
The Relevance of SOCS1 Methylation and Epigenetic Therapy in Diverse Cell Populations of Hepatocellular Carcinoma

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples
2.1.1. Human Tissue Samples
2.1.2. Cell Lines
2.2. Flow Cytometry
2.3. Isolation of Genomic DNA and Bisulfite Conversion
2.4. Methylation-Specific PCR (MS-PCR)
2.5. Treatment In Vitro
2.6. Western Blot Analysis
2.7. Reverse Transcription-Quantitative Real-Time PCR (RT-qPCR)
2.8. Statistical Analysis
3. Results
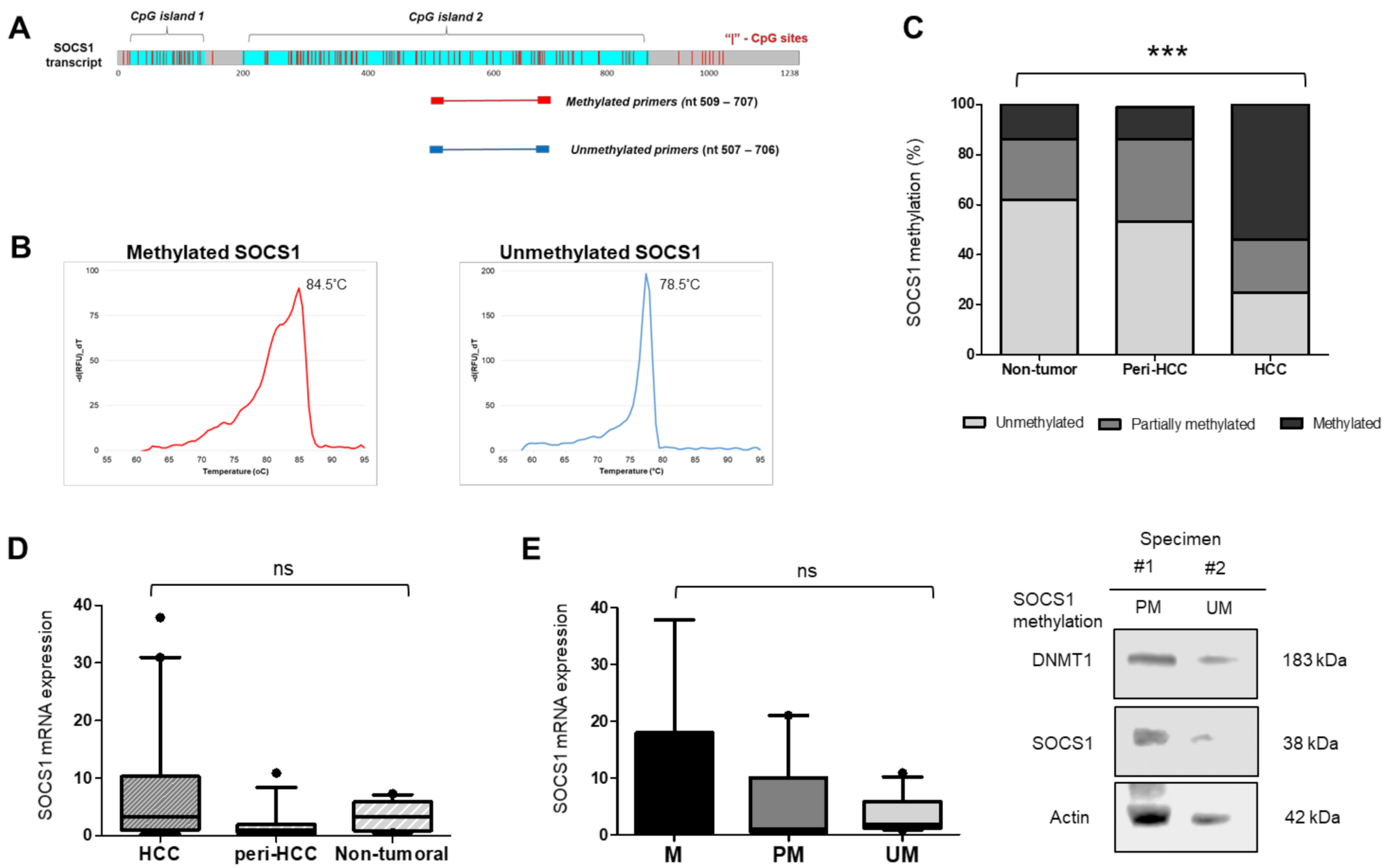
3.1. SOCS1 Methylation Status in HCC Specimens
3.2. SOCS1 mRNA Expression Is Not Correlated with DNA Methylation
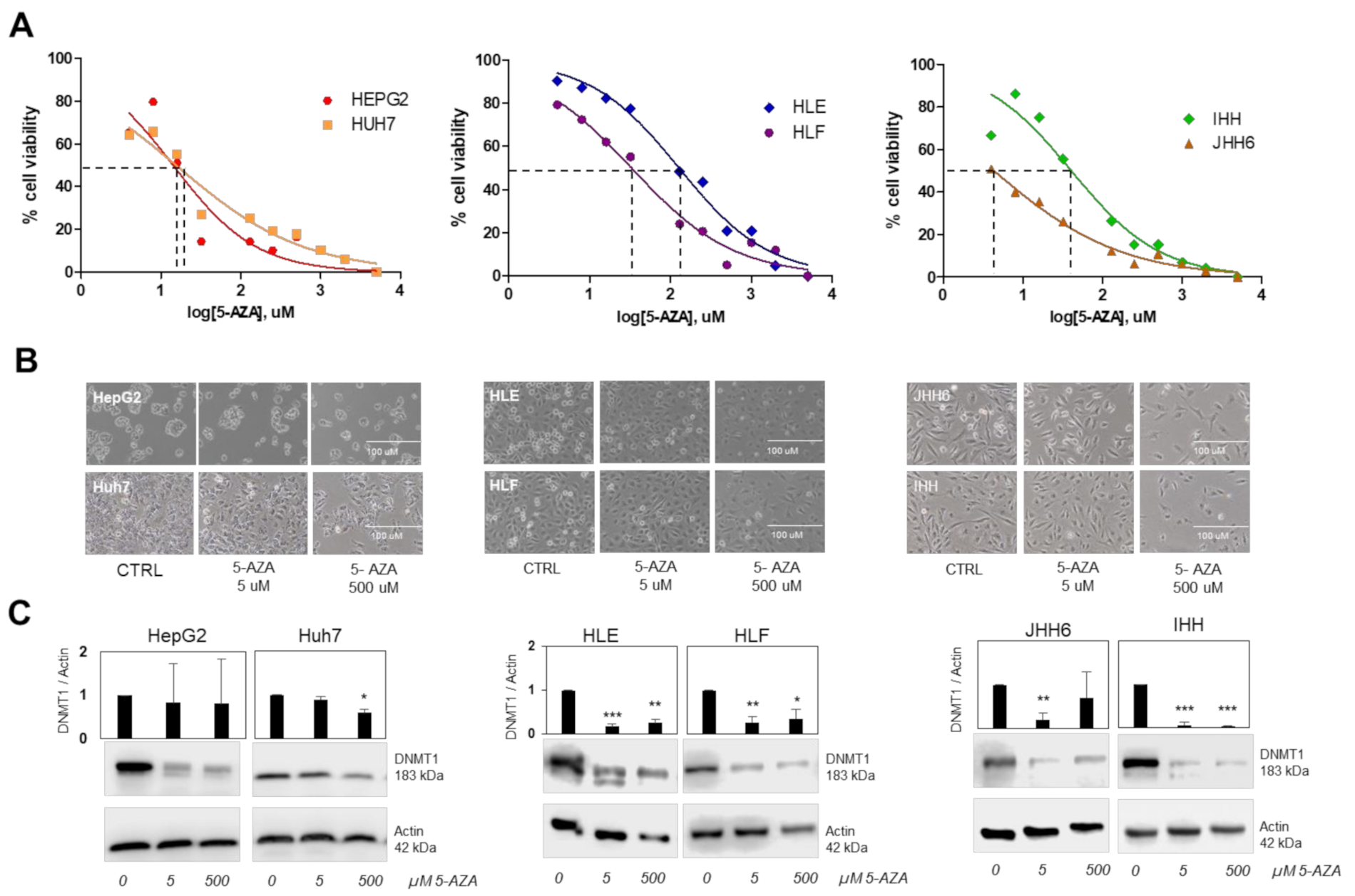
3.3. Demethylation by 5-AZA in Different Cell Types
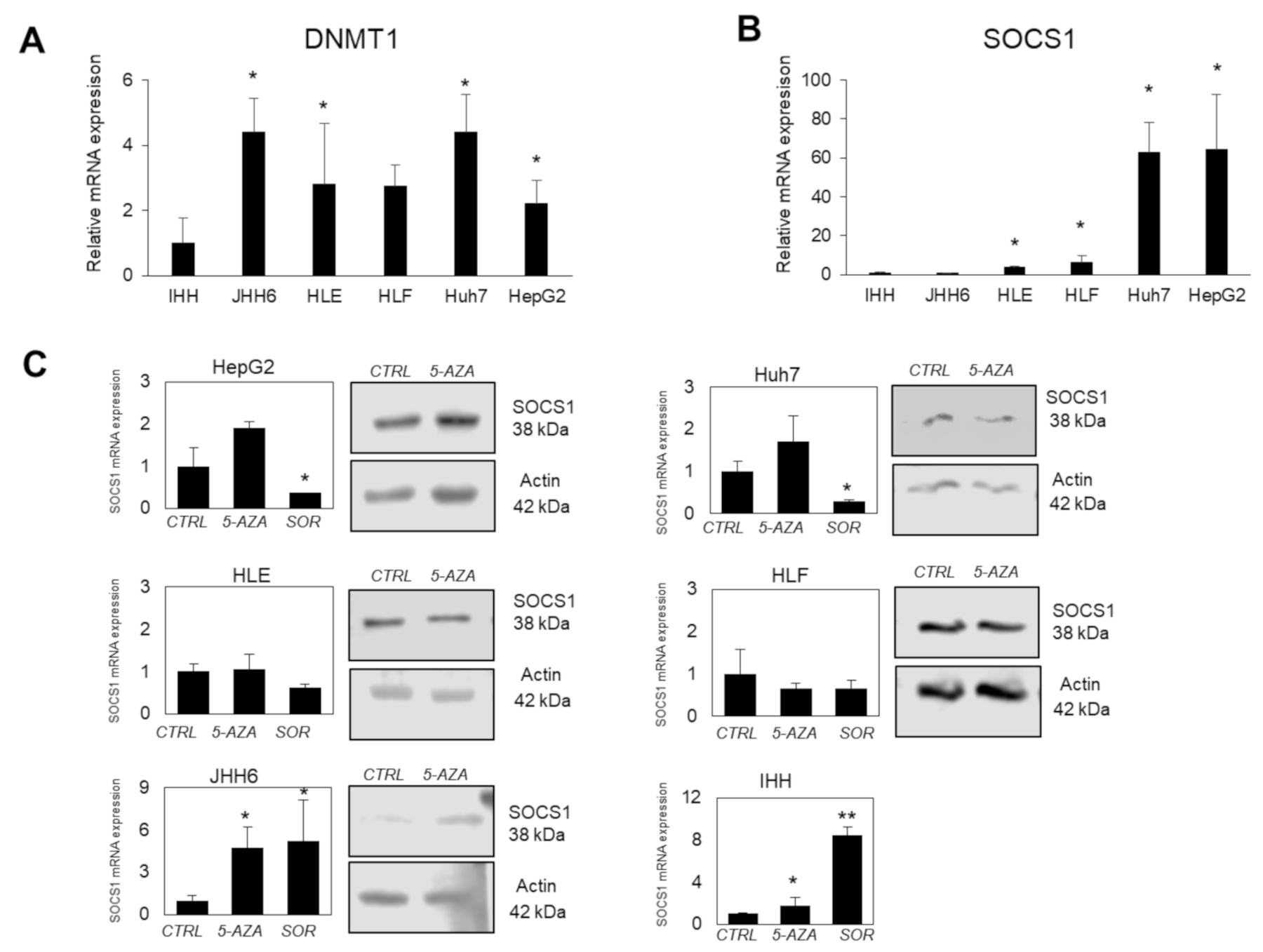
3.4. SOCS1 Modulation Following Demethylation by 5-AZA
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Sukowati, C.H.C. Heterogeneity of Hepatic Cancer Stem Cells. Adv. Exp. Med. Biol. 2019, 1139, 59–81. [Google Scholar] [CrossRef]
- Caruso, S.; O’Brien, D.R.; Cleary, S.P.; Roberts, L.R.; Zucman-Rossi, J. Genetics of Hepatocellular Carcinoma: Approaches to Explore Molecular Diversity. Hepatology 2021, 73, 14–26. [Google Scholar] [CrossRef]
- Boyault, S.; Rickman, D.S.; De Reyniès, A.; Balabaud, C.; Rebouissou, S.; Jeannot, E.; Hérault, A.; Saric, J.; Belghiti, J.; Franco, D.; et al. Transcriptome classification of HCC is related to gene alterations and to new therapeutic targets. Hepatology 2006, 45, 42–52. [Google Scholar] [CrossRef] [Green Version]
- Hoshida, Y.; Nijman, S.M.; Kobayashi, M.; Chan, J.A.; Brunet, J.-P.; Chiang, D.; Villanueva, A.; Newell, P.; Ikeda, K.; Hashimoto, M.; et al. Integrative Transcriptome Analysis Reveals Common Molecular Subclasses of Human Hepatocellular Carcinoma. Cancer Res. 2009, 69, 7385–7392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamashita, T.; Forgues, M.; Wang, W.; Kim, J.W.; Ye, Q.; Jia, H.; Budhu, A.; Zanetti, K.A.; Chen, Y.; Qin, L.-X.; et al. EpCAM and α-Fetoprotein Expression Defines Novel Prognostic Subtypes of Hepatocellular Carcinoma. Cancer Res. 2008, 68, 1451–1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wu, W. Chapter 5—DNA Methylation Alterations in Human Cancers. In Epigenetics in Human Disease, 2nd ed.; Tollefsbol, T.O., Ed.; Translational Epigenetics; Academic Press: San Diego, CA, USA, 2018; Volume 6, pp. 109–139. [Google Scholar]
- Pradhan, S.; Bacolla, A.; Wells, R.D.; Roberts, R. Recombinant Human DNA (Cytosine-5) Methyltransferase: I. Expression, Purification, and Comparison of de novo and Maintenance Methylation. J. Biol. Chem. 1999, 274, 33002–33010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, G.; Tu, Y.; Chen, C.; Sun, H.; Wan, C.; Cai, X. DNA methylation biomarkers for hepatocellular carcinoma. Cancer Cell Int. 2018, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Mah, W.-C.; Thurnherr, T.; Chow, P.K.H.; Chung, A.Y.F.; Ooi, L.L.P.J.; Toh, H.C.; Teh, B.T.; Saunthararajah, Y.; Lee, C.G.L. Methylation Profiles Reveal Distinct Subgroup of Hepatocellular Carcinoma Patients with Poor Prognosis. PLoS ONE 2014, 9, e104158. [Google Scholar] [CrossRef]
- Sun, N.; Zhang, J.; Zhang, C.; Shi, Y.; Zhao, B.; Jiao, A.; Chen, B. Using Illumina Infinium HumanMethylation 450K BeadChip to explore genome-wide DNA methylation profiles in a human hepatocellular carcinoma cell line. Mol. Med. Rep. 2018, 18, 4446–4456. [Google Scholar] [CrossRef]
- Liu, A.; Wu, Q.; Peng, D.; Ares, I.; Anadón, A.; Torres, B.L.; Martínez-Larrañaga, M.; Wang, X.; Martínez, M. A novel strategy for the diagnosis, prognosis, treatment, and chemoresistance of hepatocellular carcinoma: DNA methylation. Med. Res. Rev. 2020, 40, 1973–2018. [Google Scholar] [CrossRef] [PubMed]
- Kaminskas, E.; Farrell, A.; Abraham, S.; Baird, A.; Hsieh, L.-S.; Lee, S.-L.; Leighton, J.K.; Patel, H.; Rahman, A.; Sridhara, R.; et al. Approval Summary: Azacitidine for Treatment of Myelodysplastic Syndrome Subtypes. Clin. Cancer Res. 2005, 11, 3604–3608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, N.; George, T.L.; Otterson, G.A.; Verschraegen, C.; Wen, H.; Carbone, D.; Herman, J.; Bertino, E.M.; He, K. Advances in epigenetic therapeutics with focus on solid tumors. Clin. Epigenetics 2021, 13, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Silverman, L.R.; Demakos, E.P.; Peterson, B.L.; Kornblith, A.B.; Holland, J.C.; Odchimar-Reissig, R.; Stone, R.M.; Nelson, D.; Powell, B.L.; DeCastro, C.M.; et al. Randomized Controlled Trial of Azacitidine in Patients With the Myelodysplastic Syndrome: A Study of the Cancer and Leukemia Group B. J. Clin. Oncol. 2002, 20, 2429–2440. [Google Scholar] [CrossRef] [PubMed]
- Gnyszka, A.; Jastrzebski, Z.; Flis, S. DNA methyltransferase inhibitors and their emerging role in epigenetic therapy of cancer. Anticancer Res. 2013, 33, 2989–2996. [Google Scholar] [PubMed]
- Naka, T.; Narazaki, M.; Hirata, M.; Matsumoto, T.; Minamoto, S.; Aono, A.; Nishimoto, N.; Kajita, T.; Taga, T.; Yoshizaki, K.; et al. Structure and function of a new STAT-induced STAT inhibitor. Nat. Cell Biol. 1997, 387, 924–929. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, H.; Matsubara, K.; Qian, G.-S.; Jackson, E.P.; Groopman, J.D.; Manning, J.E.; Harris, C.C.; Herman, J.G. SOCS-1, a negative regulator of the JAK/STAT pathway, is silenced by methylation in human hepatocellular carcinoma and shows growth-suppression activity. Nat. Genet. 2001, 28, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Guo, M.; Herman, J.G.; Clark, D.P. Aberrant Promoter Methylation Profiles of Tumor Suppressor Genes in Hepatocellular Carcinoma. Am. J. Pathol. 2003, 163, 1101–1107. [Google Scholar] [CrossRef] [Green Version]
- Li, L.-C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 18, 1427–1431. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Wang, F.; Yang, L.; Guo, C.; Wan, R.; Ke, A.; Xu, L.; Hu, G.; Xu, X.; Shen, J.; et al. Expression of DNMT1 and DNMT3a Are Regulated by GLI1 in Human Pancreatic Cancer. PLOS ONE 2011, 6, e27684. [Google Scholar] [CrossRef] [Green Version]
- Toh, T.B.; Lim, J.J.; Chow, E.K. Epigenetics of hepatocellular carcinoma. Clin. Transl. Med. 2019, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.K.; Li, Y.; Pandit, H.; Li, S.; Pulliam, Z.; Zheng, Q.; Yu, Y.; Martin, R.C. Epigenetic modulation enhances immunotherapy for hepatocellular carcinoma. Cell. Immunol. 2019, 336, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Gailhouste, L.; Liew, L.C.; Yasukawa, K.; Hatada, I.; Tanaka, Y.; Nakagama, H.; Ochiya, T. Differentiation Therapy by Epigenetic Reconditioning Exerts Antitumor Effects on Liver Cancer Cells. Mol. Ther. 2018, 26, 1840–1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, H.-C.; Li, H.; Van Neste, L.; Cai, Y.; Robert, C.; Rassool, F.V.; Shin, J.J.; Harbom, K.M.; Beaty, R.; Pappou, E.; et al. Transient Low Doses of DNA-Demethylating Agents Exert Durable Antitumor Effects on Hematological and Epithelial Tumor Cells. Cancer Cell 2012, 21, 430–446. [Google Scholar] [CrossRef] [Green Version]
- Bagnyukova, T.V.; Tryndyak, V.P.; Muskhelishvili, L.; Ross, S.A.; Beland, F.A.; Pogribny, I.P. Epigenetic down-regulation of the suppressor of cytokine signaling 1 (Socs1) gene is associated with the STAT3 activation and development of hepatocellular carcinoma induced by methyl-deficiency in rats. Cell Cycle 2008, 7, 3202–3210. [Google Scholar] [CrossRef] [Green Version]
- Galm, O.; Yoshikawa, H.; Esteller, M.; Osieka, R.; Herman, J.G. SOCS-1, a negative regulator of cytokine signaling, is frequently silenced by methylation in multiple myeloma. Blood 2003, 101, 2784–2788. [Google Scholar] [CrossRef] [Green Version]
- Fujitake, S.; Hibi, K.; Okochi, O.; Kodera, Y.; Ito, K.; Akiyama, S.; Nakao, A. Aberrant methylation of SOCS-1 was observed in younger colorectal cancer patients. J. Gastroenterol. 2004, 39, 120–124. [Google Scholar] [CrossRef]
- Fukushima, N.; Sato, N.; Sahin, F.; Su, G.; Hruban, R.H.; Goggins, M. Aberrant methylation of suppressor of cytokine signalling-1 (SOCS-1) gene in pancreatic ductal neoplasms. Br. J. Cancer 2003, 89, 338–343. [Google Scholar] [CrossRef] [Green Version]
- Khan, G.M.; Ghosh, A.; Variya, B.; Santharam, M.A.; Ihsan, A.U.; Ramanathan, S.; Ilangumaran, S. Prognostic significance of SOCS1 and SOCS3 tumor suppressors and oncogenic signaling pathway genes in hepatocellular carcinoma. BMC Cancer 2020, 20, 1–18. [Google Scholar] [CrossRef]
- Zhao, R.-C.; Zhou, J.; He, J.-Y.; Wei, Y.-G.; Qin, Y.; Li, B. Aberrant promoter methylation of SOCS-1 gene may contribute to the pathogenesis of hepatocellular carcinoma: A meta-analysis. J. BU ON. Off. J. Balk. Union Oncol. 2016, 21, 142–151. [Google Scholar]
- Sanaei, M.; Kavoosi, F.; Esmi, Z. The Effect of 5-Aza-2′-Deoxycytidine in Combination to and in Comparison with Vorinostat on DNA Methyltransferases, Histone Deacetylase 1, Glutathione S-Transferase 1 and Suppressor of Cytokine Signaling 1 Genes Expression, Cell Growth Inhibition and Apoptotic Induction in Hepatocellular LCL-PI 11 Cell Line. Int. J. Hematol. Oncol. Stem Cell Res. 2020, 14, 45–55. [Google Scholar] [PubMed]
- Sanaei, M.; Kavoosi, F.; Pourahmadi, M. Effect of Decitabine (5-aza-2′-deoxycytidine, 5-aza-CdR) in Comparison with Vorinostat (Suberoylanilide Hydroxamic Acid, SAHA) on DNMT1, DNMT3a and DNMT3b, HDAC 1-3, SOCS 1, SOCS 3, JAK2, and STAT3 Gene Expression in Hepatocellular Carcinoma HLE and LCL-PI 11 Cell Lines. Asian Pac. J. Cancer Prev. 2021, 22, 2089–2098. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Li, J.; Huang, T.; Duan, S.; Dai, D.; Jiang, D.; Sui, X.; Li, D.; Chen, Y.; Ding, F.; et al. Meta-analysis of DNA methylation biomarkers in hepatocellular carcinoma. Oncotarget 2016, 7, 81255–81267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, P.-Y.; Yeh, C.-M.; Hsu, N.C.; Chang, Y.-S.; Chang, J.-G.; Yeh, K.-T. Epigenetic alteration of the SOCS1 gene in hepatocellular carcinoma. Swiss Med. Wkly. 2010, 140, w13065. [Google Scholar] [CrossRef]
- Llovet, J.M.; Ricci, S.; Mazzaferro, V.; Hilgard, P.; Gane, E.; Blanc, J.F.; De Oliveira, A.C.; Santoro, A.; Raoul, J.L.; Forner, A.; et al. Sorafenib in Advanced Hepatocellular Carcinoma. N. Engl. J. Med. 2008, 359, 378–390. [Google Scholar] [CrossRef]
- Li, Y.; Chen, G.; Han, Z.; Cheng, H.; Qiao, L.; Li, Y. IL-6/STAT3 Signaling Contributes to Sorafenib Resistance in Hepatocellular Carcinoma Through Targeting Cancer Stem Cells. OncoTargets Ther. 2020, ume 13, 9721–9730. [Google Scholar] [CrossRef]
- Meng, F.; Glaser, S.S.; Francis, H.; DeMorrow, S.; Han, Y.; Passarini, J.D.; Stokes, A.; Cleary, J.P.; Liu, X.; Venter, J.; et al. Functional analysis of microRNAs in human hepatocellular cancer stem cells. J. Cell. Mol. Med. 2011, 16, 160–173. [Google Scholar] [CrossRef]
- Murakami, K.; Sakukawa, R.; Ikeda, T.; Matsuura, T.; Hasumura, S.; Nagamori, S.; Yamada, Y.; Saiki, I. Invasiveness of Hepatocellular Carcinoma Cell Lines: Contribution of Membrane-Type 1 Matrix Metalloproteinase. Neoplasia 1999, 1, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Sukowati, C.H.C.; Anfuso, B.; Torre, G.; Francalanci, P.; Crocè, L.S.; Tiribelli, C. The Expression of CD90/Thy-1 in Hepatocellular Carcinoma: An In Vivo and In Vitro Study. PLOS ONE 2013, 8, e76830. [Google Scholar] [CrossRef]
- Venturelli, S.; Berger, A.; Weiland, T.; Essmann, F.; Waibel, M.; Nuebling, T.; Häcker, S.; Schenk, M.; Schulze-Osthoff, K.; Salih, H.R.; et al. Differential Induction of Apoptosis and Senescence by the DNA Methyltransferase Inhibitors 5-Azacytidine and 5-Aza-2′-Deoxycytidine in Solid Tumor Cells. Mol. Cancer Ther. 2013, 12, 2226–2236. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Target | Sequence F (5′ → 3′) | Sequence R (5′ → 3′) | Ref. |
|---|---|---|---|
| MS-PCR | |||
| SOCS1-methylated | ATGGTTTCGGGATTTACGAGT | TAACCACGATACGCTAACGAC | ts |
| SOCS1-unmethylated | AGATGGTTTTGGGATTTATGAGT | AACCACAATACACTAACAACA | ts |
| Gene Expression | |||
| ACTB | CGCCGCCAGCTCACCATG | CACGATGGAGGGGAAGACGG | ts |
| SOCS1 | CCCTTCCAGATTTGACCG | ATGGTTCCAGGCAAGTAA | ts |
| DNMT1 | CCATCAGGCATTCTACCA | CGTTCTCCTTGTCTTCTCT | [21] |
| SOCS1 Methylation in HCC Tissues | |||||
|---|---|---|---|---|---|
| M (%) | PM/UM (%) | All (%) | p | ||
| Sex [F:M] | 2:10 (17:83) | 2:9 (18:82) | 4:19 (17:83) | 0.9999 | |
| Age [year, mean ± std] | 70.9 ± 7.6 | 64 ± 6.9 | 67.6 ± 7.9 | 0.0177 * | |
| Tumor size [cm, mean ± std] | 4.9 ± 3.7 | 3.9 ± 2.1 | 4.4 ± 3.0 | 0.2399 | |
| AFP [median ng/mL, min–max] | 7.4 (2–5094) | 7.4 (3–139) | 7.4 (2–5094) | 0.1129 | |
| Etiology | HCV | 3 (25) | 6 (55) | 9 (39) | 0.2310 |
| HBV | 2 (17) | 0 (0) | 2 (9) | ||
| Metabolic | 6 (50) | 3 (27) | 9 (39) | ||
| no | 1 (8) | 2 (18) | 3 (13) | ||
| Histological grading | ES1 | 1 (9) | 6 (60) | 7 (33) | 0.0465 * |
| ES2 | 7 (64) | 3 (30) | 10 (48) | ||
| ES3-4 | 3 (27) | 1 (10) | 4 (19) | ||
| CTP | A | 10 (83) | 8 (73) | 18 (78) | 0.5379 |
| B-C | 2 (17) | 3 (27) | 5 (22) | ||
| BCLC | 0 | 8 (73) | 10 (91) | 18 (82) | 0.5865 |
| 1–2 | 3 (27) | 1 (9) | 4 (18) | ||
| Recurrence (m) | <12 m | 3 (75) | 6 (43) | 9 (50) | 0.5765 |
| >12 m | 1 (25) | 8 (67) | 9 (50) | ||
| Subtypes | Cell Line | EpCAM | CD133 | CD90 | CD24 | CD13 | CD45 |
|---|---|---|---|---|---|---|---|
| Hepatocytes | IHH | - | - | +/− | - | + | - |
| Stromal HCC | HLE | - | - | +/− | + | - | - |
| HLF | - | - | +/− | + | - | - | |
| JHH6 | - | - | - | - | +/− | - | |
| CSC HCC | HepG2 | + | - | - | - | +/− | - |
| Huh7 | + | + | - | + | +/− | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabral, L.K.D.; Reyes, P.A.C.; Crocè, L.S.; Tiribelli, C.; Sukowati, C.H.C. The Relevance of SOCS1 Methylation and Epigenetic Therapy in Diverse Cell Populations of Hepatocellular Carcinoma. Diagnostics 2021, 11, 1825. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11101825
Cabral LKD, Reyes PAC, Crocè LS, Tiribelli C, Sukowati CHC. The Relevance of SOCS1 Methylation and Epigenetic Therapy in Diverse Cell Populations of Hepatocellular Carcinoma. Diagnostics. 2021; 11(10):1825. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11101825
Chicago/Turabian StyleCabral, Loraine Kay D., Peter Andrew C. Reyes, Lory S. Crocè, Claudio Tiribelli, and Caecilia H. C. Sukowati. 2021. "The Relevance of SOCS1 Methylation and Epigenetic Therapy in Diverse Cell Populations of Hepatocellular Carcinoma" Diagnostics 11, no. 10: 1825. https://0-doi-org.brum.beds.ac.uk/10.3390/diagnostics11101825