.L.M.; Vieira, I.B.; Reis, T.N.V.; Vasconcelos, E.R.T.P.P.; Paes, E.T. Fitobentos da Plataforma Continental de Sergipe e de Alagoas. In Plataforma Continental de Sergipe e Alagoas: Geoquímica Sedimentar e Comunidade Bêntica; de Lara Palmeira de Macedo Arguelho, M.E.R.C.e.M., Ed.; Editora UFS; Coleção Projeto MARSEAL: São Cristóvão, Brazil, 2018; Volume 2, pp. 196–220. [Google Scholar]
- Figueiredo, M.A.O.; Eide, I.; Reynier, M.; Villas-Bôas, A.B.; Tâmega, F.T.S.; Ferreira, C.G.; Nilssen, I.; Coutinho, R.; Johnsen, S. The effect of sediment mimicking drill cuttings on deep-water rhodoliths in a flow-through system: Experimental work and modeling. Mar. Pollut. Bull. 2015, 95, 81–88. [Google Scholar] [CrossRef]
- Amado-Filho, G.M.; Moura, R.L.; Bastos, A.C.; Salgado, L.T.; Sumida, P.Y.G.; Guth, A.Z.; Francini-Filho, R.B.; Pereira-Filho, G.H.; Abrantes, D.P.; Brasileiro, P.S.; et al. Rhodolith beds are major CaCO3 bio-factories in the tropical South West Atlantic. PLoS ONE 2012, 7, e35171. [Google Scholar] [CrossRef] [Green Version]
- Amado-Filho, G.M.; Bahia, R.G.; Pereira-Filho, G.H.; Longo, L.L. South Atlantic rhodolith beds: Latitudinal distribution, species composition, structure and ecosystem functions, threats and conservation status. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Geerbestrasse, Switzerland, 2017; Volume 15, pp. 299–317. [Google Scholar]
- Guimarães, C.R.P. Composição e distribuição dos sedimentos superficiais e da fauna bêntica na plataforma continental de Sergipe. Ph.D. Thesis, Instituto de Geociências, Universidade Federal da Bahia, Salvador-Bahia, Brazil, 2010. [Google Scholar]
- Nascimento, A.A. Sedimentação holocênica na plataforma continental de Sergipe, nordeste do Brasil. Master’s Thesis, Instituto de Geociências, Universidade Federal da Bahia, Salvador–Bahia, Brazil, 2011. [Google Scholar]
- Fontes, L.C.S.; Santos, J.R.; Santos, L.A.; Mendonça, J.B.S.; Santos, M.S. Sedimentos superficiais da plataforma continental de Sergipe-Alagoas. In Geologia e Geomorfologia da Bacia de Sergipe-Alagoas; Carneiro, M.E.R., Ed.; Editora UFS; Coleção Projeto MARSEAL: São Cristóvão, Brazil, 2017; Volume 1, pp. 64–96. [Google Scholar]
- Santos, J.R.; Souza, R.M.; Andrade, E.; Fontes, L.C.S. Biogenic components as environmental indicators of the Continental Platform of the state Sergipe and south of Alagoas. Geociências 2019, 38, 409–425. [Google Scholar] [CrossRef]
- Santos, R.S.; Santos, L.A.; Fontes, L.C.S. Geomorphological and sedimentary mapping of paleo-lines of coast in the continental platform South of Alagoas. GeoNordeste J. 2019, 1, 60–79. [Google Scholar] [CrossRef]
- Souza-Lima, W.; Andrade, A.J.; Bengtson, P.; Galm, P.C. A Bacia de Sergipe-Alagoas: Evolução geológica, estratigrafia e conteúdo fóssil. Fund. Paleontológica Phoenix Aracaju 2002, 1, 1–34. [Google Scholar]
- Coutinho, P.N. Geologia Marinha da Plataforma Continental Alagoas-Sergipe. Ph.D. Thesis, Centro de Tecnologia, Universidade Federal de Pernambuco, Recife, Brazil, 1976. [Google Scholar]
- Santos, J.R. Feições morfológicas e biofacies como indicadores evolutivos da Plataforma Continental de Sergipe e sul de Alagoas. Ph.D. Thesis, Centro de Tecnologia, Universidade Federal de Sergipe, Recife, Brazil, 2019. [Google Scholar]
- Knoppers, B.A.; Carneiro, M.E.R.; Fontes, L.C.S.; Souza, W.F.L.; Medeiros, P.R.P. Plataforma Continental de Sergipe e Alagoas. In Plataforma Continental de Sergipe e Alagoas: Geoquímica Sedimentar e Comunidade Bêntica; de Lara Palmeira de Macedo Arguelho, M.E.R.C.e.M., Ed.; Editora UFS; Coleção Projeto MARSEAL: São Cristóvão, Brazil, 2018; Volume 2, pp. 11–38. [Google Scholar]
- Bezerra, F.H.R.; Barreto, A.M.F.; Suguio, K. Holocene sea level history on the Rio Grande do Norte State Coastal, Brazil. Mar. Geol. 2003, 196, 73–89. [Google Scholar] [CrossRef]
- Caldas, L.H.O.; Stattegger, K.; Vital, H. Holocene sea-level history and coastal evolution: Evidences from coastal sediments of the northern Rio Grande do Norte coast, NE Brazil. Mar. Geol. 2006, 228, 39–53. [Google Scholar] [CrossRef]
- Medeiros, P.R.P.; Knoppers, B.A.; Santos, R.C.; Souza, W.F.L. Aporte fluvial e dispersão de matéria particulada em suspensão na zona costeira do rio São Francisco (SE/AL). Geochim. Bras. 2007, 21, 212–231. [Google Scholar]
- Oliveira, E.N.; Knoppers, B.A.; Lorenzzetti, J.A.; Medeiros, P.R.P.; Carneiro, M.E.; Souza, W.F.L. A satellite view of riverine turbidity plumes on the NE-E Brazilian coastal zone. Braz. J. Oceanogr. 2012, 60, 283–298. [Google Scholar] [CrossRef] [Green Version]
- Knoppers, B.; Medeiros, P.R.P.; Souza, W.F.L.; Jennerjahn, T. The São Francisco Estuary, Brazil. In Estuaries, Pollution; Wangersky, P., Ed.; Springer Handbook of Environmental Chemistry: Berlin, Germany, 2006; Volume 5, pp. 51–70. [Google Scholar]
- Bittencourt, A.C.S.P.; Dominguez, J.M.L.; Fontes, L.C.S.; Sousa, D.L.; Silva, I.R.; Silva, F.R. Wave refraction, river damming and episodes of severe shoreline erosion: The Sao Francisco River Mouth, Northeastern Brazil. J. Coast. Res. 2007, 23, 930–938. [Google Scholar] [CrossRef] [Green Version]
- Medeiros, P.R.P.; Knoppers, B.; Souza, W.F.L.; Oliveira, E.N. Aporte de material em suspensão no Baixo Rio São Francisco (SE/AL), em diferentes condições hidrológicas. Braz. J. Aquat. Sci. Technol. 2011, 15, 42–53. [Google Scholar] [CrossRef] [Green Version]
- Dominguez, J.M.L.; Guimarães, J.K. Effects of Holocene climate changes and anthropogenic river regulation in the development of a wave-dominated delta: The São Francisco River (Eastern Brazil). Mar. Geol. 2021, 435, 106456. [Google Scholar] [CrossRef]
- Coutinho, P.N. Oceanografia Geológica. In Levantamento do Estado da Arte da Pesquisa dos Recursos Vivos Marinhos do Brasil (Programa REVIZEE); Coutinho, P.N., Ed.; Ministério do Meio Ambiente dos Recursos Hídricos e da Amazônia Legal, Secretaria de Coordenação dos Assuntos do Meio Ambiente: Brasília, Brazil, 2000; p. 75. [Google Scholar]
- Oliveira, M.B. Caracterização Integrada da Linha de Costa do Estado de Sergipe–Brasil. Master’s Thesis, Instituto de Geociências, Universidade Federal da Bahia, Salvador, Brazil, 2003. [Google Scholar]
- Bittencourt, A.C.S.P.; Oliveira, M.B.; Dominguez, J.M.L. Erosão e Progradação do Litoral Brasileiro–Sergipe. In Erosão e Progradação do Litoral Brasileiro; Muehe, D., Ed.; Ministério do Meio Ambiente: Brasília, Brazil, 2006; Volume 2, pp. 213–218. [Google Scholar]
- Fontes, L.C.S.; Santos, J.R.; Santos, L.A.; Mendonça, J.B.S.; Santos, M.S. Geomorfologia da Plataforma Continental de Sergipe-Alagoas. In Geologia e Geomorfologia da Bacia de Sergipe-Alagoas; Fontes, L.C.S., Kowsmann, R.O., Puga-Bernabéu, A., Eds.; Editora UFS: São Cristóvão, Brazil, 2017; Volume 1, pp. 25–61. [Google Scholar]
- PETROBRAS. Projeto de Caracterização Regional da Bacia de Sergipe-Alagoas, Relatório IBAMA; Etapa 3; Volume 3, Algas Calcárias, Corais e Moluscos da Bacia Sergipe-Alagoas; PETROBRAS: Rio de Janeiro, Brasil, 2015. [Google Scholar]
- Carneiro, M.E.R.; Moreira, D.L.; Oliveira, P.; Omena, E.; Garcia, C.A.B.; Alexandre, M.R.; Carreira, R.; Santos, N.C.; Politano, A.T. Delineamento amostral, métodos de campo e análise dos dados. In Plataforma Continental de Sergipe e Alagoas: Geoquímica Sedimentar e Comunidade Bêntica; de Lara Palmeira de Macedo Arguelho, M.E.R.C.e.M., Ed.; Editora UFS; Coleção Projeto MARSEAL: São Cristóvão, Brazil, 2018; Volume 2, pp. 40–59. [Google Scholar]
- Basso, D. Deep rhodolith distribution in the Pontian Islands, Italy: A model for the paleoecology of a temperate sea. Palaeogeogr. Palaeoclim. Palaeoecol. 1998, 137, 173–187. [Google Scholar] [CrossRef]
- Woelkerling, W.J.; Irvine, L.M.; Harvey, A.S. Growth-forms in non-geniculate coralline red algae (Corallinales, Rhodophyta). Aust. Syst. Bot. 1993, 6, 277–293. [Google Scholar] [CrossRef]
- Basso, D.; Nalin, R.; Nelson, C.S. Shallow-water Sporolithon rhodoliths from North Island (New Zealand). Palaios 2009, 24, 92–103. [Google Scholar] [CrossRef]
- Bosence, D.W.J. Description and classification of rhodoliths (rhodoids, rhodolites). In Coated grains; Peryt, T.M., Ed.; Springer: Berlin, Germany, 1983; pp. 217–224. [Google Scholar]
- Graham, D.J.; Midglay, N.G. Graphical representation of a particle shape using triangular diagrams: An Excel spread sheet method. Earth Surf. Processes Landf. 2000, 25, 1473–1477. [Google Scholar] [CrossRef]
- Maneveldt, G.W.; Van Der Merwe, E. Heydrichia cerasina sp. nov. (Sporolithales, Corallinophycidae, Rhodophyta) from the southernmost tip of Africa. Phycologia 2012, 51, 11–21. [Google Scholar] [CrossRef]
- Bahia, R.G. Algas Coralináceas Formadoras de Rodolitos da Plataforma Continental Tropical e Ilhas Oceânicas do Brasil: Levantamento florístico e taxonomia. Ph.D. Thesis, Escola Nacional de Botânica Tropical, Rio de Janeiro, Brazil, 2014. [Google Scholar]
- Leão, L.A.S.; Bahia, R.G.; Jesionek, M.B.; Adey, W.H.; Johnson, G.; Salgado, L.T.; Pereira, R.C. Sporolithon franciscanum sp. nov. (Sporolithales, Rhodophyta), a New Rhodolith-Forming Species from Northeast Brazil. Diversity 2020, 12, 199. [Google Scholar] [CrossRef]
- Costa, I.O.; Jesus, P.B.; Jesus, T.S.; Souza, P.S.; Horta, P.A.; Nunes, J.M.D.C. Reef-building coralline algae from the Southwest Atlantic: Filling gaps with the recognition of Harveylithon (Corallinaceae, Rhodophyta) on the Brazilian Coast. J. Phycol. 2019, 55, 1370–1385. [Google Scholar] [CrossRef]
- Bernard, G.; Romero-Ramirez, A.; Tauran, A.; Pantalos, M.; Deflandre, B.; Grall, J.; Grémare, A. Declining maerl vitality and habitat complexity across a dredging gradient: Insights from in situ sediment profile imagery (SPI). Sci. Rep. 2019, 9, 16463. [Google Scholar] [CrossRef]
- Dunham, R.J. Classification of carbonate rocks according to depositional texture. In Classification of Carbonate Rocks; Ham, W.E., Ed.; AAPG Mem: Tulsa, OK, USA, 1962; pp. 108–121. [Google Scholar]
- Embry, A.F.; Klovan, J.E. Absolute water depth limits of late Devonian paleoecological zones. Geol. Rundsch. 1972, 61, 672–686. [Google Scholar] [CrossRef]
- Bassi, D.; Iryu, Y.; Humblet, M.; Matsuda, H.; Machiyama, H.; Sasaki, K.; Matsuda, S.; Arai, K.; Inoue, T. Recent macroids on the Kikai-jima shelf, Central Ryukyu Islands, Japan. Sedimentology 2012, 59, 2024–2041. [Google Scholar] [CrossRef]
- Beijbom, O.; Edmunds, P.J.; Roelfsema, C.; Smith, J.; Kline, D.I.; Neal, B.; Dunlap, M.J.; Moriarty, V.; Fan, T.-Y.; Tan, C.-J.; et al. Towards automated annotation of benthic survey images: Variability of human experts and operational modes of automation. PLoS ONE 2015, 10, e0130312. [Google Scholar] [CrossRef]
- Nitsch, F.; Nebelsick, J.H.; Bassi, D. Constructional and Destructional patterns—Void classification of rhodoliths from Giglio Island, Italy. Palaios 2015, 30, 680–691. [Google Scholar] [CrossRef]
- Stuiver, M.; Reimer, P.J.; Reimer, R.W. CALIB 8.2. 2020. Available online: http://calib.org (accessed on 28 August 2020).
- Heaton, T.J.; Köhler, P.; Butzin, M.; Bard, E.; Reimer, R.W.; Austin, W.E.N.; Bronk Ramsey, C.; Hughen, K.A.; Kromer, B.; Reimer, P.J.; et al. Marine20-the marine radiocarbon age calibration curve (0–55,000 cal BP). Radiocarbon 2020, 62, 779–820. [Google Scholar] [CrossRef]
- Holz, V.L.; Bahia, R.G.; Karez, C.S.; Vieira, F.V.; Moraes, F.C.; Vale, N.F.; Sudatti, D.B.; Salgado, L.T.; Moura, R.L.; Amado-Filho, G.M.; et al. Structure of Rhodolith Beds and Surrounding Habitats at the Doce River Shelf (Brazil). Diversity 2020, 12, 75. [Google Scholar] [CrossRef] [Green Version]
- Stelzer, P.S.; Mazzuco, A.C.A.; Gomes, L.E.; Martins, J.; Netto, S.; Bernardino, A.F. Taxonomic and functional diversity of benthic macrofauna associated with rhodolith beds in SE Brazil. PeerJ 2021, 9, e11903. [Google Scholar] [CrossRef]
- Moura, R.L.; Amado-Filho, G.M.; Moraes, F.C.; Brasileiro, P.S.; Salomon, P.S.; Mahiques, M.M.; Bastos, A.C.; Almeida, M.G.; Silva, J.M.; Araujo, B.F.; et al. An extensive reef system at the Amazon River mouth. Sci. Adv. 2016, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Vale, N.F.L.; Amado-Filho, G.M.; Braga, J.C.; Brasileiro, P.S.; Karez, C.S.; Moraes, F.C.; Bahia, R.G.; Bastos, A.C.; Moura, R.L. Structure and composition of rhodoliths from the Amazon River mouth, Brazil. J. South Am. Earth Sci. 2018, 84, 149–159. [Google Scholar] [CrossRef]
- Brasileiro, P.S.; Braga, J.C.; Amado-Filho, G.M.; Leal, R.N.; Bassi, D.; Franco, T.; Bastos, A.C.; Moura, R.L. Burial rate determines Holocene rhodolith development on the Brazilian Shelf. Palaios 2018, 33, 464–477. [Google Scholar] [CrossRef]
- Pereira-Filho, G.H.; Shintate, G.S.; Kitahara, M.V.; Moura, R.L.; Amado-Filho, G.M.; Bahia, R.G.; Moraes, F.C.; Neves, L.M.; Francini, C.L.B.; Gibran, F.Z.; et al. The southernmost Atlantic coral reef is off the subtropical island of Queimada Grande (24° S), Brazil. Bull. Mar. Sci. 2019, 95, 277–287. [Google Scholar] [CrossRef]
- Sissini, M.N.; Koerich, G.; de Barros-Barreto, M.B.; Coutinho, L.M.; Gomes, F.P.; Oliveira, W.; Costa, I.O.; Nunes, J.M.C.; Henriques, M.C.; Vieira-Pinto, T.; et al. Diversity, distribution, and environmental drivers of coralline red algae: The major reef builders in the Southwestern Atlantic. Coral Reefs 2021. [Google Scholar] [CrossRef]
- Vieira, F.V.; Bastos, A.C.; Quaresma, V.S.; Leite, M.D.; Costa, A., Jr.; Oliveira, K.S.S.; Dalvi, C.F.; Bahia, R.G.; Holz, V.L.; Moura, R.L.; et al. Along-shelf changes in mixed carbonate-siliciclastic sedimentation patterns. Cont. Shelf Res. 2019, 187, 103964. [Google Scholar] [CrossRef]
- Ribeiro, R.F.; Dominguez, J.M.L.; Santos, A.A.; Rangel, A.G.A.N. Continuous canyon-river connection on a passive margin: The case of São Francisco Canyon (eastern Brazil). Geomorphology 2021, 375, 107549. [Google Scholar] [CrossRef]
- Pereira-Filho, G.H.; Veras, P.C.; Francini-Filho, R.B.; Moura, R.L.; Pinheiro, H.T.; Gibran, F.Z.; Matheus, Z.; Neves, L.M.; Amado-Filho, G.M. Effects of the sand tilefish Malacanthus plumieri on the structure and dynamics of a rhodolith bed in the Fernando de Noronha Archipelago, tropical West Atlantic. Mar. Ecol. Progr. Ser. 2015, 541, 65. [Google Scholar] [CrossRef]
- Amado-Filho, G.M.; Moura, R.L.; Bastos, A.C.; Francini-Filho, R.B.; Pereira-Filho, G.H.; Bahia, R.G.; Moraes, F.C.; Motta, F.S. Mesophotic ecosystems of the unique South Atlantic atoll are composed by rhodolith beds and scattered consolidated reefs. Mar. Biodiv. 2016, 46, 933–936. [Google Scholar] [CrossRef]
- Prager, E.J.; Ginsburg, R.N. Carbonate nodule growth on Florida’s outer shelf and its implications for fossil interpretations. Palaios 1989, 4, 310–317. [Google Scholar] [CrossRef]
- Pérès, J.; Picard, J.M. Nouveau manuel de bionomie benthique de la mer Méditerranée. Recl. Trav. Stn. Mar. Endoume 1964, 31, 1–131. [Google Scholar]
- Ballesteros, E. Mediterranean coralligenous assemblages: A synthesis of present knowledge. Oceanogr. Mar. Biol. Annu. Rev. 2006, 44, 123–195. [Google Scholar]
- Nalin, R.; Basso, D.; Massari, F. Pleistocene coralline algal build-ups (coralligène de plateau) and associated bioclastic deposits in the sedimentary cover of Cutro marine terrace (Calabria, southern Italy). In Cool-Water Carbonates: Depositional Systems and Palaeoenvironmental Controls; Pedley, H.M., Carannante, G., Eds.; Geological Society, Special Publications: London, UK, 2006; Volume 255, pp. 11–22. [Google Scholar]
- Basso, D.; Nalin, R.; Massari, F. Genesis and composition of the Pleistocene Coralligène de plateau of the Cutro Terrace (Calabria, southern Italy). Neues Jahrbuch für Geologie und Paläontologie–Abhandlungen 2007, 244, 173–182. [Google Scholar] [CrossRef]
- Benisek, M.F.; Marcano, G.; Betzler, C.; Mutti, M. Facies and stratigraphic architecture of a Miocene warm-temperate to tropical fault-block carbonate platform, Sardinia (Central Mediterranean Sea). In Carbonate Systems during the Oligocene-Miocene Climatic Transition; Mutti, M., Piller, W.E., Betzler, C., Eds.; IAS Special Publications: Gent, Belgium, 2010; Volume 42, pp. 129–148. [Google Scholar]
- Braga, J.C.; Martín, J.M. Neogene coralline-algal growth-forms and their palaeoenvironments in the Almanzora River Valley (Almeria, S.E. Spain). Palaeogeogr. Palaeoclim. Palaeoecol. 1988, 67, 285–303. [Google Scholar] [CrossRef]
- Wilson, S.; Blake, C.; Berges, J.A.; Maggs, C.A. Environmental tolerances of free-living coralline algae (maerl): Implications for European marine conservation. Biol. Conserv. 2004, 120, 279–289. [Google Scholar] [CrossRef]
- Ehrhold, A.; Jouet, G.; Le Roy, P.; Jorry, S.J.; Grall, J.; Reixach, T.; Lambert, C.; Gregoire, G.; Goslin, J.; Roubi, A.; et al. Fossil maerl beds as coastal indicators of late Holocene palaeo-environmental evolution in the Bay of Brest (Western France). Palaeogeogr. Palaeoclim. Palaeoecol. 2021, 577, 110525. [Google Scholar] [CrossRef]
- Pardo, C.; Guillemin, M.L.; Pena, V.; Bárbara, I.; Valero, M.; Barreiro, R. Local coastal configuration rather than latitudinal gradient shape clonal diversity and genetic structure of Phymatolithon calcareum maerl Beds in north European Atlantic. Front. Mar. Sci. 2019, 6, 149. [Google Scholar] [CrossRef] [Green Version]
- Steller, D.L.; Hernandez-Ayon, J.M.; Riosmena-Rodriguez, R.; Cabello-Pasini, A. Effect of temperature on photosynthesis, growth and calcification rates of the free- living coralline alga Lithophyllum margaritae. Cienc. Mar. 2007, 33, 441–456. [Google Scholar] [CrossRef] [Green Version]
- Toscano, F.; Vigliotti, M.; Simone, L. Variety of coralline algal deposits (rhodalgal facies) from the Bays of Naples and Pozzuoli (northern Tyrrhenian Sea, Italy). In Cool-Water Carbonates: Depositional Systems and Paleoenvironmental Controls; Pedley, H.M., Carannante, G., Eds.; Geological Society, Special Publications: London, UK, 2006; Volume 225, pp. 85–94. [Google Scholar] [CrossRef]
- Millar, K.R.; Gagnon, P. Mechanisms of stability of rhodolith beds: Sedimentological aspects. Mar. Ecol. Prog. Ser. 2018, 594, 65–83. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, L.G.; James, N.P.; Doubell, M.; Middleton, J.F.; Luick, J.; Currie, D.R.; Bone, Y. Oceanographic controls on shallow-water temperate carbonate sedimentation: Spencer Gulf, South Australia. Sedimentology 2016, 63, 105–135. [Google Scholar] [CrossRef]
- Basso, D. Study of living calcareous algae by a paleontological approach: The non-geniculate Corallinaceac (Rhodophyta) of the soft bottoms of the Tyrrhenian Sea (Western Mediterranean). The genera Phymamlitum Foslie and Mesophyllum Lemoine. Rivista Italiana di Paleontologia e Stratigrafia 1994, 100, 575–596. [Google Scholar] [CrossRef]
- Basso, D. Living calcareous algae by a paleontological approach: The genus Lithothamnion Heydrich nom. cons. from the soft bottoms of the Tyrrhenian Sea (Mediterranean). Rivista Italiana di Paleontologia e Stratigrafia 1995, 101, 349–366. [Google Scholar] [CrossRef]
- Di Geronimo, R.; Giaccone, G. Le alghe calcaree nel Detritico Costiero di Lampedusa (lsole Pelagie). Boll. Acc. Gioenia Sci. Nat. 1994, 27, 5–25. [Google Scholar]
- Braga, J.C. Neogene rhodoliths in the Mediterranean Basins. In Rhodolith/Maërl Beds: A Global Perspective; Riosmena-Rodríguez, R., Nelson, W., Aguirre, J., Eds.; Springer International Publishing: Geerbestrasse, Switzerland, 2017; Volume 15, pp. 169–193. [Google Scholar] [CrossRef]
- McMaster, R.W.; Conover, J.P. Recent algal stromatolites from the Canary Islands. J. Geol. 1966, 74, 647–652. [Google Scholar] [CrossRef]
- Focke, J.W.; Gebelein, C.D. Marine lithification of reef rock and rhodolites at a fore-reef slope locality (−50m) off Bermuda. Geol. Mijnb. 1978, 57, 163–171. [Google Scholar]
- Reid, R.P.; Macintyre, I.G. Foraminiferal-algal nodules from the Eastern Caribbean: Growth history and implications on the value of nodules as paleoenvironmental indicators. Palaios 1988, 3, 424–435. [Google Scholar] [CrossRef]
- Minnery, G.A. Crustose coralline algae from the Flower Garden Banks, northwestern Gulf of Mexico: Controls on distribution and growth morphology. J. Sed. Petrol. 1990, 60, 992–1007. [Google Scholar]
- Littler, M.M.; Littler, D.S.; Hanisak, M.D. Deep-water rhodolith, productivity, and growth history at sites of formation and subsequent degradation. J. Exp. Mar. Biol. Ecol. 1991, 150, 163–182. [Google Scholar] [CrossRef]
- Checconi, A.; Bassi, D.; Carannante, G.; Monaco, P. Re-deposited rhodoliths in the middle Miocene hemipelagic deposits of vitulano (Southern Apennines, Italy): Coralline assemblage characterization and related trace fossils. Sediment. Geol. 2010, 225, 50–66. [Google Scholar] [CrossRef]
- Coutinho, L.M.; Gomes, F.P.; Sissini, M.N.; Vieira-Pinto, T.; de Oliveira Henriques, M.C.M.; Oliveira, M.C.; Horta, P.A.; de Barros Barreto, M.B.B. Cryptic diversity in non-geniculate coralline algae: A new genus Roseolithon (Hapalidiales, Rhodophyta) and seven new species from the Western Atlantic. Eur. J. Phycol. 2021, 1–24. [Google Scholar] [CrossRef]
- Tâmega, F.T.S.; Bassi, D.; Figueiredo, M.A.; Cherkinsky, A. Deep-water rhodolith bed from central Brazilian continental shelf, Campos Basin: Coralline algal and faunal taxonomic composition. J. Coral Reef Stud. 2014, 16, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.; Hall-Spencer, J.M. Effects of Ocean Warming and Acidification on Rhodolith/Maërl Beds. In Rhodolith/Maerl Beds: A Global Perspective; Riosmena-Rodrigues, R., Nelson, W., Aguirre, J., Eds.; Coastal Research Library, Springer: New York, NY, USA, 2017; Volume 15, pp. 55–85. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Sector | Water Depth (m) | Latitude | Longitude | Samples (n) |
|---|---|---|---|---|---|
| 1 | North | 30 | −10.2789 | −35.9792 | 07 |
| 2 | North | 30 | −10.4875 | −36.1245 | 173 |
| 3 | Central | 27 | −10.8223 | −36.6118 | 04 |
| 4 | Central | 47 | −10.8694 | −36.5472 | 141 |
| 5 | South | 47 | −11.1642 | −36.8839 | 05 |
| 6 | South | 54 | −11.3389 | −37.0556 | 18 |
| 7 | South | 50 | −11.5989 | −37.2384 | 56 |
| Site | Sector | (n) | Size | CCA Growth-Forms | Rhodolith Shapes (%) | Rhodolith-Forming Coralline Algae Species | |||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| L | M | S | EN | EW | FR | L | SP | SSP | EL | SEL | DI | SDI | H | HR | HC | Hsp | Lisp | PF | Spsp | LtP | Ltsp | Lsp | Rs | ME | Msp | SFr | Ssp | Psp | |||
| 1 | N | 7 | 7 | 0 | 0 | 4 | 1 | 0 | 2 | 14.29 | 28.57 | 0 | 0 | 14.29 | 42.86 | x | x | x | x | x | x | ||||||||||
| 2 | N | 173 | 3 | 1 | 169 | 2 | 0 | 169 | 2 | 75 | 25 | 0 | 0 | 0 | 0 | x | x | x | x | x | x | ||||||||||
| 3 | C | 4 | 4 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 50 | x | x | ||||||||||||||
| 4 | C | 141 | 0 | 0 | 141 | 2 | 7 | 130 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | x | x | x | |||||||||||||
| 5 | S | 5 | 4 | 1 | 0 | 4 | 0 | 0 | 1 | 60 | 40 | 0 | 0 | 0 | 0 | x | x | xx | x | x | x | x | xx | xx | |||||||
| 6 | S | 18 | 18 | 0 | 0 | 11 | 1 | 0 | 6 | 22.22 | 38.89 | 5.56 | 16.67 | 0 | 16.67 | x | x | x | x | x | xx | xx | x | x | |||||||
| 7 | S | 56 | 54 | 2 | 0 | 36 | 8 | 0 | 12 | 39.29 | 53.57 | 0 | 1.79 | 0 | 5.36 | x | x | xx | x | xx | xx | xx | x | x | x | xx | |||||
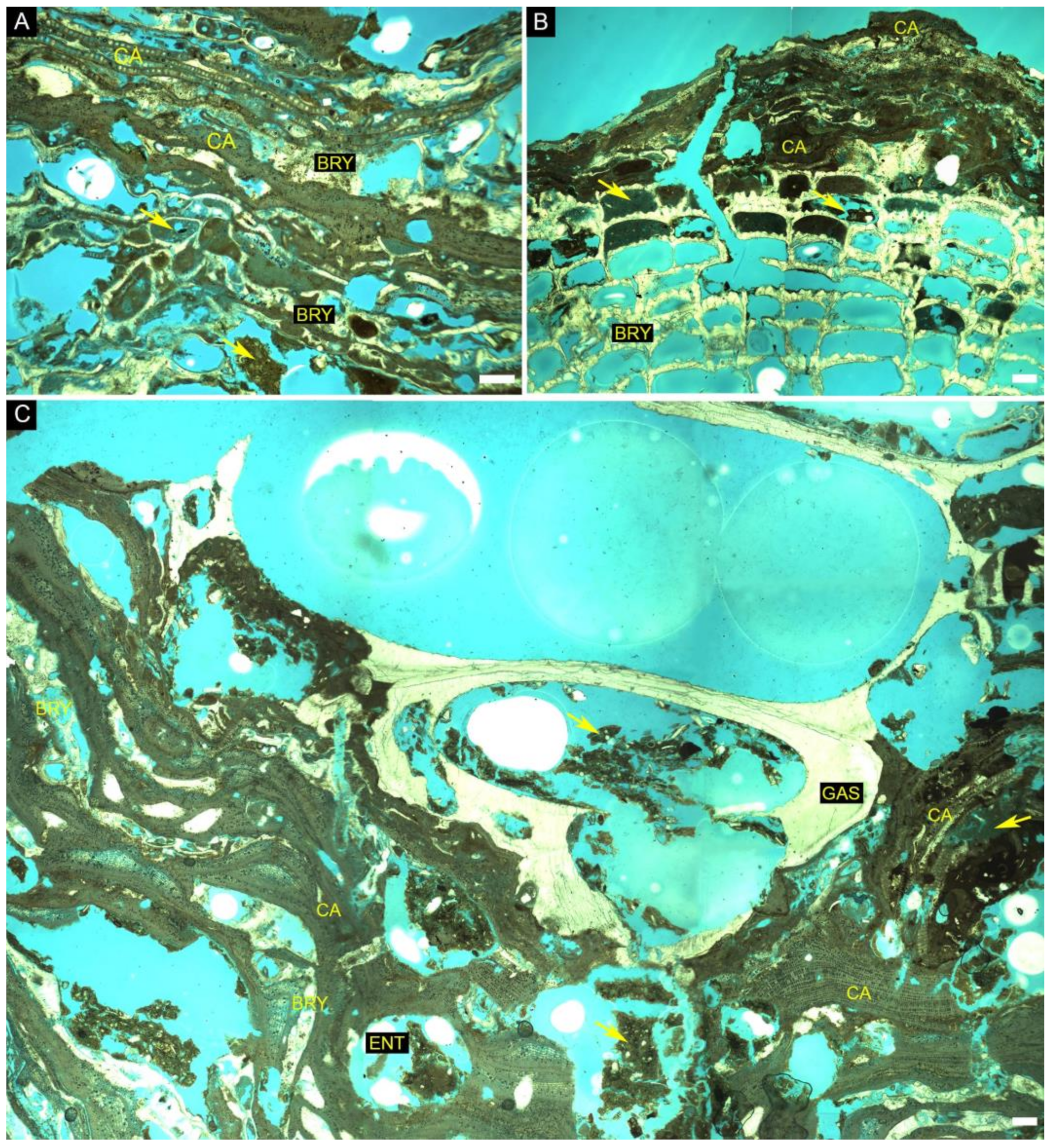
| North Sector (Site 1) | Central Sector (Site 3) | South Sector (Site 5) | South Sector (Site 6) | South Sector (Site 7) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample number | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8-1gp | 8-2gp | 9 | 10-1gp | 10-2gp |
| Bryozoan | 3.94 | 12.29 | 7.54 | 0.58 | 8.4 | 5.62 | 47.4 | 17.26 | 37.27 | 16.38 | 26.25 | 7.75 |
| Coral | 20.15 | 0 | 0.91 | 52.15 | 2.31 | 8.51 | 0 | 0 | 0 | 0 | 0 | 0 |
| Geniculate coralline algae | 1.34 | 0 | 0 | 0 | 0.8 | 0.35 | 0 | 0 | 0 | 0 | 0 | 0 |
| Non-geniculate coralline Algae | 19.28 | 56.53 | 48.36 | 6.92 | 31.36 | 52.7 | 21.46 | 23.47 | 32.38 | 38.19 | 28.75 | 59.41 |
| Echinodermata | 2.13 | 0.45 | 0.88 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Foraminifera | 0.42 | 0.44 | 0.46 | 0 | 1.38 | 0 | 0 | 0.39 | 0.71 | 0 | 0 | 0.64 |
| Gastropod | 0 | 0 | 0.24 | 0.1 | 0 | 0 | 0 | 13.4 | 0.36 | 9.93 | 7.5 | 1.57 |
| Serpulid | 0 | 1.67 | 0 | 0.83 | 0.83 | 0.6 | 0 | 0 | 0.7 | 0.42 | 0 | 0 |
| Sponge spicules | 0 | 0 | 0.54 | 3.44 | 0 | 5.03 | 0 | 0 | 0 | 0 | 0 | 1.77 |
| Filling sediments | 33.16 | 6.69 | 5.54 | 9.15 | 13.02 | 5.36 | 1.9 | 6.28 | 3.68 | 5.15 | 17.5 | 9.95 |
| Siliciclastic grains | 5.48 | 2.23 | 0 | 3.68 | 4.65 | 0 | 1.83 | 2.14 | 0 | 0 | 0 | 0 |
| Borings | 13.7 | 24.17 | 16.25 | 18.06 | 20.6 | 32.5 | 11.67 | 19.6 | 17.5 | 16.67 | 17.5 | 17.3 |
| Constructional voids | 0 | 0 | 9.51 | 0.91 | 0.9 | 0.42 | 14.47 | 16.32 | 4.71 | 11.04 | 2.5 | 0.21 |
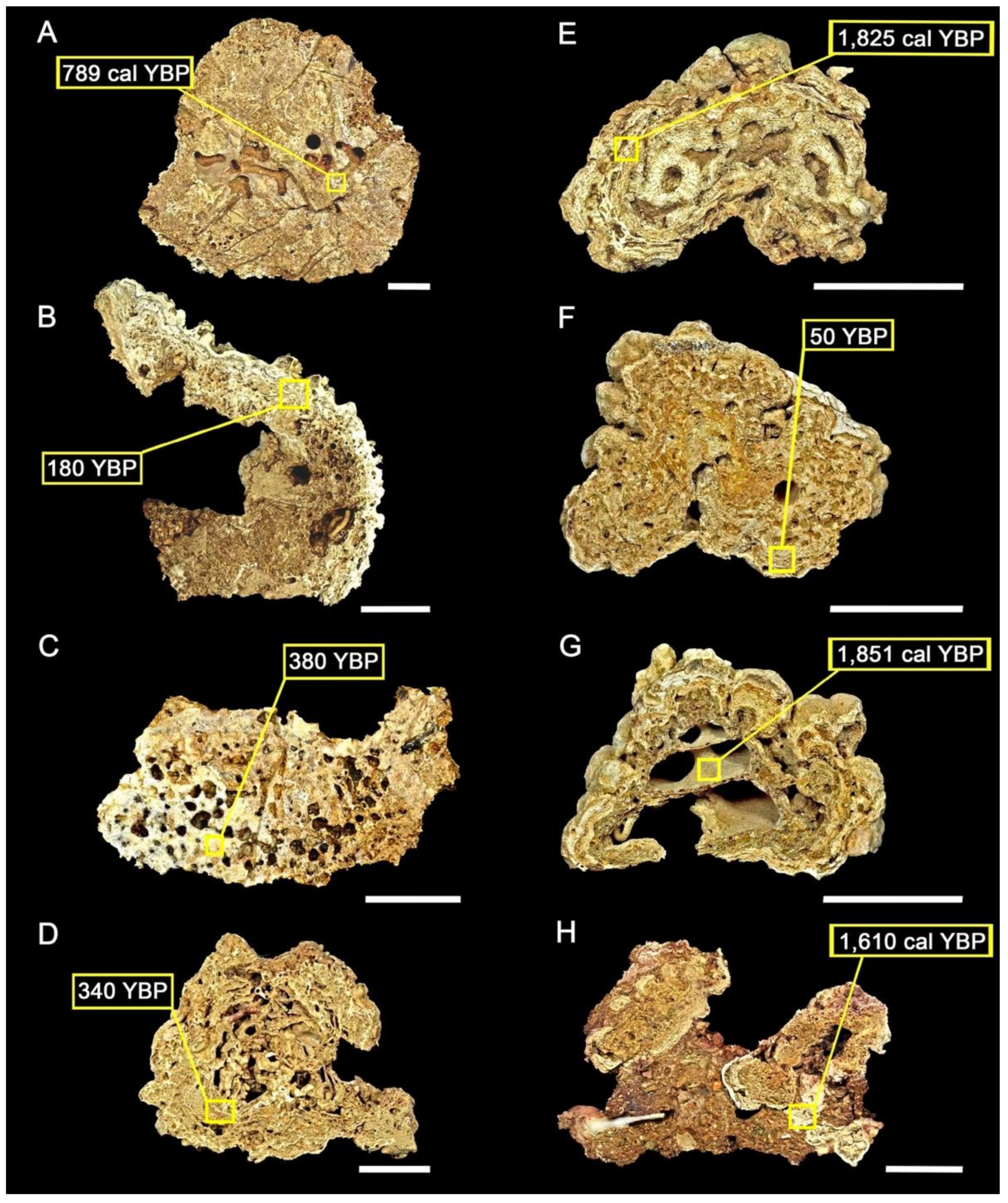
| Site | Sector | Latitude | Longitude | Water Depth (m) | Dated Component | Conventional Age (BP) | Calibrated 14C Age (Cal yr BP) | 95% Range (cal BP) |
|---|---|---|---|---|---|---|---|---|
| 1 | N | −10.2789 | −35.9792 | 30 | Coral | 1840 ± 200 | 789 | 666–922 |
| 1 | N | −10.2789 | −35.9792 | 30 | Coralline algae | 180 * ± 200 | - | - |
| 3 | C | −10.8223 | −36.6118 | 27 | Coral | 380 * ± 250 | - | - |
| 5 | S | −11.1642 | −36.8839 | 47 | Coralline algae | 340 * ± 200 | - | - |
| 6 | S | −11.3389 | −37.0556 | 54 | Coralline algae | 760 ± 200 | 1825 | 1700–1950 |
| 7 | S | −11.5989 | −37.2384 | 50 | Coralline algae | 50 * ± 200 | - | - |
| 7 | S | −11.5989 | −37.2384 | 50 | Gastropod | 4040 ± 250 | 1851 | 1690–2014 |
| 7 | S | −11.5989 | −37.2384 | 50 | Bryozoans | 960 ± 250 | 1610 | 1476–1755 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vale, N.F.L.; Braga, J.C.; Bastos, A.C.; Moraes, F.C.; Karez, C.S.; Bahia, R.G.; Leão, L.A.; Pereira, R.C.; Amado-Filho, G.M.; Salgado, L.T. Structure and Composition of Rhodolith Beds from the Sergipe-Alagoas Basin (NE Brazil, Southwestern Atlantic). Diversity 2022, 14, 282. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040282
Vale NFL, Braga JC, Bastos AC, Moraes FC, Karez CS, Bahia RG, Leão LA, Pereira RC, Amado-Filho GM, Salgado LT. Structure and Composition of Rhodolith Beds from the Sergipe-Alagoas Basin (NE Brazil, Southwestern Atlantic). Diversity. 2022; 14(4):282. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040282
Chicago/Turabian StyleVale, Nicholas F. L., Juan C. Braga, Alex C. Bastos, Fernando C. Moraes, Claudia S. Karez, Ricardo G. Bahia, Luis A. Leão, Renato C. Pereira, Gilberto M. Amado-Filho, and Leonardo T. Salgado. 2022. "Structure and Composition of Rhodolith Beds from the Sergipe-Alagoas Basin (NE Brazil, Southwestern Atlantic)" Diversity 14, no. 4: 282. https://0-doi-org.brum.beds.ac.uk/10.3390/d14040282