
Using Long-Term Removal Data to Manage a Crown-of-Thorns Starfish Population

Abstract
:1. Introduction
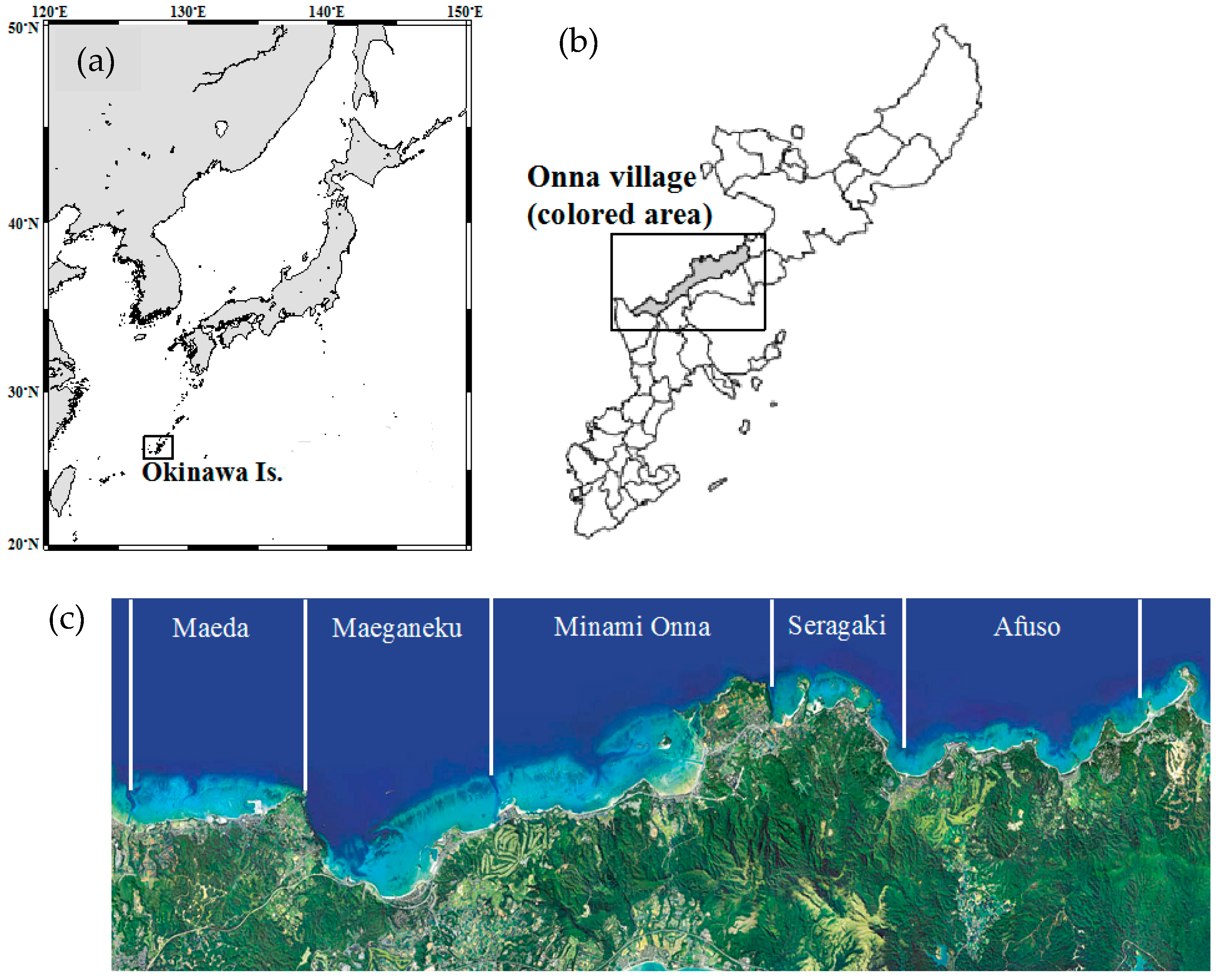
2. Materials and Methods
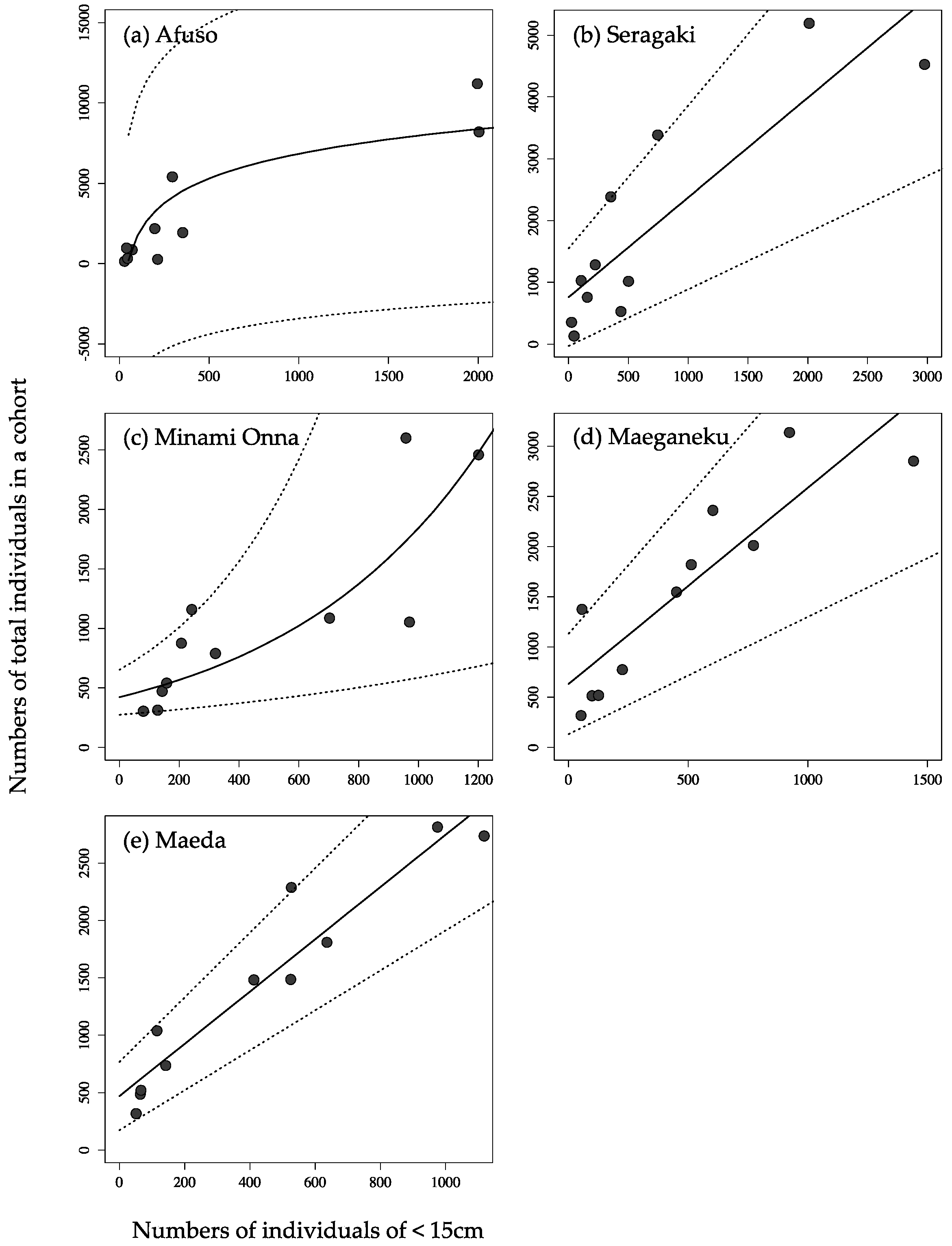
3. Results and Discussion
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Pratchett, M.S.; Caballes, C.F.; Rivera-Posada, J.A.; Sweatman, H.P.A. Limits to understanding and managing outbreaks of Crown-of-Thorns starfish (Acanthaster spp.). Oceanogr. Mar. Biol. 2014, 52, 133–200. [Google Scholar]
- De'ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [PubMed]
- Pratchett, M.S.; Schenk, T.J.; Baine, M.; Syms, C.; Baird, A.H. Selective coral mortality associated with outbreaks of Acanthaster planci L. in Bootless Bay, Papua New Guinea. Mar. Environ. Res. 2009, 67, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Baird, A.H.; Pratchett, M.S.; Hoey, A.S.; Herdiana, Y.; Campbell, S.J. Acanthaster planci is a major cause of coral mortality in Indonesia. Coral Reefs 2013, 32, 803–812. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; Schwieder, H.; Heiden, J.; Weiand, L.; Wild, C.; Jompa, J.; Ferse, S.C.A.; Teichberg, M. A recent outbreak of crown-of-thorns starfish (Acanthaster planci) in the Spermonde Archipelago, Indonesia. Reg. Environ. Chang. 2015, 15, 1157–1162. [Google Scholar] [CrossRef]
- Yamaguchi, M. Acanthaster planci infestations of reefs and coral assemblages in Japan: A retrospective analysis of control efforts. Coral Reefs 1986, 5, 23–30. [Google Scholar] [CrossRef]
- Nakamura, M.; Okaji, K.; Higa, Y.; Yamagawa, E.; Mitarai, S. Spatial and temporal population dynamics of the crown-of-thorns starfish, Acanthaster planci, over a 24-year period along the central west coast of Okinawa Island, Japan. Mar. Biol. 2014, 161, 2521–2530. [Google Scholar] [CrossRef]
- Bos, A.R. Crown-of-thorns outbreak at the Tubbataha Reef UNESCO World Heritage Site. Zool. Stud. 2010, 49, 124. [Google Scholar]
- Adjeroud, M.; Michonneau, F.; Edmounds, P.J.; Chancerelle, Y.; Lison de Loma, T.; Penin, L.; Thibaut, L.; Vidal-Dupiol, J.; Salvat, B.; Galzin, R. Recurrent disturbances recovery trajectories, and resilience of coral assemblages on a South Central Pacific reef. Coral Reefs 2009, 28, 775–780. [Google Scholar] [CrossRef]
- Kayal, M.; Vercelloni, J.; de Loma, T.L.; Bosserelle, P.; Chancerelle, Y.; Geoffroy, S.; Stievenart, C.; Michonneau, F.; Penin, L.; Planes, S.; et al. Predator crown-of-thorns starfish (Acanthaster planci) outbreak, mass mortality of corals, and cascading effects on reef fish and benthic communities. PLoS ONE 2012, 7, e47363. [Google Scholar] [CrossRef] [PubMed]
- Roche, R.C.; Pratchett, M.S.; Carr, P.; Turner, J.R.; Wagner, D.; Head, C.; Sheppard, C.R.C. Localized outbreaks of Acanthaster planci at an isolated and unpopulated reef atoll in the Chagos Archipelago. Mar. Biol. 2015, 162, 1695–1704. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Okaji, K.; De'ath, G. Three lines of evidence to link outbreaks of the corwn-of-thorns seastar Acanthaster planci to the release of larval food limitation. Coral Reefs 2010, 29, 593–605. [Google Scholar] [CrossRef]
- Bostrom-Einarsson, L.; Rivera-Posada, J. Controlling outbreaks of the coral-eating crown-of-thorns starfish using a single injection of common household vinegar. Coral Reefs 2016, 35, 223–228. [Google Scholar] [CrossRef]
- Bos, A.R.; Gumanao, G.S.; Mueller, B.; Saceda-Cardoza, M.M.E. Management of crown-of-thorns sea star (Acanthaster planci L.) outbreaks: Removal success depends on reef topography and timing within the reproduction cycle. Ocean Coast Manag. 2013, 71, 116–122. [Google Scholar] [CrossRef]
- Yokochi, H. The early detection of Acanthaster outbreaks by monitoring the algae-eating juvenile starfish. Midoriishi 1998, 9, 12–14. (In Japanese) [Google Scholar]
- Research Institute for Subtropical Regions. Manual for Monitoring of Juvenile Crown-of-Thorns Starfish; Research Institute for Subtropical Regions: Naha, Japan, 2006; p. 29. (In Japanese) [Google Scholar]
- Yamaguchi, M. Growth of juvenile Acanthaster planci (L) in the laboratory. Pac. Sci. 1974, 28, 123–138. [Google Scholar]
- Lucas, J.S. Growth, maturation and effects of diet in Acanthaster planci (L.) (Asteroidae) and hybrids reared in the laboratory. J. Exp. Mar. Biol. Ecol. 1984, 79, 129–147. [Google Scholar] [CrossRef]
- Birkeland, C.; Lucas, J.S. Acanthaster planci: Major Management Problems of Coral Reefs; CRC Press, Inc.: Florida, FL, USA, 1990. [Google Scholar]
- Babcook, R.C.; Mundy, C.N. Reproductive biology, spawning and field fertilization rates of Acanthaster planci. Aust. J. Mar. Freshw. Res. 1992, 43, 525–534. [Google Scholar] [CrossRef]
- Pratchett, M.S. Dynamics of an outbreak population of Acanthaster planci at Lizard Island, northern Great Barrier Reef (1995–1999). Coral Reefs 2005, 24, 453–462. [Google Scholar] [CrossRef]
- Nakamura, M.; Kumagai, N.H.; Sakai, K.; Okaji, K.; Ogasawara, K.; Mitarai, S. Spatial variability in recruitment of acroporid corals and predatory starfish along the Onna coast, Okinawa, Japan. Mar. Ecol. Prog. Ser. 2015, 540, 1–12. [Google Scholar] [CrossRef]
- Moran, P.J.; De'ath, G. Estimates of the abundance of the crown-of-thorns starfish Acanthaster planci in outbreaking and non-outbreaking populations on reefs within the Great Barrier Reef. Mar. Biol. 1992, 113, 509–515. [Google Scholar] [CrossRef]
- Yasuda, N.; Ogasawara, K.; Kajiwara, K.; Ueno, M.; Oki, K.; Taniguchi, H.; Kakuma, S.; Okaji, K.; Nadaoka, K. Latitudinal differentiation in the reproduction patterns of the crown-of-thorns starfish Acanthaster planci through the Ryukyu Island Archipelago. Plankton Benthos Res. 2010, 5, 156–164. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Estimate | Std. Error | t-value | p-value | |
|---|---|---|---|---|
| Intercept | 1035 | 208.5 | 4.961 | 8.83 × 10−6 |
| 1st year class (<10 cm) | 6.355 | 1.486 | 4.277 | 8.74 × 10−5 |
| 1st year class (<10 cm)_AF | 14.81 | 2.321 | 6.384 | 5.98 × 10−8 |
| 1st year class (<10 cm)_MO | −3.278 | 2.810 | −1.167 | 0.249 |
| 1st year class (<10 cm)_MG | −2.946 | 2.454 | −1.201 | 0.236 |
| 1st year class (<10 cm)_MD | −2.421 | 2.974 | −0.814 | 0.420 |
| AIC | 948.1 |
| Estimate | Std. Error | t-value | p-value | |
|---|---|---|---|---|
| Intercept | 637.4 | 161.6 | 3.943 | 25.5 × 10−5 |
| 1st year class (<15 cm) | 1.680 | 0.243 | 6.927 | 8.61 × 10−9 |
| 1st year class (<15 cm)_AF | 2.552 | 0.325 | 7.841 | 3.35 × 10−10 |
| 1st year class (<15 cm)_MO | −0.475 | 0.493 | −0.964 | 0.340 |
| 1st year class (<15 cm)_MG | 0.267 | 0.475 | 0.562 | 0.577 |
| 1st year class (<15 cm)_MD | 0.367 | 0.533 | 0.688 | 0.495 |
| AIC | 905.8 |
| Sector | Models | ||
|---|---|---|---|
| Linear | Logarithmic | Exponential | |
| Afuso | n.a. | 0.808 | 0.791 |
| Seragaki | 0.753 | 0.716 | 0.549 |
| Minami Onna | 0.720 | 0.687 | 0.723 |
| Maeganeku | 0.779 | 0.739 | 0.567 |
| Maeda | 0.911 | 0.897 | 0.783 |
| Sector | Best Model |
|---|---|
| Afuso | Y = 2218 log (X) − 8484 |
| Seragaki | Y = 1.615 X + 758.4 |
| Minami Onna | Y = 422.2 × 1.001x |
| Maeganeku | Y = 1.955 X + 631.2 |
| Maeda | Y = 2.277 X + 470.0 |
| Sector | Number of Individuals that Escaped Removal | |
|---|---|---|
| Cohort 2013 | Cohort 2014 | |
| Afuso | 577 | 6350 |
| Seragaki | 0 | 1315 |
| Minami Onna | 0 | 357 |
| Maeganeku | 0 | 1057 |
| Maeda | 508 | 644 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakamura, M.; Higa, Y.; Kumagai, N.H.; Okaji, K. Using Long-Term Removal Data to Manage a Crown-of-Thorns Starfish Population. Diversity 2016, 8, 24. https://0-doi-org.brum.beds.ac.uk/10.3390/d8040024
Nakamura M, Higa Y, Kumagai NH, Okaji K. Using Long-Term Removal Data to Manage a Crown-of-Thorns Starfish Population. Diversity. 2016; 8(4):24. https://0-doi-org.brum.beds.ac.uk/10.3390/d8040024
Chicago/Turabian StyleNakamura, Masako, Yoshimi Higa, Naoki H. Kumagai, and Ken Okaji. 2016. "Using Long-Term Removal Data to Manage a Crown-of-Thorns Starfish Population" Diversity 8, no. 4: 24. https://0-doi-org.brum.beds.ac.uk/10.3390/d8040024