Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications


US Environmental Protection Agency, Duluth, MN 55804, USA
Ecologies 2021, 2(1), 43-70; https://0-doi-org.brum.beds.ac.uk/10.3390/ecologies2010004
Submission received: 18 December 2020
/
Revised: 30 December 2020
/
Accepted: 4 January 2021
/
Published: 8 January 2021
(This article belongs to the Special Issue Feature Papers of Ecologies 2021)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Adaptative traits enable organisms to survive and reproduce. Though these traits are often innate features (ones that may or may not exhibit variability in response to environmental cues or originate from horizontal gene transfer), this is not always the case. Many species endure natural selection not with the traits they possess intrinsically but with exogenous substances and abilities that they acquire from other species, via ecological interactions akin to outsourcing, pillaging, and fraud. Here, I review the mechanisms of this exogenous trait acquisition and highlight some of their repercussions and usefulness for natural resource management, industry, and human health.
1. Introduction
Phylogenetic constraints on adaptation can lead to population decline and extinction in the face of natural selection [1]. Yet, in some cases, what species lack intrinsically, they can compensate for with the use of exogenous substances and abilities. For example, although unable to metabolize the toxins present in the fruits they eat (e.g., alkaloids such as strychnine and quinine), Peruvian blue-headed parrots (Pionus menstruus L.) successfully bind and expunge these toxins by ingesting clay that is rich in detoxifying minerals such as smectite and kaolin [2]. For ecologists, knowing the secret behind exogenous traits such as the parrots’ tolerance of fruit toxins can be important from the standpoint of natural resource management, as can understanding how these traits compare to endogenous ones in terms of their ecological costs and benefits and their sensitivities to environmental change (e.g., metabolic expression of the cytochrome P-450 gene in rats and other animals [3,4]). Such insights would clarify, for instance, how the parrots (and all the organisms the parrots interact with) might be impacted by the loss of access to suitable clay licks due to urban and agricultural development, acid rain, and soil erosion [5,6,7]. In ecology, data regarding species traits are used not only to determine species susceptibility to toxic exposures and over-exploitation [8,9] but also to interpret species distributions across space and time [10,11,12] and anticipate population declines [13]. They provide the means to assess the “functional redundancy” of ecological communities (the extent to which different species contribute similarly to ecological processes such as nitrogen fixation or the transfer of carbon and energy from one level of a food chain to another), which is often a reliable indicator of the communities’ “resilience” (ability to remain healthy and recognizable following disastrous perturbations such as oil spills and invasive species introductions; [14,15,16].
Species can acquire exogenous traits not only from their abiotic environment, as in the above example of the parrots, but also from other species. One could argue that this includes via genetic exchange mechanisms such as horizontal gene transfer [17]. However, once foreign genes are assimilated into a species’ genome and expressed, the traits they produce are, for all intents and purposes, intrinsic (endogenous). Similarly, inbuilt traits that exhibit context-dependent variability, such as “developmental” or “inducible” traits [18,19], are phenotypic expressions of the species (genotypes) that utilize them and, therefore, still endogenous. Truer examples of species acquiring exogenous traits from other species are ones that are analogous to outsourcing (or to pillaging and fraud, depending on the equity and intensity of the ecological exchanges involved). Just as relying on vendors or third-party experts for outsourced goods and services might enable financial institutions to reduce financial costs, achieve tasks that cannot be completed internally, and focus on core business activities [20], relying on other species for exogenous traits can enable organisms to reduce metabolic expenditure, overcome phylogenetic constraints, and undergo adaptive specialization [21,22].
Myriad community ecological interactions can enable species to acquire exogenous traits from other species, ranging from protracted, intimate, mutually beneficial relationships to ones that are transient and one-sidedly exploitative. Some of these have been thoroughly investigated, while others are lesser known. Here, I provide illustrative examples of these interactions that reveal both the ubiquity of the acquisition of exogenous traits from other species and its ability to generate lasting ecological effects in diverse environments. Furthermore, I highlight some of the implications of exogenous trait acquisition for natural resource management, industry, and human health and discuss how it has been (or might in future be) incorporated into the theory and practice of ecology and evolution.
2. Community Ecological Interactions Involving Exogenous Trait Acquisition
2.1. Competition
Competitive interactions underly some of the most pressing global environmental issues of our time, including the spread of invasive species [23] and the formation of harmful algal blooms [24]. They are defined as interactions in which competitors (organisms that utilize the same finite resources, which might include growth-limiting nutrients) harm one another. Generally, competitive interactions fall within one or the other of two main categories: “exploitative competition” and “interference”. In exploitative competition, competitors harm each other merely through their utilization of the resource itself (one competitor cannot use what the other has already consumed). In interference, competitors employ direct assaults.
Despite the well-established ecological and evolutionary importance of exploitative competition [25,26], I have chosen to omit examples of this category of competition from my review, with the rationale that it would be challenging and perhaps meaningless to attempt to qualify the assertion that a competitively superior species’ exogenous traits are what enable it to assimilate more resources (all adaptive traits, regardless of their actual function, must, by definition, ultimately sustain or increase the ability of a species to assimilate resources over time—at the population level, if not the individual level). Instead, my focus in this section is on the modes of interference. These modes include the following, which I define in the ensuing sections: (1) “preemption”, (2) “territoriality”, (3) “overgrowth”, and (4) “allelopathy” (Figure 1) [27]. As I touch on below, species can rely on interspecific associations providing exogenous traits to engage in each of these four modes of interference “indirectly”.
2.1.1. Indirect Preemption
Preemption is the blockage of access to resources via occupation of space or creation of barriers (à la the children’s game Musical Chairs [32]), which may include altering the nature of the resource so that it is unsuitable for or inaccessible to competitors. This is the strategy employed by both Wolbachia (a genus of endosymbiotic bacteria) and dengue virus (the cause of dengue fever). Each of these pathogens is able to manipulate their host, the yellow-fever mosquito (Aedes aegypti L.), in various ways. When one of them alters the regulation of the host’s DNA methyltransferase gene (causing it to be suppressed, in the case of Wolbachia, and to be overexpressed, in the case of dengue virus), it inhibits the other’s ability to replicate within the host [33]. Bacteria in the genus Enterobacter engage in a similar form of indirect preemption. Upon infecting the finger millet (Eleusine coracana (L.) Gaertn.), these bacteria disrupt cell membranes and cause the formation of multilayer physical barriers inside their host that limit the infection rate and growth of fungal competitors such as Fusarium graminearum (a.k.a. Gibberella zeae (Schwein.) Petch) [34,35]. Investigation of the mechanisms underlying such dynamics (e.g., modes of signaling between host and pathogen cells) might reveal ways to evaluate and mitigate the risks to humans, wildlife, and crops associated with coinfecting pathogens [36,37]. Additionally, since A. aegypti is a known vector of disease in humans and other animals [38], deeper understanding of when and how pathogens employ exogenous barricades within this particular host to exclude competitors could lead to human and animal health-related applications that complement existing approaches to modifying vectors to control disease [39].
Another way that exogenous trait acquisition can mediate preemption is by providing species on the receiving end of preemption with a countermeasure. Cyanobacteria of the genus Dolichospermum (formerly Anabaena) are thought to be at a disadvantage against competing cyanobacteria that are more motile or able to regulate their buoyancy, since these traits enable the competitors to gain better access to sunlight and nutrients within the water column and to evade certain kinds of grazers [40,41]. However, Dolichospermum spp. can compensate for their lack of motility and buoyancy regulation via symbioses with ciliates of the genus Vorticella, from which they gain sufficient mobility to appear as if their colonies are swimming (and potentially increase their rate of nutrient sequestration due to the ciliates’ vacuum-cleaner-like mode of feeding) [42,43]. Since Dolichospermum species are common constituents of harmful algal blooms worldwide [44], it may be worthwhile to explore the extent to which their exogenous mobility enables them to proliferate at the expense of less pestilent competitors.
2.1.2. Indirect Territoriality
Territoriality is the staking of claims via aggressive behavior and use of warning signals such as scent markings [45]. Cases of territoriality made possible (or enhanced) by acquisition of exogenous traits include that which is seen in mixed-species groups of tamarin monkeys (Saguinus spp.). These groups are often more successful in repelling intruders from their joint territories than monospecific groups, due not only to the former often comprising more individuals but also to differences among species in body size, sensory perception, aggressiveness, and timings of daily activities [46,47]. The same has been found to be true for mixed-species flocks of neotropical forest understory birds [48]. While it is realistic to expect there to be trade-offs and socially enforced deterrents to “cheating” (lack of reciprocity and fidelity) among the interspecific relationships that allow for indirect territoriality [49,50], this does not mean that the relationships must be mutualistic or commensal [51,52]. Regardless, since territoriality often evolves where there is scarcity of resources [53,54], species loss in these systems (e.g., due to poaching and deforestation) may give rise to cascades of extinction, as resources become even scarcer and the ability to rely on the traits of other species to secure these resources diminishes. The prospect of such dependency-exacerbated extinctions has been evaluated in the context of plant-pollinator networks [55,56] but has received little attention in the case of other similar forms of association.
Acquisition of exogenous traits can also play a role in the cases of territoriality that involve scent marking and other demarcation behaviors. Scent marking in adult meerkats (Suricata suricatta Schreber), for example, is supported by bacteria obtained from the meerkats’ immediate environment/territory. Meerkats whose glandular pastes have amassed similar bacteria are able to showcase similar smells, whereas the pastes of outsiders and intruders smell different to meerkats and elicit different responses (generally, aggressive territorial defense behaviors) [57]. There is presently little by way of theory to explain why the bacterial communities of one meerkat habitat should be easily distinguishable from those of a neighboring meerkat habitat, a knowledge gap that may owe something to the popularity of the paradigm that bacteria and other microbes are capable of virtually unlimited dispersal and gene flow (“everything is everywhere”) [58,59]. Moreover, for the purposes of meerkat conservation, there may be a need, in certain areas, to consider how meerkat territoriality and mating success might be affected by the presence of antibiotics and other microbe-impacting contaminants in the environment [60,61].
Most examples of territoriality found in nature are directed not at other species but at conspecifics (members of the same species). Exogenous trait acquisition can play a role even in the cases tied to courtship and contest for mates. For example, there are various animals whose symbionts trigger the release of aphrodisiac scents or induce cannibalism [62,63] and animals that incorporate prey or once-living ornaments into their courtship rituals [64,65]. These animals, like the others described above, may, by virtue of their reliance on other species, be indirectly susceptible to environmental conditions that alter the size or structure of their associates’ populations.
2.1.3. Indirect Overgrowth
Overgrowth is the physical ouster of competitors from spaces where resources can be obtained [66]. A case where acquiring exogenous traits enables organisms to overgrow their competitors is seen in kelp forests, wherein red algae out-compete other species of phytoplankton in their vicinity via the capabilities of their piscine symbiont, the garibaldi (Hypsypops rubicundus (Girard) Gill). As part of a set of ritualized behaviors used to attract females, male garibaldis build nests, grooming and tending a rocky surface for several days in preparation for spawning. In so doing, the males remove all the invertebrates and algae from the chosen surface, leaving only a ring of red algae around the groomed site. The sparser the ring of red algae surrounding this surface, the less likely the males are to successfully court passing females; thus, the males generally attempt to maintain the cover of red algae in high abundance while weeding out all other phytoplankton from the vicinity [67]. It is possible that the garibaldis’ behavior might inhibit the spread of invasive species, such as lacy crust bryozoans (Membranipora membranacea L.) [68], just as grazing by other species of fish (e.g., the senorita, Oxyjulis californica Günther) and by nudibranchs (e.g., Polycera quadrilineata Müller and Onchidoris muricata Müller) has been shown to do in other locations [69,70]. Alternatively, the mutualistic interaction might enable the red algae themselves to become invasive or cause other problems, as similar mutualistic interactions are thought to have done in other systems [71]. To my knowledge, no study has explored these possibilities.
Certain virulent food-borne microbial pathogens employ the strategy of indirect overgrowth, as well, via the capabilities of a host. From the pathogens’ perspective, the surfaces of the host’s intestines are complex habitats that are often already occupied by billions of resident species. Invading and establishing within these habitats can be a tremendous challenge. In fact, the immense resident diversity of a host’s intestines may well aid the host in combating invasive pathogens [72]. Pathogens such as Salmonella enterica serovar Typhimurium, however, are able to overcome this obstacle by inducing a novel immune response to which they have superior resistance. As their resident competitors are rarely preadapted to this immune response, the pathogens are able to eliminate and supplant these competitors in one fell swoop [73,74]. Whereas other strains of S. enterica are known for causing typhoid fever, S. enterica serovar Typhimurium is among the few emerging invasive strains of non-typhoidal Salmonella, as a direct result of this indirect overgrowth strategy [75].
Other kinds of endosymbiotic microbes (besides pathogens) that engage in overgrowth via the capabilities of their hosts include the obligate bacterial endosymbionts of the marine sponge Aplysina aerophoba Nardo. The sponge is a selective particle-feeder that draws these bacteria in from its environment and houses them (often in large, densely packed populations) in its “mesohyl” (an acellular gel layer between the epidermis and the inner cavity known as the “spongocoel”). Certain physical and chemical properties of these bacteria signal the sponge to retain them as endosymbionts, even while it proceeds to digest the bulk of the endosymbionts’ microbial competitors [76]. A. aerophoba has been used as an indicator of aquaculture-and sewage-related environmental impacts [77], wherein its responses to factors such as dissolved nitrogen are often a function of its endosymbionts’ metabolic activities and requirements [78].
Free-living, macroscopic organisms also rely on other species’ differential feeding capabilities to engage in overgrowth. Two such organisms are the striped poison-fang blenny (Meiacanthus grammistes Valenciennes) and the white-tailed deer (Odocoileus virginianus Zimmermann). The blenny relies on potential predators, which it positions to make short work of competing fish by envenomating these competitors with substances that disorient them and make it difficult for them to coordinate movements such as swimming [79]. In contrast, the deer relies on the parasitic meningeal helminth Parelaphostrongylus tenuis Dougherty. Although the deer itself is mostly unaffected by this helminth, the helminth is potentially fatal to moose (Alces alces (L.) Gray) and other cervids. This parasite has caused widespread population declines in areas where white-tailed deer have invaded the ranges of other cervids. The majority of attempts to reintroduce other cervids to ranges now occupied by infected white-tailed deer have failed [80]. Some evidence suggests that the phenomenon of competitors engaging in indirect overgrowth through use of parasites may be widespread and may exhibit consistent patterns across various habitats, an idea that has been termed the “geographic-range hypothesis” [81,82].
2.1.4. Indirect Allelopathy
Allelopathy is the use of chemical weaponry (poison) against competitors [27]. It was first discovered and is best known in plants [83]. Some plants, however, have essentially evolved biological warfare in place of chemical warfare, launching pathogens and symbiont-derived banes against their competitors in lieu of or in combination with endogenous poison. The tropical invasive weed, Chromolaena odorata (L.) King and Rob., for instance, has been found to inhibit the growth of numerous native and naturalized competing plant species by preferentially stimulating the growth of the fungus Fusarium semitectum (Desm.) Sacc., a generalist plant pathogen (to which C. odorata is mostly immune), in the soil of its rhizosphere [84]. Another invasive sessile organism, the bryozoan Celleporaria brunnea Hincks, employs a similar form of indirect allelopathy. It provides hydroids of the genus Zanclea with a scaffold on which to form polyps and protects these polyps with skeletal material. In return, the hydroids enable the bryozoan to outgrow adjacent competitors by stinging these competitors into a weakened state [85]. The spread of C. odorata and C. brunnea has not only resulted in losses of biodiversity and ecosystem function but also, respectively, caused damage to commercial plantations and boats [86,87]. Insights into how these invasive species initiate and coordinate their activities with the species from which they acquire their indirect allelopathy may create opportunities for mitigating their effects.
Organisms need not be sessile to employ indirect allelopathy. For example, fungus-growing ants (Formicidae: Attini) employ indirect allelopathy when competing with microfungal parasites of the genus Escovopsis. These microfungal parasites occupy the (ant-processed) leafy substrate intended for a fungal cultivar that serves as the ants’ primary food source, and they infect this fungal cultivar when it is present. The ants rid their fungal gardens of these parasites by relying on a streptomycete bacterium that inhabits their cuticles and produces a potent antibiotic [88]. Similar adaptations have evolved in other fungus-farming insects, such as termites (Isoptera) [89] and ambrosia beetles (Ambrosiophilus spp.) [90]. The ambrosia beetles, however, instead of relying on the antibiotics of mutualistic endosymbionts, utilize ethanol produced by the weakened trees that the beetles exploit as beds for their fungal gardens [91]. Deeper understanding of the workings of such allelopathy-conferring associations might aid in the prevention of outbreaks of diseases caused by the organisms that employ the allelopathy (e.g., Dutch Elm Disease and Laurel Wilt Disease) [92,93].
Other motile organisms that engage in indirect allelopathy can be found among protozoa. Upon assimilating the phage-like bacterial endosymbionts of the genus Caedibacter (previously known as “kappa bodies”), certain ciliates of the genus Paramecium transform into “killer cells” capable of paralyzing and disintegrating closely related Paramecium species that do not possess the symbiont [94]. The kappa bodies produce paramecin, a toxin that is lethal to various Paramecium species, and seems to release the toxin into the surrounding environment while protecting the host from the toxin’s effects (possibly by modifying the host’s cell membrane, though this has yet to be determined) [95]. Additionally, Chen (1955) and Müller et al. (2011) have reported evidence to suggest that a Paramecium species that does not typically harbor Caedibacter, P. bursaria Ehrenberg, is able to utilize toxic compounds produced by its facultative microalgal symbiont Chlorella vulgaris Beijerinck in its competitive interactions with other Paramecium species and with fellow bacterivorous ciliates Colpidium striatum (Losana) Ganner and Foissner and Tetrahymena pyriformis [96,97]. It would be a challenging task to determine how susceptible the cells of Caedibacter- and Chlorella-housing paramecia are to one another’s symbiont-derived weaponry, but it also would be intriguing in its own right (as an opportunity to observe a sort of “rivals’ match”) and provide insights into the mechanics of susceptibility and endosymbiosis in eukaryotes.
2.2. Mutualism
Mutualistic interactions are those in which organisms engage in mutually beneficial exchanges with other species that cause either no harm or negligible/outweighed harm to the participants [98,99,100]. The various benefits they can provide include nutritional benefits [101], dispersal benefits [102], and reproductive benefits [103]. Mutualistic interactions need not be symbioses, but many of the most famous and arguably most consequential mutualistic interactions are symbioses. For example, all known eukaryotic organisms are thought to have originated from once free-living bacteria and archaea that became endosymbionts. Exchanging genetic material and optimizing divisions of labor, they became inseparable, interdependent, and able to perform tasks and tolerate conditions that their free-living ancestors could not have performed and tolerated (a process referred to by some as “symbiogenesis”) [104,105,106]. Similarly, if not for the mutualistic interactions of mycorrhizal fungi and plants, terrestrial life on Earth might not have been possible [107,108].
Other examples of mutualistic interaction are less far-reaching and recognizable but no less fascinating and potentially useful. One that was initially described more than a century ago but is still poorly understood and considered extremely unique among animals is that of the symbiosis between the spotted salamander (Ambystoma maculatum Shaw) and the green algae that populate the salamander’s eggs. Studies have shown that salamander embryos benefit from the increased oxygen concentrations that their algal symbionts produce through photosynthesis and that the algae, in turn, may benefit from ammonia excreted by the embryos. More recently, it has been shown that the algae do not simply grow within the egg capsule (outer shell of the eggs) but actually penetrate salamander embryonic tissues [109] (Figure 2A). Apart from providing insights into the ecology, evolution, and ontogeny of each of the species involved (and perhaps the processes of ecology, evolution, and ontogeny in general), closer examination of this relationship might allow humans to advance certain cutting-edge experimental medical techniques, such as the use of cyanobacteria to treat coronary artery disease [110] or the use of marine algae-derived bone-forming material for grafting [111].
When mutualistic interactions happen to enhance one or both partners’ ability to respond to the threat of natural enemies, it is easy to see how the acquisition of exogenous traits might impact consumer–resource dynamics. This, however, I will address in a later section. Here, I restrict my focus to showing how the acquisition of exogenous traits from other species can be the basis of mutualistic synergisms that enable species to tolerate and respond to abiotic stress.
2.2.1. Neighborhood Habitat Amelioration (“Adversity Makes Strange Bedfellows”)
Neighborhood habitat amelioration (a.k.a. “bidirectional facilitation”) is where organisms that would otherwise compete strongly with one another have net positive effects on one another, instead [100,112,113]. Two conditions must be met for this to occur: (1) both organisms must be simultaneously exposed to one or more abiotic stresses they cannot cope with on their own (not necessarily ones to which they both share susceptibility), and (2) each must possess traits that either enable the other to cope with its respective stresses or that combine with the other’s traits to create emergent coping mechanisms for both. In other words, the organisms must “make a good team” (functionally complement one another) under conditions of abiotic stress. For example, certain plants cannot simultaneously acclimatize to the stresses of shade and drought due to physiological trade-offs. Yet, paradoxically, in dry areas, these same plants, as seedlings, are often found to be restricted to shady sites under the canopy of other, already established plants. Holmgren et al. (1997) determined that the effects of the canopy and the plants’ combined root structures create microhabitat conditions of sufficient water retention/availability for the plants to survive, turning what would have otherwise been water-monopolizing competitors for the seedlings into life-saving “nurse plants”. For their part, the nurse plants benefit from the greater volume and more consistent accessibility of water, as well [114].
Multispecies biofilms are another form of symbiosis that might entail or promote neighborhood habitat amelioration. These non-randomly structured, spatially defined communities of bacteria and other microbes are omnipresent in natural environments and often found in clinical settings. Some have argued that biofilms provide important insights into the evolution of multicellularity and could be considered multicellular organisms in their own right (or, at least, “pseudo-” or “proto-multicellular” organisms) [115,116]. Biofilm constituents may overlap significantly in their resource requirements, which would make them intense competitors under other circumstances. Yet, under the conditions that trigger biofilm formation and persistence, interactions among the constituent species often lead to emergent properties that benefit them all, such as resistance to antibiotics, heavy metals, and host immune responses [117,118]. In clinical settings and in the food industry, these acquired exogenous benefits can be deadly [119,120] and could perhaps be effectively dealt with using applied community ecological principles [121].
2.2.2. Association-Derived Heat and Cold Tolerance
Certain organisms can acquire increased tolerance of extreme heat through the manipulations of their endosymbionts. The cucumber (Cucumis sativus L.), for example, grows better at high temperatures when it houses the endophytic fungus Paecilomyces formosus LHL10 than when it does not, possibly due to the endosymbiont facilitating heat shock protein expression and down-regulating certain endogenous cucumber hormones [122,123]. Other strains of P. formosus have been found to be pathogenic on plants such as pistachio (Pistacia vera L.) [124], but the exogenous heat-tolerance conferred by this particular strain could be applied beneficially where heat stress among crops is a concern (e.g., where cucumbers are double-cropped with tomatoes under black mulch [125]). On the opposite side of the temperature spectrum are organisms that rely on endosymbionts to enhance their cold tolerance. This includes the deer tick (Ixodes scapularis Say), host to Anaplasma phagocytophilum Foggie, the bacterial cause of human granulocytic anaplasmosis. The bacterium increases the ability of its host to survive the harsh winters of the northeastern and upper midwestern United States by causing the tick to produce an antifreeze glycoprotein [126]. This exogenous cold tolerance has a bearing on the prevalence of human granulocytic anaplasmosis in these areas and may also affect that of other tick-borne diseases such as Lyme Disease (caused by the bacterium Borrelia burgdorferi Burgdorfer), depending on the as-yet poorly understood dynamics of coinfection in tick and human hosts [127].
Another way that organisms might rely on the traits of other species for help in regulating their temperature is by engaging in or modifying social behaviors (this is referred to as “social thermoregulation”). One such behavior is huddling for warmth in communal groups [128]. Though the benefit of thermoregulation is not necessarily the main reason, such communal groups are frequently heterospecific, as seen in birds, bats, flying squirrels, mongooses, and slugs [129,130,131,132,133]. On the cooling side of social thermoregulation, one finds behaviors such as fanning in mixed-species bee colonies (Apis spp.) [134] and tree-hugging in the koala Phascolarctos cinereus Goldfuss (where the koala seeks out and presses its body against species of trees capable of retaining cooler surfaces than others during heat waves) [135]. At present, it is unclear precisely how important these behavior-based exogenous benefits are to the recipients’ survival and reproduction or how susceptible they are to the influences of human activity and species loss, but the fact that these species routinely utilize them is perhaps telling.
2.2.3. Cooperative Shelter Building
Physiological responses, even when they are (as in the examples above) derived from another species, might provide inadequate or prohibitively costly protection against the elements. A potential solution in this scenario is to build shelter. Mutualistic interactions can provide non-human architects with the equivalent of “construction co-workers” and “building material suppliers”. For instance, Asiatic honeybees (Apis cerana Fabricius) and European honeybees (Apis mellifera L.) have been shown to cooperate in comb building [136]. In another bee-related example, the cadaghi (Corymbia torelliana (Mueller) Hill and Johnson, an Australian eucalypt) supplies the sugarbag bee (Tetragonula carbonaria Smith) with its eucalypt resin in return for seed dispersal benefits. This resin is not only a source of nutrients and energy but also an adaptable building material, with which the bee can construct and waterproof its signature spiral hive and fashion sticky traps to prevent the infiltration of its hive by ants and other would-be intruders [137] (Figure 2B). It is yet to be determined how vulnerable these relationships are to the global spread of infectious disease pathogens such as Israeli Acute Paralysis Virus [138] and the fungal species within the genus Quambalaria that infect Corymbia spp. [139] or whether these relationships can be manipulated to control these pathogens.
2.3. Consumer–Resource Interactions
Consumer–resource interactions can be viewed as a battle of opposing forces: defenses versus offenses. Defenses are traits used to avoid, deter, or eliminate consumers (natural enemies such as predators and parasites), whereas offenses are traits used to acquire, secure, or assimilate resources (prey or hosts). The armor-like scales of the arapaima are an endogenous defense [140] (Figure 3A), while the gun-like tongue and gills of the archer fish are an endogenous offense [141] (Figure 3B). Also endogenous, despite their effectiveness being dependent on the co-occurrence or effect of another species, are most forms of Batesian and aggressive mimicry (wherein the success of the mimic depends on the proximity or past evolutionary influence of its model) [142,143] and many cases of “inducible” defense and offense (phenotypic transformations in organisms triggered by the cues of natural enemies or resources, respectively) [144,145].
Distinct from the above-mentioned endogenous defenses and offenses are exogenous defenses and offenses that are obtained from other species. Hereafter, I employ the term “associational defenses” (which is already in common use [148,149]) to refer to the defensive exogenous traits and term the offensive exogenous traits “associational offenses”.
2.3.1. Associational Defenses (“The Enemy of My Enemy Is My Ally”)
Associational defenses, as a group, have been the centerpiece of numerous studies addressing globally important topics such as climate change [150,151] and ecological succession [152,153,154]. They have occasionally been incorporated into general mathematical models of predation and competition [155,156,157]. Authors such as Sabelis et al. (2001) and Hay et al. (2004) have tied them to the popular adage “the enemy of my enemy is my ally”, thus bestowing on them, simultaneously, a touch of familiarity and air of mystique [158,159]. Many associational defenses have long been well-known. Some, such as the various toxins that lepidopterans sequester from poisonous plants [160,161] and the formic acid that birds procure via “anting” [162], are acquired in ways that harm the recipients’ benefactors. Others originate from associations that could rightly be categorized as mutualisms, including the improved awareness and capacity for vigilance that organisms gain from dwelling and foraging in heterospecific groups [163,164,165], the ectoparasite-dislodging services that fish and terrestrial animals solicit from cleaning symbionts [166,167], and the herbivore-repelling behaviors that terrestrial and aquatic plants elicit in sympatric predators and parasitoids [159,168,169].
The above example of plants relying on predators and parasitoids to repel herbivores is often mediated by signaling cues such as the volatile organic compounds that the plants release in response to herbivory which betray the location of the herbivores to the herbivores’ predators and parasitoids [170]. A parallel to this mechanism exists in oceans, wherein marine dinoflagellates respond to their nocturnal predators by emitting flashes of blue light that draw the attention of larger predators [171]. At the molecular level, both plant volatiles and dinoflagellate bioluminescence (production of light) are byproducts of essential metabolic processes. The former are still commonly referred to as “secondary metabolites” [172], and the latter appears to stem from photo-oxidation of chlorophyll [173]. Discovery of the compounds and pathways involved in these metabolic processes has given rise to practical applications, such as use in agricultural pest management [174], in the cases where the associational defenses and their underlying mechanisms are already known.
Other examples of organisms acquiring associational defenses involve less active recruitment than those described above, arising, instead, from intimate and protracted symbioses. The source of the toxicity of marine organisms such as the pufferfish (Arothron hispidus L.), for instance, is endosymbiotic bacteria within the genus Vibrio [146] (Figure 3C). Marine isopods [175], harvestman [176], jewel box clams [177], crabs [178], and (possibly) three-toed sloths [179] acquire camouflage and other mechanical and chemical defenses from ectosymbiotic algae, cyanobacteria, and sponges. There are also defense-conferring symbioses that represent a sort of middle ground between recruited association and near-complete fusion, including those of “myrmecophytes” (plants that bear specialized structures to support colonies of “bodyguard” ants) [180,181,182] and those of gobies with burrowing shrimp (wherein the shrimp permit the gobies to share their dens in return for “sentry duty”) [183]. Such symbiosis-derived associational defenses can also be directed at parasites, as is the case with the mutualistic symbiosis that New Zealand cockles (Austrovenus stutchburyi Wood) share with a fellow intertidal bivalve, the mud flat anemone (Anthopleura aureoradiata Carlgren). The anemone relies on the cockles’ shells as requisite hard substrates for attachment and prevents the cockles from being infected by the trematode Curtuteria australis Allison [184].
2.3.2. Associational Offenses (“Enemies Can Have Allies, Too”)
If associational defenses should inspire us to recall that “the enemy of my enemy is my ally”, then associational offenses should prompt us to ask, “What if my enemy has allies, too?” Unlike their defensive counterparts, associational offenses have been reported sparsely and often anecdotally. One could argue that some cases are well represented (or, at least, reasonably approximated) in models of phenomena such as parasite-mediated trophic transmission [185,186], vector-pathogen mutualism [187], cooperative hunting [188,189], and indirect effects on predators by mutualists [156]. However, there have been few, if any, deliberate attempts to incorporate associational offenses as a group into ecological and evolutionary theory. As such, for the purpose of introduction, I provide, below, an overview of the phenomenon slightly more detailed than that which I have provided for the other exogenous traits in this review. While not an exhaustive listing, the examples I include are intended to represent the remarkable diversity of associational offenses and associational offense users found in nature.
Just as competitors engage in indirect allelopathy and prey or hosts defend themselves using exogenous toxins, various consumers have been found to subdue their prey or hosts using exogenous venom. Among these are blue-ringed octopuses [147] (Figure 3D), entomophagous nematodes [190], leaf-chewing insects [191], and parasitoid insects [192]. Each of these consumers relies on microbial endosymbionts to create, supplement, or serve as the deadly substance. Other consumers depend on microbial endosymbionts for the opposite purpose: neutralization. These include specialist herbivores such as the cigarette beetle (Lasioderma serricorne Fabricius) [193] and the greater sage-grouse (Centrocercus urophasianus Bonaparte) [194], which use the enzymes of their respective gut microbiota (as opposed to the clay or the cytochrome P-450 genes mentioned in the Introduction) to break down plant chemical defenses.
Numerous aquatic predators possess microbial endosymbionts that provide associational offenses in the form of bioluminescence. The anglerfish (Melanocetus murrayi Günther) uses this bioluminescence to lure its prey [195]. As does the cookie-cutter shark (Isistius brasiliensis Quoy and Gaimard), though in the case of the shark, the prey are often larger predators than itself [196]. The Hawaiian bobtail squid (Euprymna scolopes Berry) uses bioluminescence to create disruptive color patterns, which enable it to ambush prey from above by breaking up its silhouette and eliminating its shadow [197]. The loose-jawed dragonfish (Malacosteus niger Ayres) uses bioluminescence (in its case, a unique far-red frequency) to detect (illuminate) its prey without being detected [198,199].
Aside from substances and bodily attributes, associational offenses can also take the form of services (behaviors). For example, some foraging animals routinely risk or forfeit nutritional and energetic rewards to retain the scouting and flushing services of other species [200,201,202,203]. Among these are giant moray eels (Gymnothorax javanicus Bleeker), which rely on coral groupers (Plectropomus pessuliferus Fowler) to alert them and guide them to the location of small fish hiding in narrow crevices beyond the groupers’ reach [202]. Interspecific cooperative foraging relationships such as these can entail surprisingly sophisticated displays of communication and coordinated mutual aid [204,205]. In addition, consumers with limited endogenous motility or speed compared to their prey or hosts often rely on phoretic partners to come within range of these prey or hosts, as is the case with mesostigmatid mites (Poecilochirus necrophori Canestrini and Canestrini) [206], fig nematodes (Schistonchus spp. and Parasitodiplogaster spp.) [207], and kangaroo leeches (Marsupiobdella africana) [208]. Arguably, such phoresy-utilizing consumers could also include all known vector-transmitted pathogens, especially those that actively recruit their vectors via manipulations of their hosts (e.g., malarial plasmodia and Dutch elm disease fungi manipulating host odor to attract mosquitos and bark beetles, respectively [209,210]).
Even the plant kingdom, despite being mostly photoautotrophic, houses some examples of associational offense users. Among these are holoparasites such as Cytinus hypocistis L., which commandeers the hyphae of its host’s mycorrhizal fungi to increase its efficiency in appropriating host nutrients [211], and carnivorous plants such as those of the genus Roridula, which obtain nutrients from insects trapped in their sticky leaves by relying on the digestive enzymes of the predatory hemipteran, Pameridea roridulae Reuter [212]. The Roridula-hempiteran symbiosis is, in many ways, the associational offense counterpart of myrmecophyte-ant symbioses (complete with evidence of intense coevolution, some of which I touch upon in the next section). In addition, like those of associational defense users, the ranks of associational offense users include organisms that actively maim or kill their associates to acquire the victims’ traits. Among these are blanket octopuses (Tremoctopus violaceus Chiaje, which can use the severed stinging tentacles of its prey to subdue additional prey) [213], aphid lions (which encase themselves in the shed exoskeletons and dead bodies of their prey to infiltrate nests and ambush more prey) [214], green-backed herons (which capture insects and other small organisms to use as bait for catching fish) [215], dolphins (which fit marine sponges onto their beaks while foraging to expand their diet breadths) [216], and complex life cycle parasites (which alter the behavior or physiology of their intermediate hosts to gain access to their definitive hosts) [217,218].
Finally, lest one forgets, we humans are perhaps the most regular associational offense users of all. Despite our extensive array of synthetic tools and elixirs and our widespread establishment of agricultural settlements, we have relied on the aid of other species in foraging throughout our known evolutionary history and continue to do so in modern times. For instance, natives of the tropical forests of Central and South America use the bark of the liana plant, Strychnos toxifera L. [219], and the secretions of poison dart frogs of the genus Phyllobates [220] to hunt for food. Members of the nomadic Boran tribe of Kenya rely on the greater honeyguide (Indicator indicator) to lead them to honeybee nests [200,221]. The Mayangna and Miskito tribes of Nicaragua rely on hunting dogs to expose and chase after potential prey [222]. Members of the Dukha tribe of Mongolia hunt with the aid of reindeer (as steeds) and golden eagles (as slayers) [223].
Moreover, use of associational offenses is not restricted to these supposedly primitive and remotely isolated groups of humans. Even in economically developed industrial societies, one finds the consumption of herbs as antidotes for hepatotoxic substances in food and drink [224,225], of peppers and other spicy plant materials to ward off food-borne pathogens [226,227], and of live bacterial and yeast cultures or decomposer enzymes to aid digestion [228,229,230]. There are the uses of live bait [231,232] and cooperation with dolphins [233] for fishing, as well as the use of tracking mammals to uncover edible fungi [234] and of pig-tailed macaques (Macaca nemestrina L.) to harvest coconuts [235]. Humans also rely on other organisms to enhance the palatability or novelty of certain foods, as in the cases of palm civets (Paradoxurus hermaphroditus Pallas) being used to predigest coffee cherries [236], yeast and bacteria being used to ferment tea leaves [237] and cocoa beans [238], and the fungus Botrytis cinerea Pers. being used to produce “botrytized wines” via “noble rot” [239]. It is unknown whether these various human associational offenses have evolved similarly to their non-human counterparts and have similar ecological consequences or if they are as different as trophy hunting is from predation [240].
Above, I have stated that associational offenses have not been deliberately incorporated as a group into ecological and evolutionary theory, despite the numerous examples that have been documented. This could be due to skepticism as to whether the effort would result in any meaningful scientific contributions, especially given that, as I have mentioned, associational offenses might be well represented or, at least, reasonably approximated in models of other related phenomena. It is fair to say that attempts to categorize phenomena using new or repurposed terminology (such as “associational offenses”) have often spawned unnecessary confusion and counter-productive semantic debates in science [241,242]. Is there, in fact, any reason to think that the dynamics of ecological interactions would change if mediated by associational offenses instead of endogenous ones? Should one expect associational offenses to generally differ from their endogenous counterparts in terms of how they evolve?
In truth, the answers to these questions are currently a matter of speculation. What we can say is that what associational offenses and endogenous offenses have in common is that they enhance a consumer’s ability to “attack and handle” its resource (Figure 4, left), and what distinguishes them is that the former require the involvement of a third species: the consumer’s offense-conferring associate. The relationship between the associate population and the measurements one would assign to the offense will most certainly vary, case to case (e.g., a venomous consumer might hypothetically need to house a thousand cells of its offense-conferring endosymbiont to produce one milliliter of venom per day and require five milliliters to kill an individual prey, or it might need much fewer endosymbionts and much less venom). The users and their associates might be so adjoined and interdependent as to merit their being regarded as a single biological unit [243]. In this case, whatever distinction one would make between associational offense users and endogenous offense users could be ecologically moot (one could argue, for instance, that tetrodotoxin is tetrodotoxin, regardless of whether it has been synthesized by an octopus, a bacterium, or a chemist).
However, environmental conditions may exclusively impact the associate, in which case the distinction between “associational” and “endogenous” offense becomes relevant even in the most intimate cases of symbiosis. For example, exposure to antibiotics would be unlikely to affect the ability of the endogenously venomous spider Zodarion nitidum Audouin to envenomate its victims [244], but it would most likely nullify that of the parasitic nematode Steinernema carpocapsae, due to the nematode being dependent on endosymbiotic bacteria for its venom [245]. This caveat has potential implications for pest control and other applications of natural resource management, as I elucidate in a later section. It is also worth considering that, when consumers are associational offense users, reciprocal co-evolutionary adaptations among consumers and resources (e.g., “Red Queen” dynamics [246,247] become reciprocal adaptations among consumers, resources, and offense-conferring associates. For example, plant immune responses to the saliva of the potato aphid (Macrosiphum euphorbiae Thomas) are triggered by proteins such as the chaperonin GroEL, which are produced by the aphid’s endosymbiotic proteobacterium Buchnera aphidicola Munson et al. [248]. Similarly, when envenomated with the deadly gut bacteria of entomopathogenic nematodes, larvae of the common cockchafer (Melolontha melolontha Fabricius) might be able to counteract the threat using associational defenses they gain from their own gut bacteria [249], making this scenario a four-player co-evolutionary dynamic (between the nematodes, the cockchafers, the nematodes’ gut bacteria, and the cockchafers’ gut bacteria).
To see how one might begin to incorporate associational offenses into existing ecological and evolutionary theory, consider the staple of modern ecological curricula known as “trophic cascades” [250]. Trophic cascades are interactions in which populations at a given “trophic level” (position in a food chain) are indirectly affected by populations two or more levels above or below theirs. A classic example is that of wolves (Canis lupus L.) promoting the growth of aspens (Populus tremuloides Michx.) by killing or frightening away elk (Cervus canadensis Erxleben), which would otherwise feed on the aspens [251]. Hypothetically, if the wolves were able to rely on a sympatric species such as the common raven (Corvus corax L.) for scouting, flushing, or lookout services, this would increase the wolves’ chances of success and reduce the costs associated with attack and handling [252,253,254,255]. The trophic cascade from wolves to aspens would thereby be less sporadic; its rate of occurrence would increase.
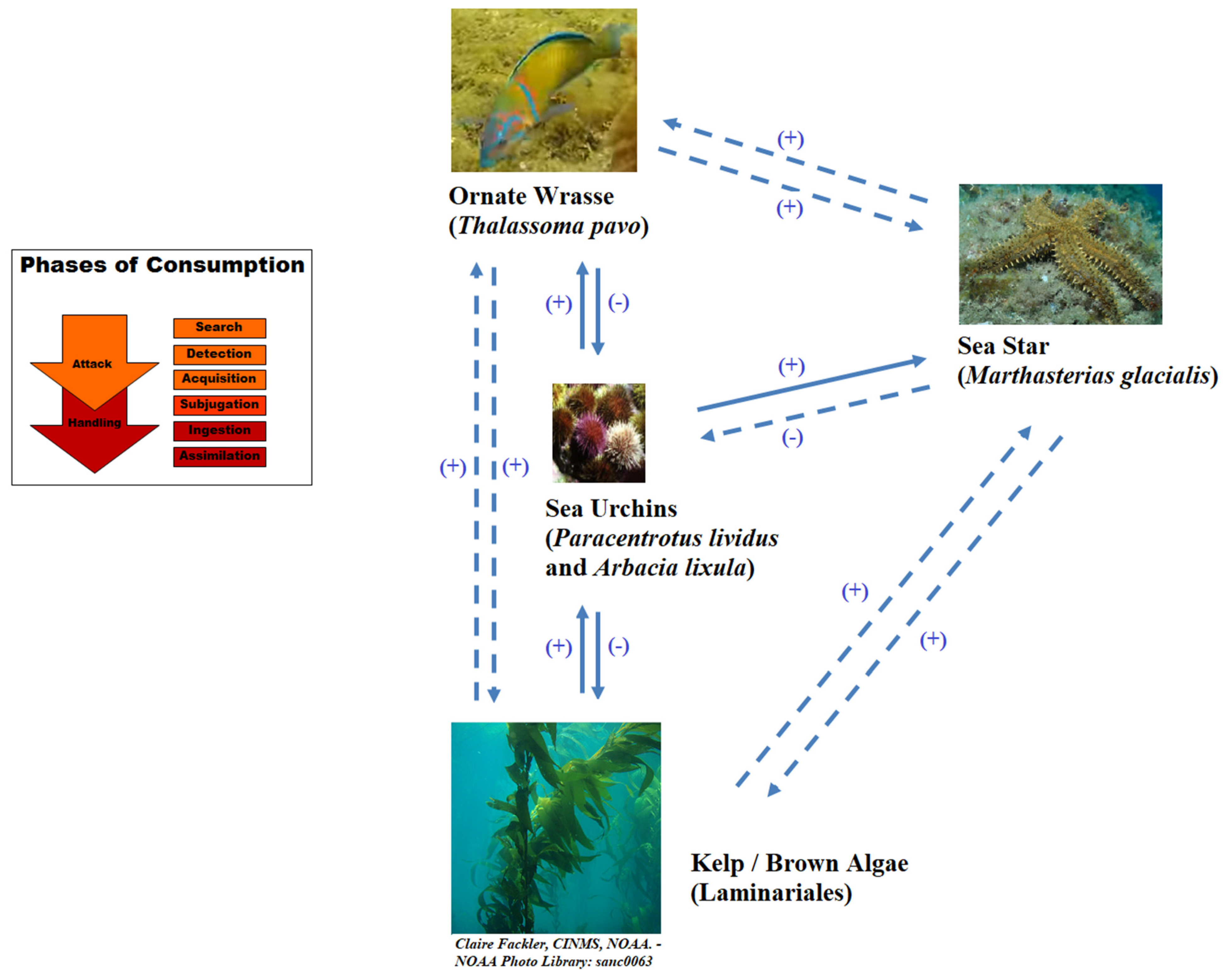
A real-life example of this kind of association-mediated trophic cascade is seen in the effect of the ornate wrasse (Thalassoma pavo L.) on the brown algae (Order Laminariales) that sustain shallow reef kelp forests in the Mediterranean Sea. A study by Galasso et al. (2015) found that a previously unknown synergism between the wrasse and the sea star Marthasterias glacialis L. led to algal recovery in Marine Protected Areas that had earlier been deforested. This prior kelp deforestation had been due to the prevalence of sea urchins (Paracentrotus lividus Lamarck and Arbacia lixula L.), which, in turn, seemed to have been caused by the overfishing of top predators. Manipulative experiments and field observations revealed that the proximity of the sea star triggers a sea urchin escape response for which the sea urchins must extend their tube feet. The wrasse exploits this behavior by feeding on the exposed tube feet, thus impairing urchin movement and making them vulnerable to predation by the sea stars [256] (Figure 4, right).
The effects of associational offenses on trophic cascades could be incorporated into existing mathematical models of predator–prey relationships (such as those of the Rosenzweig-MacArthur class [257]) as coefficients modifying attack, handling, or conversion rate terms. Rather than being constants, however, the coefficients may have to be functions of the size of the associate population (or, rather, of the participating subset of the associate population). Furthermore, the functions would have to account for case-specific variation in the associate–offense relationship (in the case of the sea star–sea urchin–kelp example, the variation would pertain to things such as how many sea stars it would take per wrasse to increase the wrasses’ encounter rate of sea urchins by one sea urchin and how many sea stars would represent the point of diminishing returns). This approach might enable the determination of the conditions under which associational offenses might destabilize food chains (e.g., drive sea urchin populations to local extinction).
2.4. Mixed Ecological Interactions
In the previous sections, I have endeavored to categorize all the main ecological interactions mediated by exogenous trait acquisition as “competition”, “mutualism”, and “consumer–resource interactions”. However, as convenient as these categories may be for organization, they are merely constructs. There are ecological relationships that fall somewhere between or beside these categories (e.g., “commensal” and “amensal” interactions [258,259]). Additionally, boundary lines between separate categories may blur or disappear completely, depending on environmental conditions [260] and on how well one accounts for trade-offs [261] and long-term population-, community-, and ecosystem-level trends [262]. For instance, precocious chicks of the brood-parasitic giant cowbird (Molothrus oryzivorus Gmelin) can provide oropendolas (Psarocolius spp., fellow birds whose nests they infiltrate) with an associational defense against botfly-driven nestling mortality, but this benefit becomes null and void in the absence of the botflies, the presence of more cost-effective defenders such as wasps, or the scarcity of oropendola eggs [263].
Moreover, it would be incorrect to assume that a trait (exogenous or otherwise) can only be used for one purpose or another. The predatory nudibranch Aeolidia papillosa L. can use the stinging cells it appropriates from sea anemones to enhance both its ability to deter predators and its ability to subdue prey [264]. Trees of the genus Cecropia, via their myrmecophytic symbioses with ants (Azteca spp.), not only gain associational defenses against herbivores and pathogens but also a competitive advantage against vines and nearby saplings [265]. Similarly, Roridula plants rely on their hemipteran symbionts not only for associational offenses, as previously mentioned, but also for facilitation of self-fertilization [266]. Additionally, interactions involving exogenous traits can overlap or be coupled. By releasing chemical cues that trigger sporulation in the fungus Neozygites tanajoae, the cassava plant (Manihot esculenta Crantz) not only staves off its predator, the green mite (Mononychellus tanajoa Bondar), but also enables its mite-pathogenic associate (the fungus) to detect and acquire new hosts (Hountondji et al. 2006) [267], thereby simultaneously giving rise to both an associational defense (from the perspective of the plant) and an associational offense (from the perspective of the fungus). Symbiotic bacteria in the skin of red-backed salamanders produce emergent metabolites that may play roles in bacterial allelopathy but also provide the salamanders with an associational defense against the chytrid fungus Batrachochytrium dendrobatidis Berger (the cause of an on-going mass extinction of amphibians throughout the world [268]) [269].
2.5. Implications of Exogenous Trait Acquisition for Management and Theory
The strategy of acquiring exogenous traits through community ecological interactions with other species is neither unusual nor trivial. As I have touched on throughout this review, this strategy is employed by a vast array of organisms in various ways and may represent important opportunities and concerns for natural resource management, human health, industry, and ecological and evolutionary theory. Some of the organisms that employ association-derived exogenous traits have become invasive and devastated ecosystems such as grasslands [154,270], temperate forests [209,271], and coral reefs [149]. Some have been linked to epidemics of diseases, such as avian vacuolar myelinopathy, a cause of recurrent die-offs in bald eagles [272]. Others are not pestilent but are, nonetheless, equally important in the context of natural resource management, including some widely used biological control agents of herbivorous insects among entomophagous nematodes and parasitoid wasps [273,274].
Failure to consider the acquisition of exogenous traits in our analyses and projections might result in significant gaps in our understanding of ecological communities, as well as impair our ability to fulfill commercial and natural resource management objectives. There has already been some real-world evidence of it having delayed our understanding of the basis for and means of controlling pestilence and disease, especially in the contexts of pesticide and antimicrobial resistance [275]. For example, although development of insecticide resistance has historically been attributed to evolutionary changes in pest insect genomes (e.g., alteration of drug target sites, up-regulation of degrading enzymes, and enhancement of drug excretion), Kikuchi et al. (2012) demonstrated that insecticide resistance can be established immediately via the insecticide-degrading bacterial symbionts that nymphal stages of these pest insects acquire from the soil every generation [276]. Similarly, Xu et al. (2014) discovered that mutualistic viruses such as the novel densovirus HaDNV-1 can provide their hosts with resistance to both chemical pesticides such as Bacillus thuringiensis (Bt) toxin and pathogenic biological control agents like baculovirus HaNPV [277]. In the same vein, a review by Banerji et al. (2019) elucidated the role of ecological interactions in creating or promoting antimicrobial resistance among bacteria and other microbes, even in the absence of known antimicrobial resistance genes (“ARGs”) [121].
To be clear, the phenomenon of exogenous trait acquisition (and occurrence of ecological synergism in general) does not invalidate or undermine the relevance of studies that have focused on endogenous traits and their underlying genetics, nor does it necessitate that researchers check for exogenous trait acquisition in all studies pertaining to biodiversity and ecological community dynamics. Recognition of exogenous trait acquisition is not cause to dismiss what we have already learned. If anything, it is the opposite: a reminder not to ignore or overlook the fundamentals of ecology and evolution as we make progress in our understanding of the genetic and epigenetic mechanisms that give rise to characteristics of living beings. In fact, assuming that there are acquisitions of exogenous traits in a given system (e.g., because of “old wives’ tales”) can occasionally be worse from the standpoint of conservation and natural resource management than discounting the possibility outright. Dean et al. (1990) have articulated, for example, how some ill-conceived approaches to management of African nature reserves and some subsequent human rights violations against indigenous tribes were all partly fueled by the charisma of an unsubstantiated (yet long-perpetuated) account of honeyguides escorting juvenile ratels (Mellivora capensis Schreber) to beehives (as they do humans and other mammals) [200].
A productive way forward would be to capitalize on the prospect of adapting the tools that have been used to study endogenous traits to explore the mechanisms and broader impacts of exogenous trait acquisition in nature. This would not only help to ensure that one does not misjudge organisms’ ranges of tolerance and misinterpret their population dynamics but also provide a solid conceptual framework to support the development of new hypotheses and techniques. In the context of integrated pest management, one could extend the tried-and-tested methods of targeting vectors to control disease to include targeting other kinds of defense- or offense-conferring associates. For instance, one might follow the example set by Mauldin et al. (1981) and others (Waller 1996) of using antibiotic- or pathogen-infused bait wood to combat termites [278,279]. In said example, the antibiotics and pathogens disable the termites’ ability to safely ingest wood by killing or supplanting the endosymbionts that confer this ability [280]. Tarascoa et al. (2011) have similarly demonstrated how one might quantify and predict the combined action of differing biological control agents that employ associational and endogenous offenses against the same target pests. In the case of their study, the agents were the entomopathogenic nematode Steinernema ichnusae (which, like S. carpocapsae, envenomates its prey with virulent endosymbiotic bacteria) and the entomopathogenic fungus Beauveria bassiana (Bals.-Criv.) Vuill., both feeding on larvae of the honeycomb moth Galleria mellonella Fabricius [281].
One might also consider bioengineering defense- or offense-conferring symbionts [282] or introducing probiotics and dietary supplements to bolster the effectiveness of biological control agents. The latter would be akin to introducing alternative prey or hosts to enhance the effectiveness of biological agents through “apparent competition” [145,283]. Similarly, for the purposes of conservation or restoration, one might design landscapes to enhance or reduce neighborhood habitat amelioration, as has been recommended for initiatives intended to stem dryland degradation [284]. One might also introduce organisms capable of occupying a critical niche that was lost due to the recent extinction of an associate, which would fit the scheme of the proposed substitution of the extinct dodo bird (Raphus cucullatus L.) with the extant turkey bird (Meleagris gallopavo L.) to preserve the tambalacoque tree (Sideroxylon grandiflorum A.DC., formerly Calvaria major) [285] (but see [286] for a critical review of this particular example).
There are, in addition, the innovations in microscopy [287,288], computer processing [289], and high-throughput sequencing [290,291,292] that, over the past decade, have permitted scientists to perform species classifications and detect and quantify (in bulk) traits such as morphology, behavior, and metabolic capability with greater speed and precision than ever before [293,294,295]. Some of these modern tools and techniques could be employed not only to track functional traits across whole ecological communities but also to identify what each participant in an exogenous trait-conferring association contributes when the trait is a product of synergism (i.e., where none of the species involved possess the genetic machinery to produce the trait by themselves) [268,296]. The latter has been conducted to a limited extent in studies of the symbiotic associations between the fungi and photoautotrophic microbes that create lichen [295,297,298].
3. Conclusions
Studies attempting to uncover, quantify, and evaluate traits of interest (e.g., antimicrobial resistance and metal resistance) often start with the working premise that these traits are endogenous (e.g., phenotypic expressions of the species’ intrinsic ARGs and metal resistance genes) [299]. This is an intuitive application of the Occam’s razor principle. It may also be that the growing speed and precision with which one can now isolate endogenous traits and their underlying genetic bases have encouraged the proliferation of studies dedicated to this task [300,301,302], as have the success and profitability of applications in bioprospecting [303]. However, as previous authors have underscored, even for these purposes, it is necessary to put species traits in their proper ecological context, accounting for the life-history strategies of the organisms that possess them [304,305] and for the possibility of ecological communities exhibiting “functional complementarity” [306,307].
As I have expounded in this review, exogenous traits that are acquired through interspecific association can be as important and worthwhile to consider as endogenous traits. Updating conceptual and mathematical models in ecology and evolution to explicitly incorporate them may bring to fruition exciting prospects for interdisciplinary collaboration and creative new avenues of applied science. Future research should seek to optimize the value of such endeavors, as well as reflect on the insights to refine past interpretations of important ecological and evolutionary trends.
Funding
This research received no external funding.
Acknowledgments
The author gratefully acknowledges Peter Morin, Cara Faillace, Ken Fritz, Nina Fefferman, Emily Griffiths, Dennis Hansen, Owen Petchey, Hank Stevens, Sydney Greenlee, Holly Vuong, Carrie Norin, and Timon McPhearson for their insights and encouragement.
Disclaimer
All photographic images were obtained from the public domain (Wikimedia Commons and YouTube Creative Commons). Any opinions expressed in this paper are those of the author and do not necessarily reflect the views of the U.S. Environmental Protection Agency; therefore, no official endorsement should be inferred. Any mention of trade names or commercial products does not constitute endorsement or recommendation for use.
Conflicts of Interest
The author declares no conflict of interest.
References
- McKitrick, M.C. Phylogenetic constraint in evolutionary theory: Has it any explanatory power? Ann. Rev. Ecol. Syst. 1993, 24, 307–330. [Google Scholar] [CrossRef]
- Diamond, J.M. Dirty eating for healthy living. Nat. Cell Biol. 1999, 400, 120–121. [Google Scholar] [CrossRef] [PubMed]
- Pyerin, W.; Taniguchi, H. Phosphorylation of hepatic phenobarbital-inducible cytochrome P-450. EMBO J. 1989, 8, 3003–3010. [Google Scholar] [CrossRef]
- Tanimoto, Y.; Kaneko, H.; Ohkuma, T.; Oguri, K.; Yoshimura, H. Site-selective oxidation of strychnine by phenobarbital inducible cytochrome P-450. J. Pharm. Dyn. 1991, 14, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Rampazzo, N.; Blum, W.E.H. Changes in chemistry and mineralogy of forest soils by acid rain. Water Air Soil Pollut. 1992, 61, 209–220. [Google Scholar] [CrossRef]
- Freebairn, D.; Loch, R.; Silburn, D. Chapter 9 Soil erosion and soil conservation for vertisols. Dev. Pedotransfer Funct. Soil Hydrol. 1996, 24, 303–362. [Google Scholar] [CrossRef]
- Molina, E.; Espelta, J.M.; Pino, J.; Bagaria, G.; Armenteras, D. Influence of clay licks on the diversity and structure of an Amazonian forest. Biotropica 2018, 50, 740–749. [Google Scholar] [CrossRef]
- Walsh, M.R.; Munch, S.B.; Chiba, S.; Conover, D.O. Maladaptive changes in multiple traits caused by fishing: Impediments to population recovery. Ecol. Lett. 2005, 9, 142–148. [Google Scholar] [CrossRef]
- Baird, D.J.; Rubach, M.N.; Van den Brinkt, P.J. Trait-based ecological risk assessment (TERA): The new frontier? Integr. Environ. Assess. Manag. 2009, 4, 2–3. [Google Scholar] [CrossRef]
- Baker, R.H.A.; Cannon, R.; Bartlett, P.; Barker, I. Novel strategies for assessing and managing the risks posed by invasive alien species to global crop production and biodiversity. Ann. Appl. Biol. 2005, 146, 177–191. [Google Scholar] [CrossRef]
- Tremlová, K.; Münzbergová, Z. Importance of species traits for species distribution in fragmented landscapes. Ecology 2007, 88, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Halliday, F.W.; Rohr, J.R. Measuring the shape of the biodiversity-disease relationship across systems reveals new findings and key gaps. Nat. Commun. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Lips, K.R.; Reeve, J.D.; Witters, L.R. Ecological traits predicting amphibian population declines in Central America. Conserv. Biol. 2003, 17, 1078–1088. [Google Scholar] [CrossRef] [Green Version]
- Laliberté, E.; Wells, J.A.; Declerck, F.; Metcalfe, D.J.; Catterall, C.P.; Queiroz, C.; Aubin, I.; Bonser, S.P.; Ding, Y.; Fraterrigo, J.M.; et al. Land-use intensification reduces functional redundancy and response diversity in plant communities. Ecol. Lett. 2010, 13, 76–86. [Google Scholar] [CrossRef]
- Butterfield, B.J.; Callaway, R.M. A functional comparative approach to facilitation and its context dependence. Funct. Ecol. 2013, 27, 907–917. [Google Scholar] [CrossRef]
- Morris, L.; O’Brien, A.; Natera, S.H.; Lutz, A.; Roessner, U.; Long, S.M. Structural and functional measures of marine microbial communities: An experiment to assess implications for oil spill management. Mar. Pollut. Bull. 2018, 131, 525–529. [Google Scholar] [CrossRef]
- Sun, D. Pull in and push out: Mechanisms of horizontal gene transfer in bacteria. Front. Microbiol. 2018, 9, 2154. [Google Scholar] [CrossRef]
- Dufty, A.M., Jr.; Clobert, J.; Møller, A.P. Hormones, developmental plasticity and adaptation. Trends Ecol. Evol. 2002, 17, 190–196. [Google Scholar] [CrossRef]
- Fusco, G.; Minelli, A. Phenotypic plasticity in development and evolution: Facts and concepts. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Belcourt, M. Outsourcing-the benefits and the risks. Hum. Res. Manag. Rev. 2006, 16, 269–279. [Google Scholar] [CrossRef]
- Sapp, J. Evolution by Association: A History of Symbiosis; Oxford University Press: New York, NY, USA, 1994. [Google Scholar]
- Flower, T.P.; Child, M.F.; Ridley, A.R. The ecological economics of kleptoparasitism: Pay-offs from self-foraging versus kleptoparasitism. J. Anim. Ecol. 2012, 82, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Paini, D.R.; Funderburk, J.E.; Reitz, S.R. Competitive exclusion of a worldwide invasive pest by a native. Quantifying competition between two phytophagous insects on two host plant species. J. Anim. Ecol. 2008, 77, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Van De Waal, D.B.; Verspagen, J.M.H.; Finke, J.F.; Vournazou, V.; Immers, A.K.; Kardinaal, W.E.A.; Tonk, L.; Becker, S.; Van Donk, E.; Visser, P.M.; et al. Reversal in competitive dominance of a toxic versus non-toxic cyanobacterium in response to rising CO2. ISME J. 2011, 5, 1438–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Bourlot, V.; Tully, T.; Claessen, D. Interference versus exploitative competition in the regulation of size-structured populations. Am. Nat. 2014, 184, 609–623. [Google Scholar] [CrossRef] [PubMed]
- Wignall, V.R.; Harry, I.C.; Davies, N.L.; Kenny, S.D.; McMINN, J.K.; Ratnieks, F.L.W. Seasonal variation in exploitative competition between honeybees and bumblebees. Oecologia 2019, 192, 351–361. [Google Scholar] [CrossRef] [Green Version]
- Connell, J.H. On the prevalence and relative importance of interspecific competition: Evidence from field experiments. Am. Nat. 1983, 122, 661–696. [Google Scholar] [CrossRef]
- Halpern, C.B.; Lutz, J.A. Canopy closure exerts weak controls on understory dynamics: A 30-year study of overstory–understory interactions. Ecol. Monogr. 2013, 83, 221–237. [Google Scholar] [CrossRef] [Green Version]
- Oda, R. Scent marking and contact call production in ring-tailed Lemurs (Lemur catta). Folia Primatol. 1999, 70, 121–124. [Google Scholar] [CrossRef]
- Wegener, C.; Martin, B.; Didden, C.; Edmunds, P.J. Overgrowth of Caribbean octocorals by milleporid hydrocorals. Invertebr. Biol. 2017, 137, 29–37. [Google Scholar] [CrossRef]
- Prati, D.; Bossdorf, O. Allelopathic inhibition of germination by Alliaria petiolata (Brassicaceae). Am. J. Bot. 2004, 91, 285–288. [Google Scholar] [CrossRef] [Green Version]
- Samhouri, J.F.; Vance, R.R.; Forrester, G.E.; Steele, M.A. Musical chairs mortality functions: Density-dependent deaths caused by competition for unguarded refuges. Oecologia 2009, 160, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia uses a host microRNA to regulate transcripts of a methyltransferase, contributing to dengue virus inhibition in Aedes aegypti. Proc. Natl. Acad. Sci. USA 2013, 110, 10276–10281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mousa, W.K.; Shearer, C.; Limay-Rios, V.; Ettinger, C.L.; Eisen, J.A.; Raizada, M.N. Root-hair endophyte stacking in finger millet creates a physicochemical barrier to trap the fungal pathogen Fusarium graminearum. Nat. Microbiol. 2016, 1, 16167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, A.S.; Moffat, C.S.; Lopez-Ruiz, F.J.; Gibberd, M.R.; Hamblin, J.; Zerihun, A. Host-multi-pathogen warfare: Pathogen interactions in co-infected plants. Front. Plant Sci. 2017, 8, 1806. [Google Scholar] [CrossRef] [Green Version]
- Cézilly, F.; Perrot-Minnot, M.-J.; Rigaud, T. Cooperation and conflict in host manipulation: Interactions among macro-parasites and micro-organisms. Front. Microbiol. 2014, 5, 248. [Google Scholar] [CrossRef]
- Hilker, F.M.; Allen, L.J.S.; Bokil, V.; Briggs, C.J.; Feng, Z.; Garrett, K.A.; Gross, L.J.; Hamelin, F.; Jeger, M.J.; Manore, C.A.; et al. Modeling virus coinfection to inform management of maize lethal necrosis in Kenya. Phytopathology 2017, 107, 1095–1108. [Google Scholar] [CrossRef] [Green Version]
- Powell, J.R. Mosquito-borne human viral diseases: Why Aedes aegypti? Am. J. Trop. Med. Hyg. 2018, 98, 1563–1565. [Google Scholar] [CrossRef]
- Nazareth, T.; Craveiro, I.; Moutinho, A.; Seixas, G.; Gonçalves, C.; Gonçalves, L.; Teodósio, R.; Sousa, C.A. What happens when we modify mosquitoes for disease prevention? A systematic review. Emerg. Microbes Infect. 2020, 9, 348–365. [Google Scholar] [CrossRef]
- Chu, Z.; Jin, X.; Yang, B.; Zeng, Q. Buoyancy regulation of Microcystis flos-aquae during phosphorus-limited and nitrogen-limited growth. J. Plankton Res. 2007, 29, 739–745. [Google Scholar] [CrossRef] [Green Version]
- Wilde, A.; Mullineaux, C.W. Motility in cyanobacteria: Polysaccharide tracks and Type IV pilus motors. Mol. Microbiol. 2015, 98, 998–1001. [Google Scholar] [CrossRef]
- Pratt, J.R.; Rosen, B.H. Associations of species of Vorticella (Peritrichida) and Planktonic Algae. Trans. Am. Microsc. Soc. 1983, 102, 48. [Google Scholar] [CrossRef]
- Canter, H.M.; Walsby, A.; Kinsman, R.; Ibelings, B. The effect of attached vorticellids on the buoyancy of the colonial cyanobacteriumAnabaena lemmermannii. Br. Phycol. J. 1992, 27, 65–74. [Google Scholar] [CrossRef] [Green Version]
- Chia, M.A.; Jankowiak, J.G.; Kramer, B.J.; Goleski, J.A.; Huang, I.-S.; Zimba, P.V.; Bittencourt-Oliveira, M.D.C.; Gobler, C. Succession and toxicity of microcystis and Anabaena (Dolichospermum) blooms are controlled by nutrient-dependent allelopathic interactions. Harmful Algae 2018, 74, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Murray, B.G., Jr. The ecological consequences of interspecific territorial behavior in birds. Ecology 1971, 52, 414–423. [Google Scholar] [CrossRef]
- Waser, P.M. Interactions among primate species. In Primate Societies; Smuts, B.B., Cheny, D.L., Seyfarth, R.M., Wrangham, R.W., Struhsaker, T.T., Eds.; University of Chicago Press: Chicago, IL, USA, 1986; pp. 210–226. [Google Scholar]
- Stensland, E.; Angerbjörn, A.; Berggren, P. Mixed species groups in mammals. Mammal. Rev. 2003, 33, 205–223. [Google Scholar] [CrossRef]
- Jullien, M.; Thiollay, J.-M. Multi-species territoriality and dynamic of neotropical forest understory bird flocks. J. Anim. Ecol. 1998, 67, 227–252. [Google Scholar] [CrossRef]
- Bshary, R.; Grutter, A.S. Punishment and partner switching cause cooperative behaviour in a cleaning mutualism. Biol. Lett. 2005, 1, 396–399. [Google Scholar] [CrossRef]
- Weyl, E.G.; Frederickson, M.E.; Yu, D.W.; Pierce, N.E. Economic contract theory tests models of mutualism. Proc. Natl. Acad. Sci. USA 2010, 107, 15712–15716. [Google Scholar] [CrossRef] [Green Version]
- Groom, M.J. Sand-colored nighthawks parasitize the antipredator behavior of three nesting bird species. Ecology 1992, 73, 785–793. [Google Scholar] [CrossRef]
- Nolen, M.T.; Lucas, J.R. Asymmetries in mobbing behaviour and correlated intensity during predator mobbing by nuthatches, chickadees and titmice. Anim. Behav. 2009, 77, 1137–1146. [Google Scholar] [CrossRef]
- Brown, J.L. The evolution of diversity in avian territorial systems. Wilson Bull. 1964, 76, 160–169. [Google Scholar]
- Hinsch, M.; Komdeur, J. Defence, intrusion and the evolutionary stability of territoriality. J. Theor. Biol. 2010, 266, 606–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezende, E.L.; Lavabre, J.E.; Guimarães, P.R.; Jordano, P.; Bascompte, J. Non-random coextinctions in phylogenetically structured mutualistic networks. Nat. Cell Biol. 2007, 448, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.C.; Almeida-Neto, M. A simple stochastic model for complex coextinctions in mutualistic networks: Robustness decreases with connectance. Ecol. Lett. 2015, 18, 144–152. [Google Scholar] [CrossRef] [PubMed]
- LeClaire, S.; Jacob, S.; Greene, L.K.; Dubay, G.R.; Drea, C.M. Social odours covary with bacterial community in the anal secretions of wild meerkats. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Cohan, F.M. Bacterial species and speciation. Syst. Biol. 2001, 50, 513–524. [Google Scholar] [CrossRef] [PubMed]
- Fondi, M.; Karkman, A.; Tamminen, M.V.; Bosi, E.; Virta, M.; Fani, R.; Alm, E.; McInerney, J.O. “Every gene is everywhere but the environment selects”: Global geolocalization of gene sharing in environmental samples through network analysis. Genome Biol. Evol. 2016, 8, 1388–1400. [Google Scholar] [CrossRef]
- Giller, K.E.; Witter, E.; McGrath, S.P. Toxicity of heavy metals to microorganisms and microbial processes in agricultural soils: A review. Soil Biol. Biochem. 1998, 30, 1389–1414. [Google Scholar] [CrossRef]
- Daghrir, R.; Drogui, P. Tetracycline antibiotics in the environment: A review. Environ. Chem. Lett. 2013, 11, 209–227. [Google Scholar] [CrossRef]
- Adamo, S.A. Parasitic aphrodisiacs: Manipulation of the hosts’ behavioral defenses by sexually transmitted parasites. Integr. Comp. Biol. 2014, 54, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Sadeh, A.; Northfield, T.D.; Rosenheim, J.A. The epidemiology and evolution of parasite transmission through cannibalism. Ecology 2016, 97, 2003–2011. [Google Scholar] [CrossRef] [PubMed]
- Veiga, J.P.; Polo, V.; Viñuela, J. Nest green plants as a male status signal and courtship display in the spotless starling. Ethology 2006, 112, 196–204. [Google Scholar] [CrossRef]
- Albo, M.J.; Toft, S.; Bilde, T. Female spiders ignore condition-dependent information from nuptial gift wrapping when choosing mates. Anim. Behav. 2012, 84, 907–912. [Google Scholar] [CrossRef]
- Airoldi, L. Effects of disturbance, life histories, and overgrowth on coexistence of algal crusts and turfs. Ecology 2000, 81, 798–814. [Google Scholar] [CrossRef]
- Sikkel, P.C. Effects of nest quality on male courtship and female spawning-site choice in an algal-nesting damselfish. Bull. Mar. Sci. 1995, 57, 682–689. [Google Scholar]
- Denley, D.; Metaxas, A. Effects of intrinsic and extrinsic factors on reproduction of an ecologically significant invasive bryozoan: Implications for invasion success. Mar. Biol. 2017, 164, 145. [Google Scholar] [CrossRef]
- Yoshioka, P.M. Role of planktonic and benthic factors in the population dynamics of the Bryozoan Membranipora membranacea. Ecology 1982, 63, 457–468. [Google Scholar] [CrossRef]
- Pratt, M.C.; Grason, E.W. Invasive species as a new food source: Does a nudibranch predator prefer eating an invasive bryozoan? Biol. Invasions 2006, 9, 645–655. [Google Scholar] [CrossRef]
- Green, P.T.; O’Dowd, D.J.; Abbott, K.L.; Jeffery, M.; Retallick, K.; Mac Nally, R. Invasional meltdown: Invader–invader mutualism facilitates a secondary invasion. Ecology 2011, 92, 1758–1768. [Google Scholar] [CrossRef]
- Dillon, R.J.; Vennard, C.T.; Buckling, A.; Charnley, A.K. Diversity of locust gut bacteria protects against pathogen invasion. Ecol. Lett. 2005, 8, 1291–1298. [Google Scholar] [CrossRef]
- Brown, S.P.; Le Chat, L.; Taddei, F. Evolution of virulence: Triggering host inflammation allows invading pathogens to exclude competitors. Ecol. Lett. 2007, 11, 44–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stecher, B.; Robbiani, R.; Walker, A.W.; Westendorf, A.M.; Barthel, M.; Kremer, M.; Chaffron, S.; MacPherson, A.J.; Buer, J.; Parkhill, J.; et al. Salmonella enterica Serovar Typhimurium exploits inflammation to compete with the intestinal microbiota. PLoS Biol. 2007, 5, e244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okoro, C.K.; Kingsley, R.A.; Connor, T.R.; Harris, S.R.; Parry, C.M.; Al-Mashhadani, M.N.; Kariuki, S.; Msefula, C.L.; Gordon, M.A.; De Pinna, E.; et al. Intracontinental spread of human invasive Salmonella Typhimurium pathovariants in sub-Saharan Africa. Nat. Genet. 2012, 44, 1215–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkinson, C.R.; Garrone, R.; Vacelet, J. Marine sponges discriminate between food bacteria and bacterial symbionts: Electron microscope radioautography and in situ evidence. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1984, 220, 519. [Google Scholar]
- Dolenec, T.; Lojen, S.; Kniewald, G.; Dolenec, M.; Rogan, N. Nitrogen stable isotope composition as a tracer of fish farming in invertebrates Aplysina aerophoba, Balanus perforatus and Anemonia sulcata in central Adriatic. Aquaculture 2007, 262, 237–249. [Google Scholar] [CrossRef]
- Bayer, K.; Schmitt, S.; Hentschel, U. Physiology, phylogeny and in situ evidence for bacterial and archaeal nitrifiers in the marine sponge Aplysina aerophoba. Environ. Microbiol. 2008, 10, 2942–2955. [Google Scholar] [CrossRef]
- Harris, R.J.; Jenner, R.A. Evolutionary ecology of fish venom: Adaptations and consequences of evolving a venom system. Toxins 2019, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, O.J.; Nudds, T.D. Parasite-mediated competition in deer and moose: How strong is the effect of Meningeal worm on moose? Ecol. Appl. 1994, 4, 91–103. [Google Scholar] [CrossRef]
- Price, P.W.; Westoby, M.; Rice, B. Parasite-mediated competition: Some predictions and tests. Am. Nat. 1988, 131, 544–555. [Google Scholar] [CrossRef] [Green Version]
- Mlynarek, J.J.; Knee, W.; Forbes, M.R. Host phenology, geographic range size and regional occurrence explain interspecific variation in damselfly-water mite associations. Ecography 2014, 38, 670–680. [Google Scholar] [CrossRef]
- Willis, R.J. The History of Allelopathy; Springer: New York, NY, USA, 2007. [Google Scholar]
- Mangla, S.; Callaway, R.M. Exotic invasive plant accumulates native soil pathogens which inhibit native plants. J. Ecol. 2007, 96, 58–67. [Google Scholar] [CrossRef]
- Osman, R.W.; Haugsness, J.A. Mutualism among sessile invertebrates: A mediator of competition and predation. Science 1981, 211, 846–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodall, J.; Erasmus, D. Review of the status and integrated control of the invasive alien weed, Chromolaena odorata, in South Africa. Agric. Ecosyst. Environ. 1996, 56, 151–164. [Google Scholar] [CrossRef]
- Canning-Clode, J.; Souto, J.; McCann, L. First record of Celleporaria brunnea (Bryozoa: Lepraliellidae) in Portugal and in the East Atlantic. Mar. Biodivers. Rec. 2013, 6, 108. [Google Scholar] [CrossRef]
- Haeder, S.; Wirth, R.; Herz, H.; Spiteller, D. Candicidin-producing Streptomyces support leaf-cutting ants to protect their fungus garden against the pathogenic fungus Escovopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 4742–4746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otani, S.; Challinor, V.L.; Kreuzenbeck, N.B.; Kildgaard, S.; Christensen, S.K.; Larsen, L.L.M.; Aanen, D.K.; Rasmussen, S.A.; Beemelmanns, C.; Poulsen, M. Disease-free monoculture farming by fungus-growing termites. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kasson, M.T.; Wickert, K.L.; Stauder, C.M.; Macias, A.M.; Berger, M.C.; Simmons, D.R.; Short, D.P.; DeVallance, D.B.; Hulcr, J. Mutualism with aggressive wood-degrading Flavodon ambrosius (Polyporales) facilitates niche expansion and communal social structure in Ambrosiophilus ambrosia beetles. Fungal Ecol. 2016, 23, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Ranger, C.M.; Biedermann†, P.H.W.; Phuntumart, V.; Beligala, G.U.; Ghosh, S.; Palmquist, D.E.; Mueller, R.; Barnett, J.; Schultz, P.B.; Reding, M.E.; et al. Symbiont selection via alcohol benefits fungus farming by ambrosia beetles. Proc. Natl. Acad. Sci. USA 2018, 115, 4447–4452. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Epsky, N.D. An uncertain future for American Lauraceae: A lethal threat from redbay ambrosia beetle and laurel wilt disease (a review). Am. J. Plant Sci. 2013, 4, 727–738. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C.; Hulcr, J.; Wingfield, M.J.; De Beer, Z.W. Destructive tree diseases associated with ambrosia and bark beetles: Black swan events in tree pathology? Plant Dis. 2013, 97, 856–872. [Google Scholar] [CrossRef] [Green Version]
- Kusch, J.; Stremmel, M.; Breiner, H.-W.; Adams, V.; Schweikert, M.; Schmidt, H.J. The toxic symbiont Caedibacter caryophila in the cytoplasm of Paramecium novaurelia. Microb. Ecol. 2000, 40, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Schrallhammer, M. The killer trait of Paramecium and its causative agents. Palaeodiversity 2010, 3, 79–88. [Google Scholar]
- Chen, T.-T. Paramecin 34, A Killer substance produced by Paramecium bursaria. Exp. Biol. Med. 1955, 88, 541–543. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.P.; Hauzy, C.; Hulot, F.D. Ingredients for protist coexistence: Competition, endosymbiosis and a pinch of biochemical interactions. J. Anim. Ecol. 2011, 81, 222–232. [Google Scholar] [CrossRef]
- Bronstein, J.L. Our current understanding of mutualism. Q. Rev. Biol. 1994, 69, 31–51. [Google Scholar] [CrossRef]
- Connor, R.C. The benefits of mutualism: A conceptual framework. Biol. Rev. 1995, 70, 427–457. [Google Scholar] [CrossRef] [Green Version]
- Stachowicz, J.J. Mutualism, facilitation, and the structure of ecological communities: Positive interactions play a critical, but underappreciated, role in ecological communities by reducing physical or biotic stresses in existing habitats and by creating new habitats on which many species depend. BioScience 2001, 51, 235–246. [Google Scholar]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect-Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [Green Version]
- Gammans, N.; Bullock, J.M.; Schönrogge, K. Ant benefits in a seed dispersal mutualism. Oecologia 2005, 146, 43–49. [Google Scholar] [CrossRef]
- Peoples, B.K.; Frimpong, E.A. Context-dependent outcomes in a reproductive mutualism between two freshwater fish species. Ecol. Evol. 2016, 6, 1214–1223. [Google Scholar] [CrossRef]
- Bull, L.; Fogarty, T.C. Artificial symbiogenesis. Artif. Life 1995, 2, 269–292. [Google Scholar] [CrossRef] [PubMed]
- Chapman, M.J.; Margulis, L. Morphogenesis by symbiogenesis. Int. Microbiol. 1998, 1, 319–326. [Google Scholar] [PubMed]
- Martin, W.F.; Garg, S.; Zimorski, V. Endosymbiotic theories for eukaryote origin. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corning, P.A. “The synergism hypothesis”: On the concept of synergy and its role in the evolution of complex systems. J. Soc. Evol. Syst. 1998, 21, 133–172. [Google Scholar] [CrossRef]
- Feijen, F.A.A.; Vos, R.A.; Nuytinck, J.; Merckx, V.S.F.T. Evolutionary dynamics of mycorrhizal symbiosis in land plant diversification. Sci. Rep. 2018, 8, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Kerney, R.R. Symbioses between salamander embryos and green algae. Symbiosis 2011, 54, 107–117. [Google Scholar] [CrossRef]
- Cohen, J.E.; Goldstone, A.B.; Paulsen, M.J.; Shudo, Y.; Steele, A.N.; Edwards, B.B.; Patel, J.B.; MacArthur, J.W., Jr.; Hopkins, M.S.; Burnett, C.E.; et al. An innovative biologic system for photon-powered myocardium in the ischemic heart. Sci. Adv. 2017, 3, e1603078. [Google Scholar] [CrossRef] [Green Version]
- Ewers, R. Maxilla sinus grafting with marine algae derived bone forming material: A clinical report of long-term results. J. Oral Maxillofac. Surg. 2005, 63, 1712–1723. [Google Scholar] [CrossRef]
- Bertness, M.D.; Callaway, R. Positive interactions in communities. Trends Ecol. Evol. 1994, 9, 191–193. [Google Scholar] [CrossRef]
- Holzapfel, C. Bidirectional facilitation and interference between shrubs and annuals in the Mojave Desert. Ecology 1999, 80, 1747–1761. [Google Scholar] [CrossRef]
- Holmgren, M.; Scheffer, M.; Huston, M.A. The interplay of facilitation and competition in plant communities. Ecology 1997, 78, 1966–1975. [Google Scholar] [CrossRef]
- Webb, J.S.; Givskov, M.C.; Kjelleberg, S. Bacterial biofilms: Prokaryotic adventures in multicellularity. Curr. Opin. Microbiol. 2003, 6, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Claessen, D.; Rozen, D.E.; Kuipers, O.P.; Søgaard-Andersen, L.; Van Wezel, G.P. Bacterial solutions to multicellularity: A tale of biofilms, filaments and fruiting bodies. Nat. Rev. Genet. 2014, 12, 115–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teitzel, G.M.; Parsek, M.R. Heavy metal resistance of biofilm and planktonic Pseudomonas aeruginosa. Appl. Environ. Microbiol. 2003, 69, 2313–2320. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Røder, H.L.; Madsen, J.S.; Bjarnsholt, T.; Sørensen, S.J.; Burmølle, M. Interspecific bacterial interactions are reflected in multispecies biofilm spatial organization. Front. Microbiol. 2016, 7, 1366. [Google Scholar] [CrossRef] [Green Version]
- Donlan, R.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Banerji, A.; Jahne, M.; Herrmann, M.; Brinkman, N.; Keely, S. Bringing community ecology to bear on the issue of antimicrobial resistance. Front. Microbiol. 2019, 10, 2626. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Hamayun, M.; Radhakrishnan, R.; Waqas, M.; Kang, S.-M.; Kim, Y.; Shin, J.-H.; Choo, Y.-S.; Kim, J.-G.; Lee, I.-J. Mutualistic association of Paecilomyces formosus LHL10 offers thermotolerance to Cucumis sativus. Antonie Leeuwenhoek 2011, 101, 267–279. [Google Scholar] [CrossRef]
- Bilal, S.; Shahzad, R.; Imran, M.; Jan, R.; Kim, K.M.; Lee, I.-J. Synergistic association of endophytic fungi enhances Glycine max L. resilience to combined abiotic stresses: Heavy metals, high temperature and drought stress. Ind. Crop. Prod. 2020, 143, 111931. [Google Scholar] [CrossRef]
- Torabi, A.; Bonjar, G.H.S.; Abdolshahi, R.; Pournamdari, M.; Saadoun, I.; Barka, E.A. Biological control of Paecilomyces formosus, the causal agent of dieback and canker diseases of pistachio by two strains of Streptomyces misionensis. Biol. Control. 2019, 137, 104029. [Google Scholar] [CrossRef]
- Hanna, H. Black polyethylene mulch does not reduce yield of cucumbers double-cropped with tomatoes under heat stress. HortScience 2000, 35, 190–191. [Google Scholar] [CrossRef] [Green Version]
- Neelakanta, G.; Sultana, H.; Fish, D.; Anderson, J.F.; Fikrig, E. Anaplasma phagocytophilum induces Ixodes scapularis ticks to express an antifreeze glycoprotein gene that enhances their survival in the cold. J. Clin. Investig. 2010, 120, 3179–3190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loebermann, M.; Fingerle, V.; Lademann, M.; Fritzsche, C.; Reisinger, E.C. Borrelia burgdorferi and Anaplasma phagocytophilum coinfection. Emerg. Infect. Dis. 2006, 12, 353–355. [Google Scholar] [CrossRef]
- Ruf, T.; Bieber, C. Use of social thermoregulation fluctuates with mast seeding and reproduction in a pulsed resource consumer. Oecologia 2020, 192, 919–928. [Google Scholar] [CrossRef] [Green Version]
- Cook, A. Huddling and the control of water loss by the slug Limax pseudoflavus Evans. Anim. Behav. 1981, 29, 289–298. [Google Scholar] [CrossRef]
- Eiserer, L.A. Communal roosting in birds. Bird Behav. 1984, 5, 61–80. [Google Scholar]
- Waterman, J.M.; Roth, J.D. Interspecific associations of Cape ground squirrels with two mongoose species: Benefit or cost? Behav. Ecol. Sociobiol. 2007, 61, 1675–1683. [Google Scholar] [CrossRef]
- Ancillotto, L.; Allegrini, C.; Serangeli, M.T.; Jones, G.; Russo, D. Sociality across species: Spatial proximity of newborn bats promotes heterospecific social bonding. Behav. Ecol. 2014, 26, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Olson, M.N.; Bowman, J.; Burness, G. Social thermoregulation does not explain heterospecific nesting in North American flying squirrels. Biol. J. Linn. Soc. 2018, 123, 805–813. [Google Scholar] [CrossRef]
- Egley, R.L.; Breed, M.D. The fanner honey bee: Behavioral variability and environmental cues in workers performing a specialized task. J. Insect Behav. 2012, 26, 238–245. [Google Scholar] [CrossRef]
- Briscoe, N.J.; Handasyde, K.A.; Griffiths, S.R.; Porter, W.P.; Krockenberger, A.; Kearney, M.R. Tree-hugging koalas demonstrate a novel thermoregulatory mechanism for arboreal mammals. Biol. Lett. 2014, 10, 20140235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.-X.; Tan, K.; Radloff, S.E.; Phiancharoen, M.; Hepburn, H.R. Comb construction in mixed-species colonies of honeybees, Apis cerana and Apis mellifera. J. Exp. Biol. 2010, 213, 1659–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonhardt, S.D.; Baumann, A.-M.; Wallace, H.M.; Brooks, P.; Schmitt, T. The chemistry of an unusual seed dispersal mutualism: Bees use a complex set of olfactory cues to find their partner. Anim. Behav. 2014, 98, 41–51. [Google Scholar] [CrossRef]
- Chen, Y.P.; Pettis, J.S.; Corona, M.; Chen, W.P.; Li, C.J.; Spivak, M.; Visscher, P.K.; DeGrandi-Hoffman, G.; Boncristiani, H.; Zhao, Y.; et al. Israeli acute paralysis virus: Epidemiology, pathogenesis and implications for honey bee health. PLoS Pathog. 2014, 10, e1004261. [Google Scholar] [CrossRef]
- Pegg, G.S.; Webb, R.I.; Carnegie, A.J.; Wingfield, M.J.; Drenth, A. Infection and disease development of Quambalaria spp. on Corymbia and Eucalyptus species. Plant Pathol. 2009, 58, 642–654. [Google Scholar] [CrossRef] [Green Version]
- Meyers, M.A.; Lin, Y.S.; Olevsky, E.; Chen, P.-Y. Battle in the Amazon: Arapaima versus Piranha. Adv. Eng. Mater. 2012, 14, B279–B288. [Google Scholar] [CrossRef]
- Schuster, S. Hunting in archerfish-An ecological perspective on a remarkable combination of skills. J. Exp. Biol. 2018, 221, jeb159723. [Google Scholar] [CrossRef] [Green Version]
- Nelson, X.J.; Jackson, R.R. Aggressive use of Batesian mimicry by an ant-like jumping spider. Biol. Lett. 2009, 5, 755–757. [Google Scholar] [CrossRef] [Green Version]
- Cheney, K.L. Multiple selective pressures apply to a coral reef fish mimic: A case of Batesian–aggressive mimicry. Proc. R. Soc. B Biol. Sci. 2010, 277, 1849–1855. [Google Scholar] [CrossRef] [Green Version]
- Mougi, A.; Kishida, O.; Iwasa, Y. Coevolution of phenotypic plasticity in predator and prey: Why are inducible offenses rarer than inducible defenses? Evolution 2010, 65, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Banerji, A.; Morin, P.J. Trait-mediated apparent competition in an intraguild predator-prey system. Oikos 2013, 123, 567–574. [Google Scholar] [CrossRef]
- Campbell, S.; Harada, R.; DeFelice, S.; Bienfang, P.K.; Li, Q. Bacterial production of tetrodotoxin in the pufferfish Arothron hispidus. Nat. Prod. Res. 2009, 23, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.F.; Arakawa, O.; Saito, T.; Noguchi, T.; Simidu, U.; Tsukamoto, K.; Shida, Y.; Hashimoto, K. Tetrodotoxin-producing bacteria from the blue-ringed octopus Octopus maculosus. Mar. Biol. 1989, 100, 327–332. [Google Scholar] [CrossRef]
- Gochfeld, D.J. Territorial damselfishes facilitate survival of corals by providing an associational defense against predators. Mar. Ecol. Prog. Ser. 2010, 398, 137–148. [Google Scholar] [CrossRef] [Green Version]
- Smith, T.B.; Fong, P.; Kennison, R.; Smith, J. Spatial refuges and associational defenses promote harmful blooms of the alga Caulerpa sertularioides onto coral reefs. Oecologia 2010, 164, 1039–1048. [Google Scholar] [CrossRef]
- Steinke, M.; Malin, G.; Liss, P.S. Trophic interactions in the sea: An ecological role for climate relevant volatiles?1. J. Phycol. 2002, 38, 630–638. [Google Scholar] [CrossRef]
- Yuan, J.S.; Himanen, S.J.; Holopainen, J.K.; Chen, F.; Stewart, C.N., Jr. Smelling global climate change: Mitigation of function for plant volatile organic compounds. Trends Ecol. Evol. 2009, 24, 323–331. [Google Scholar] [CrossRef]
- Hay, M.E. Associational plant defenses and the maintenance of species diversity: Turning competitors into accomplices. Am. Nat. 1986, 128, 617–641. [Google Scholar] [CrossRef]
- Clay, K.; Holah, J. Fungal endophyte symbiosis and plant diversity in successional fields. Science 1999, 285, 1742–1744. [Google Scholar] [CrossRef]
- Rudgers, J.A.; Mattingly, W.B.; Koslow, J.M. Mutualistic fungus promotes plant invasion into diverse communities. Oecologia 2005, 144, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Rai, B.; Freedman, H.; Addicott, J.F. Analysis of three species models of mutualism in predator-prey and competitive systems. Math. Biosci. 1983, 65, 13–50. [Google Scholar] [CrossRef]
- Addicott, J.F.; Freedman, H. On the structure and stability of mutualistic systems: Analysis of predator-prey and competition models as modified by the action of a slow-growing mutualist. Theor. Popul. Biol. 1984, 26, 320–339. [Google Scholar] [CrossRef]
- van Rijn, P.C.J.; van Houten, Y.M.; Sabelis, M.W. How plants benefit from providing food to predators even when it is also edible to herbivores. Ecology 2002, 83, 2664–2679. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Janssen, A.; Kant, M.R. ECOLOGY: Enhanced: The enemy of my enemy is my ally. Science 2001, 291, 2104–2105. [Google Scholar] [CrossRef] [Green Version]
- Hay, M.E.; Parker, J.D.; Burkepile, D.E.; Caudill, C.C.; Wilson, A.E.; Hallinan, Z.P.; Chequer, A.D. Mutualisms and aquatic community structure: The enemy of my enemy is my friend. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 175–197. [Google Scholar] [CrossRef] [Green Version]
- Nishida, R. Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 2002, 47, 57–92. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.L.; Brodie, E.D., Jr.; Brodie, E.D., III. A resistant predator and its toxic prey: Persistence of newt toxin leads to poisonous (not venomous) snakes. J. Chem. Ecol. 2004, 30, 1901–1919. [Google Scholar] [CrossRef]
- Bush, S.E.; Clayton, D.H. Anti-parasite behaviour of birds. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170196. [Google Scholar] [CrossRef]
- Noë, R.; Bshary, R. The formation of red colobus-Diana monkey associations under predation pressure from chimpanzees. Proc. Biol. Sci. 1997, 264, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Semeniuk, C.A.D.; Dill, L.M. Anti-predator benefits of mixed-species groups of Cowtail Stingrays (Pastinachus sephen) and whiprays (Himantura uarnak) at rest. Ethology 2006, 112, 33–43. [Google Scholar] [CrossRef]
- Hetrick, S.A.; Sieving, K.E. Antipredator calls of tufted titmice and interspecific transfer of encoded threat information. Behav. Ecol. 2011, 23, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Marcus, M.J. Feeding associations between capybaras and jacanas: A case of interspecific grooming and possibly mutualism. Ibis 2008, 127, 240–243. [Google Scholar] [CrossRef]
- Cote, I.M. Evolution and ecology of cleaning symbioses in the sea. Oceanogr. Mar. Biol. 2000, 38, 311–355. [Google Scholar]
- Frederickson, M.E. Ant species confer different partner benefits on two neotropical myrmecophytes. Oecologia 2005, 143, 387–395. [Google Scholar] [CrossRef]
- Sabelis, M.W.; Janssen, A.; Takabayashi, J. Can plants evolve stable alliances with the enemies’ enemies? J. Plant Interact. 2011, 6, 71–75. [Google Scholar] [CrossRef]
- Kessler, A.; Baldwin, I.-T. Defensive function of herbivore-induced plant volatile emissions in nature. Science 2001, 291, 2141–2144. [Google Scholar] [CrossRef]
- Cusick, K.D.; Widder, E.A. Intensity differences in bioluminescent dinoflagellates impact foraging efficiency in a nocturnal predator. Bull. Mar. Sci. 2014, 90, 797–811. [Google Scholar] [CrossRef]
- Verpoorte, R. Secondary metabolism. In Metabolic Engineering of Plant Secondary Metabolism; Verpoorte, R., Alfermann, A.W., Eds.; Springer: Dordrecht, The Netherlands, 2000. [Google Scholar]
- Valiadi, M.; Iglesias-Rodriguez, D. Understanding bioluminescence in dinoflagellates-How far have we come? Microorganisms 2013, 1, 3–25. [Google Scholar] [CrossRef] [Green Version]
- Williams, L.; Rodriguez-Saona, C.; Del Conte, S.C.C. Methyl jasmonate induction of cotton: A field test of the ‘attract and reward’ strategy of conservation biological control. AoB Plants 2017, 9, 032. [Google Scholar] [CrossRef] [Green Version]
- Lindquist, N.; Barber, P.H.; Weisz, J.B. Episymbiotic microbes as food and defence for marine isopods: Unique symbioses in a hostile environment. Proc. Biol. Sci. 2005, 272, 1209–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proud, D.N.; Wade, R.R.; Rock, P.; Townsend, V.R., Jr.; Jiménez Chavez, D.J. Epizoic cyanobacteria associated with a Neotropical harvestman (Opiliones: Sclerosomatidae) from Costa Rica. J. Arachnol. 2012, 40, 259–261. [Google Scholar] [CrossRef]
- Vance, R.R. A mutualistic interaction between a sessile marine clam and its epibionts. Ecology 1978, 59, 679–685. [Google Scholar] [CrossRef]
- Maldonado, M.; Uriz, M.J. Relationships between sponges and crabs: Patterns of epibiosis on Inachus aguiarii (Decapoda: Majidae). Mar. Biol. 1992, 113, 281–286. [Google Scholar]
- Suutari, M.; Majaneva, M.; Fewer, D.P.; Voirin, B.; Aiello, A.; Friedl, T.; Chiarello, A.G.; Blomster, J. Molecular evidence for a diverse green algal community growing in the hair of sloths and a specific association with Trichophilus welckeri (Chlorophyta, Ulvophyceae). BMC Evol. Biol. 2010, 10, 86. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, A.G. The role of the extrafloral nectaries of catalpa speciosa in limiting herbivory and increasing fruit production. Ecology 1982, 63, 663–669. [Google Scholar] [CrossRef]
- Chanam, J.; Borges, R.M. Cauline domatia of the ant-plant Humboldtia brunonis (Fabaceae). Flora Morphol. Distrib. Funct. Ecol. Plants 2017, 236, 58–66. [Google Scholar] [CrossRef]
- Fonseca-Romero, M.A.; Fornoni, J.; Del-Val, E.; Boege, K. Ontogenetic trajectories of direct and indirect defenses of myrmecophytic plants colonized either by mutualistic or opportunistic ant species. Oecologia 2019, 190, 857–865. [Google Scholar] [CrossRef]
- Stiefel, K.M.; Reyes, R.B., Jr. The marine goby-Alpheid shrimp symbiosis does not correlate with larger fish eye size. BioRxiv 2018. [Google Scholar] [CrossRef]
- Mouritsen, K.N.; Poulin, R. The mud flat anemone-cockle association: Mutualism in the intertidal zone? Oecologia 2003, 135, 131–137. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, K.D. The evolution of trophic transmission. Parasitol. Today 1999, 15, 111–115. [Google Scholar] [CrossRef]
- Fenton, A.; Rands, S.A. The impact of parasite manipulation and predator foraging behavior on predator-prey communities. Ecology 2006, 87, 2832–2841. [Google Scholar] [CrossRef]
- Schneider, D.S.; James, A.A. Bridging the gaps in vector biology. EMBO Rep. 2006, 7, 259–262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chow, Y.; Jang, S.; Wang, H. Cooperative hunting in a discrete predator–prey system II. J. Biol. Dyn. 2018, 13, 247–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grinsted, L.; Schou, M.F.; Settepani, V.; Holm, C.; Bird, T.L.; Bilde, T. Prey to predator body size ratio in the evolution of cooperative hunting-A social spider test case. Dev. Genes Evol. 2019, 230, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Mlot, C. A Soil Story. Sci. News 1997, 152, 58–59. [Google Scholar] [CrossRef]
- Chung, S.H.; Rosa, C.; Scully, E.D.; Peiffer, M.; Tooker, J.F.; Hoover, K.; Luthe, D.S.; Felton, G.W. Herbivore exploits orally secreted bacteria to suppress plant defenses. Proc. Natl. Acad. Sci. USA 2013, 110, 15728–15733. [Google Scholar] [CrossRef] [Green Version]
- Shiga, D. Poisonous partnership: Parasitoid wasps use viruses as a weapon. Sci. News 2005, 167, 136–137. [Google Scholar] [CrossRef]
- Dowd, P.F.; Shen, S.K. The contribution of symbiotic yeast to toxin resistance of the cigarette beetle (Lasioderma serricorne). Entomol. Exp. Appl. 1990, 56, 241–248. [Google Scholar] [CrossRef]
- Kohl, K.D.; Connelly, J.W.; Dearing, M.D.; Forbey, J.S. Microbial detoxification in the gut of a specialist avian herbivore, the Greater Sage-Grouse. FEMS Microbiol. Lett. 2016, 363, 144. [Google Scholar] [CrossRef] [Green Version]
- Hulet, W.H.; Musil, G. Intracellular bacteria in the light organ of the deep sea angler fish, Melanocetus murrayi. Copeia 2006, 1968, 506. [Google Scholar] [CrossRef]
- Widder, E.A. A predatory use of counterillumination by the squaloid shark, Isistius brasiliensis. Environ. Boil. Fishes 1998, 53, 267–273. [Google Scholar] [CrossRef]
- Jones, B.W.; Nishiguchi, M.K. Counterillumination in the Hawaiian bobtail squid, Euprymna scolopes Berry (Mollusca: Cephalopoda). Mar. Biol. 2004, 144, 1151–1155. [Google Scholar] [CrossRef]
- Herring, P.J.; Cope, C. Red bioluminescence in fishes: On the suborbital photophores of Malacosteus, Pachystomias and Aristostomias. Mar. Biol. 2005, 148, 383–394. [Google Scholar] [CrossRef]
- Sutton, T. Trophic ecology of the deep-sea fish Malacosteus niger (Pisces: Stomiidae): An enigmatic feeding ecology to facilitate a unique visual system? Deep. Sea Res. Part I Oceanogr. Res. Pap. 2005, 52, 2065–2076. [Google Scholar] [CrossRef] [Green Version]
- Dean, W.R.J.; Siegfried, W.R.; Macdonald, I.A.W. The fallacy, fact, and fate of guiding behavior in the greater honeyguide. Conserv. Biol. 1990, 4, 99–101. [Google Scholar] [CrossRef]
- Minta, S.C.; Minta, K.A.; Lott, D.F. Hunting associations between badgers (Taxidea taxus) and coyotes (Canis latrans). J. Mammal. 1992, 73, 814–820. [Google Scholar] [CrossRef]
- Bshary, R.; Hohner, A.; Ait-El-Djoudi, K.; Fricke, H. Interspecific communicative and coordinated hunting between groupers and giant moray eels in the Red Sea. PLoS Biol. 2006, 4, e431. [Google Scholar] [CrossRef]
- D’Angelo, G.B.; Sazima, I. Commensal association of piscivorous birds with foraging otters in southeastern Brazil, and a comparison with such a relationship of piscivorous birds with cormorants. J. Nat. Hist. 2013, 48, 241–249. [Google Scholar] [CrossRef]
- Lönnstedt, O.M.; Ferrari, M.C.O.; Chivers, D.P. Lionfish predators use flared fin displays to initiate cooperative hunting. Biol. Lett. 2014, 10, 20140281. [Google Scholar] [CrossRef]
- Sridhar, H.; Guttal, V. Friendship across species borders: Factors that facilitate and constrain heterospecific sociality. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170014. [Google Scholar] [CrossRef] [Green Version]
- Wilson, D.S.; Knollenberg, W.G. Adaptive indirect effects: The fitness of burying beetles with and without their phoretic mites. Evol. Ecol. 1987, 1, 139–159. [Google Scholar] [CrossRef]
- Krishnan, A.; Muralidharan, S.; Sharma, L.; Borges, R.M. A hitchhiker’s guide to a crowded syconium: How do fig nematodes find the right ride? Funct. Ecol. 2010, 24, 741–749. [Google Scholar] [CrossRef]
- Badets, M.; Du Preez, L. Phoretic interaction between the kangaroo leech Marsupiobdella africana (Hirudinea: Glossiphoniidae) and the cape river crab Potamonautes perlatus (Decapoda: Potamonautidae). Int. J. Parasitol. Parasites Wildl. 2014, 3, 6–11. [Google Scholar] [CrossRef] [Green Version]
- McLeod, G.; Gries, R.; Von Reuss, S.H.; Rahe, J.E.; McIntosh, R.; König, W.A.; Gries, G. The pathogen causing Dutch elm disease makes host trees attract insect vectors. Proc. Biol. Sci. 2005, 272, 2499–2503. [Google Scholar] [CrossRef]
- Robinson, A.; Busula, A.O.; Voets, M.A.; Beshir, K.B.; Caulfield, J.C.; Powers, S.J.; Verhulst, N.O.; Winskill, P.; Muwanguzi, J.; Birkett, M.A.; et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, E4209–E4218. [Google Scholar] [CrossRef] [Green Version]
- de Vega, C.; Arista, M.; Ortiz, P.L.; Talavera, S. Anatomical relations among endophytic holoparasitic angiosperms, autotrophic host plants and mycorrhizal fungi: A novel tripartite interaction. Am. J. Bot. 2010, 97, 730–737. [Google Scholar] [CrossRef]
- Anderson, B. Adaptations to foliar absorption of faeces: A pathway in plant carnivory. Ann. Bot. 2005, 95, 757–761. [Google Scholar] [CrossRef] [Green Version]
- Jones, E.C. Tremoctopus violaceus uses physalia tentacles as weapons. Science 1963, 139, 764–766. [Google Scholar] [CrossRef]
- Tauber, C.A.; De León, T.; Penny, N.D.; Tauber, M.J. The Genus Ceraeochrysa (Neuroptera: Chrysopidae) of America north of Mexico: Larvae, adults, and comparative biology. Ann. Entomol. Soc. Am. 2000, 93, 1195–1221. [Google Scholar] [CrossRef]
- Robinson, S.K. Use of bait and lures by green-backed herons in Amazonian Peru. Wilson Bull. 1994, 106, 567–569. [Google Scholar]
- Krützen, M.; Kreicker, S.; MacLeod, C.D.; Learmonth, J.; Kopps, A.M.; Walsham, P.; Allen, S.J. Cultural transmission of tool use by Indo-Pacific bottlenose dolphins (Tursiops sp.) provides access to a novel foraging niche. Proc. Biol. Sci. 2014, 281, 20140374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefèvre, T.; Lebarbenchon, C.; Gauthier-Clerc, M.; Missé, D.; Poulin, R.; Thomas, F. The ecological significance of manipulative parasites. Trends Ecol. Evol. 2009, 24, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Poulin, R. Chapter 5-Parasite manipulation of host behavior: An update and frequently asked questions. Adv. Stud. Behav. 2010, 41, 151–186. [Google Scholar]
- Nedergaard, O.A. Curare: The flying death. Pharmacol. Toxicol. 2003, 92, 154–155. [Google Scholar] [CrossRef]
- Myers, C.W.; Daly, J.W.; Malkin, B. A dangerously toxic new frog (Phyllobates) used by Emberá Indians of western Colombia, with discussion of blowgun fabrication and dart poisoning. Bull. Am. Mus. Nat. Hist. 1978, 161. Available online: http://hdl.handle.net/2246/1286 (accessed on 6 January 2021).
- Spottiswoode, C.N.; Begg, K.S.; Begg, C.M. Reciprocal signaling in honeyguide-human mutualism. Science 2016, 353, 387–389. [Google Scholar] [CrossRef] [Green Version]
- Koster, J. The impact of hunting with dogs on wildlife harvests in the Bosawas Reserve, Nicaragua. Environ. Conserv. 2008, 35, 211–220. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, M.J.; Surovell, T.A. Spatial expression of kinship among the Dukha reindeer herders of Northern Mongolia. Arct. Anthr. 2017, 54, 110–119. [Google Scholar] [CrossRef]
- Park, K.J.; Lee, M.J.; Kang, H.; Kim, K.S.; Lee, S.-H.; Cho, I.; Lee, H.H. Saeng-Maek-San, a medicinal herb complex, protects liver cell damage induced by alcohol. Biol. Pharm. Bull. 2002, 25, 1451–1455. [Google Scholar] [CrossRef] [Green Version]
- Jaeschke, H.; Williams, C.D.; McGill, M.R.; Farhood, A. Herbal extracts as hepatoprotectants against acetaminophen hepatotoxicity. World J. Gastroenterol. 2010, 16, 2448–2450. [Google Scholar] [CrossRef] [PubMed]
- Sherman, P.W.; Flaxman, S.M. Protecting ourselves from food: Spices and morning sickness may shield us from toxins and microorganisms in the diet. Am. Sci. 2001, 89, 142–151. [Google Scholar] [CrossRef]
- Marini, E.; Magi, G.; Mingoia, M.; Pugnaloni, A.; Facinelli, B. Antimicrobial and anti-virulence activity of capsaicin against erythromycin-resistant, cell-invasive group a Streptococci. Front. Microbiol. 2015, 6, 1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lettieri, J.T.; Dain, B. Effects of beano on the tolerability and pharmacodynamics of acarbose. Clin. Ther. 1998, 20, 497–504. [Google Scholar] [CrossRef]
- Czerucka, D.; Piche, T.; Rampal, P. Review article: Yeast as probiotics-Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2007, 26, 767–778. [Google Scholar] [CrossRef]
- Burton, K.J.; Rosikiewicz, M.; Pimentel, G.; Bütikofer, U.; von Ah, U.; Voirol, M.-J.; Croxatto, A.; Aeby, S.; Drai, J.; McTernan, P.G.; et al. Probiotic yogurt and acidified milk similarly reduce postprandial inflammation and both alter the gut microbiota of healthy, young men. Br. J. Nutr. 2017, 117, 1312–1322. [Google Scholar] [CrossRef] [Green Version]
- Beukemaj, J.J. Acquired hook-avoidance in the pike Esox lucius L. fished with artificial and natural baits. J. Fish Biol. 1970, 2, 155–160. [Google Scholar] [CrossRef]
- Drake, D.A.R.; Mandrak, N.E. Ecological risk of live bait fisheries: A new angle on selective fishing. Fisheries 2014, 39, 201–211. [Google Scholar] [CrossRef]
- Daura-Jorge, F.G.; Cantor, M.; Ingram, S.N.; Lusseau, S.M.; Simões-Lopes, P.C. The structure of a bottlenose dolphin society is coupled to a unique foraging cooperation with artisanal fishermen. Biol. Lett. 2012, 8, 702–705. [Google Scholar] [CrossRef] [Green Version]
- Talou, T.; Gaset, A.; Delmas, M.; Kulifaj, M.; Montant, C. Dimethyl sulphide: The secret for black truffle hunting by animals? Mycol. Res. 1990, 94, 277–278. [Google Scholar] [CrossRef]
- Ruslin, F.; Azmi, M.A.; Matsuda, I.; Amir, R.; Md-Zain, B.M. Monkey school: Training phases for coconut-picking macaques (Macaca nemestrina). Malay. Nat. J. 2017, 69, 301–306. [Google Scholar]
- Marcone, M. Composition and properties of Indonesian palm civet coffee (Kopi Luwak) and Ethiopian civet coffee. Food Res. Int. 2004, 37, 901–912. [Google Scholar] [CrossRef]
- Pasha, C.; Reddy, G. Nutritional and medicinal improvement of black tea by yeast fermentation. Food Chem. 2005, 89, 449–453. [Google Scholar] [CrossRef]
- Schwan, R.F.; Wheals, A.E. The microbiology of cocoa fermentation and its role in chocolate quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M. Yeasts in botrytized wine making. In Yeasts in the Production of Wine; Romano, P., Ciani, M., Fleet, G., Eds.; Springer: New York, NY, USA, 2019. [Google Scholar]
- Zeckhauser, R.J. Human hunters and nonhuman predators: Fundamental differences. Proc. Natl. Acad. Sci. USA 2016, 114, 13–14. [Google Scholar] [CrossRef] [Green Version]
- Dicke, M.; Sabelis, M.W. Infochemical terminology: Based on cost-benefit analysis rather than origin of compounds? Funct. Ecol. 1988, 2, 131. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.C.; Di Bitetti, M.S.; Janson, C.; Slobodkin, L.B.; Valenzuela, N. An “Audience effect” for ecological terminology: Use and misuse of jargon. Oikos 1997, 80, 632. [Google Scholar] [CrossRef] [Green Version]
- Queller, D.C.; Strassmann, J.E. Problems of multi-species organisms: Endosymbionts to holobionts. Biol. Philos. 2016, 31, 855–873. [Google Scholar] [CrossRef] [Green Version]
- Michálek, O.; Kuhn-Nentwig, L.; Natarajan, R. High specific efficiency of venom of two prey-specialized spiders. Toxins 2019, 11, 687. [Google Scholar] [CrossRef] [Green Version]
- Yadav, S.; Shokal, U.; Forst, S.; Eleftherianos, I. An improved method for generating axenic entomopathogenic nematodes. BMC Res. Notes 2015, 8, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Persoons, A.; Hayden, K.J.; Fabre, B.; Frey, P.; De Mita, S.; Tellier, A.; Halkett, F. The escalatory Red Queen: Population extinction and replacement following arms race dynamics in poplar rust. Mol. Ecol. 2017, 26, 1902–1918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strotz, L.C.; Simões, M.; Girard, M.G.; Breitkreuz, L.; Kimmig, J.; Lieberman, B.S. Getting somewhere with the Red Queen: Chasing a biologically modern definition of the hypothesis. Biol. Lett. 2018, 14, 20170734. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Atamian, H.S.; Shen, Z.; Briggs, S.P.; Kaloshian, I. GroEL from the endosymbiont Buchnera aphidicola betrays the aphid by triggering plant defense. Proc. Natl. Acad. Sci. USA 2014, 111, 8919–8924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skowronek, M.; Sajnaga, E.; Pleszczyńska, M.; Kazimierczak, W.; Lis, M.; Wiater, A. Bacteria from the midgut of common cockchafer (Melolontha melolontha L.) larvae exhibiting antagonistic activity against bacterial symbionts of entomopathogenic nematodes: Isolation and molecular identification. Int. J. Mol. Sci. 2020, 21, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polis, G.A.; Sears, A.L.; Huxel, G.R.; Strong, D.R.; Maron, J. When is a trophic cascade a trophic cascade? Trends Ecol. Evol. 2000, 15, 473–475. [Google Scholar] [CrossRef]
- Fortin, D.; Beyer, H.L.; Boyce, M.S.; Smith, D.W.; Duchesne, T.; Mao, J.S. Wolves influence elk movements: Behavior shapes a trophic cascade in Yellowstone National Park. Ecology 2005, 86, 1320–1330. [Google Scholar] [CrossRef] [Green Version]
- Berger, J. Anthropogenic extinction of top carnivores and interspecific animal behaviour: Implications of the rapid decoupling of a web involving wolves, bears, moose and ravens. Proc. Biol. Sci. 1999, 266, 2261–2267. [Google Scholar] [CrossRef] [Green Version]
- Stahler, D.; Heinrich, B.; Smith, D. Common ravens, Corvus corax, preferentially associate with grey wolves, Canis lupus, as a foraging strategy in winter. Anim. Behav. 2002, 64, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Despret, V. The enigma of the raven. Angelaki 2015, 20, 57–72. [Google Scholar] [CrossRef]
- Walker, L.E.; Marzluff, J.M.; Metz, M.C.; Wirsing, A.J.; Moskal, L.M.; Stahler, D.R.; Smith, D.W. Population responses of common ravens to reintroduced gray wolves. Ecol. Evol. 2018, 8, 11158–11168. [Google Scholar] [CrossRef] [Green Version]
- Galasso, N.M.; Bonaviri, C.; Di Trapani, F.; Picciotto, M.; Gianguzza, P.; Agnetta, D.; Badalamenti, F. Fish-seastar facilitation leads to algal forest restoration on protected rocky reefs. Sci. Rep. 2015, 5, srep12409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myerscough, M.R.; Darwen, M.; Hogarth, W. Stability, persistence and structural stability in a classical predator-prey model. Ecol. Model. 1996, 89, 31–42. [Google Scholar] [CrossRef]
- Samways, M.J. Asymmetrical competition and amensalism through soil dumping by the ant, Myrmicaria natalensis. Ecol. Entomol. 1983, 8, 191–194. [Google Scholar] [CrossRef]
- Hulme-Beaman, A.; Dobney, K.; Cucchi, T.; Searle, J.B. An ecological and evolutionary framework for commensalism in anthropogenic environments. Trends Ecol. Evol. 2016, 31, 633–645. [Google Scholar] [CrossRef] [Green Version]
- Leung, T.L.F.; Poulin, R. Parasitism, commensalism, and mutualism: Exploring the many shades of symbioses. Life Environ. 2008, 58, 107–115. [Google Scholar]
- Dutton, E.M.; Luo, E.Y.; Cembrowski, A.R.; Shore, J.S.; Frederickson, M.E. Three’s a crowd: Trade-offs between attracting pollinators and ant bodyguards with nectar rewards in Turnera. Am. Nat. 2016, 188, 38–51. [Google Scholar] [CrossRef]
- Shankar, U. Predation risk of pollinators on flowers. Bee World 2015, 92, 84–85. [Google Scholar] [CrossRef]
- Webster, M.S. Interspecific brood parasitism of Montezuma oropendolas by giant cowbirds: Parasitism or mutualism? Condor 1994, 96, 794–798. [Google Scholar] [CrossRef]
- Greenwood, P.G. Acquisition and use of nematocysts by cnidarian predators. Toxicon 2009, 54, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Janzen, D.H. Allelopathy by myrmecophytes: The ant azteca as an allelopathic agent of cecropia. Ecology 2006, 50, 147–153. [Google Scholar] [CrossRef]
- Anderson, B.; Midgley, J.J.; Stewart, B.A. Facilitated selfing offers reproductive assurance: A mutualism between a hemipteran and carnivorous plant. Am. J. Bot. 2003, 90, 1009–1015. [Google Scholar] [CrossRef] [PubMed]
- Hountondji, F.C.; Sabelis, M.W.; Hanna, R.; Janssen, A. Herbivore-induced plant volatiles trigger sporulation in entomopathogenic fungi: The case of Neozygites tanajoae infecting the cassava green mite. J. Chem. Ecol. 2005, 31, 1003–1021. [Google Scholar] [CrossRef] [PubMed]
- Loudon, A.H.; Holland, J.A.; Umile, T.P.; Burzynski, E.A.; Minbiole, K.P.C.; Harris, R.N. Interactions between amphibians’ symbiotic bacteria cause the production of emergent anti-fungal metabolites. Front. Microbiol. 2014, 5, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, M.C.; Garner, T.W.J. Chytrid fungi and global amphibian declines. Nat. Rev. Genet. 2020, 18, 332–343. [Google Scholar] [CrossRef]
- Saikkonen, K.; Saari, S.; Helander, M. Defensive mutualism between plants and endophytic fungi? Fungal Divers. 2010, 41, 101–113. [Google Scholar] [CrossRef]
- Karnosky, D.F. Dutch elm disease: A review of the history, environmental implications, control, and research needs. Environ. Conserv. 1979, 6, 311–322. [Google Scholar] [CrossRef]
- Wilde, S.B.; Johansen, J.R.; Wilde, H.D.; Jiang, P.; Bartelme, B.; Haynie, R.S. Aetokthonos hydrillicola gen. et sp. nov.: Epiphytic cyanobacteria on invasive aquatic plants implicated in Avian Vacuolar Myelinopathy. Phytotaxa 2014, 181, 243–260. [Google Scholar] [CrossRef] [Green Version]
- Fleming, J.-A.G. The integration of polydnavirus genomes in parasitoid genomes: Implications for biocontrol and genetic analyses of parasitoid wasps. Biol. Control. 1991, 1, 127–135. [Google Scholar] [CrossRef]
- Georgis, R.; Gaugler, R. Predictability in biological control using entomopathogenic nematodes. J. Econ. Entomol. 1991, 84, 713–720. [Google Scholar] [CrossRef]
- Orzech, K.M.; Nichter, M. From resilience to resistance: Political ecological lessons from antibiotic and pesticide resistance. Annu. Rev. Anthr. 2008, 37, 267–282. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Hayatsu, M.; Hosokawa, T.; Nagayama, A.; Tago, K.; Fukatsu, T. Symbiont-mediated insecticide resistance. Proc. Natl. Acad. Sci. USA 2012, 109, 8618–8622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Liu, Y.; Graham, R.I.; Wilson, K.; Wu, K. Densovirus is a mutualistic symbiont of a global crop pest (Helicoverpa armigera) and protects against a baculovirus and Bt biopesticide. PLoS Pathog. 2014, 10, e1004490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauldin, J.K.; Carter, F.L.; Rich, N.M. Protozoan populations of Reticulitermes flavipes (Kollar) exposed to heart-wood blocks of 21 American species. Mater. Org. 1981, 16, 15–28. [Google Scholar]
- Waller, D.A. Ampicillin, tetracycline and urea as protozoicides for symbionts of Reticulitermes flavipes and R. virginicus (Isoptera: Rhinotermitidae). Bull. Entomol. Res. 1996, 86, 77–81. [Google Scholar] [CrossRef]
- Esenther, G.R.; Beal, R.H. Termite control: Decayed wood bait. Sociobiology 1979, 4, 215–222. [Google Scholar]
- Tarasco, E.; Alvarez, C.S.; Triggiani, O.; Moraga, E.Q. Laboratory studies on the competition for insect haemocoel between Beauveria bassiana and Steinernema ichnusae recovered in the same ecological niche. Biol. Control. Sci. Technol. 2011, 21, 693–704. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Thönen, L.; Arce, C.C.M.; Theepan, V.; Prada, F.; Wüthrich, D.; Robert, C.A.M.; Vogiatzaki, E.; Shi, Y.-M.; Schaeren, O.P.; et al. Engineering bacterial symbionts of nematodes improves their biocontrol potential to counter the western corn rootworm. Nat. Biotechnol. 2020, 38, 600–608. [Google Scholar] [CrossRef]
- Karban, R.; Hougen-Eitzmann, D.; English-Loeb, G. Predator-mediated apparent competition between two herbivores that feed on grapevines. Oecologia 1994, 97, 508–511. [Google Scholar] [CrossRef]
- Faist, A.; Antoninka, A.; Belnap, J.; Bowker, M.A.; Duniway, M.C.; Garcia-Pichel, F.; Nelson, C.; Reed, S.C.; Giraldo-Silva, A.; Ayuso, S.V.; et al. Inoculation and habitat amelioration efforts in biological soil crust recovery vary by desert and soil texture. Restor. Ecol. 2020, 28. [Google Scholar] [CrossRef]
- Temple, S.A. Plant-animal mutualism: Coevolution with dodo leads to near extinction of plant. Science 1977, 197, 885–886. [Google Scholar] [CrossRef]
- Witmer, M.C.; Cheke, A.S. The Dodo and the Tambalacoque tree: An obligate mutualism reconsidered. Oikos 1991, 61, 133. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.Y.; Collinson, M.E.; Rudall, P.J.; Simpson, D.A.; Marone, F.; Stampanoni, M. Virtual taphonomy using synchrotron tomographic microscopy reveals cryptic features and internal structure of modern and fossil plants. Proc. Natl. Acad. Sci. USA 2009, 106, 12013–12018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinseel, E.; Vesela, J.; Scharfen, V.; Urbánková, P.; Van De Vijver, B.; Vyverman, W. Extensive Cryptic Diversity in the Terrestrial Diatom Pinnularia borealis (Bacillariophyceae). Protist 2018, 170, 121–140. [Google Scholar] [CrossRef]
- Hahn, A.S.; Konwar, K.M.; Louca, S.; Hanson, N.W.; Hallam, S.J. The information science of microbial ecology. Curr. Opin. Microbiol. 2016, 31, 209–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roumpeka, D.D.; Wallace, R.J.; Escalettes, F.; Fotheringham, I.; Watson, M. A review of bioinformatics tools for bio-prospecting from metagenomic sequence data. Front. Genet. 2017, 8, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagley, M.; Pilgrim, E.; Knapp, M.; Yoder, C.; Domingo, J.S.; Banerji, A. High-throughput environmental DNA analysis informs a biological assessment of an urban stream. Ecol. Indic. 2019, 104, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Titcomb, G.; Jerde, C.L.; Young, H.S. High-throughput sequencing for understanding the ecology of emerging infectious diseases at the wildlife-human interface. Front. Ecol. Evol. 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- van Tienderen, P.H.; de Haan, A.A.; van der Linden, C.G.; Vosman, B. Biodiversity assessment using markers for ecologically important traits. Trends Ecol. Evol. 2002, 17, 577–582. [Google Scholar] [CrossRef]
- Dinsdale, E.A.; Edwards, R.A.; Hall, D.; Angly, F.; Breitbart, M.; Brulc, J.M.; Furlan, M.; Desnues, C.; Haynes, M.; Li, L.; et al. Functional metagenomic profiling of nine biomes. Nat. Cell Biol. 2008, 452, 629–632. [Google Scholar] [CrossRef]
- Peršoh, D. Plant-associated fungal communities in the light of meta’omics. Fungal Divers. 2015, 75, 1–25. [Google Scholar] [CrossRef]
- Porras-Alfaro, A.; Bayman, P. Hidden fungi, emergent properties: Endophytes and microbiomes. Annu. Rev. Phytopathol. 2011, 49, 291–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grube, M.; Cernava, T.; Soh, J.; Fuchs, S.; Aschenbrenner, I.A.; Lassek, C.; Wegner, U.; Becher, D.; Riedel, K.; Sensen, C.W.; et al. Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. ISME J. 2015, 9, 412–424. [Google Scholar] [CrossRef] [Green Version]
- Scherlach, K.; Hertweck, C. Mediators of mutualistic microbe–microbe interactions. Nat. Prod. Rep. 2018, 35, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Arango-Argoty, G.; Dai, D.; Pruden, A.; Vikesland, P.J.; Heath, L.S.; Zhang, W.-F. NanoARG: A web service for detecting and contextualizing antimicrobial resistance genes from nanopore-derived metagenomes. Microbiome 2019, 7, 88. [Google Scholar] [CrossRef] [PubMed]
- Kildegaard, H.F.; Baycin-Hizal, D.; Lewis, N.E.; Bettenbaugh, M.J. The emerging CHO systems biology era: Harnessing the ‘omics revolution for biotechnology. Curr. Opin. Biotechnol. 2013, 24, 1102–1107. [Google Scholar] [CrossRef] [PubMed]
- Pirih, N.; Kunej, T. Toward a taxonomy for multi-omics science? Terminology development for whole genome study approaches by omics technology and hierarchy. OMICS A J. Integr. Biol. 2017, 21, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Zorin, A.; Dass, G.; Vu, M.-T.; Xu, P.; Glont, M.; Vizcaíno, J.A.; Jarnuczak, A.F.; Petryszak, R.; Ping, P.; et al. Quantifying the impact of public omics data. Nat. Commun. 2019, 10, 3512. [Google Scholar] [CrossRef] [Green Version]
- Ratnaparkhe, A.; Tiwari, A. Bio-prospecting approach for biocatalysts from extremophiles. Int. J. Biomed. Res. 2011, 2, 1–17. [Google Scholar] [CrossRef] [Green Version]
- McGill, B.J.; Enquist, B.J.; Weiher, E.; Westoby, M. Rebuilding community ecology from functional traits. Trends Ecol. Evol. 2006, 21, 178–185. [Google Scholar] [CrossRef]
- Verberk, W.C.E.P.; Van Noordwijk, C.G.E.; Hildrew, A.G. Delivering on a promise: Integrating species traits to transform descriptive community ecology into a predictive science. Freshw. Sci. 2013, 32, 531–547. [Google Scholar] [CrossRef] [Green Version]
- Jones, M.; Twieg, B.D.; Ward, V.; Barker, J.; Durall, D.M.; Simard, S.W. Functional complementarity of Douglas-fir ectomycorrhizas for extracellular enzyme activity after wildfire or clearcut logging. Funct. Ecol. 2010, 24, 1139–1151. [Google Scholar] [CrossRef]
- Blüthgen, N.; Klein, A.-M. Functional complementarity and specialisation: The role of biodiversity in plant-pollinator interactions. Basic Appl. Ecol. 2011, 12, 282–291. [Google Scholar] [CrossRef]
Figure 1.
Competitive interactions. (A) In closed-canopy forests, overstory trees engage in “preemption” against understory herbs and shrubs by blocking sunlight [28]. (B) The ring-tailed lemur (Lemur catta L.) engages in “territoriality” by using its scent-producing glands to broadcast olfactory signals [29]. (C) The hydrocoral Millepora alcicornis L. engages in “overgrowth” when it encrusts arborescent octocorals of the genus Gorgonia [30]. (D) Garlic mustard engages in “allelopathy” when it inhibits germination of competing plants by releasing toxic root exudates into the soil [31].
Figure 1.
Competitive interactions. (A) In closed-canopy forests, overstory trees engage in “preemption” against understory herbs and shrubs by blocking sunlight [28]. (B) The ring-tailed lemur (Lemur catta L.) engages in “territoriality” by using its scent-producing glands to broadcast olfactory signals [29]. (C) The hydrocoral Millepora alcicornis L. engages in “overgrowth” when it encrusts arborescent octocorals of the genus Gorgonia [30]. (D) Garlic mustard engages in “allelopathy” when it inhibits germination of competing plants by releasing toxic root exudates into the soil [31].

Figure 2.
Mutualistic interactions. (A) Developing embryos of the spotted salamander (Ambystoma maculatum Shaw) share a mutually beneficial endosymbiosis with green algae, the likes of which have yet to be found in any other terrestrial vertebrate [109]. (B) The sugarbag bee (Tetragonula carbonaria Smith) disperses the seeds of the cadaghi tree (Corymbia torelliana Hill and Johnson) in return for resin, with which it can construct and waterproof its spiral hive [137].
Figure 2.
Mutualistic interactions. (A) Developing embryos of the spotted salamander (Ambystoma maculatum Shaw) share a mutually beneficial endosymbiosis with green algae, the likes of which have yet to be found in any other terrestrial vertebrate [109]. (B) The sugarbag bee (Tetragonula carbonaria Smith) disperses the seeds of the cadaghi tree (Corymbia torelliana Hill and Johnson) in return for resin, with which it can construct and waterproof its spiral hive [137].

Figure 3.
Defenses and offenses among resources and consumers. (A) The durable scales of the arapaima (Arapaima gigas Schinz) provide an endogenous defense against predators such as piranhas (Serrasalminae) [140]. (B) The archer fish (Toxotes jaculatrix Pallas) uses its tongue and gills as an endogenous offense to shoot down insect prey [141]. (C) The endosymbiont-produced tetrodotoxin of the pufferfish (Arothron hispidus L.) is an associational defense against predators [146]. (D) The endosymbiont-produced tetrodotoxin of blue-ringed octopuses (Hapalochlaena spp.) is an associational offense the octopuses use to envenomate their prey [147].
Figure 3.
Defenses and offenses among resources and consumers. (A) The durable scales of the arapaima (Arapaima gigas Schinz) provide an endogenous defense against predators such as piranhas (Serrasalminae) [140]. (B) The archer fish (Toxotes jaculatrix Pallas) uses its tongue and gills as an endogenous offense to shoot down insect prey [141]. (C) The endosymbiont-produced tetrodotoxin of the pufferfish (Arothron hispidus L.) is an associational defense against predators [146]. (D) The endosymbiont-produced tetrodotoxin of blue-ringed octopuses (Hapalochlaena spp.) is an associational offense the octopuses use to envenomate their prey [147].

Figure 4.
A trophic cascade mediated by an associational offense. Here, the influence of the sea star benefits the wrasse in terms of one or more of the phases of consumption (shown on the left) in return for a portion of the kill. Solid blue arrows denote direct positive (+) and negative (-) interspecific effects, while dashed arrows represent indirect effects.
Figure 4.
A trophic cascade mediated by an associational offense. Here, the influence of the sea star benefits the wrasse in terms of one or more of the phases of consumption (shown on the left) in return for a portion of the kill. Solid blue arrows denote direct positive (+) and negative (-) interspecific effects, while dashed arrows represent indirect effects.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Banerji, A. Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications. Ecologies 2021, 2, 43-70. https://0-doi-org.brum.beds.ac.uk/10.3390/ecologies2010004
AMA Style
Banerji A. Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications. Ecologies. 2021; 2(1):43-70. https://0-doi-org.brum.beds.ac.uk/10.3390/ecologies2010004
Chicago/Turabian StyleBanerji, Aabir. 2021. "Acquisition of Adaptive Traits via Interspecific Association: Ecological Consequences and Applications" Ecologies 2, no. 1: 43-70. https://0-doi-org.brum.beds.ac.uk/10.3390/ecologies2010004