
Xanthine Oxidase Inhibition by Aqueous Extract of Limonium brasiliense (Plumbaginaceae) †



1
Departamento de Química, INQUISUR, UNS, B8000CPB Bahía Blanca, Argentina
2
Laboratorio de Parasitología, IIAA, USC, E-15782 Santiago de Compostela, Spain
*
Author to whom correspondence should be addressed.
†
Presented at the 24th International Electronic Conference on Synthetic Organic Chemistry, 15 November–15 December 2020; Available online: https://ecsoc-24.sciforum.net/ .
Chem. Proc. 2021, 3(1), 123; https://0-doi-org.brum.beds.ac.uk/10.3390/ecsoc-24-08410
Published: 14 December 2020
(This article belongs to the Proceedings of The 24th International Electronic Conference on Synthetic Organic Chemistry)
Abstract
:Limonium brasiliense (Boiss.) Kuntze (Plumbaginaceae) is a medicinal plant that is popularly used in the folk medicine. In this study, we examined xanthine oxidase (XO) inhibitory activity of the aqueous fraction of Limonium brasiliense extract and its polar bioactive constituents using the enzymatic hypoxanthine (HX)–XO system in vitro. Seven active compounds isolated from the aqueous extract proved to be strong XO inhibitors. Prodelphinidin B1-3,3′-digallate (1) showed the most potent inhibitory activity with an IC50 value of 6.61 µM, comparable to allopurinol. This study provides basis for the potential use of the aqueous fraction of L. brasiliense against hyperuricemia, displaying potent XO inhibitory activity with an IC50 value of 48.3 µg/mL. This inhibitory effect is compatible with the presence of (1) in this extract.
1. Introduction
Xanthine oxidase (XO) is an enzyme that catalyzes the oxidation of hypoxanthine and xanthine to uric acid; the inhibition of xanthine oxidase decreases uric acid accumulation in the joints, controlling the disease called gout, and is relevant for the treatment of hyperuricemia. Currently, gout is treated with the drug allopurinol (hypoxanthine analog). Prolonged use of allopurinol leads to adverse effects for health, since it controls but does not eliminate the uric acid overproduction [1], making it necessary to find new alternatives for control and gout prevention. XO inhibitors are also being investigated for management of reperfusion injury.
Oxidative stress, defined as an imbalance between levels of various oxidant molecules as reactive oxygen species (ROS) and antioxidants, leads to many biochemical changes and, consequently, serious disorders in the human organism. Oxidative stress can result in damage of biomolecules, such as lipids, proteins, and DNA, leading to cytotoxic and genotoxic effects. Although ROS are involved in host defense, overproduction of these species may contribute to the pathogenesis of inflammatory diseases and other immunopathological phenomena related to oxidative stress, such as rheumatoid arthritis, myocardial reperfusion injury, respiratory disorders, hepatocellular oxidative DNA injury, and pathogenesis of CNS neurons [2,3,4,5,6,7].
The inhibitory activity of xanthine oxidase is a combination of uric acid formation inhibition and superoxide anion (O2•−) generation suppression. The inhibition of uric acid formation by XO is useful for the screening of natural compounds that prevent gout, while the suppression of O2•− generation is useful for treating oxidative stress. The plants contain abundant phenolic compounds, and some of these compounds, such as flavonoid, have shown to inhibit XO and reactive oxygen species [8]. In recent studies, it has been demonstrated that XO inhibitory activity and the suppression of O2•− generation are used to characterize natural products [9].
Limonium brasiliense (Boiss.) Kuntze (Plumbaginaceae) is a medicinal plant known by its common name, “guaycurú”, that grows mostly in saline soils distributed in Argentina, Uruguay, and South of Brazil. Infusion from the roots is popularly used in the treatment of hemorrhage, menstrual disorders, and rheumatism, and it is believed to have cardioprotective properties [10].
In previous studies, we evaluated the methanolic extract from the roots of L. brasiliense that showed antioxidant activity evidenced by the reduction of the stable free radical 2,2-diphenyl-1-picrylhydrazyl (DPPH) [11]. For this reason, we examined XO inhibitory activities of the aqueous fraction of L. brasiliense extract and its polar bioactive constituents using the enzymatic HX–XO system in vitro. Our results suggest that the antioxidant activity of the aqueous fraction of L. brasiliense extract may be partially explained by an O2•− scavenging effect and by inhibition of XO activity.
For the first time, this study provides a new potential alternative for the use of L. brasiliense against hyperuricemia and for control and gout prevention.
2. Material and Methods
2.1. General
1H and 13C NMR spectra were recorded in DMSO-d6 using tetramethylsilane (TMS) asinternal standard on a Bruker ARX 300 multinuclear spectrometer at 300 MHz and 75 MHz, respectively. UV spectra were recorded on a GBC spectral UV-VIS spectrophoto-meter.
Silica gel 60 (70–230 mesh, Fluka) was used for column chromatography, Dry Column Vacuum Chromatography (DCVC) with C18 silica, and semipreparative HPLC with a C18 Phenomenex Gemini column.
UV spectra were recorded on a GBC Spectral UV-VIS spectrophotometer and microplate UV/VIS reader (Titertek Multiscan, Flow Laboratories, Helsinki, Finland). Fluorescence emission was then determined in a microplate fluorescence reader (Bio-Tek Instruments, Jenway, United Kingdom; excitation 488 nm, detection 530 nm) over a 55 min period and microplate ELISA reader (Fluo-star Optima, BMG LABTECH, Offenburg, Germany).
Nitroblue tetrazolium (NBT), hypoxanthine (HX), xanthine, superoxide dismutase (SOD; from bovine erythrocytes), phosphate buffer (KH2PO4-KOH), Ethylenediaminetetraacetic acid disodium salt dihydrate (EDTA-Na2), allopurinol, and XO enzyme (from buttermilk) were all purchased from Sigma. Appropriate dilutions of the above drugs were prepared every day immediately before use in phosphate-buffered saline–PBS (XO assays) and/or in deionized water (for the other experiments) from the following concentrated stock solutions kept at −20 °C: SOD (1 U/mL) in deionized water, HX and xanthine (10 mM) in a 0.1% w/v aqueous KOH solution, all other compounds (100 mM) in deionized water. XO was dissolved daily before the experiments in PBS.
2.2. Plant Material
Roots of L. brasiliense were collected at Puerto Cuatreros (38°45′ S, 62°22′ O), General Cerri, Buenos Airesprovince, Argentina, in February 2010. A voucher specimen was identified by Dr. María Gabriela Murray and was deposited in the Herbarium of Universidad Nacional del Sur (BBB) in Bahía Blanca, Argentina, under the number Murray, M.G. 421.
2.3. Obtention of Aqueous Extract and Its Active Compounds
Dried roots (124.3 g) were macerated overnight with ethanol (EtOH) (1 L). The ethanolic extract (EE) was evaporated in vacuo to yield 78.2 g of a brown residue that was dissolved in water (200 mL) and partitioned with hexane (HF, 3 × 150 mL). The aqueous fraction (AF) was evaporated, and the residue was lyophilized for 3 days (45.04 g). A portion of AF (0.7 g) was submitted to DCVC chromatography with C18 silica and H2O:MeOH as eluent (100:0, 90:10, 80:20, 70:30, 50:50, 30:70, 0:100). This procedure was repeated 5 times. Chromatographic separation was monitored by TLC, and the fractions were pooled according to their chromatographic profile in seven fractions, F1–F7. Fractions F2, F3, and F4 were selected for further chromatographic separation due to their activity in the DPPH assay. These fractions were submitted to semipreparative HPLC with a C18 Phenomenex Gemini column (5 µm C18 110 Å, 250 × 10 mm) and MeOH:H2O (50:50) as mobile phase (1 mL/min). Absorbance was registered at λ = 254 nm. Compound 7 (8.2 mg) was isolated from the F2/A fraction and compound 1 (31.9 mg) from the F2/C fraction, with retention times of 3.1 and 11.2 min, respectively. Compound 6 (20.3 mg) was isolated from the F3/A fraction and compound 2 (29.4 mg) from the F/B fraction, with retention times of 8.7 and 10.8 min, respectively.
Compound 3 (61.4 mg) was obtained from F3/C and F4/C fractions, with a retention time of 25.2 min. Compounds 4 (30.5 mg) and 5 (6.1 mg) were obtained from F4/A and F4/B fractions, with retention times of 16.5 and 18.3 min, respectively. All compounds were characterized by 1H and 13C NMR.
2.4. Xanthine Oxidase Activity
Generation of O2•− by use of the enzymatic hypoxanthine oxidase the HX-XO system.
The superoxide ion (O2•−) was generated enzymatically in an HX-XO system and quantified by the spectrophotometric measurement of the product of the reduction of NBT, essentially following the procedure described by Orallo et al. [12] using 250 μL of test solution of 1 mM EDTA-Na2, 100 μM HX, 100 μM NBT in phosphate buffer (50 mM KH2PO4-KOH, pH 7.4) and EE or AF extracts (0.1–250 μg/mL) and compounds 1–7 (0.1–500 μM). Control experiments were carried out simultaneously using the same test solution but without extract. The reaction was started with the test solution already in a Cobas Fara 22–3123 AutoAnalyzer (Roche) by adding XO in phosphate buffer (0.066 U/mL) and continued at room temperature (22–24 °C) for 10 min. The rate of NBT reduction was estimated from the difference in absorbance at 560 nm between the test solution and a blank solution in which the XO was replaced by buffer solution. In some experiments, the sensitivity of the method was evaluated by assessing the influence of SOD (1 U/mL), a known scavenger of O2•−.
Determination of XO activity by use of the XO system.
The potential effect of L. brasiliense extracts and compounds 1–7 on XO activity was investigated by measuring uric acid formation, as previously described (Orallo et al. 2002). Briefly, 1 mL of test solutions of phosphate buffer (50 mM KH2PO4-KOH, pH 7.4) containing the EE or the AF extract at different concentrations (0.1–250 μg/mL) and compounds 1–7 at different concentrations (0.1–500 μM), EDTA-Na2 (1 mM), and 0.066 U of XO were incubated for 15 min at room temperature. The reaction was started by adding xanthine in phosphate buffer (100 μM), and the rate of uric acid production was estimated from the difference in absorbance at 295 nm (measured at room temperature for 10 min in a UV-visible absorption spectrophotometer, Shimadzu UV-240, Duisburg, Germany) between the test solution and a blank solution in which XO was replaced by buffer solution. In some assays, the validity of the method was confirmed by assessing the influence of allopurinol (a well-known XO inhibitor) on uric acid formation.
2.5. Antioxidant Activity
Determination of DPPH assay.
The antioxidant activity was evaluated through the ability as free radical scavenger of extracts, fractions, and/or pure compounds, as described Bors et al. [13]. The preliminary test was performed with a rapid TLC screening method using the 2,2-diphenyl-1-picrylhydrazyl radical. Analytical TLC on reverse plates was developed with appropriate conditions after application of 5 μL of each test compound solution (1 mg/mL), dried, and sprayed with DPPH solution (0.2%, MeOH). A total of 5 min later, active compounds appeared as yellow spots against a purple background. The purple stable free radical 2,2-diphenyl-1-picrylhydrazyl was reduced to the yellow colored diphenylpicryl hydrazine. Quercetin was used as positive control.
2.6. Statistical Analysis
Statistical analysis was performed using Tukey–Kramer correction of a one-way ANOVA test using Graph-Pad Prism 5.0 software. The results are expressed as means standard errors of the means (SEM). The concentration of drugs that caused a 50% inhibition of the response (IC50) was estimated by nonlinear regression analysis (GraphPad Prism software; GraphPad, San Diego, CA, USA) from the concentration–response curves obtained in each case.
3. Results and Discussion
3.1. Extraction and Isolation
Dried roots of L. brasiliense were milled and macerated with ethanol to obtain ethanolic extract (EE) that was dissolved in water (AF) and partitioned with hexane (HF). AF was submitted to chromatographic separations to render seven pure compounds that were identified as prodelphinidin B1-3,3′-digallate (1), myricetin (2), apigenin (3), taxifolin (4), 3-O-acetyltaxifolin (5), myricetin-3-O-α-rhamnopyranoside (6), and gallic acid (7). Identification was achieved on the basis of their 1H and 13C NMR spectra and by comparison with literature data [14,15,16]
3.2. Xhanthine Oxidase Activity
XO catalyzes the formation of uric acid and ROS during the catabolism of purines, and excess of uric acid can lead to increased oxidative stress, mutagenesis, and possibly to cancer development [17]. In recent studies, it has been demonstrated that XO inhibitory activity and the suppression of O2•− generation are used to characterize natural products. It has been reported that XO converts HX or xanthine to uric acid, H2O2, and O2•− [18]. O2•− generated by this system reacts with NBT to produce formazan (NBT is reduced in the reaction). The formation of this colored compound (formazan) and the amount of O2•− generated enzymatically may be measured spectrophotometrically [19]. When a drug lowers the amount of O2•− (i.e., the reduction of NBT) and at the same time does not affect the formation of uric acid, it is considered to be a selective scavenger of O2•−. On the other hand, if a drug inhibits XO activity, both uric acid and O2•− concentrations are diminished.
XO is a key enzyme which can catalyze hypoxanthine and xanthine to uric acid causing hyperuricemia in humans. In this study, we examined XO inhibitory activities of the aqueous fraction of L. brasiliense extract and its polar bioactive constituents using the enzymatic HX–XO system in vitro, which can be measured spectrophotometrically.
Superoxide-scavenging activity of the aqueous fraction and isolated compounds of L. Brasiliense.
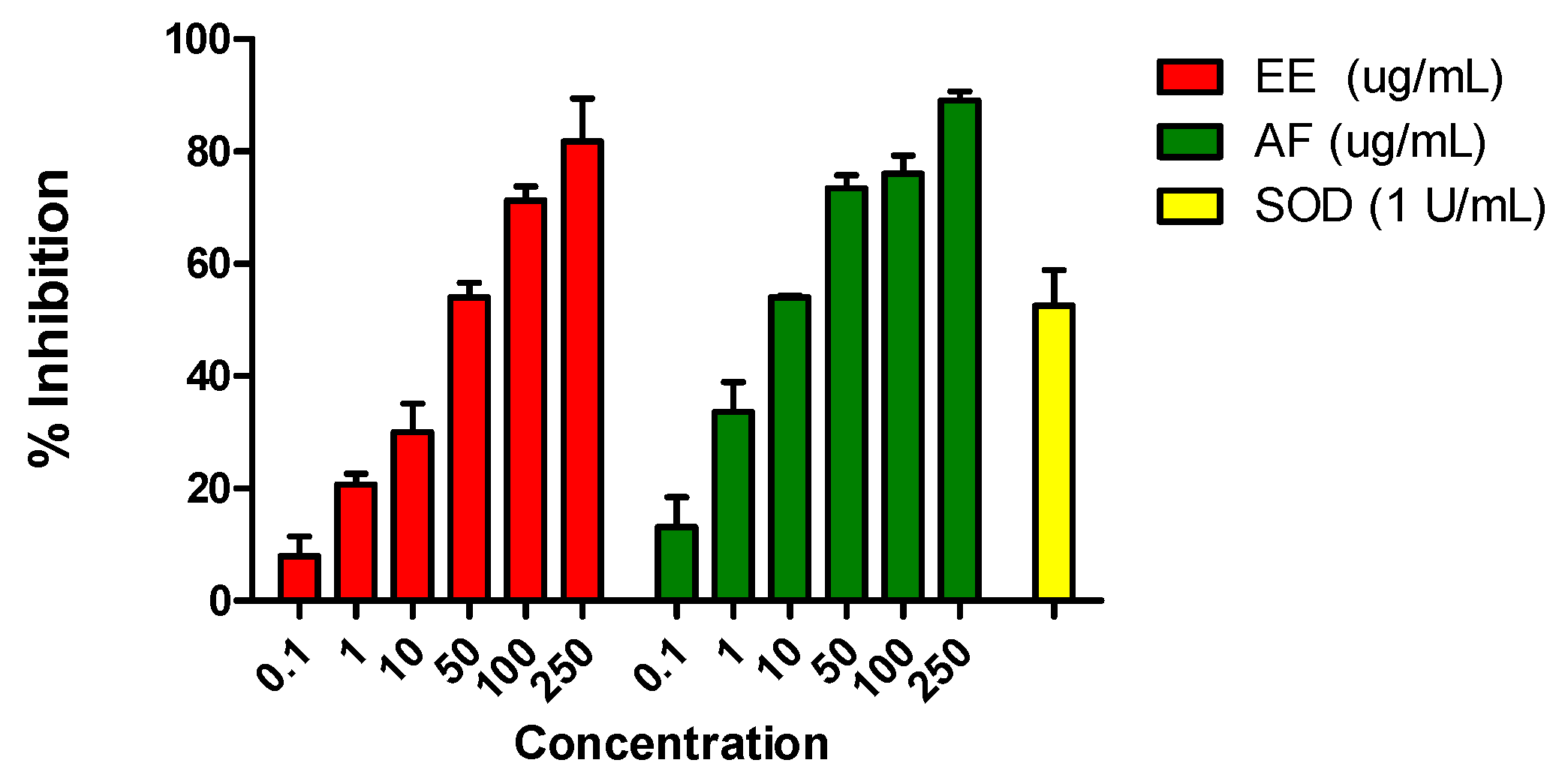
To investigate the possible superoxide scavenging activity, we performed assays in which O2•− was generated by an enzymatic system (HX-XO). EE and AF at concentrations between 0.1 and 250 μg/mL significantly reduced medium concentrations of O2•−, with the inhibitory effect with IC50 values of 42.03 ± 1.04 µg/mL and 10.86 ± 1.25 μg/mL, respectively (Figure 1).
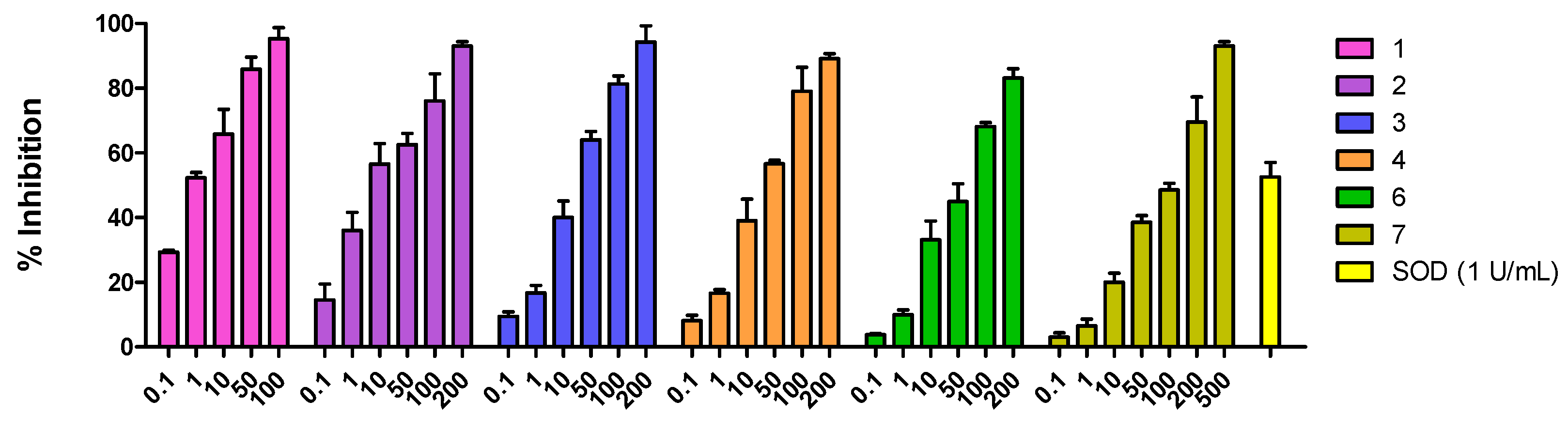
In this assay, myricetin (2) and myricetin-3-O-α-rhamnopyranoside (6) showed a significant superoxide scavenging activitywith IC50 values of 6.04 ± 1.51 µM and 18.96 ± 1.36 µM, respectively. A moderate effect was observed for taxifolin (4) with an IC50 = 35.93 ± 1.65 µM. Neither apigenin (3) (IC50 67.21 ± 1.35 µM) nor gallic acid (7) (IC50 126.13 ± 2.06 µM) had significant effects.
Prodelphinidin B1-3,3′-digallate (1) showed the most potent superoxide scavenging activity with an IC50 value of 1.58 ± 0.34 µM (Figure 2).
Effects of the aqueous fraction and compounds 1–7 on XO activity.
The effects of aqueous fraction of L. brasiliense extract and its polar bioactive constituents (1–7) on XO activity were investigated by measuring their ability to affect the formation of uric acid from xanthine.
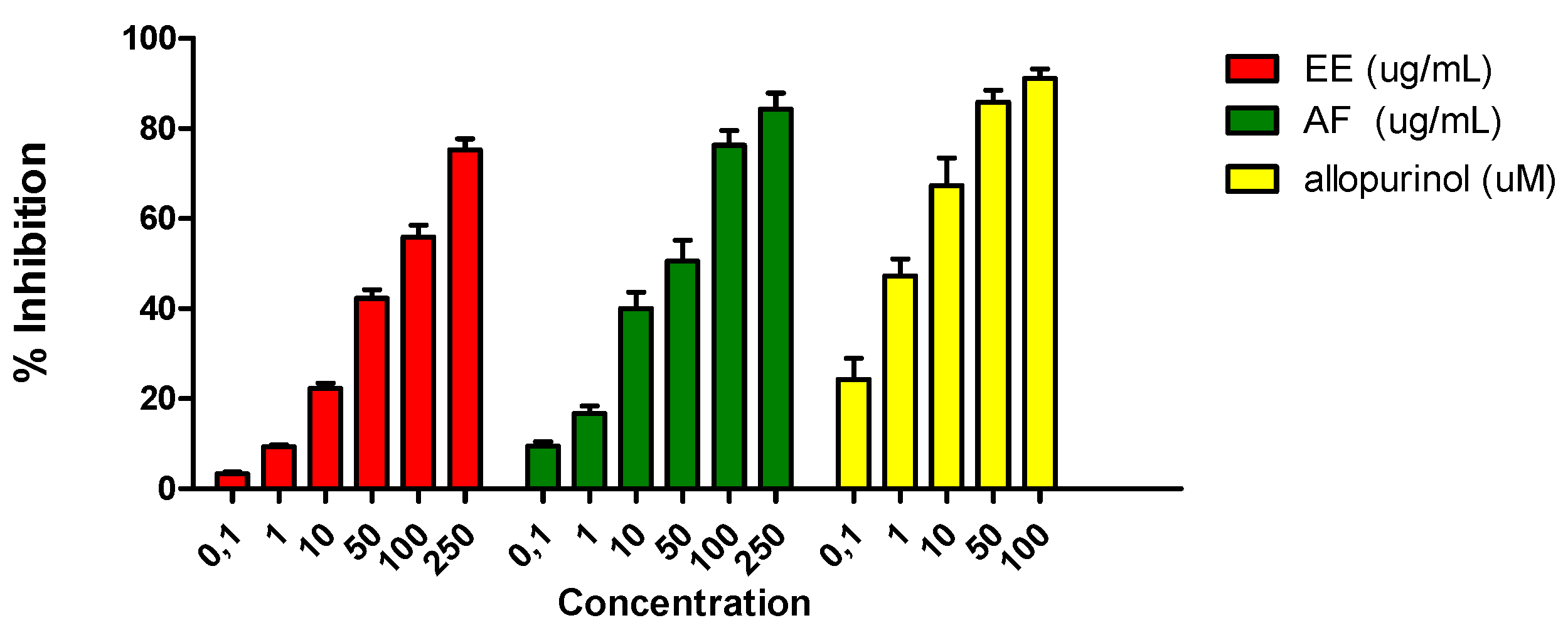
Only AF showed inhibition of uric acid formation and thus XO activity with an IC50 value of 48.30 ± 1.63 μg/mL, while EE showed a moderate inhibition of uric acid formation (IC50 = 96.14 ± 2.09 μg/mL) (Figure 3).
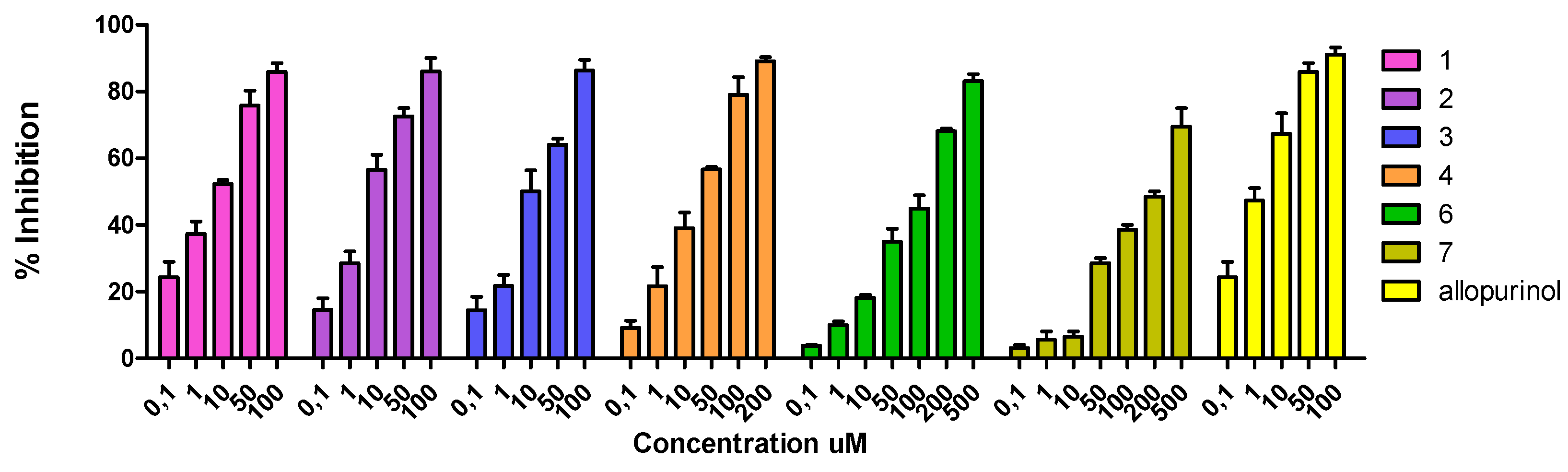
As is shown in Figure 4, neither myricetin-3-O-α-rhamnopyranoside (6) (IC50 167.02 ± 1.02 µM) nor gallic acid (7) (IC50 213.24 ± 1.61 µM) had significant effects, while taxifolin (4) elicited a moderate inhibition of uric acid formation (IC50 = 31.58 ± 0.36 μg/mL). On the other hand, myricetin (2) and apigenin (3) showed a significative inhibition of XO activity with IC50 values of 16.89 ± 1.03 µM and 19.01 ± 1.10 µM, respectively.
Prodelphinidin B1-3,3′-digallate (1) showed the most potent inhibitory activity with an IC50 value of 6.61 ± 0.13 µM, comparable to allopurinol, the drug of choice for inhibition of XO in gout patients.
4. Conclusions
In the present study, the effects of the aqueous fraction of L. brasiliense extract, and its polar bioactive constituents on XO activity were investigated by measuring their ability to affect the formation of uric acid from xanthine using the enzymatic HX–XO system in vitro and oxidative stress inhibition using O2•− scavenging effect.
The aqueous fraction of L. brasiliense displayed potent XO inhibitory activity with an IC50 value of 48.3 ± 1.63 µg/mL. Our result suggests that the antioxidant activity of the aqueous fraction of L. brasiliense extract previously reported [11] may be partially explained by an O2•− scavenging effect (IC50 10.86 ± 1.25 μg/mL) and by inhibition of xanthine oxidase activity.
This inhibitory effect is compatible with the presence of flavonoid compounds in this aqueous fraction. It has been previously demonstrated that myricetin (2) and apigenin (3) can inhibit XO activity [20].
Prodelphinidin B1-3,3′-digallate (1) was effective for scavenging superoxide radicals (O2•−) (IC50 1.58 ± 0.13 µM), generated enzymatically by a hypoxanthine/xanthine oxidase system and for inhibiting XO activity. This compound showed the most potent XO inhibitory activity with an IC50 value of 6.61 ± 0.13 µM, at the same level as allopurinol, the drug of choice for inhibition of XO in gout patients. These results clearly indicate that 1 displays selective O2•− scavenging and direct inhibitory XO properties.
In conclusion, the aqueous fraction of L. brasiliense extract significantly inhibits ROS production, both by inhibiting XO and by scavenging ROS already produced. It seems likely that these activities are attributable to prodelphinidin B1-3,3′-digallate (1) present in the extract.
For the first time, this study provides a new alternative for the use of aqueous fraction of L. brasiliense against hyperuricemia and for control and gout prevention. Thus, flavonoids could be used as a preventive and therapeutic agent for gout. In addition, they might be useful as a biochemical probe, taking advantage of their potent inhibitory activities.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
References
- Keenan, R.T.; O’Brien, W.R.; Lee, K.H.; Crittenden, D.B.; Fisher, M.C.; Goldfarb, D.S.; Krasnokutsky, S.; Oh, C.; Pillinger, M.H. Prevalence of Contraindications and Prescription of Pharmacologic Therapies for Gout. Am. J. Med. 2011, 124, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Darley-Usmar, V.; Wiseman, H.; Halliwell, B. Nitric oxide and oxygen radicals: A question of balance. FEBS Lett. 1995, 309, 131–135. [Google Scholar] [CrossRef]
- Klimiuk, P.A.; Sierakowski, S.; Latosiewicz, R.; Cylwik, B.; Skowronski, J.; Chwiecko, J. Serum cytokines in different histological variants of rheumatoid arthritis. J. Rheumatol. 2001, 28, 1211–1217. [Google Scholar] [PubMed]
- Bell, D.; Jackson, M.; Nicoll, J.J. Inflammatory response, neutrophil activation, and free radical production after acute myocardial infarction: Effect of thrombolytic treatment. Br. Heart J. 1990, 63, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Jackson, R.M. Reactive species mechanisms of cellular hypoxia-reoxygenation injury. Am. J. Physiol. Cell Physiol. 2002, 282, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, N.; Miura, S.; Zeki, S.; Ishii, H. Hepatocellular oxidative DNA injury induced by macrophage-derived nitric oxide. Free Radical. Biol. Med. 2001, 30, 1019–1028. [Google Scholar] [CrossRef]
- Satoh, T.; Numakawa, T.; Abiru, Y. Production of reactive oxygen species and release of L-glutamate during superoxide anion-induced cell death of cerebellar granule neurons. J. Neurochem. 1998, 70, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Nagao, A.; Seki, M.; Kobayashi, H. Inhibition of Xanthine Oxidase by Flavonoids. Biosci. Biotechnol. Biochem. 1999, 63, 1787–1790. [Google Scholar] [CrossRef] [PubMed]
- Nessa, F.; Saeed, A.K. Evaluation of antioxidant and xanthine oxidase inhibitory activity of different solvent extracts of leaves of Citrullus colocynthis. Pharmacogn. Res. 2014, 3, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.P. 270 Plantas Medicinales Iberoamericanas; CYTED-SECAB: Bogotá, Colombia, 1995; pp. 21–24. [Google Scholar]
- Murray, A.P.; Rodriguez, S.; Frontera, M.A.; Tomas, M.A.; Mulet, M.C. Antioxidant metabolites from Limonium brasiliense (Boiss.) kuntze. Z. Nat. Sect C J. Biosci. 2004, 59, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Orallo, F.; Alvarez, E.; Camina, M.; Leiro, J.M.; Gomez, E.; Fernandez, P. The possible implication of trans-resveratrol in the cardioprotective effects of long-term moderate wine consumption. Mol. Pharmacol. 2002, 61, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Bors, W.; Saran, M.; Eltsner, E.F. Screening for plants antioxidants. In Modern Methods of Plant Analysis New Series; Plant Toxin Analysis; Linskens, H.F., Jackson, J.F., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; Volume 13, pp. 277–295. [Google Scholar]
- Agrawal, P.K. Carbon-13 NMR of Flavonoids; Elsevier: Amsterdam, The Netherlands, 1989. [Google Scholar]
- Markham, K.R.; Ternai, B.; Stanley, R.; Geiger, H.; Mabry, T.J. Carbon-13 NMR studies of flavonoids—III: Naturally occurring flavonoid glycosides and their acylated derivatives. Tetrahedron 1978, 34, 1389–1397. [Google Scholar] [CrossRef]
- Shen, C.C.; Chang, Y.S.; Hott, L.K. Nuclear magnetic resonance studies of 5,7-dihydroxyflavonoids. Phytochemistry 1993, 34, 843–845. [Google Scholar] [CrossRef]
- Aucamp, J.; Gaspar, A.; Hara, Y.; Apostolides, Z. Inhibition of xanthine oxidase by catechins from tea (Camellia sinensis). Anticancer Res. 1997, 17, 4381–4385. [Google Scholar] [PubMed]
- Wippich, N.I.C.O.; Peschke, D.O.R.O.T.H.E.E.; Peschke, E.L.M.A.R.; Holtz, J.Ü.R.G.E.N.; Bromme, H.J. Comparison between xanthine oxidases from buttermilk and microorganisms regarding their ability to generate reactive oxygen species. Int. J. Mol. Med. 2001, 7, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Huang, L.; Sun, C.; Zhao, D.; Tang, H. Studies on the structure-activity relationship and interaction mechanism of flavonoids and xanthine oxidase through enzyme kinetics, spectroscopy methods and molecular simulations. Food Chem. 2020, 323, 126807. [Google Scholar] [CrossRef] [PubMed]
- Mohos, V.; Fliszár-Nyúl, E.; Poór, M. Inhibition of Xanthine Oxidase-Catalyzed Xanthine and 6-Mercaptopurine Oxidation by Flavonoid Aglycones and Some of Their Conjugates. Int. J. Mol. Sci. 2020, 21, 3256–3266. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of the ethanolic extract (EE) and the aqueous fraction (AF) (0.1–250 μg/mL) of L. brasiliense on reduction of nitroblue tetrazolium (NBT) by O2•− generated by xanthine oxidase (XO) in the presence of 100 μM hypoxanthine (HX). The rightmost bar represents assays in which the enzyme superoxide dismutase (SOD; 1 U/mL) was added to cultures instead of EE and AF; this enzyme abolishes superoxide generation in the HX-XO system. Values shown are mean percentages of the control response (n = 3) ± SEM. Asterisks indicate statistical significance (p < 0.001).
Figure 1.
Effects of the ethanolic extract (EE) and the aqueous fraction (AF) (0.1–250 μg/mL) of L. brasiliense on reduction of nitroblue tetrazolium (NBT) by O2•− generated by xanthine oxidase (XO) in the presence of 100 μM hypoxanthine (HX). The rightmost bar represents assays in which the enzyme superoxide dismutase (SOD; 1 U/mL) was added to cultures instead of EE and AF; this enzyme abolishes superoxide generation in the HX-XO system. Values shown are mean percentages of the control response (n = 3) ± SEM. Asterisks indicate statistical significance (p < 0.001).

Figure 2.
Effects of the compounds 1–7 (0.1–500 μM) on reduction of NBT by O2•− generated by XO in the presence of 100 μM HX. The rightmost bar represents assays in which the enzyme superoxide dismutase (SOD; 1 U/mL) was added to cultures instead of EE and AF; this enzyme abolishes superoxide generation in the HX-XO system. Values shown are mean percentages of the control response (n = 3) ± SEM. Asterisks indicate statistical significance (p < 0.001).
Figure 2.
Effects of the compounds 1–7 (0.1–500 μM) on reduction of NBT by O2•− generated by XO in the presence of 100 μM HX. The rightmost bar represents assays in which the enzyme superoxide dismutase (SOD; 1 U/mL) was added to cultures instead of EE and AF; this enzyme abolishes superoxide generation in the HX-XO system. Values shown are mean percentages of the control response (n = 3) ± SEM. Asterisks indicate statistical significance (p < 0.001).

Figure 3.
Effects of EE and AF (0.1–250 μg/mL) of L. brasiliense on uric acid production by XO in the presence of 100 μM xanthine. The rightmost bar represents assays in which the XO inhibitor allopurinol was added. Values shown are means percentages of the control response (n = 3) ± SEM. Asterisks indicate statistical significance (p < 0.001).
Figure 3.
Effects of EE and AF (0.1–250 μg/mL) of L. brasiliense on uric acid production by XO in the presence of 100 μM xanthine. The rightmost bar represents assays in which the XO inhibitor allopurinol was added. Values shown are means percentages of the control response (n = 3) ± SEM. Asterisks indicate statistical significance (p < 0.001).

Figure 4.
Effects of the compounds 1–7 (0.1–500 μM) of L. brasiliense on uric acid production by XO in the presence of 100 μM xanthine. The rightmost bar represents assays in which the XO inhibitor allopurinol was added. Values shown are means percentages of the control response (n = 3) ±SEM. Asterisks indicate statistical significance (p < 0.001).
Figure 4.
Effects of the compounds 1–7 (0.1–500 μM) of L. brasiliense on uric acid production by XO in the presence of 100 μM xanthine. The rightmost bar represents assays in which the XO inhibitor allopurinol was added. Values shown are means percentages of the control response (n = 3) ±SEM. Asterisks indicate statistical significance (p < 0.001).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
IC50 values SOD and XO of extracts and isolated compounds obtained from L. brasiliense.
| Antioxidants | IC50 SOD a | IC50 XO a |
|---|---|---|
| allopurinol b | N.T | 3.61 ± 0.05 c |
| EE | 42.03 ± 1.04 d | 96.14 ± 2.09 d |
| AF | 10.86 ± 1.25 d | 48.3 ± 1.63 d |
| prodelphinidin B1-3,3′-digallate (1) | 1.58 ± 0.34 c | 6.61 ± 0.13 c |
| myricetin (2) | 6.04 ± 1.51 c | 16.89 ± 1.03 c |
| apigenin (3) | 67.21 ± 1.35 c | 19.01 ± 1.10 c |
| taxifolin (4) | 35.93 ± 1.65 c | 31.58 ± 0.36 c |
| myricetin-3-O-α-rhamnopyranoside (6) | 18.96 ± 1.36 c | 167.02 ± 1.02 c |
| gallic acid (7) | 126.13 ± 2.06 c | 213.24 ± 1.61 c |
a Values are the mean ± S.E.M. of three replicas. b Positive control c µM. d µg/mL. NT not tested.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rodriguez, S.A.; Murray, A.P.; Leiro, J.M. Xanthine Oxidase Inhibition by Aqueous Extract of Limonium brasiliense (Plumbaginaceae). Chem. Proc. 2021, 3, 123. https://0-doi-org.brum.beds.ac.uk/10.3390/ecsoc-24-08410
AMA Style
Rodriguez SA, Murray AP, Leiro JM. Xanthine Oxidase Inhibition by Aqueous Extract of Limonium brasiliense (Plumbaginaceae). Chemistry Proceedings. 2021; 3(1):123. https://0-doi-org.brum.beds.ac.uk/10.3390/ecsoc-24-08410
Chicago/Turabian StyleRodriguez, Silvana Andrea, Ana Paula Murray, and José Manuel Leiro. 2021. "Xanthine Oxidase Inhibition by Aqueous Extract of Limonium brasiliense (Plumbaginaceae)" Chemistry Proceedings 3, no. 1: 123. https://0-doi-org.brum.beds.ac.uk/10.3390/ecsoc-24-08410