
Interactive Effects of Endogenous and Exogenous Nutrition on Larval Development for Crown-Of-Thorns Starfish


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Collection and Maintenance of Specimens
2.2. Maternal Feeding Treatments
2.3. Spawning Induction and Oocyte Metrics
2.4. Fertilization and Larval Rearing
2.5. Statistical Analyses
3. Results
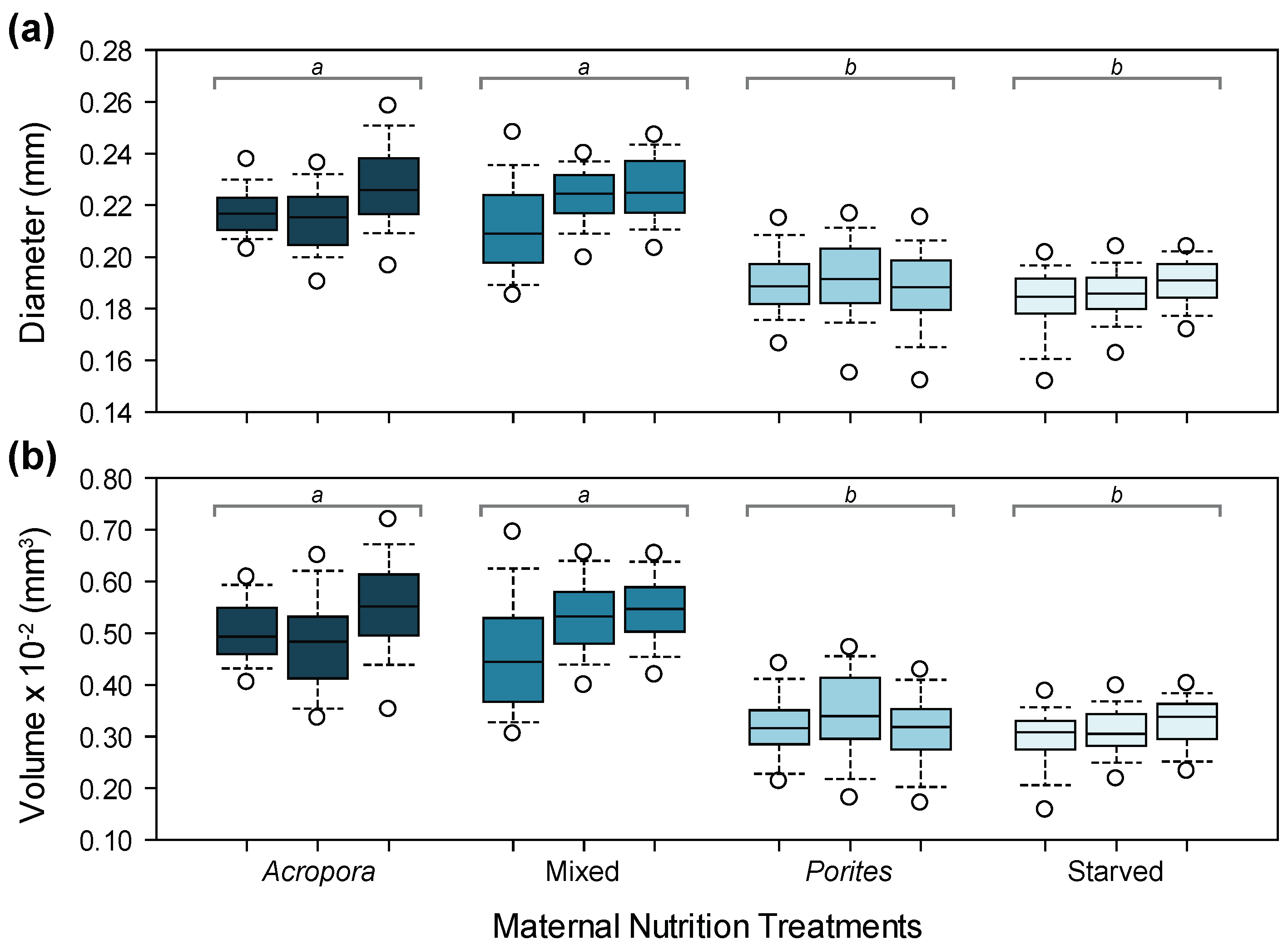
3.1. Maternal and Oocyte Metrics
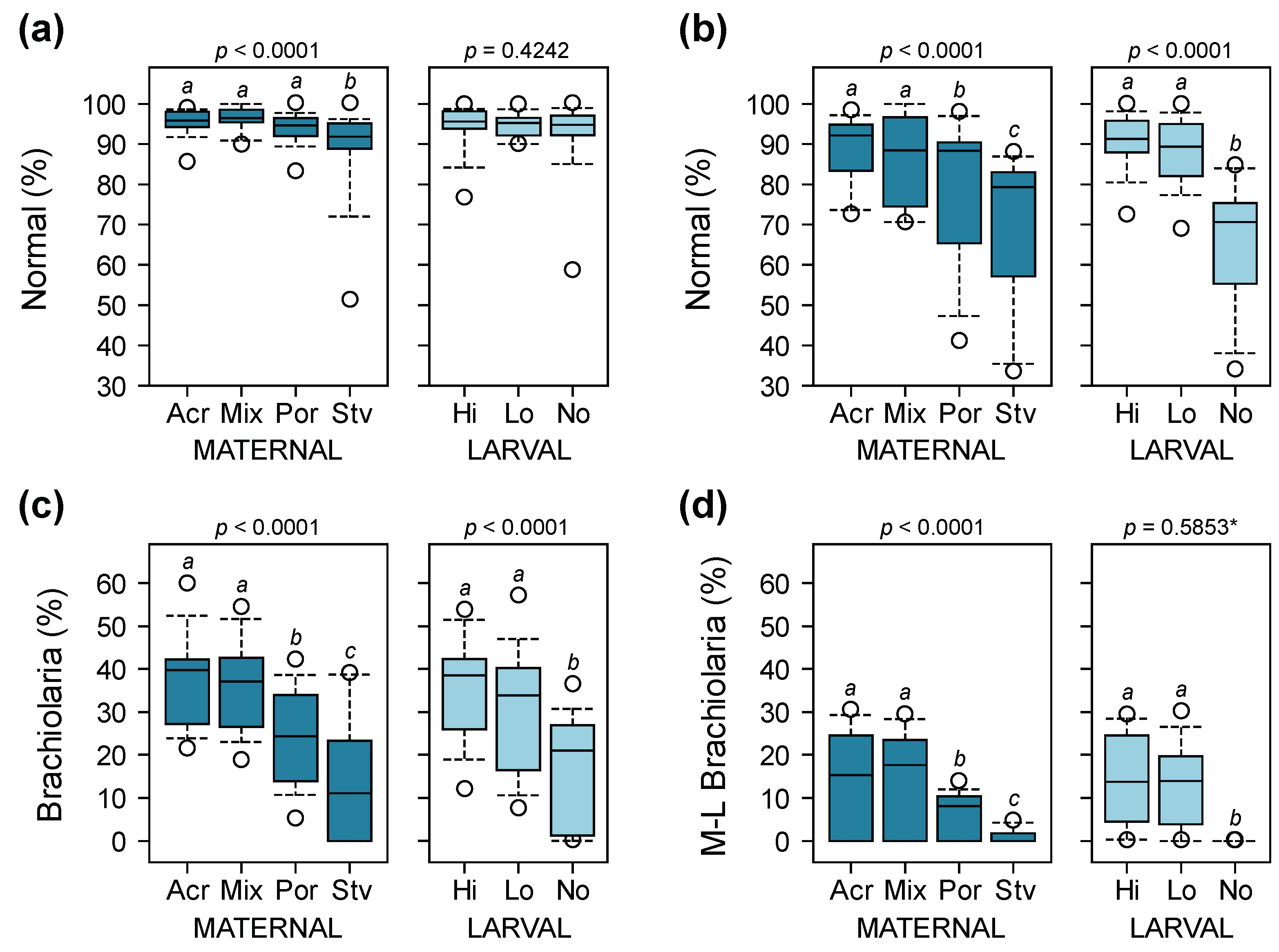
3.2. Larval Survival
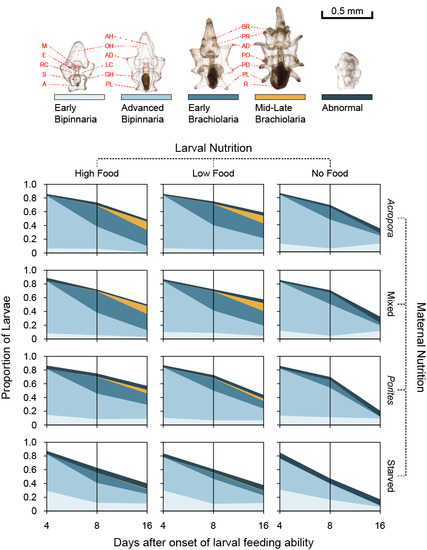
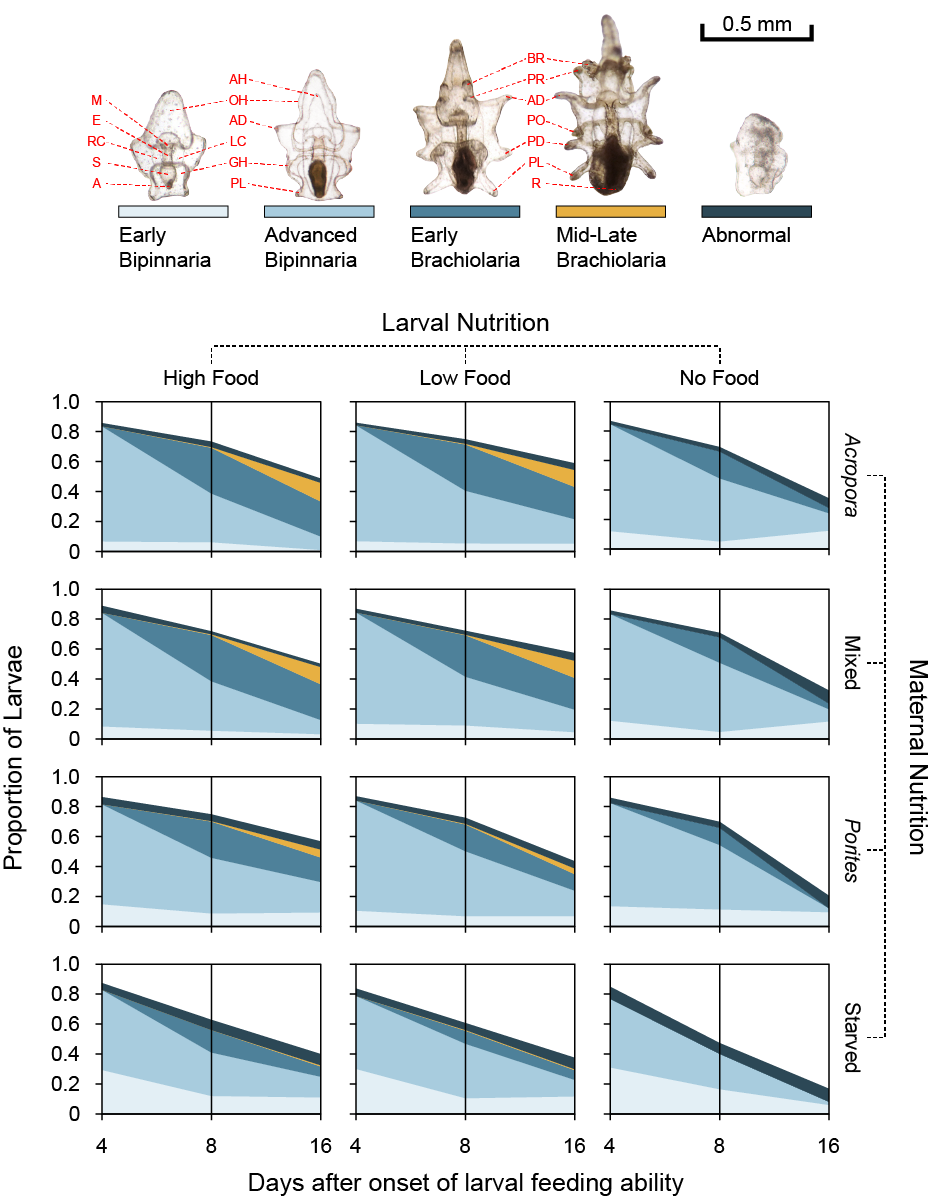
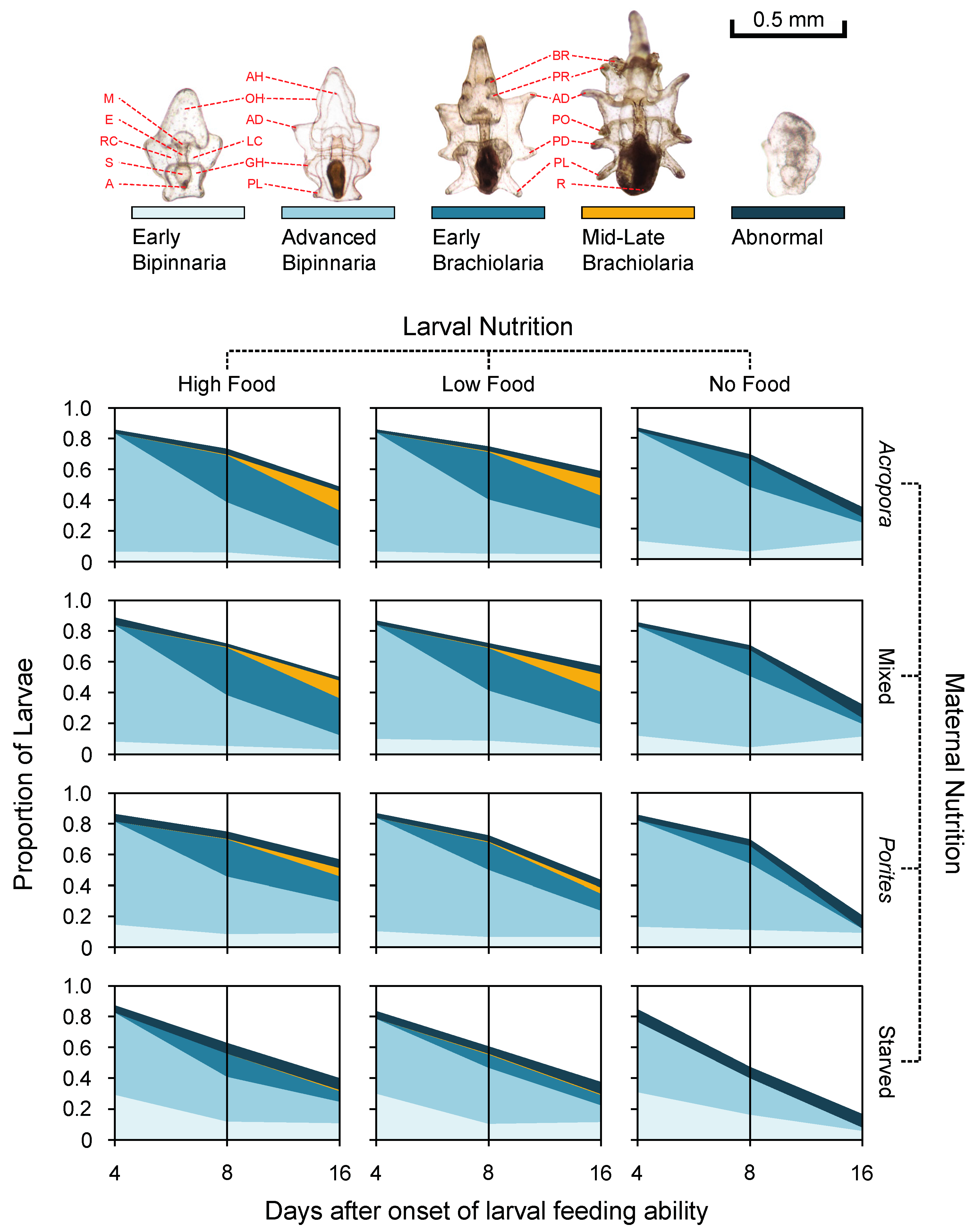
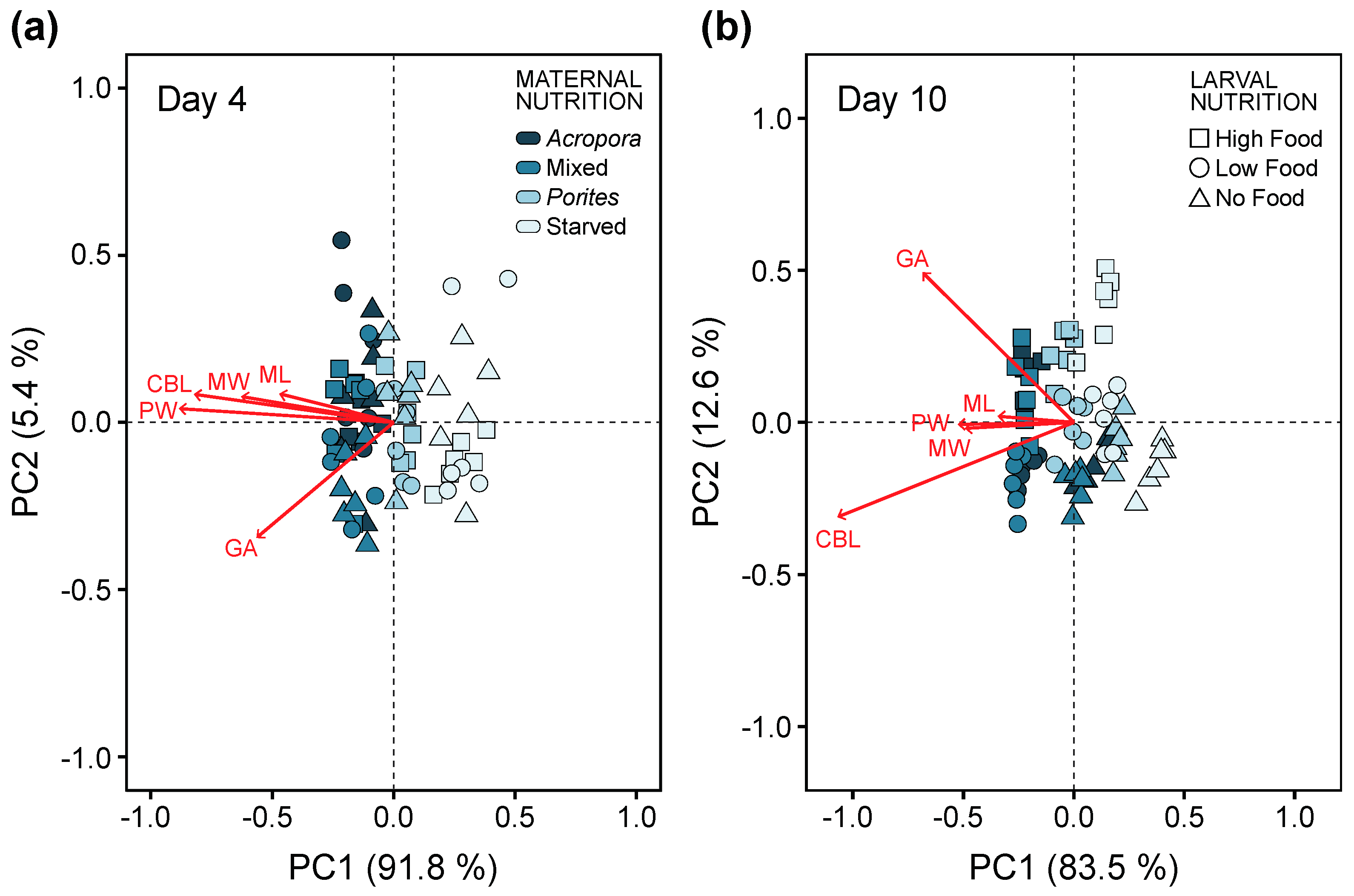
3.3. Larval Development
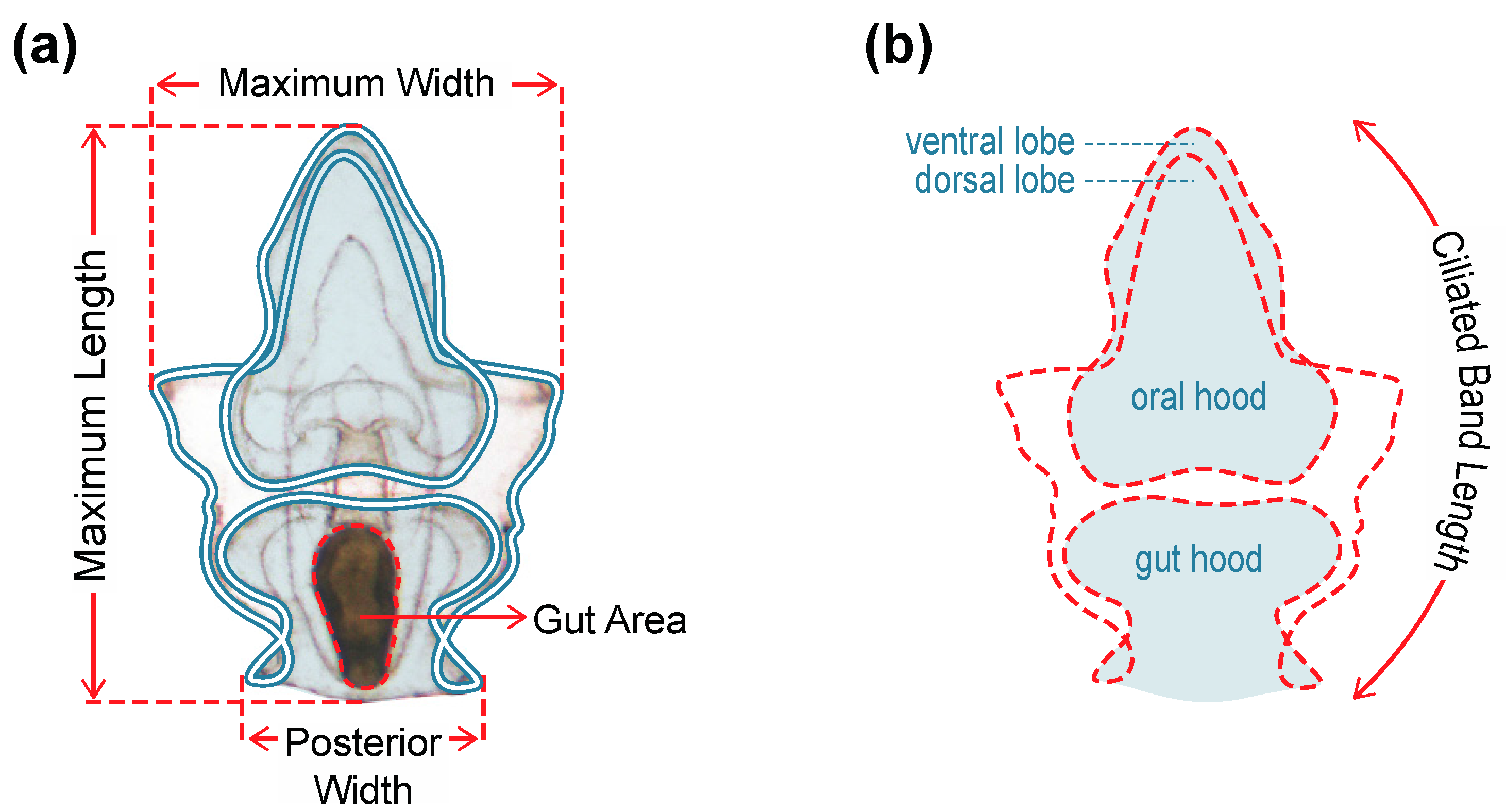
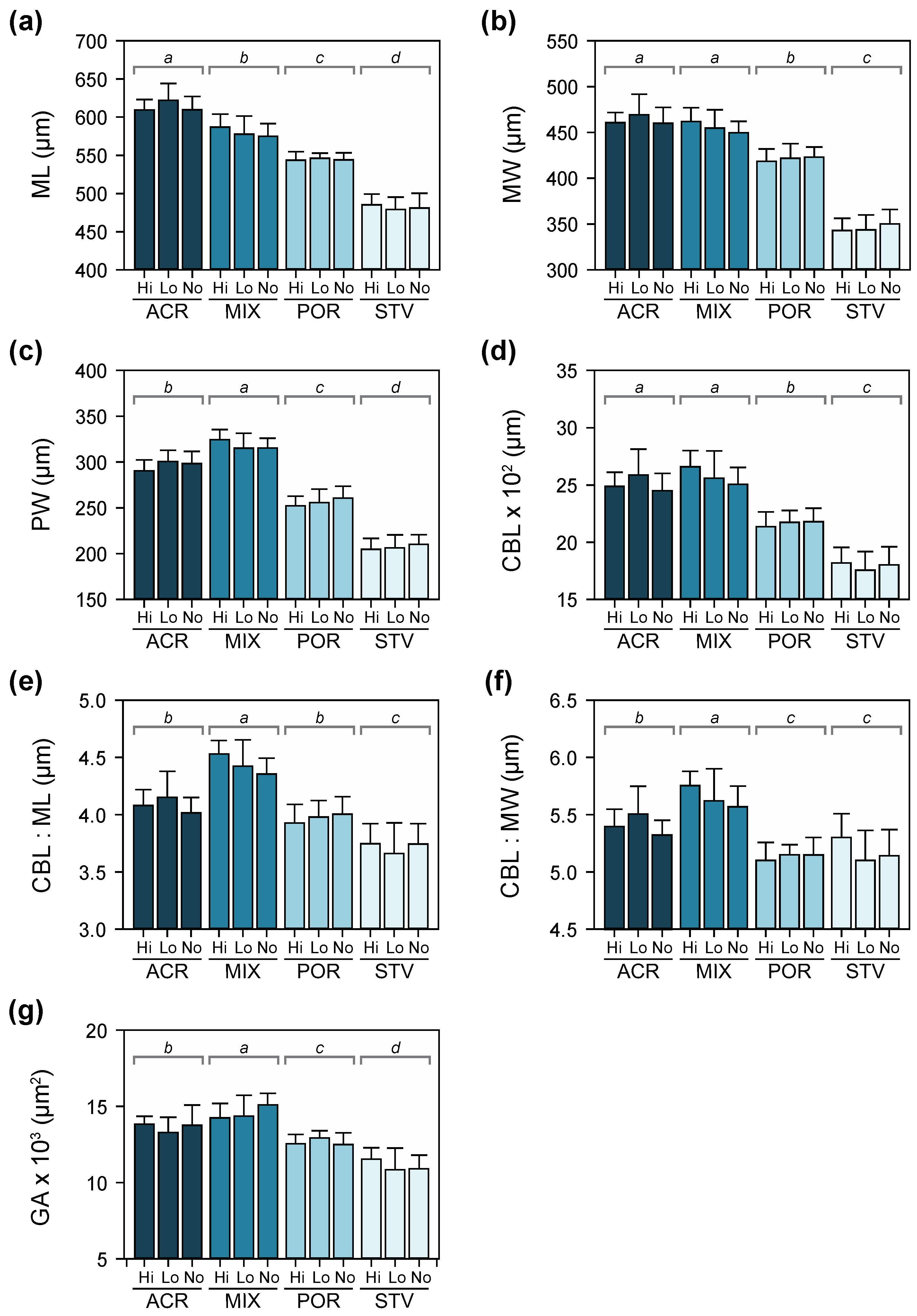
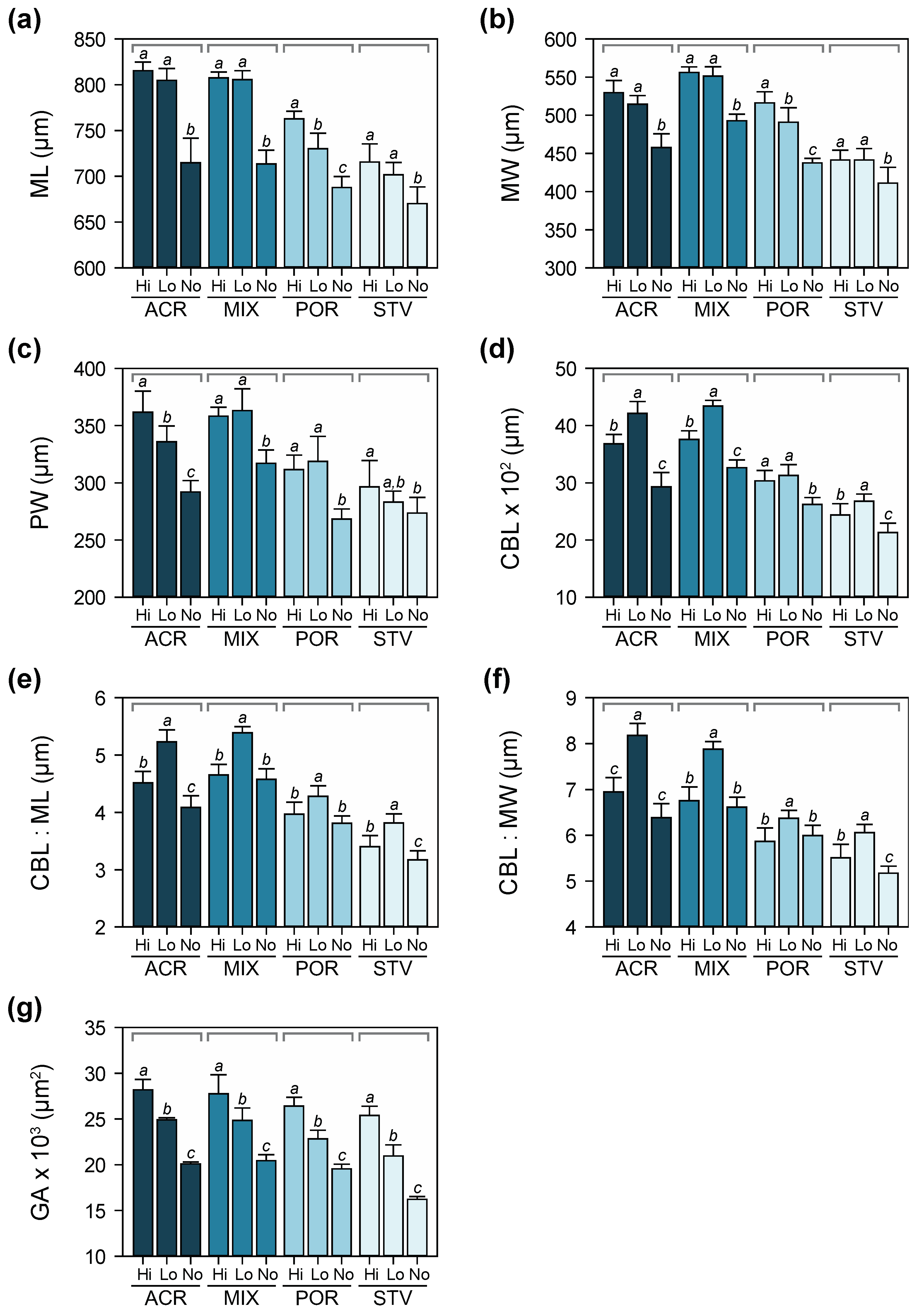
3.4. Larval Growth and Morphometry
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Graham, N.A.J.; Jackson, J.B.C.; Mumby, P.J.; Steneck, R.S. Rising to the challenge of sustaining coral reef resilience. Trends Ecol. Evol. 2010, 25, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Pratchett, M.S.; Caballes, C.F.; Rivera-Posada, J.A.; Sweatman, H.P.A. Limits to understanding and managing outbreaks of crown-of-thorns starfish (Acanthaster spp.). Oceanogr. Mar. Biol. Annu. Rev. 2014, 52, 133–200. [Google Scholar]
- Westcott, D.A.; Fletcher, C.S.; Babcock, R.C.; Plaganyi-Lloyd, E. A Strategy to Link Research and Management of Crown-of-Thorns Starfish on the Great Barrier Reef: An Integrated Pest Management Approach; National Environment Science Programme: Cairns, Australia, 2016. [Google Scholar]
- Hoey, J.; Campbell, M.; Hewitt, C.; Gould, B.; Bird, R. Acanthaster planci invasions: Applying biosecurity practices to manage a native boom and bust coral pest in Australia. Manag. Biol. Invasions 2016, 7, 213–220. [Google Scholar] [CrossRef]
- Moore, R.J. Persistent and transient populations of the crown-of-thorns starfish, Acanthaster planci. Lect. Notes Biomath. 1990, 88, 236–277. [Google Scholar]
- Babcock, R.C.; Milton, D.A.; Pratchett, M.S. Relationships between size and reproductive output in the crown-of-thorns starfish. Mar. Biol. 2016, 163, 234. [Google Scholar] [CrossRef]
- Uthicke, S.; Schaffelke, B.; Byrne, M. A boom–bust phylum? Ecological and evolutionary consequences of density variations in echinoderms. Ecol. Monogr. 2009, 79, 3–24. [Google Scholar] [CrossRef]
- Pratchett, M.S. Dynamics of an outbreak population of Acanthaster planci at Lizard Island, northern Great Barrier Reef (1995–1999). Coral Reefs 2005, 24, 453–462. [Google Scholar] [CrossRef]
- Brodie, J.E.; Fabricius, K.E.; De’ath, G.; Okaji, K. Are increased nutrient inputs responsible for more outbreaks of crown-of-thorns starfish? An appraisal of the evidence. Mar. Pollut. Bull. 2005, 51, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Wooldridge, S.A.; Brodie, J.E. Environmental triggers for primary outbreaks of crown-of-thorns starfish on the Great Barrier Reef, Australia. Mar. Pollut. Bull. 2015, 101, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Houk, P.; Raubani, J. Acanthaster planci outbreaks in Vanuatu coincide with ocean productivity, furthering trends throughout the Pacific Ocean. J. Oceanogr. 2010, 66, 435–438. [Google Scholar] [CrossRef]
- Houk, P. The transition zone chlorophyll front can trigger Acanthaster planci outbreaks in the Pacific Ocean: Historical confirmation. J. Oceanogr. 2007, 63, 149–154. [Google Scholar] [CrossRef]
- Fabricius, K.E.; Okaji, K.; De’ath, G. Three lines of evidence to link outbreaks of the crown-of-thorns seastar Acanthaster planci to the release of larval food limitation. Coral Reefs 2010, 29, 593–605. [Google Scholar] [CrossRef]
- Wolfe, K.; Graba-Landry, A.; Dworjanyn, S.A.; Byrne, M. Larval starvation to satiation: Influence of nutrient regime on the success of Acanthaster planci. PLoS ONE 2015, 10, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pratchett, M.S.; Dworjanyn, S.; Mos, B.; Caballes, C.F.; Thompson, C.; Blowes, S. Larval survivorship and settlement of crown-of-thorns starfish (Acanthaster cf. solaris) at varying algal cell densities. Diversity 2017, 9, 2. [Google Scholar] [CrossRef]
- McEdward, L.R. Reproductive strategies of marine benthic invertebrates revisited: Facultative feeding by planktotrophic larvae. Am. Nat. 1997, 150, 48–72. [Google Scholar] [CrossRef] [PubMed]
- Bertram, D.F.; Strathmann, R.R. Effects of maternal and larval nutrition on growth and form of planktotrophic larve. Ecology 1998, 79, 315–327. [Google Scholar] [CrossRef]
- Byrne, M.; Sewell, M.A.; Prowse, T.A.A. Nutritional ecology of sea urchin larvae: Influence of endogenous and exogenous nutrition on echinopluteal growth and phenotypic plasticity in Tripneustes gratilla. Funct. Ecol. 2008, 22, 643–648. [Google Scholar] [CrossRef]
- Lucas, J.S. Quantitative studies of feeding and nutrition during larval development of the coral reef asteroid Acanthaster planci (L.). J. Exp. Mar. Biol. Ecol. 1982, 65, 173–193. [Google Scholar] [CrossRef]
- Okaji, K.; Ayukai, T.; Lucas, J.S. Selective feeding by larvae of the crown-of-thorns starfish, Acanthaster planci (L.). Coral Reefs 1997, 16, 47–50. [Google Scholar] [CrossRef]
- Uthicke, S.; Logan, M.; Liddy, M.; Francis, D.S.; Hardy, N.; Lamare, M.D. Climate change as an unexpected co-factor promoting coral eating seastar (Acanthaster planci) outbreaks. Sci. Rep. 2015, 5, 8402. [Google Scholar] [CrossRef] [PubMed]
- Birkeland, C. Terrestrial runoff as a cause of outbreaks of Acanthaster planci (Echinodermata: Asteroidea). Mar. Biol. 1982, 69, 175–185. [Google Scholar] [CrossRef]
- Caballes, C.F.; Pratchett, M.S.; Kerr, A.M.; Rivera-Posada, J.A. The role of maternal nutrition on oocyte size and quality, with respect to early larval development in the coral-eating starfish, Acanthaster planci. PLoS ONE 2016, 11, e0158007. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.S. Growth, maturation and effects of diet in Acanthaster planci (L.) (Asteroidea) and hybrids reared in the laboratory. J. Exp. Mar. Biol. Ecol. 1984, 79, 129–147. [Google Scholar] [CrossRef]
- Kayal, M.; Vercelloni, J.; Lison de Loma, T.; Bosserelle, P.; Chancerelle, Y.; Geoffroy, S.; Stievenart, C.; Michonneau, F.; Penin, L.; Planes, S.; et al. Predator crown-of-thorns starfish (Acanthaster planci) outbreak, mass mortality of corals, and cascading effects on reef fish and benthic communities. PLoS ONE 2012, 7, e47363. [Google Scholar] [CrossRef] [PubMed]
- Ormond, R.F.G.; Hanscomb, N.J.; Beach, D.H. Food selection and learning in the crown-of-thorns starfish, Acanthaster planci (L.). Mar. Behav. Physiol. 1976, 4, 93–105. [Google Scholar] [CrossRef]
- Revelante, N.; Gilmartin, M. Dynamics of phytoplankton in the Great Barrier Reef lagoon. J. Plankton Res. 1982, 4, 47–76. [Google Scholar] [CrossRef]
- McEdward, L.R. Comparative morphometrics of echinoderm larvae. II. Larval size, shape, growth, and the scaling of feeding and metabolism in echinoplutei. J. Exp. Mar. Biol. Ecol. 1986, 96, 267–286. [Google Scholar] [CrossRef]
- Hart, M.W.; Strathmann, R.R. Functional consequences of phenotypic plasticity in echinoid larvae. Biol. Bull. 1994, 186, 291–299. [Google Scholar] [CrossRef]
- George, S.B. Phenotypic plasticity in the larvae of Luidia foliolata (Echinodermata: Asteroidea). In Echinoderms Through Time; David, B., Guille, A., Féral, J.-P., Roux, M., Eds.; August Aimé Balkema: Rotterdam, The Netherlands, 1994; p. 20. [Google Scholar]
- Wolfe, K.; Graba-Landry, A.; Dworjanyn, S.A.; Byrne, M. Larval phenotypic plasticity in the boom-and-bust crown-of-thorns seastar, Acanthaster planci. Mar. Ecol. Prog. Ser. 2015, 539, 179–189. [Google Scholar] [CrossRef]
- Caballes, C.F.; Pratchett, M.S. Reproductive biology and early life history of the crown-of-thorns starfish. In Echinoderms: Ecology, Habitats and Reproductive Biology; Whitmore, E., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2014; pp. 101–146. [Google Scholar]
- Pratchett, M.S. Influence of coral symbionts on feeding preferences of crown-of thorns starfish Acanthaster planci in the western Pacific. Mar. Ecol. Prog. Ser. 2001, 214, 111–119. [Google Scholar] [CrossRef]
- Lucas, J.S. Reproductive and larval biology of Acanthaster planci (L.) in Great Barrier Reef Waters. Micronesica 1973, 9, 197–203. [Google Scholar]
- Conand, C. Distribution, reproductive cycle and morphometric relationships of Acanthaster planci (Echinodermata: Asteroidea) in New Caledonia, western tropical Pacific. In Proceedings of the 5th International Echinoderm Conference, Galway, Ireland, 24–29 September 1984; pp. 499–506.
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; DeBose, J.L.; Ajani, P.; Teixeira da Silva, E.; Petus, C.; Brodie, J.E. Phytoplankton in the Great Barrier Reef: Microscopy analysis of community structure in high flow events. In Report to the National Environmental Research Program; Reef and Rainforest Research Centre Limited: Cairns, Australia, 2013; p. 68. [Google Scholar]
- Yamaguchi, M. Early life histories of coral reef asteroids, with special reference to Acanthaster planci (L.). In Biology and Geology of Coral Reefs; Jones, O.A., Endean, R., Eds.; Academic Press, Inc.: New York, NY, USA, 1973; Volume 2, pp. 369–387. [Google Scholar]
- Byrne, M.; Barker, M.F. Embryogenesis and larval development of the asteroid Patiriella regularis viewed by light and scanning electron microscopy. Biol. Bull. 1991, 180, 332–345. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.R-project.org/ (accessed on 28 February 2017).
- Agresti, A. Introduction to Categorical Data Analysis; John Wiley & Sons Ltd.: New York, NY, USA, 1996. [Google Scholar]
- Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: New York, NY, USA, 2002. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar]
- Anderson, M.J. Permutation tests for univariate or multivariate analysis of variance and regression. Can. J. Fish. Aquat. Sci. 2001, 58, 626–639. [Google Scholar] [CrossRef]
- George, S.B. Echinoderm egg and larval quality as a function of adult nutritional state. Oceanol. Acta 1996, 19, 297–308. [Google Scholar]
- George, S.B. Population differences in maternal size and offspring quality for Leptasterias epichlora (Brandt) (Echinodermata: Asteroidea). J. Exp. Mar. Biol. Ecol. 1994, 175, 121–131. [Google Scholar] [CrossRef]
- De Jong-Westman, M.; Qian, P.-Y.; March, B.E.; Carefoot, T.H. Artificial diets in sea urchin culture: Effects of dietary protein level and other additives on egg quality, larval morphometrics, and larval survival in the green sea urchin, Strongylocentrotus droebachiensis. Can. J. Zool. 1995, 73, 2080–2090. [Google Scholar] [CrossRef]
- George, S.B. Egg quality, larval growth and phenotypic plasticity in a forcipulate seastar. J. Exp. Mar. Biol. Ecol. 1999, 237, 203–224. [Google Scholar] [CrossRef]
- Herrera, J.C.; McWeeney, S.K.; McEdward, L.R. Diversity of energetic strategies among echinoid larvae and the transition from feeding to nonfeeding development. Oceanol. Acta 1996, 19, 313–321. [Google Scholar]
- Keesing, J.K. Feeding Biology of the Crown-of-Thorns Starfish, Acanthaster planci (Linnaeus). Ph.D. Thesis, James Cook University, Townsville, Australia, 1990. [Google Scholar]
- Prowse, T.A.A.; Sewell, M.A.; Byrne, M. Fuels for development: Evolution of maternal provisioning in asterinid sea stars. Mar. Biol. 2008, 153, 337–349. [Google Scholar] [CrossRef]
- Stimson, J.S. Location, quantity and rate of change in quantity of lipids in tissue of Hawaiian hermatypic corals. Bull. Mar. Sci. 1987, 41, 889–904. [Google Scholar]
- George, S.B.; Cellario, C.; Fenaux, L. Population differences in egg quality of Arbacia lixula (Echinodermata: Echinoidea): Proximate composition of eggs and larval development. J. Exp. Mar. Biol. Ecol. 1990, 141, 107–118. [Google Scholar] [CrossRef]
- Patton, J.S.; Battey, J.F.; Rigler, M.W.; Porter, J.W.; Black, C.C.; Burris, J.E. A comparison of the metabolism of bicarbonate 14C and acetate 1-14C and the variability of species lipid compositions in reef corals. Mar. Biol. 1983, 75, 121–130. [Google Scholar] [CrossRef]
- Harland, A.D.; Navarro, J.C.; Spencer Davies, P.; Fixter, L.M. Lipids of some Caribbean and Red Sea corals: total lipid, wax esters, triglycerides and fatty acids. Mar. Biol. 1993, 117, 113–117. [Google Scholar] [CrossRef]
- Yamashiro, H.; Oku, H.; Higa, H.; Chinen, I.; Sakai, K. Composition of lipids, fatty acids and sterols in Okinawan corals. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1999, 122, 397–407. [Google Scholar] [CrossRef]
- Giese, A.C. Lipids in the economy of marine invertebrates. Physiol. Rev. 1966, 46, 244–298. [Google Scholar] [PubMed]
- Arai, I.; Kato, M.; Heyward, A.; Ikeda, Y.; Iizuka, T.; Maruyama, T. Lipid composition of positively buoyant eggs of reef building corals. Coral Reefs 1993, 12, 71–75. [Google Scholar] [CrossRef]
- Imbs, A.B.; Yakovleva, I.M. Dynamics of lipid and fatty acid composition of shallow-water corals under thermal stress: an experimental approach. Coral Reefs 2012, 31, 41–53. [Google Scholar] [CrossRef]
- Shilling, F.M. Morphological and physiological responses of echinoderm larvae to nutritive signals. Am. Zool. 1995, 35, 399–411. [Google Scholar] [CrossRef]
- Miner, B.G. Larval feeding structure plasticity during pre-feeding stages of echinoids: Not all species respond to the same cues. J. Exp. Mar. Biol. Ecol. 2007, 343, 158–165. [Google Scholar] [CrossRef]
- Sinervo, B.; McEdward, L.R. Developmental consequences of an evolutionary change in egg size: An experimental test. Evolution 1988, 42, 885. [Google Scholar] [CrossRef]
- Strathmann, R.R. Length of pelagic period in echinoderms with feeding larvae from the Northeast Pacific. J. Exp. Mar. Biol. Ecol. 1978, 34, 23–27. [Google Scholar] [CrossRef]
- Lane, D.J.W. Acanthaster planci impact on coral communities at permanent transect sites on Bruneian reefs, with a regional overview and a critique on outbreak causes. J. Mar. Biol. Assoc. UK 2011, 92, 803–809. [Google Scholar] [CrossRef]
- Roche, R.C.; Pratchett, M.S.; Carr, P.; Turner, J.R.; Wagner, D.; Head, C.; Sheppard, C.R.C. Localized outbreaks of Acanthaster planci at an isolated and unpopulated reef atoll in the Chagos Archipelago. Mar. Biol. 2015, 162, 1695–1704. [Google Scholar] [CrossRef]
- Suzuki, G.; Yasuda, N.; Ikehara, K.; Fukuoka, K.; Kameda, T.; Kai, S.; Nagai, S.; Watanabe, A.; Nakamura, T.; Kitazawa, S.; et al. Detection of a high-density brachiolaria-stage larval population of crown-of-thorns sea star (Acanthaster planci) in Sekisei Lagoon (Okinawa, Japan). Diversity 2016, 8, 9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SOURCE | df | χ2 | p | Post Hoc |
|---|---|---|---|---|
| Day 4 | ||||
| Maternal Nutrition | 3 | 2.58 | 0.9349 | |
| Larval Nutrition | 2 | 2.12 | 0.8396 | |
| Maternal Nutrition × Larval Nutrition | 6 | 4.71 | 0.9927 | |
| Day 8 | ||||
| Maternal Nutrition | 3 | 140.00 | 0.0001 | Acr = Mix = Por > Stv |
| Larval Nutrition | 2 | 27.92 | 0.1281 | |
| Maternal Nutrition × Larval Nutrition | 6 | 15.63 | 0.8901 | |
| Day 12 | ||||
| Maternal Nutrition | 3 | 98.03 | 0.0035 | Acr = Mix = Por > Stv |
| Larval Nutrition | 2 | 60.24 | 0.0152 | Hi = Lo > No |
| Maternal Nutrition × Larval Nutrition | 6 | 20.05 | 0.8349 | |
| Day 16 | ||||
| Maternal Nutrition | 3 | 61.00 | 0.0072 | Acr = Mix = Por > Stv |
| Larval Nutrition | 2 | 435.23 | <0.0001 | Hi = Lo > No |
| Maternal Nutrition × Larval Nutrition | 6 | 5.81 | 0.9794 |
| Source | df | χ2 | p | Post Hoc |
|---|---|---|---|---|
| Day 4 | ||||
| Maternal Nutrition | 36 | 82.80 | <0.0001 | Acr = Mix = Por ≠ Stv |
| Larval Nutrition | 32 | 7.34 | 0.9663 | |
| Maternal Nutrition × Larval Nutrition | 24 | 4.94 | 1.0000 | |
| Day 8 | ||||
| Maternal Nutrition | 36 | 79.62 | <0.0001 | Acr = Mix ≠ Por = Stv |
| Larval Nutrition 1 | 32 | 51.55 | 0.0157 | |
| Maternal Nutrition × Larval Nutrition | 24 | 22.05 | 0.5763 | |
| Day 16 | ||||
| Maternal Nutrition | 36 | 103.49 | <0.0001 | Acr = Mix ≠ Por = Stv |
| Larval Nutrition | 32 | 143.85 | <0.0001 | Hi = Lo ≠ No |
| Maternal Nutrition × Larval Nutrition | 24 | 12.38 | 0.9753 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caballes, C.F.; Pratchett, M.S.; Buck, A.C.E. Interactive Effects of Endogenous and Exogenous Nutrition on Larval Development for Crown-Of-Thorns Starfish. Diversity 2017, 9, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/d9010015
Caballes CF, Pratchett MS, Buck ACE. Interactive Effects of Endogenous and Exogenous Nutrition on Larval Development for Crown-Of-Thorns Starfish. Diversity. 2017; 9(1):15. https://0-doi-org.brum.beds.ac.uk/10.3390/d9010015
Chicago/Turabian StyleCaballes, Ciemon Frank, Morgan S. Pratchett, and Alexander C. E. Buck. 2017. "Interactive Effects of Endogenous and Exogenous Nutrition on Larval Development for Crown-Of-Thorns Starfish" Diversity 9, no. 1: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/d9010015