
Natural Killer Cell Mobilization in Breast and Prostate Cancer Survivors: The Implications of Altered Stress Hormones Following Acute Exercise



{kind=link}
{kind=link}
Abstract
:1. Overview of Natural Killer Cells
2. NK Cells and Hormone-Dependent Cancers
3. Side Effects of Breast and Prostate Cancer Treatments
3.1. General Side Effects
3.2. Endocrine Therapy Side Effects
3.3. Endocrine Therapy Helps Mediate Chemotherapy and Radiation Side Effects on Immunity
3.4. Androgen Deprivation Therapy Side Effects on Immunity
4. Exercise during Cancer Treatment
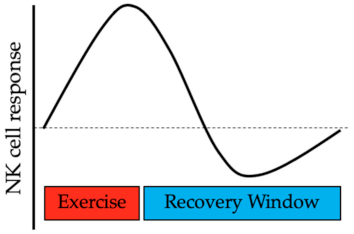
5. Mobilization of Natural Killer Cells during Hormone-Dependent Cancer
5.1. Attenuated Catecholamine Response
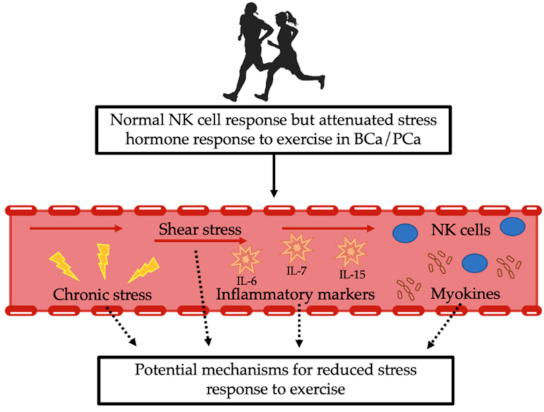
5.2. Other Potential Mechanisms
5.2.1. Shear Stress
5.2.2. Myokines
5.2.3. Chronic Stress Exposure
5.3. Interaction between Acute and Chronic Exercise
6. Implications and Future Directions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Dranoff, G. Cytokines in cancer pathogenesis and cancer therapy. Nat. Rev. Cancer 2004, 4, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Lowder, T.W.; Spielmann, G.; Bigley, A.B.; LaVoy, E.C.; Kunz, H. Exercise and the aging immune system. Ageing Res. Rev. 2012, 11, 404–420. [Google Scholar] [CrossRef] [PubMed]
- Leading Causes of Death (U.S.). Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/nchs/fastats/leading-causes-of-death.htm (accessed on 15 March 2021).
- De Maria, A.; Bozzano, F.; Cantoni, C.; Moretto, L. Revisiting human natural killer cell subset function revealed cytolytic CD56(dim)CD16+ NK cells as rapid producers of abundant IFN-gamma on activation. Proc. Natl. Acad. Sci. USA 2011, 108, 728–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bigley, A.B.; Rezvani, K.; Chew, C.; Sekine, T.; Pistillo, M.; Crucian, B.; Bollard, C.M.; Simpson, R.J. Acute exercise preferentially redeploys NK-cells with a highly-differentiated phenotype and augments cytotoxicity against lymphoma and multiple myeloma target cells. Brain Behav. Immun. 2014, 39, 160–171. [Google Scholar] [CrossRef]
- Gayoso, I.; Sanchez-Correa, B.; Campos, C.; Alonso, C.; Pera, A.; Casado, J.G.; Morgado, S.; Tarazona, R.; Solana, R. Immunosenescence of human natural killer cells. J. Innate. Immun. 2011, 3, 337–343. [Google Scholar] [CrossRef]
- Campbell, J.P.; Turner, J.E. Debunking the Myth of Exercise-Induced Immune Suppression: Redefining the Impact of Exercise on Immunological Health Across the Lifespan. Front. Immunol. 2018, 9, 648. [Google Scholar] [CrossRef]
- Peake, J.M.; Neubauer, O.; Walsh, N.P.; Simpson, R.J. Recovery of the immune system after exercise. J. Appl. Physiol. (1985) 2017, 122, 1077–1087. [Google Scholar] [CrossRef]
- Walsh, N.P.; Gleeson, M.; Shephard, R.J.; Gleeson, M.; Woods, J.A.; Bishop, N.C.; Fleshner, M.; Green, C.; Pedersen, B.K.; Hoffman-Goetz, L.; et al. Position statement. Part one: Immune function and exercise. Exerc. Immunol. Rev. 2011, 17, 6–63. [Google Scholar]
- Evans, E.S.; Hackney, A.C.; McMurray, R.G.; Randell, S.H.; Muss, H.B.; Deal, A.M.; Battaglini, C.L. Impact of Acute Intermittent Exercise on Natural Killer Cells in Breast Cancer Survivors. Integr. Cancer Ther. 2015, 14, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Hanson, E.D.; Sakkal, S.; Que, S.; Cho, E.; Spielmann, G.; Kadife, E.; Violet, J.A.; Battaglini, C.L.; Stoner, L.; Bartlett, D.B.; et al. Natural killer cell mobilization and egress following acute exercise in men with prostate cancer. Exp. Physiol. 2020, 105, 1524–1539. [Google Scholar] [CrossRef]
- Rooney, B.V.; Bigley, A.B.; LaVoy, E.C.; Laughlin, M.; Pedlar, C.; Simpson, R.J. Lymphocytes and monocytes egress peripheral blood within minutes after cessation of steady state exercise: A detailed temporal analysis of leukocyte extravasation. Physiol. Behav. 2018, 194, 260–267. [Google Scholar] [CrossRef] [Green Version]
- Nieman, D.C.; Miller, A.R.; Henson, D.A.; Warren, B.J.; Gusewitch, G.; Johnson, R.L.; Davis, J.M.; Butterworth, D.E.; Herring, J.L.; Nehlsen-Cannarella, S.L. Effect of high- versus moderate-intensity exercise on lymphocyte subpopulations and proliferative response. Int. J. Sports Med. 1994, 15, 199–206. [Google Scholar] [CrossRef]
- Anane, L.H.; Edwards, K.M.; Burns, V.E.; Drayson, M.T.; Riddell, N.E.; van Zanten, J.J.; Wallace, G.R.; Mills, P.J.; Bosch, J.A. Mobilization of gammadelta T lymphocytes in response to psychological stress, exercise, and beta-agonist infusion. Brain Behav. Immun. 2009, 23, 823–829. [Google Scholar] [CrossRef]
- Natale, V.M.; Brenner, I.K.; Moldoveanu, A.I.; Vasiliou, P.; Shek, P.; Shephard, R.J. Effects of three different types of exercise on blood leukocyte count during and following exercise. Sao Paulo Med. J. 2003, 121, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Hanson, E.D.; Bates, L.C.; Bartlett, D.B.; Campbell, J.P. Does exercise attenuate age- and disease-associated dysfunction in unconventional T cells? Shining a light on overlooked cells in exercise immunology. Eur. J. Appl. Physiol. 2021. [Google Scholar] [CrossRef]
- Standish, L.J.; Sweet, E.S.; Novack, J.; Wenner, C.A.; Bridge, C.; Nelson, A.; Martzen, M.; Torkelson, C. Breast cancer and the immune system. J. Soc. Integr. Oncol. 2008, 6, 158–168. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Kruijsen-Jaarsma, M.; Revesz, D.; Bierings, M.B.; Buffart, L.M.; Takken, T. Effects of exercise on immune function in patients with cancer: A systematic review. Exerc. Immunol. Rev. 2013, 19, 120–143. [Google Scholar]
- Khosravi, N.; Stoner, L.; Farajivafa, V.; Hanson, E.D. Exercise training, circulating cytokine levels and immune function in cancer survivors: A meta-analysis. Brain Behav. Immun. 2019, 81, 92–104. [Google Scholar] [CrossRef]
- Campbell, K.L.; Winters-Stone, K.M.; Wiskemann, J.; May, A.M.; Schwartz, A.L.; Courneya, K.S.; Zucker, D.S.; Matthews, C.E.; Ligibel, J.A.; Gerber, L.H.; et al. Exercise Guidelines for Cancer Survivors: Consensus Statement from International Multidisciplinary Roundtable. Med. Sci. Sports Exerc. 2019, 51, 2375–2390. [Google Scholar] [CrossRef] [Green Version]
- von Blanckenburg, P.; Schuricht, F.; Albert, U.S.; Rief, W.; Nestoriuc, Y. Optimizing expectations to prevent side effects and enhance quality of life in breast cancer patients undergoing endocrine therapy: Study protocol of a randomized controlled trial. BMC Cancer 2013, 13, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramchand, S.K.; Lim, E.; Grossmann, M. Adjuvant endocrine therapy in women with oestrogen-receptor-positive breast cancer: How should the skeletal and vascular side effects be assessed and managed? Clin. Endocrinol. 2016, 85, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Choo, S.B.; Saifulbahri, A.; Zullkifli, S.N.; Fadzil, M.L.; Redzuan, A.M.; Abdullah, N.; Bustamam, R.S.A.; Ahmad, H.Z.; Shah, N.M. Adjuvant endocrine therapy side-effects among postmenopausal breast cancer patients in Malaysia. Climacteric 2019, 22, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Berkowitz, M.J.; Thompson, C.K.; Zibecchi, L.T.; Lee, M.K.; Streja, E.; Berkowitz, J.S.; Wenziger, C.M.; Baker, J.L.; DiNome, M.L.; Attai, D.J. How patients experience endocrine therapy for breast cancer: An online survey of side effects, adherence, and medical team support. J. Cancer Surviv. 2021, 15, 29–39. [Google Scholar] [CrossRef]
- Condorelli, R.; Vaz-Luis, I. Managing side effects in adjuvant endocrine therapy for breast cancer. Expert Rev. Anticancer. Ther. 2018, 18, 1101–1112. [Google Scholar] [CrossRef]
- Bargiota, A.; Oeconomou, A.; Zachos, I.; Samarinas, M.; Pisters, L.L.; Tzortzis, V. Adverse effects of androgen deprivation therapy in patients with prostate cancer: Focus on muscle and bone health. J. BUON 2020, 25, 1286–1294. [Google Scholar]
- Agarwal, M.; Canan, T.; Glover, G.; Thareja, N.; Akhondi, A.; Rosenberg, J. Cardiovascular Effects of Androgen Deprivation Therapy in Prostate Cancer. Curr. Oncol. Rep. 2019, 21, 91. [Google Scholar] [CrossRef]
- Fang, D.; Zhou, L. Androgen deprivation therapy in nonmetastatic prostate cancer patients: Indications, treatment effects, and new predictive biomarkers. Asia Pac. J. Clin. Oncol. 2019, 15, 108–120. [Google Scholar] [CrossRef] [Green Version]
- Edmunds, K.; Tuffaha, H.; Scuffham, P.; Galvao, D.A.; Newton, R.U. The role of exercise in the management of adverse effects of androgen deprivation therapy for prostate cancer: A rapid review. Support. Care Cancer 2020, 28, 5661–5671. [Google Scholar] [CrossRef]
- Mohamad, N.V.; Soelaiman, I.N.; Chin, K.Y. A Review on the Effects of Androgen Deprivation Therapy (ADT) on Bone Health Status in Men with Prostate Cancer. Endocr. Metab. Immune. Disord. Drug Targets 2017, 17, 276–284. [Google Scholar] [CrossRef]
- Solomayer, E.F.; Feuerer, M.; Bai, L.; Umansky, V.; Beckhove, P.; Meyberg, G.C.; Bastert, G.; Schirrmacher, V.; Diel, I.J. Influence of adjuvant hormone therapy and chemotherapy on the immune system analysed in the bone marrow of patients with breast cancer. Clin. Cancer Res. 2003, 9, 174–180. [Google Scholar]
- Mozaffari, F.; Lindemalm, C.; Choudhury, A.; Granstam-Bjorneklett, H.; Helander, I.; Lekander, M.; Mikaelsson, E.; Nilsson, B.; Ojutkangas, M.L.; Osterborg, A.; et al. NK-cell and T-cell functions in patients with breast cancer: Effects of surgery and adjuvant chemo- and radiotherapy. Br. J. Cancer 2007, 97, 105–111. [Google Scholar] [CrossRef] [Green Version]
- Kang, D.H.; Weaver, M.T.; Park, N.J.; Smith, B.; McArdle, T.; Carpenter, J. Significant impairment in immune recovery after cancer treatment. Nurs Res. 2009, 58, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Zavadova, E.; Vocka, M.; Spacek, J.; Konopasek, B.; Fucikova, T.; Petruzelka, L. Cellular and humoral immunodeficiency in breast cancer patients resistant to hormone therapy. Neoplasma 2014, 61, 90–98. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A.M.; Roxa, A.; Leandersson, K.; Bergenfelz, C. Impact of systemic therapy on circulating leukocyte populations in patients with metastatic breast cancer. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sabbioni, M.E.; Castiglione, M.; Hurny, C.; Siegrist, H.P.; Bacchi, M.; Bernhard, J.; Thurlimann, B.; Bonnefoi, H.; Perey, L.; Goldhirsch, A.; et al. Interaction of tamoxifen with concurrent cytotoxic adjuvant treatment affects lymphocytes and lymphocyte subsets counts in breast cancer patients. Support. Care Cancer 1999, 7, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Sheard, C.R.; Reilly, F.; Tee, D.E.; Vergani, D.; Lowe, D.; Baum, M.; Cameron, A.E. The effect of adjuvant cyclophosphamide or tamoxifen on the numbers of lymphocytes bearing T cell or NK cell markers. Br. J. Cancer 1986, 54, 705–709. [Google Scholar] [CrossRef] [Green Version]
- Gannon, P.O.; Poisson, A.O.; Delvoye, N.; Lapointe, R.; Mes-Masson, A.M.; Saad, F. Characterization of the intra-prostatic immune cell infiltration in androgen-deprived prostate cancer patients. J. Immunol. Methods 2009, 348, 9–17. [Google Scholar] [CrossRef]
- Aragon-Ching, J.B.; Williams, K.M.; Gulley, J.L. Impact of androgen-deprivation therapy on the immune system: Implications for combination therapy of prostate cancer. Front. Biosci 2007, 12, 4957–4971. [Google Scholar] [CrossRef] [Green Version]
- Morse, M.D.; McNeel, D.G. Prostate cancer patients on androgen deprivation therapy develop persistent changes in adaptive immune responses. Hum. Immunol. 2010, 71, 496–504. [Google Scholar] [CrossRef] [Green Version]
- Mercader, M.; Bodner, B.K.; Moser, M.T.; Kwon, P.S.; Park, E.S.; Manecke, R.G.; Ellis, T.M.; Wojcik, E.M.; Yang, D.; Flanigan, R.C.; et al. T cell infiltration of the prostate induced by androgen withdrawal in patients with prostate cancer. Proc. Natl. Acad. Sci. USA 2001, 98, 14565–14570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battaglini, C.L.; Mills, R.C.; Phillips, B.L.; Lee, J.T.; Story, C.E.; Nascimento, M.G.; Hackney, A.C. Twenty-five years of research on the effects of exercise training in breast cancer survivors: A systematic review of the literature. World J. Clin. Oncol 2014, 5, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Hanson, E.D.; Hurley, B.F. Intervening on the side effects of hormone-dependent cancer treatment: The role of strength training. J. Aging Res. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, S.C.; Newton, R.U.; Spence, R.R.; Galvao, D.A. The Exercise and Sports Science Australia position statement: Exercise medicine in cancer management. J. Sci. Med. Sport 2019, 22, 1175–1199. [Google Scholar] [CrossRef] [Green Version]
- Shephard, R.J. Adhesion molecules, catecholamines and leucocyte redistribution during and following exercise. Sports Med. 2003, 33, 261–284. [Google Scholar] [CrossRef]
- Sen, C.K.; Roy, S. Antioxidant regulation of cell adhesion. Med. Sci. Sports Exerc. 2001, 33, 377–381. [Google Scholar] [CrossRef]
- Dusseaux, M.; Martin, E.; Serriari, N.; Peguillet, I.; Premel, V.; Louis, D.; Milder, M.; Le Bourhis, L.; Soudais, C.; Treiner, E.; et al. Human MAIT cells are xenobiotic-resistant, tissue-targeted, CD161hi IL-17-secreting T cells. Blood 2011, 117, 1250–1259. [Google Scholar] [CrossRef]
- Kay, N.E.; Oken, M.M.; Kyle, R.; Van Ness, B.; Kalish, L.; Leong, T.; Greipp, P. Sequential phenotyping of myeloma patients on chemotherapy: Persistence of activated T-cells and natural killer cells. Leuk. Lymphoma 1995, 16, 351–354. [Google Scholar] [CrossRef]
- Waidhauser, J.; Schuh, A.; Trepel, M.; Schmalter, A.K.; Rank, A. Chemotherapy markedly reduces B cells but not T cells and NK cells in patients with cancer. Cancer Immunol. Immunother. 2020, 69, 147–157. [Google Scholar] [CrossRef]
- Zimmer, P.; Baumann, F.T.; Bloch, W.; Zopf, E.M.; Schulz, S.; Latsch, J.; Schollmayer, F.; Shimabukuro-Vornhagen, A.; von Bergwelt-Baildon, M.; Schenk, A. Impact of a half marathon on cellular immune system, pro-inflammatory cytokine levels, and recovery behavior of breast cancer patients in the aftercare compared to healthy controls. Eur. J. Haematol. 2016, 96, 152–159. [Google Scholar] [CrossRef]
- Shek, P.N.; Sabiston, B.H.; Buguet, A.; Radomski, M.W. Strenuous exercise and immunological changes: A multiple-time-point analysis of leukocyte subsets, CD4/CD8 ratio, immunoglobulin production and NK cell response. Int. J. Sports Med. 1995, 16, 466–474. [Google Scholar] [CrossRef]
- Zouhal, H.; Jacob, C.; Delamarche, P.; Gratas-Delamarche, A. Catecholamines and the effects of exercise, training and gender. Sports Med. 2008, 38, 401–423. [Google Scholar] [CrossRef]
- Graff, R.M.; Kunz, H.E.; Agha, N.H.; Baker, F.L.; Laughlin, M.; Bigley, A.B.; Markofski, M.M.; LaVoy, E.C.; Katsanis, E.; Bond, R.A.; et al. beta2-Adrenergic receptor signaling mediates the preferential mobilization of differentiated subsets of CD8+ T-cells, NK-cells and non-classical monocytes in response to acute exercise in humans. Brain Behav. Immun. 2018, 74, 143–153. [Google Scholar] [CrossRef]
- Evans, E.S.; Battaglini, C.L.; Groff, D.G.; Hackney, A.C. Aerobic exercise intensity in breast cancer patients: A preliminary investigation. Integr. Cancer Ther. 2009, 8, 139–147. [Google Scholar] [CrossRef]
- Tosti, K.P.; Hackney, A.C.; Battaglini, C.L.; Evans, E.S.; Groff, D. Exercise in patients with breast cancer and healthy controls: Energy substrate oxidation and blood lactate responses. Integr. Cancer Ther. 2011, 10, 6–15. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.S.; Hackney, A.C.; Pebole, M.M.; McMurray, R.G.; Muss, H.B.; Deal, A.M.; Battaglini, C.L. Adrenal Hormone and Metabolic Biomarker Responses to 30 min of Intermittent Cycling Exercise in Breast Cancer Survivors. Int. J. Sports Med. 2016, 37, 921–929. [Google Scholar] [CrossRef] [Green Version]
- Hanson, E.D.; Sakkal, S.; Evans, W.S.; Violet, J.A.; Battaglini, C.L.; McConell, G.K.; Hayes, A. Altered stress hormone response following acute exercise during prostate cancer treatment. Scand. J. Med. Sci. Sports 2018, 28, 1925–1933. [Google Scholar] [CrossRef]
- Carlson, L.E.; Angen, M.; Cullum, J.; Goodey, E.; Koopmans, J.; Lamont, L.; MacRae, J.H.; Martin, M.; Pelletier, G.; Robinson, J.; et al. High levels of untreated distress and fatigue in cancer patients. Br. J. Cancer 2004, 90, 2297–2304. [Google Scholar] [CrossRef]
- Pedersen, L.; Idorn, M.; Olofsson, G.H.; Lauenborg, B.; Nookaew, I.; Hansen, R.H.; Johannesen, H.H.; Becker, J.C.; Pedersen, K.S.; Dethlefsen, C.; et al. Voluntary running suppresses tumor growth through epinephrine-and il-6-dependent nk cell mobilization and redistribution. Cell Metab. 2016, 23, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Paszkowiak, J.J.; Dardik, A. Arterial wall shear stress: Observations from the bench to the bedside. Vasc. Endovasc. Surg. 2003, 37, 47–57. [Google Scholar] [CrossRef]
- Evans, W. NK cell recruitment and exercise: Potential immunotherapeutic role of shear stress and endothelial health. Med. Hypotheses 2017, 109, 170–173. [Google Scholar] [CrossRef]
- Provost, P.; Lam, J.Y.; Lacoste, L.; Merhi, Y.; Waters, D. Endothelium-derived nitric oxide attenuates neutrophil adhesion to endothelium under arterial flow conditions. Arter. Thromb. 1994, 14, 331–335. [Google Scholar] [CrossRef] [Green Version]
- Benschop, R.J.; Nijkamp, F.P.; Ballieux, R.E.; Heijnen, C.J. The effects of beta-adrenoceptor stimulation on adhesion of human natural killer cells to cultured endothelium. Br. J. Pharm. 1994, 113, 1311–1316. [Google Scholar] [CrossRef]
- Benschop, R.J.; Oostveen, F.G.; Heijnen, C.J.; Ballieux, R.E. Beta 2-adrenergic stimulation causes detachment of natural killer cells from cultured endothelium. Eur. J. Immunol. 1993, 23, 3242–3247. [Google Scholar] [CrossRef]
- Benschop, R.J.; Schedlowski, M.; Wienecke, H.; Jacobs, R.; Schmidt, R.E. Adrenergic control of natural killer cell circulation and adhesion. Brain Behav. Immun. 1997, 11, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Capellino, S.; Claus, M.; Watzl, C. Regulation of natural killer cell activity by glucocorticoids, serotonin, dopamine, and epinephrine. Cell Mol. Immunol. 2020, 17, 705–711. [Google Scholar] [CrossRef]
- Huang, Q.; Hu, X.; He, W.; Zhao, Y.; Hao, S.; Wu, Q.; Li, S.; Zhang, S.; Shi, M. Fluid shear stress and tumor metastasis. Am. J. Cancer Res. 2018, 8, 763–777. [Google Scholar]
- Regmi, S.; Fu, A.; Luo, K.Q. High Shear Stresses under Exercise Condition Destroy Circulating Tumor Cells in a Microfluidic System. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Avvisato, C.L.; Yang, X.; Shah, S.; Hoxter, B.; Li, W.; Gaynor, R.; Pestell, R.; Tozeren, A.; Byers, S.W. Mechanical force modulates global gene expression and beta-catenin signaling in colon cancer cells. J. Cell Sci. 2007, 120, 2672–2682. [Google Scholar] [CrossRef] [Green Version]
- Schadler, K.L.; Thomas, N.J.; Galie, P.A.; Bhang, D.H.; Roby, K.C.; Addai, P.; Till, J.E.; Sturgeon, K.; Zaslavsky, A.; Chen, C.S.; et al. Tumor vessel normalization after aerobic exercise enhances chemotherapeutic efficacy. Oncotarget 2016, 7, 65429–65440. [Google Scholar] [CrossRef] [Green Version]
- Ashcraft, K.A.; Warner, A.B.; Jones, L.W.; Dewhirst, M.W. Exercise as Adjunct Therapy in Cancer. Semin. Radiat. Oncol. 2019, 29, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Idorn, M.; Hojman, P. Exercise-Dependent Regulation of NK Cells in Cancer Protection. Trends Mol. Med. 2016, 22, 565–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, B.K.; Febbraio, M.A. Muscle as an endocrine organ: Focus on muscle-derived interleukin-6. Physiol. Rev. 2008, 88, 1379–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bay, M.L.; Heywood, S.; Wedell-Neergaard, A.S.; Schauer, T.; Lehrskov, L.L.; Christensen, R.H.; Legard, G.E.; Jensen, P.O.; Krogh-Madsen, R.; Ellingsgaard, H. Human immune cell mobilization during exercise: Effect of IL-6 receptor blockade. Exp. Physiol. 2020, 105, 2086–2098. [Google Scholar] [CrossRef] [PubMed]
- Premkumar, V.G.; Yuvaraj, S.; Vijayasarathy, K.; Gangadaran, S.G.; Sachdanandam, P. Serum cytokine levels of interleukin-1beta, -6, -8, tumour necrosis factor-alpha and vascular endothelial growth factor in breast cancer patients treated with tamoxifen and supplemented with co-enzyme Q(10), riboflavin and niacin. Basic Clin. Pharm. Toxicol. 2007, 100, 387–391. [Google Scholar] [CrossRef]
- Sprod, L.K.; Palesh, O.G.; Janelsins, M.C.; Peppone, L.J.; Heckler, C.E.; Adams, M.J.; Morrow, G.R.; Mustian, K.M. Exercise, sleep quality, and mediators of sleep in breast and prostate cancer patients receiving radiation therapy. Community Oncol. 2010, 7, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Hawley, J.E.; Pan, S.; Figg, W.D.; Lopez-Bujanda, Z.A.; Strope, J.D.; Aggen, D.H.; Dallos, M.C.; Lim, E.A.; Stein, M.N.; Hu, J.; et al. Association between immunosuppressive cytokines and PSA progression in biochemically recurrent prostate cancer treated with intermittent hormonal therapy. Prostate 2020, 80, 336–344. [Google Scholar] [CrossRef]
- Hojman, P. Exercise protects from cancer through regulation of immune function and inflammation. Biochem Soc. Trans. 2017, 45, 905–911. [Google Scholar] [CrossRef]
- Dethlefsen, C.; Lillelund, C.; Midtgaard, J.; Andersen, C.; Pedersen, B.K.; Christensen, J.F.; Hojman, P. Exercise regulates breast cancer cell viability: Systemic training adaptations versus acute exercise responses. Breast Cancer Res. Treat. 2016, 159, 469–479. [Google Scholar] [CrossRef]
- Hwang, J.H.; McGovern, J.; Minett, G.M.; Della Gatta, P.A.; Roberts, L.; Harris, J.M.; Thompson, E.W.; Parker, T.J.; Peake, J.M.; Neubauer, O. Mobilizing serum factors and immune cells through exercise to counteract age-related changes in cancer risk. Exerc. Immunol. Rev. 2020, 26, 80–99. [Google Scholar]
- Antoni, M.H.; Dhabhar, F.S. The impact of psychosocial stress and stress management on immune responses in patients with cancer. Cancer 2019, 125, 1417–1431. [Google Scholar] [CrossRef]
- Pasquini, M.; Biondi, M. Depression in cancer patients: A critical review. Clin. Pr. Epidemiol Ment. Health 2007, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Wurtman, R.J. Stress and the adrenocortical control of epinephrine synthesis. Metabolism 2002, 51, 11–14. [Google Scholar] [CrossRef] [Green Version]
- Hassan, S.; Karpova, Y.; Flores, A.; D’Agostino, R., Jr.; Danhauer, S.C.; Hemal, A.; Kulik, G. A pilot study of blood epinephrine levels and CREB phosphorylation in men undergoing prostate biopsies. Int. Urol. Nephrol. 2014, 46, 505–510. [Google Scholar] [CrossRef] [Green Version]
- Thornton, L.M.; Andersen, B.L.; Blakely, W.P. The pain, depression, and fatigue symptom cluster in advanced breast cancer: Covariation with the hypothalamic-pituitary-adrenal axis and the sympathetic nervous system. Health Psychol. 2010, 29, 333–337. [Google Scholar] [CrossRef] [Green Version]
- Mausbach, B.T.; Aschbacher, K.; Mills, P.J.; Roepke, S.K.; von Kanel, R.; Patterson, T.L.; Dimsdale, J.E.; Ziegler, M.G.; Ancoli-Israel, S.; Grant, I. A 5-year longitudinal study of the relationships between stress, coping, and immune cell beta(2)-adrenergic receptor sensitivity. Psychiatry Res. 2008, 160, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Krizanova, O.; Babula, P.; Pacak, K. Stress, catecholaminergic system and cancer. Stress 2016, 19, 419–428. [Google Scholar] [CrossRef]
- Wright, J.J.; Powers, A.C.; Johnson, D.B. Endocrine toxicities of immune checkpoint inhibitors. Nat. Rev. Endocrinol 2021. [Google Scholar] [CrossRef]
- Grouthier, V.; Lebrun-Vignes, B.; Moey, M.; Johnson, D.B.; Moslehi, J.J.; Salem, J.E.; Bachelot, A. Immune Checkpoint Inhibitor-Associated Primary Adrenal Insufficiency: WHO VigiBase Report Analysis. Oncologist 2020, 25, 696–701. [Google Scholar] [CrossRef]
- Hagstrom, A.D.; Marshall, P.W.; Lonsdale, C.; Papalia, S.; Cheema, B.S.; Toben, C.; Baune, B.T.; Fiatarone Singh, M.A.; Green, S. The effect of resistance training on markers of immune function and inflammation in previously sedentary women recovering from breast cancer: A randomized controlled trial. Breast Cancer Res. Treat. 2016, 155, 471–482. [Google Scholar] [CrossRef]
- Hutnick, N.A.; Williams, N.I.; Kraemer, W.J.; Orsega-Smith, E.; Dixon, R.H.; Bleznak, A.D.; Mastro, A.M. Exercise and lymphocyte activation following chemotherapy for breast cancer. Med. Sci. Sports Exerc. 2005, 37, 1827–1835. [Google Scholar] [CrossRef]
- Nieman, D.C.; Cook, V.D.; Henson, D.A.; Suttles, J.; Rejeski, W.J.; Ribisl, P.M.; Fagoaga, O.R.; Nehlsen-Cannarella, S.L. Moderate exercise training and natural killer cell cytotoxic activity in breast cancer patients. Int. J. Sports Med. 1995, 16, 334–337. [Google Scholar] [CrossRef] [PubMed]
- Peters, C.; Lotzerich, H.; Niemeier, B.; Schule, K.; Uhlenbruck, G. Influence of a moderate exercise training on natural killer cytotoxicity and personality traits in cancer patients. Anticancer. Res. 1994, 14, 1033–1036. [Google Scholar]
- Fairey, A.S.; Courneya, K.S.; Field, C.J.; Bell, G.J.; Jones, L.W.; Mackey, J.R. Randomized controlled trial of exercise and blood immune function in postmenopausal breast cancer survivors. J. Appl. Physiol. 2005, 98, 1534–1540. [Google Scholar] [CrossRef] [Green Version]
- Galvao, D.A.; Nosaka, K.; Taaffe, D.R.; Peake, J.; Spry, N.; Suzuki, K.; Yamaya, K.; McGuigan, M.R.; Kristjanson, L.J.; Newton, R.U. Endocrine and immune responses to resistance training in prostate cancer patients. Prostate Cancer Prostatic Dis. 2008, 11, 160–165. [Google Scholar] [CrossRef]
- Khosravi, N.; Hanson, E.D.; Farajivafa, V.; Evans, W.S.; Lee, J.T.; Danson, E.; Wagoner, C.W.; Harrell, E.P.; Sullivan, S.A.; Nyrop, K.A.; et al. Exercise-induced modulation of monocytes in breast cancer survivors. Brain Behav. Immun. Health 2021. in Press. [Google Scholar]
- Hanson, E.D.; Bates, L.C.; Harrell, E.P.; Bartlett, D.B.; Lee, J.T.; Wagoner, C.W.; Alzer, M.S.; Amatuli, D.J.; Jensen, B.C.; Deal, A.M.; et al. Exercise Training Partially Rescues Impaired Mucosal Associated Invariant T-Cell Mobilization in Breast Cancer Survivors Compared to Healthy Older Women; University of North Carolina: Chapel Hill, NC, USA, Unpublished work; 2021. [Google Scholar]
- Bartlett, D.B.; Hanson, E.D.; Lee, J.T.; Wagoner, C.W.; Harrell, E.P.; Sullivan, S.A.; Bates, L.C.; Deal, A.M.; Jensen, B.C.; MacDonald, G.; et al. The Effects of 16-Weeks of Exercise Training on Neutrophil Functions in Breast Cancer Survivors; Duke University: Durham, NC, USA, Unpublished work; 2021. [Google Scholar]
- Conceicao, F.; Sousa, D.M.; Paredes, J.; Lamghari, M. Sympathetic activity in breast cancer and metastasis: Partners in crime. Bone Res. 2021, 9, 1–11. [Google Scholar] [CrossRef]
- Johnsson, A.; Demmelmaier, I.; Sjovall, K.; Wagner, P.; Olsson, H.; Tornberg, A.B. A single exercise session improves side-effects of chemotherapy in women with breast cancer: An observational study. BMC Cancer 2019, 19, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hanson, E.D.; Bates, L.C.; Moertl, K.; Evans, E.S. Natural Killer Cell Mobilization in Breast and Prostate Cancer Survivors: The Implications of Altered Stress Hormones Following Acute Exercise. Endocrines 2021, 2, 121-132. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2020012
Hanson ED, Bates LC, Moertl K, Evans ES. Natural Killer Cell Mobilization in Breast and Prostate Cancer Survivors: The Implications of Altered Stress Hormones Following Acute Exercise. Endocrines. 2021; 2(2):121-132. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2020012
Chicago/Turabian StyleHanson, Erik D., Lauren C. Bates, Kaileigh Moertl, and Elizabeth S. Evans. 2021. "Natural Killer Cell Mobilization in Breast and Prostate Cancer Survivors: The Implications of Altered Stress Hormones Following Acute Exercise" Endocrines 2, no. 2: 121-132. https://0-doi-org.brum.beds.ac.uk/10.3390/endocrines2020012