Coconut Coir as a Sustainable Nursery Growing Media for Seedling Production of the Ecologically Diverse Quercus Species

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nursery Stock Cultivation
2.2. Data Collection
2.3. Statistical Analysis
3. Results
3.1. Emergence, Growth, and Physiological Traits during Nursery Cultivation
3.2. Morphological Traits
3.3. N, P, K Concentration in Seedlings and Substrate at the End of Cultivation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wilson, B.C.; Jacobs, D.F. Quality assessment of temperate zone deciduous hardwood seedlings. New For. 2006, 31, 417–433. [Google Scholar] [CrossRef]
- Pinto, J.R.; Dumroese, R.K.; Davis, A.S.; Landis, T.D. Conducting seedling stocktype trials: A new approach to an old question. J. For. 2011, 109, 293–299. [Google Scholar]
- Dumroese, K.R.; Landis, T.D.; Pinto, J.R.; Haase, D.L.; Wilkinson, K.W.; Davis, A.S. Meeting forest restoration challenges: Using the target plant concept. Reforesta 2016, 1, 37–52. [Google Scholar] [CrossRef] [Green Version]
- Duryea, M.L. Nursery cultural practices: Impacts on seedling quality. In Forestry Nursery Manual: Production of Bareroot Seedlings; Springer: Berlin, Germany, 1984; pp. 143–164. [Google Scholar]
- Landis, T.D. The target plant concept-a history and brief overview. In National Proceedings: Forest and Conservation Nursery Associations-2010; Proc. Proc. RMRS-P-65; Riley, L.E., Haase, D.L., Pinto, J.R., Eds.; USDA Forest Service, Rocky Mountain Research Station: Fort Collins, CO, USA, 2011; Volume 65, pp. 61–66. [Google Scholar]
- Haase, D.L.; Davis, A.S. Developing and supporting quality nursery facilities and staff are necessary to meet global forest and landscape restoration needs. Reforesta 2017, 4, 69–93. [Google Scholar] [CrossRef] [Green Version]
- Pemán, J.; Chirino, E.; Espelta, J.M.; Jacobs, D.F.; Martín-Gómez, P.; Navarro-Cerrillo, R.; Oliet, J.A.; Vilagrosa, A.; Villar-Salvador, P.; Gil-Pelegrín, E. Physiological keys for natural and artificial regeneration of oaks. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Springer: Berlin, Germany, 2017; pp. 453–511. [Google Scholar]
- Landis, T.D.; Jacobs, D.F.; Wilkinson, K.M.; Luna, T. Growing Media. In Tropical Nursery Manual—A Guide to Starting and Operating a Nursery for Native and Traditional Plants; Agriculture Handbook; Wilkinson, K.M., Landis, T.D., Haase, D.L., Daley, B.F., Dumroese, R.K., Eds.; USDA Forest Service: Washington, DC, USA, 2014; Volume 732, pp. 101–122. [Google Scholar]
- Barrett, G.E.; Alexander, P.D.; Robinson, J.S.; Bragg, N.C. Achieving environmentally sustainable growing media for soilless plant cultivation systems–A review. Sci. Hortic. 2016, 212, 220–234. [Google Scholar] [CrossRef] [Green Version]
- Landis, T.D.; Tinus, R.W.; McDonald, S.E.; Barnett, J.P. Containers and Growing Media. In The Container Tree Nursery Manual; Agriculture Handbook; Landis, T.D., Nisley, R.G., Eds.; USDA Forest Service: Washington, DC, USA, 1990; Volume 2, pp. 41–85. [Google Scholar]
- Tsakaldimi, M.; Ganatsas, P. A synthesis of results on wastes as potting media substitutes for the production of native plant species. Reforesta 2016, 1, 147–163. [Google Scholar] [CrossRef]
- Caron, J.; Rochefort, L. Use of peat in growing media: State of the art on industrial and scientific efforts envisioning sustainability. Acta Hortic. 2013, 982, 15–22. [Google Scholar] [CrossRef]
- Ángeles-Argáiz, R.E.; Flores-García, A.; Ulloa, M.; Garibay-Orijel, R. Commercial Sphagnum peat moss is a vector for exotic ectomycorrhizal mushrooms. Biol. Invasions 2016, 18, 89–101. [Google Scholar] [CrossRef] [Green Version]
- Schmilewski, G. Growing media constituents used in the EU in 2013. Acta Hortic. 2017, 1168, 85–92. [Google Scholar] [CrossRef]
- Apodaca, L.E. Peat in 2015. Min. Eng. 2016, 68, 3030. [Google Scholar]
- Clarke, D.; Rieley, J. Strategy for Responsible Peatland Management, 6th ed.; International Peat Society: Jyväskylä, Finland, 2010; pp. 10–25. [Google Scholar]
- Schmilewski, G. The role of peat in assuring the quality of growing media. Mires Peat 2008, 3, article 2. [Google Scholar]
- Michel, J.-C. The physical properties of peat: A key factor for modern growing media. Mires Peat 2010, 6, article 2. [Google Scholar]
- Maher, M.; Prasad, M. Organic soilless media components. In Soilless Culture: Theory and Practice, 1st ed.; Raviv, M., Lierh, J.H., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2008; pp. 479–481. [Google Scholar]
- Alexander, P.D.; Bragg, N.C.; Meade, R.; Padelopoulos, G.; Watts, O. Peat in horticulture and conservation: The UK response to a changing world. Mires Peat 2008, 3, 10. [Google Scholar]
- Carlile, B.; Coules, A. Towards sustainability in growing media. Acta Hortic. 2013, 1013, 341–349. [Google Scholar] [CrossRef]
- Bonn, A.; Reed, M.S.; Evans, C.D.; Joosten, H.; Bain, C.; Farmer, J.; Emmer, I.; Couwenberg, J.; Moxey, A.; Artz, R. Investing in nature: Developing ecosystem service markets for peatland restoration. Ecosyst. Serv. 2014, 9, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Rochefort, L.; Lode, E. Restoration of degraded boreal peatlands. In Boreal Peatland Ecosystems; Wieder, R.K., Vitt, D.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2006; Volume 188, pp. 381–423. [Google Scholar]
- Kimmel, K.; Mander, Ü. Ecosystem services of peatlands: Implications for restoration. Prog. Phys. Geogr. 2010, 34, 491–514. [Google Scholar] [CrossRef]
- Parry, L.E.; Holden, J.; Chapman, P.J. Restoration of blanket peatlands. J. Environ. Manag. 2014, 133, 193–205. [Google Scholar] [CrossRef] [Green Version]
- Dunn, C.; Freeman, C. Peatlands: Our greatest source of carbon credits? Carbon Manag. 2011, 2, 289–301. [Google Scholar] [CrossRef]
- Ugolini, F.; Mariotti, B.; Maltoni, A.; Tani, A.; Salbitano, F.; Izquierdo, C.G.; Macci, C.; Masciandaro, G.; Tognetti, R. A tree from waste: Decontaminated dredged sediments for growing forest tree seedlings. J. Environ. Manag. 2018, 211, 269–277. [Google Scholar] [CrossRef]
- Wallace, P.; Holmes, S.; Alexander, R.; England, J.; Gaze, R. Review of Growing Media Use and Dominant Materials (Peat and Alternatives) for Growing Media in Other Countries (European and International); Final Report; DEFRA Project SP1206; Department for Environment, Food and Rural Affairs: London, UK, 2010.
- Ceglie, F.G.; Bustamante, M.A.; Amara, M.B.; Tittarelli, F. The challenge of peat substitution in organic seedling production: Optimization of growing media formulation through mixture design and response surface analysis. PLoS ONE 2015, 10, e0128600. [Google Scholar] [CrossRef]
- Gruda, N.S. Increasing sustainability of growing media constituents and stand-alone substrates in soilless culture systems. Agronomy 2019, 9, 298. [Google Scholar] [CrossRef] [Green Version]
- Handreck, K.A. Properties of coir dust, and its use in the formulation of soilless potting media. Commun. Soil Sci. Plant Anal. 1993, 24, 349–363. [Google Scholar] [CrossRef]
- Schmilewski, G. Peat covers 77 percent of the growing media production in the EU. Peatl. Int. 2008, 1, 39–43. [Google Scholar]
- Nichols, M.A. Coir—A XXIst Century sustainable growing medium. Acta Hortic. 2007, 747, 91–95. [Google Scholar] [CrossRef]
- Blok, C.; Wever, G. Experience with selected physical methods to characterize the suitability of growing media for plant growth. Acta Hortic. 2008, 779, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Poulter, R. Quantifying differences between treated and untreated coir substrate. Acta Hortic. 2014, 1018, 557–564. [Google Scholar] [CrossRef]
- Gruda, N. Current and future perspective of growing media in Europe. Acta Hortic. 2012, 960, 37–43. [Google Scholar] [CrossRef]
- Poorter, H.; Nagel, O. The role of biomass allocation in the growth response of plants to different levels of light, CO2, nutrients and water: A quantitative review. Funct. Plant Biol. 2000, 27, 1191. [Google Scholar] [CrossRef] [Green Version]
- Poorter, H.; Niklas, K.J.; Reich, P.B.; Oleksyn, J.; Poot, P.; Mommer, L. Biomass allocation to leaves, stems and roots: Meta-analyses of interspecific variation and environmental control. New Phytol. 2012, 193, 30–50. [Google Scholar] [CrossRef]
- Domínguez, M.T.; Aponte, C.; Pérez-Ramos, I.M.; García, L.V.; Villar, R.; Marañón, T. Relationships between leaf morphological traits, nutrient concentrations and isotopic signatures for Mediterranean woody plant species and communities. Plant Soil 2012, 357, 407–424. [Google Scholar] [CrossRef] [Green Version]
- Villar-Salvador, P.; Planelles, R.; Enrıquez, E.; Rubira, J.P. Nursery cultivation regimes, plant functional attributes, and field performance relationships in the Mediterranean oak Quercus ilex L. For. Ecol. Manag. 2004, 196, 257–266. [Google Scholar] [CrossRef]
- Hernández, E.I.; Vilagrosa, A.; Luis, V.C.; Llorca, M.; Chirino, E.; Vallejo, V.R. Root hydraulic conductance, gas exchange and leaf water potential in seedlings of Pistacia lentiscus L. and Quercus suber L. grown under different fertilization and light regimes. Environ. Exp. Bot. 2009, 67, 269–276. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2012, 43, 711–738. [Google Scholar] [CrossRef]
- Cortina, J.; Vilagrosa, A.; Trubat, R. The role of nutrients for improving seedling quality in drylands. New For. 2013, 44, 719–732. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; MacDonald, J.E. Seedling Quality: History, application, and plant attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.A.; Planelles, R.; Artero, F.; Jacobs, D.F. Nursery fertilization and tree shelters affect long-term field response of Acacia salicina Lindl. planted in Mediterranean semiarid conditions. For. Ecol. Manag. 2005, 215, 339–351. [Google Scholar] [CrossRef]
- Ovalle, J.F.; Arellano, E.C.; Oliet, J.A.; Becerra, P.; Ginocchio, R. Linking nursery nutritional status and water availability post-planting under intense summer drought: The case of a South American Mediterranean tree species. IForest-Biogeosci. For. 2016, 9, 758. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.A.; Salazar, J.M.; Villar, R.; Robredo, E.; Valladares, F. Fall fertilization of Holm oak affects N and P dynamics, root growth potential, and post-planting phenology and growth. Ann. For. Sci. 2011, 68, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Villar-Salvador, P.; Valladares, F.; Domínguez-Lerena, S.; Ruiz-Díez, B.; Fernández-Pascual, M.; Delgado, A.; Peñuelas, J.L. Functional traits related to seedling performance in the Mediterranean leguminous shrub Retama sphaerocarpa: Insights from a provenance, fertilization, and rhizobial inoculation study. Environ. Exp. Bot. 2008, 64, 145–154. [Google Scholar] [CrossRef]
- Lambers, H.; Shane, M.W.; Cramer, M.D.; Pearse, S.J.; Veneklaas, E.J. Root structure and functioning for efficient acquisition of phosphorus: Matching morphological and physiological traits. Ann. Bot. 2006, 98, 693–713. [Google Scholar] [CrossRef] [Green Version]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Plant morphology and root hydraulics are altered by nutrient deficiency in Pistacia lentiscus (L.). Trees 2006, 20, 334. [Google Scholar] [CrossRef]
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trubat, R.; Cortina, J.; Vilagrosa, A. Root architecture and hydraulic conductance in nutrient deprived Pistacia lentiscus L. seedlings. Oecologia 2012, 170, 899–908. [Google Scholar] [CrossRef] [PubMed]
- Lugli, L.F.; Anderson, K.M.; Aragão, L.E.; Cordeiro, A.L.; Cunha, H.F.; Fuchslueger, L.; Meir, P.; Mercado, L.M.; Oblitas, E.; Quesada, C.A.; et al. Multiple phosphorus acquisition strategies adopted by fine roots in low-fertility soils in Central Amazonia. Plant Soil 2019, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fernández, M.; Marcos, C.; Tapias, R.; Ruiz, F.; López, G. Nursery fertilisation affects the frost-tolerance and plant quality of Eucalyptus globulus Labill. cuttings. Ann. For. Sci. 2007, 64, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.A.; Planelles, R.; Artero, F.; Valverde, R.; Jacobs, D.F.; Segura, M.L. Field performance of Pinus halepensis planted in Mediterranean arid conditions: Relative influence of seedling morphology and mineral nutrition. New For. 2009, 37, 313–331. [Google Scholar] [CrossRef]
- Oliet, J.A.; Puértolas, J.; Planelles, R.; Jacobs, D.F. Nutrient loading of forest tree seedlings to promote stress resistance and field performance: A Mediterranean perspective. New For. 2013, 44, 649–669. [Google Scholar] [CrossRef]
- Egilla, J.N.; Davies, F.T.; Drew, M.C. Effect of potassium on drought resistance of Hibiscus rosa-sinensis cv. Leprechaun: Plant growth, leaf macro-and micronutrient content and root longevity. Plant Soil 2001, 229, 213–224. [Google Scholar] [CrossRef]
- Egilla, J.N.; Davies, F.T.; Boutton, T.W. Drought stress influences leaf water content, photosynthesis, and water-use efficiency of Hibiscus rosa-sinensis at three potassium concentrations. Photosynthetica 2005, 43, 135–140. [Google Scholar] [CrossRef]
- Asgharipour, M.R.; Heidari, M. Effect of potassium supply on drought resistance in sorghum: Plant growth and macronutrient content. Pak. J. Agric. Sci. 2011, 48, 197–204. [Google Scholar]
- Ragel, P.; Raddatz, N.; Leidi, E.O.; Quintero, F.J.; Pardo, J.M. Regulation of K+ nutrition in plants. Front. Plant Sci. 2019, 10, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, R.; Haase, D.L. The use of coir as a containerized growing medium for Douglas-fir seedlings. Native Plants J. 2000, 1, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Radjagukguk, B.; Soeseno, O. A comparative study of peats and other media for containerized forest tree seedlings. Acta Hortic. 1983, 150, 449–458. [Google Scholar] [CrossRef]
- Oliet, J.A.; Jacobs, D.F. Restoring forests: Advances in techniques and theory. New For. 2012, 43, 535–541. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Castro, J.; Delgado-Capel, M.J.; Molinas-González, C.; Pulgar, M.; Marañón-Jiménez, S.; Delgado-Huertas, A.; Querejeta, J.I. Restoring for the present or restoring for the future: Enhanced performance of two sympatric oaks (Quercus ilex and Quercus pyrenaica) above the current forest limit. Restor. Ecol. 2015, 23, 936–946. [Google Scholar] [CrossRef]
- Madrigal-González, J.; Ruiz-Benito, P.; Ratcliffe, S.; Rigling, A.; Wirth, C.; Zimmermann, N.E.; Zweifel, R.; Zavala, M.A. Competition Drives Oak Species Distribution and Functioning in Europe: Implications Under Global Change. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Gil-Pelegrín, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer: New York, NY, USA, 2017; pp. 513–538. [Google Scholar]
- Löf, M.; Castro, J.; Engman, M.; Leverkus, A.B.; Madsen, P.; Reque, J.A.; Villalobos, A.; Gardiner, E.S. Tamm Review: Direct seeding to restore oak (Quercus spp.) forests and woodlands. For. Ecol. Manag. 2019, 448, 474–489. [Google Scholar] [CrossRef]
- Gil-Pelegrín, E.; Saz, M.Á.; Cuadrat, J.M.; Peguero-Pina, J.J.; Sancho-Knapik, D. Oaks under Mediterranean-type climates: Functional response to summer aridity. In Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Gil-Pelegrín, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer: New York, NY, USA, 2017; pp. 137–193. [Google Scholar]
- Ducousso, A.; Bordacs, S. EUFORGEN Technical Guidelines for Genetic Conservation and Use for Pedunculate and Sessile Oaks (Quercus robur) and (Quercus petraea); International Plant Genetic Resources Institute: Rome, Italy, 2004. [Google Scholar]
- Löf, M.; Bolte, A.; Jacobs, D.F.; Jensen, A.M. Nurse trees as a forest restoration tool for mixed plantations: Effects on competing vegetation and performance in target tree species. Restor. Ecol. 2014, 22, 758–765. [Google Scholar] [CrossRef]
- ISTA. International Rules for Seed Testing; International Seed Testing Association: Bassersdorf, Switzerland, 2020. [Google Scholar]
- Strasser, R.J.; Tsimilli-Michael, M.; Srivastava, A. Analysis of the Chlorophyll a Fluorescence Transient. In Chlorophyll a Fluorescence: A Signature of Photosynthesis; Advances in Photosynthesis and Respiration; Papageorgiou, G.C., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 321–362. ISBN 978-1-4020-3218-9. [Google Scholar]
- Villar-Salvador, P.; Heredia, N.; Millard, P. Remobilization of acorn nitrogen for seedling growth in holm oak (Quercus ilex), cultivated with contrasting nutrient availability. Tree Physiol. 2010, 30, 257–263. [Google Scholar] [CrossRef]
- García-Cebrián, F.; Esteso-Martínez, J.; Gil-Pelegrín, E. Influence of cotyledon removal on early seedling growth in Quercus robur L. Ann. For. Sci. 2003, 60, 69–73. [Google Scholar] [CrossRef] [Green Version]
- Quero, J.L.; Villar, R.; Marañón, T.; Zamora, R.; Poorter, L. Seed-mass effects in four Mediterranean Quercus species (Fagaceae) growing in contrasting light environments. Am. J. Bot. 2007, 94, 1795–1803. [Google Scholar] [CrossRef] [Green Version]
- Chirino, E.; Vilagrosa, A.; Hernández, E.I.; Matos, A.; Vallejo, V.R. Effects of a deep container on morpho-functional characteristics and root colonization in Quercus suber L. seedlings for reforestation in Mediterranean climate. For. Ecol. Manag. 2008, 256, 779–785. [Google Scholar] [CrossRef]
- Mariotti, B.; Maltoni, A.; Jacobs, D.F.; Tani, A. Container effects on growth and biomass allocation in Quercus robur and Juglans regia seedlings. Scand. J. For. Res. 2015, 30, 401–415. [Google Scholar]
- Mariotti, B.; Maltoni, A.; Jacobs, D.F.; Tani, A. Tree shelters affect shoot and root system growth and structure in Quercus robur during regeneration establishment. Eur. J. For. Res. 2015, 134, 641–652. [Google Scholar] [CrossRef]
- Cabral, R.; O’Reilly, C. Physiological and field growth responses of oak seedlings to warm storage. New For. 2008, 36, 159–170. [Google Scholar] [CrossRef]
- Tsakaldimi, M.; Zagas, T.; Tsitsoni, T.; Ganatsas, P. Root Morphology, Stem Growth and Field Performance of Seedlings of Two Mediterranean Evergreen Oak Species Raised in Different Container Types. Plant Soil 2005, 278, 85–93. [Google Scholar] [CrossRef]
- Del Campo, A.D.; Navarro, R.M.; Ceacero, C.J. Seedling quality and field performance of commercial stocklots of containerized holm oak (Quercus ilex) in Mediterranean Spain: An approach for establishing a quality standard. New For. 2010, 39, 19. [Google Scholar] [CrossRef]
- Dlgs 386/03. Available online: https://www.camera.it/parlam/leggi/deleghe/03386dl.htm (accessed on 1 May 2020).
- Offord, C.A.; Muir, S.; Tyler, J.L. Growth of selected Australian plants in soilless media using coir as a substitute for peat. Aust. J. Exp. Agric. 1998, 38, 879–887. [Google Scholar] [CrossRef]
- Handreck, K.A. Immobilization of nitrogen in potting media. Acta Hortic. 1993, 342, 121–126. [Google Scholar] [CrossRef]
- Grantzau, E.; Gennrich, J.; DP, D. Mit Kokos Substrate verbessern. Gb Gw 1993, 11, 538–541. [Google Scholar]
- Noguera, P.; Abad, M.; Noguera, V.; Puchades, R.; Maquieira, A. Coconut coir waste, a new and viable ecologically-friendly peat substitute. Acta Hortic. 2000, 517, 279–286. [Google Scholar] [CrossRef]
- Meerow, A.W. Growth of two subtropical ornamentals using coir (coconut mesocarp pith) as a peat substitute. HortScience 1994, 29, 1484–1486. [Google Scholar] [CrossRef] [Green Version]
- Stamps, R.H.; Evans, M.R. Growth of Dracaena marginata and Spathiphyllum ‘Petite’in sphagnum peat-and coconut coir dust-based growing media. J. Environ. Hortic. 1999, 17, 49–52. [Google Scholar]
- Wang, M.; Zheng, Q.; Shen, Q.; Guo, S. The critical role of potassium in plant stress response. Int. J. Mol. Sci. 2013, 14, 7370–7390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripler, C.E.; Kaushal, S.S.; Likens, G.E.; Todd Walter, M. Patterns in potassium dynamics in forest ecosystems. Ecol. Lett. 2006, 9, 451–466. [Google Scholar] [CrossRef] [PubMed]
- Santiago, L.S.; Wright, S.J.; Harms, K.E.; Yavitt, J.B.; Korine, C.; Garcia, M.N.; Turner, B.L. Tropical tree seedling growth responses to nitrogen, phosphorus and potassium addition. J. Ecol. 2012, 100, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Soliveres, S.; Maestre, F.T. Plant–plant interactions, environmental gradients and plant diversity: A global synthesis of community-level studies. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 154–163. [Google Scholar] [CrossRef] [Green Version]
- Uscola, M.; Salifu, K.F.; Oliet, J.A.; Jacobs, D.F. An exponential fertilization dose–response model to promote restoration of the Mediterranean oak Quercus ilex. New For. 2015, 46, 795–812. [Google Scholar] [CrossRef] [Green Version]
- Salifu, K.F.; Jacobs, D.F. Characterizing fertility targets and multi-element interactions in nursery culture of Quercus rubra seedlings. Ann. For. Sci. 2006, 63, 231–237. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Peñuelas, J.L.; Nicolás-Peragón, J.L.; Benito, L.F.; Domínguez-Lerena, S. Is nitrogen fertilization in the nursery a suitable tool for enhancing the performance of Mediterranean oak plantations? New For. 2013, 44, 733–751. [Google Scholar] [CrossRef]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391–408. [Google Scholar] [CrossRef]
- Postma, J.A.; Dathe, A.; Lynch, J.P. The optimal lateral root branching density for maize depends on nitrogen and phosphorus availability. Plant Physiol. 2014, 166, 590–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Péret, B.; Desnos, T.; Jost, R.; Kanno, S.; Berkowitz, O.; Nussaume, L. Root architecture responses: In search of phosphate. Plant Physiol. 2014, 166, 1713–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colla, G.; Rouphael, Y.; Possanzini, G.; Cardarelli, M.; Temperini, O.; Saccardo, F.; Pierandrei, F.; Rea, E. Coconut coir as a potting media for organic lettuce transplant production. In VIII International Symposium on Protected Cultivation in Mild Winter Climates: Advances in Soil and Soilless Cultivation under Protected Environment; International Society for Horticultural Science: Leuven, Belgium, 2006; pp. 293–296. [Google Scholar]
- Chulaka, P.; Maruo, T.; Takagaki, M.; Shinohara, Y. Organic substrates of tropical origin as an alternative to growing media for chili and cucumber transplant production. Jpn. J. Trop. Agric. 2004, 48, 79–87. [Google Scholar]
- Wilson, S.B.; Muller, K.L.; Wilson, P.C.; Incer, M.R.; Stoffella, P.J.; Graetz, D.A. Evaluation of new container media for Aglaonema production. Commun. Soil Sci. Plant Anal. 2009, 40, 2673–2687. [Google Scholar] [CrossRef]
- Lloret, F.; Casanovas, C.; Penuelas, J. Seedling survival of Mediterranean shrubland species in relation to root: Shoot ratio, seed size and water and nitrogen use. Funct. Ecol. 1999, 13, 210–216. [Google Scholar] [CrossRef]
- Zida, D.; Tigabu, M.; Sawadogo, L.; Odén, P.C. Initial seedling morphological characteristics and field performance of two Sudanian savanna species in relation to nursery production period and watering regimes. For. Ecol. Manag. 2008, 255, 2151–2162. [Google Scholar] [CrossRef]
- Villar-Salvador, P.; Puértolas, J.; Cuesta, B.; Peñuelas, J.L.; Uscola, M.; Heredia-Guerrero, N.; Rey Benayas, J.M. Increase in size and nitrogen concentration enhances seedling survival in Mediterranean plantations. Insights from an ecophysiological conceptual model of plant survival. New For. 2012, 43, 755–770. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J.; Rodà, F. Plasticity of leaf morphological traits, leaf nutrient content, and water capture in the Mediterranean evergreen oak Quercus ilex subsp. ballota in response to fertilization and changes in competitive conditions. Ecoscience 2006, 13, 258–270. [Google Scholar] [CrossRef]
- Vollmar, A.; Gunderson, C. Physiological adjustments of leaf respiration to atmospheric warming in Betula alleghaniensis and Quercus rubra. J. Undergrad. Res. 2006, 6, 104–107. [Google Scholar]
- Salifu, K.F.; Apostol, K.G.; Jacobs, D.F.; Islam, M.A. Growth, physiology, and nutrient retranslocation in nitrogen-15 fertilized Quercus rubra seedlings. Ann. For. Sci. 2008, 65, 101. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.A.; Tejada, M.; Salifu, K.F.; Collazos, A.; Jacobs, D.F. Performance and nutrient dynamics of holm oak (Quercus ilex L.) seedlings in relation to nursery nutrient loading and post-transplant fertility. Eur. J. For. Res. 2009, 128, 253–263. [Google Scholar] [CrossRef]
- Berger, T.W.; Glatzel, G. Response of Quercus petraea seedlings to nitrogen fertilization. For. Ecol. Manag. 2001, 149, 1–14. [Google Scholar] [CrossRef]
- Bigg, W.L.; Schalau, J.W. Mineral nutrition and the target seedling. In Target Seedling Symposium: Proceedings, Combined Meeting of the Western Forest Nursery Associations; USDA Forest Service: Fort Collins, CO, USA, 1990; pp. 139–160. [Google Scholar]
- Folk, R.S.; Grossnickle, S.C. Stock-type patterns of phosphorus uptake, retranslocation, net photosynthesis and morphological development in interior spruce seedlings. New For. 2000, 19, 27–49. [Google Scholar] [CrossRef]
- Sardans, J.; Peñuelas, J. Increasing drought decreases phosphorus availability in an evergreen Mediterranean forest. Plant Soil 2004, 267, 367–377. [Google Scholar] [CrossRef]
- Sun, Y.; Gu, J.-C.; Zhuang, H.-F.; Wang, Z.-Q. Effects of ectomycorrhizal colonization and nitrogen fertilization on morphology of root tips in a Larix gmelinii plantation in northeastern China. Ecol. Res. 2010, 25, 295–302. [Google Scholar] [CrossRef]
- Pascual, S.; Olarieta, J.R.; Rodríguez-Ochoa, R. Development of Quercus ilex plantations is related to soil phosphorus availability on shallow calcareous soils. New For. 2012, 43, 805–814. [Google Scholar] [CrossRef]
- Andivia, E.; Fernández, M.; Vázquez-Piqué, J. Autumn fertilization of Quercus ilex ssp. ballota (Desf.) Samp. nursery seedlings: Effects on morpho-physiology and field performance. Ann. For. Sci. 2011, 68, 543. [Google Scholar] [CrossRef] [Green Version]
- Oliet, J.A.; Planelles, R.; López, M.; Artero, F. Efecto de la fertilización en vivero sobre la supervivencia en plantación de Pinus halepensis Mill. Investig. Agrar. Sist. Recur. For. 1997, 8, 207–228. [Google Scholar]
- Puértolas, J.; Gil, L.; Pardos, J.A. Effects of nutritional status and seedling size on field performance of Pinus halepensis planted on former arable land in the Mediterranean basin. Forestry 2003, 76, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Del Campo, A.D.; Hermoso, J.; Flors, J.; Lidón, A.; Navarro-Cerrillo, R.M. Nursery location and potassium enrichment in Aleppo pine stock 2. Performance under real and hydrogel-mediated drought conditions. Forestry 2011, 84, 235–245. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Importance of root growth in overcoming planting stress. New For. 2005, 30, 273–294. [Google Scholar] [CrossRef]



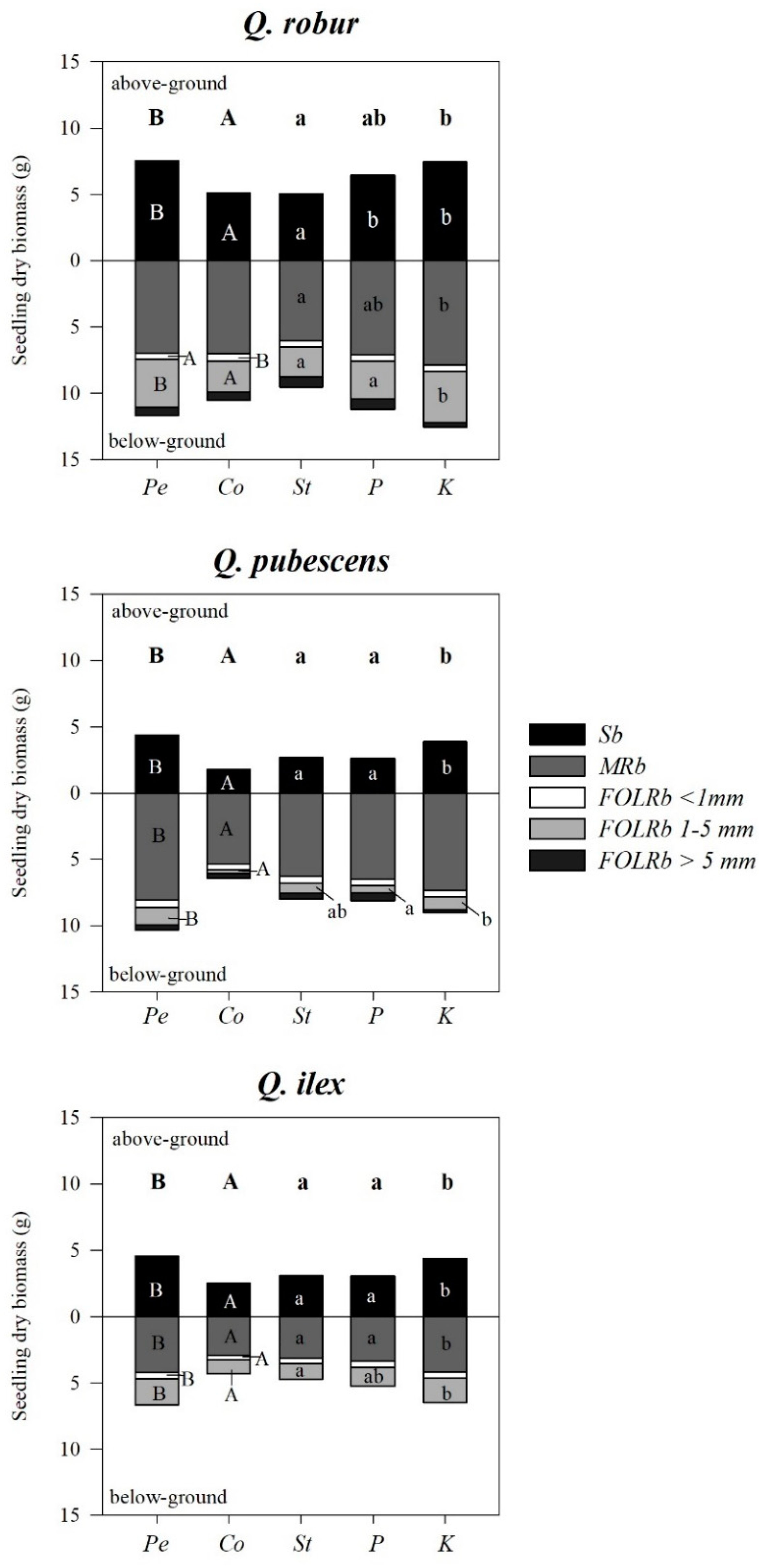{kind=link}
{kind=link}
| Content per Volume mg/L | Content per Seedling (mg per pot) | |||||
|---|---|---|---|---|---|---|
| N | P | K | N | P | K | |
| Standard | 450.0 | 270.0 | 330.0 | 292.5 | 175.5 | 214.5 |
| P enriched | 450.0 | 690.0 | 330.0 | 292.5 | 448.5 | 214.5 |
| K enriched | 440.0 | 440.0 | 720.0 | 286.0 | 286.0 | 468.0 |
| Q. robur | S | F | S × F | Pe | Co | St | P | K |
|---|---|---|---|---|---|---|---|---|
| H | <0.0001 | 0.0108 | 0.8213 | 57.6 ± 10.8 B | 45.8 ± 10.3 A | 48.3 ± 11.4 a | 51.5 ± 12.2 ab | 55.4 ± 11.8 b |
| RcD | 0.2265 | 0.0006 | 0.6354 | 11.2 ± 1.9 | 10.8 ± 1.7 | 10.1 ± 1.8 a | 11.2 ± 1.8 ab | 11.6 ± 1.6 b |
| H/RcD | <0.0001 | 0.4976 | 0.9521 | 52.6 ± 11.3 B | 42.9 ± 9.1 A | 48.4 ± 12.2 | 46.1 ± 9.8 | 48.6 ± 11.9 |
| Rv | 0.0143 | 0.0001 | 0.1355 | 28.0 ± 9.2 B | 24.6 ± 7.8 A | 21.0 ± 6.0 a | 26.9 ± 8.0 b | 31.0 ± 8.8 b |
| Rd | 0.3264 | 0.0040 | 0.0711 | 0.43 ± 0.1 | 0.44 ± 0.1 | 0.47 ± 0.1 b | 0.42 ± 0.1 ab | 0.41 ± 0.1 a |
| S/R | <0.0001 | 0.0419 | 0.6985 | 0.65 ± 0.14 B | 0.49 ± 0.15 A | 0.53 ± 0.17 a | 0.57 ± 0.14 ab | 0.61 ± 0.18 b |
| SLA | 0.0016 | 0.0120 | 0.5480 | 156.1 ± 29.2 B | 142.1 ± 18.1 A | 147.0 ± 20.2 ab | 142.2 ± 32.0 a | 157.9 ± 19.3 b |
| Ln | 0.0156 | <0.0001 | 0.7843 | 56.6 ± 26.0 B | 47.2 ± 19.7 A | 41.6 ± 13.2 a | 48.0 ± 22.0 ab | 66.1 ± 26.2 b |
| Lb | <0.0001 | <0.0001 | 0.2137 | 4.5 ± 1.5 B | 3.4 ± 1.3 A | 3.2 ± 1.1 a | 4.0 ± 1.3 b | 4.7 ± 1.6 b |
| Q. pubescens | S | F | S × F | Pe | Co | St | P | K |
| H | <0.0001 | 0.0028 | 0.4337 | 36.8 ± 11.5 B | 22.4 ± 8.2 A | 27.9 ± 10.7 ab | 27.0 ± 11.1 a | 33.9 ± 14.0 b |
| RcD | 0.0003 | 0.0024 | 0.4202 | 10.7 ± 1.9 B | 9.4 ± 2.2 A | 9.7 ± 1.9 ab | 9.4 ± 2.3 a | 10.9 ± 2.0 b |
| H/RcD | <0.0001 | 0.5218 | 0.5139 | 34.8 ± 10.3 B | 24.2 ± 7.2 A | 28.9 ± 10.6 | 28.9 ± 9.5 | 30.8 ± 11.1 |
| Rv | <0.0001 | 0.3988 | 0.0951 | 25.8 ± 10.5 B | 14.9 ± 6.1 A | 20.9 ± 12.6 | 19.1 ± 8.6 | 21.3 ± 9.0 |
| Rd | 0.0101 | 0.2732 | 0.0215 | 0.41 ± 0.1 A | 0.44 ± 0.1 B | 0.42 ± 0.1 | 0.43 ± 0.16 | 0.42 ± 0.1 |
| S/R | <0.0001 | 0.0001 | 0.0725 | 0.43 ± 0.1 B | 0.27 ± 0.1 A | 0.33 ± 0.1 a | 0.31 ± 0.1 a | 0.42 ± 0.2 b |
| SLA | 0.0600 | 0.6396 | 0.7851 | 104.6 ± 13.4 | 100.6 ± 8.7 | 101.4 ± 9.0 | 103.8 ± 13.9 | 102.7 ± 11.2 |
| Ln | <0.0001 | 0.0232 | 0.4068 | 60.5 ± 27.2 B | 34.9 ± 15.1 A | 46.5 ± 29.7 ab | 41.7 ± 20.5 a | 54.9 ± 24.0 b |
| Lb | <0.0001 | 0.0192 | 0.4390 | 4.3 ± 1.5 B | 2.1 ± 1.1 A | 3.1 ± 1.7 ab | 2.9 ± 1.4 a | 3.7 ± 1.9 b |
| Q. ilex | S | F | S × F | Pe | Co | St | P | K |
| H | <0.0001 | 0.0032 | 0.3332 | 52.7 ± 13.1 B | 35.0 ± 10.3 A | 40.9 ± 13.0 a | 41.8 ± 13.3 ab | 48.9 ± 16.7 b |
| RcD | 0.0114 | 0.1610 | 0.5929 | 8.5 ± 1.1 B | 7.8 ± 1.6 A | 8.1 ± 1.3 | 7.9 ± 1.5 | 8.5 ± 1.3 |
| H/RcD | <0.0001 | 0.0386 | 0.2352 | 62.7 ± 15.4 B | 45.1 ± 12.3 A | 50.1 ± 13.4 a | 53.6 ± 15.7 ab | 58.0 ± 19.2 b |
| Rv | <0.0001 | 0.0009 | 0.4449 | 14.9 ± 5.0 B | 10.7 ± 4.3 A | 10.8 ± 4.2 a | 12.9 ± 5.4 ab | 14.6 ± 5.1 b |
| Rd | 0.3072 | 0.1237 | 0.3875 | 0.45 ± 0.10 | 0.43 ± 0.13 | 0.45 ± 0.13 | 0.41 ± 0.08 | 0.46 ± 0.12 |
| S/R | 0.0062 | 0.3922 | 0.3637 | 0.71 ± 0.3 B | 0.59 ± 0.2 A | 0.68 ± 0.2 | 0.61 ± 0.3 | 0.67 ± 0.2 |
| SLA | 0.0104 | 0.2676 | 0.0005 | 73.6 ± 8.0 B | 70.5 ± 5.5 A | 73.2 ± 7.6 | 70.7 ± 7.1 | 72.2 ± 6.4 |
| Ln | <0.0001 | 0.0140 | 0.3323 | 49.4 ± 15.6 B | 30.0 ± 11.1 A | 38.6 ± 16.2 ab | 36.0 ± 13.6 a | 44.5 ± 19.0 b |
| Lb | <0.0001 | 0.0040 | 0.1082 | 5.2 ± 1.5 B | 2.9 ± 1.2 A | 3.7 ± 1.5 a | 3.9 ± 1.8 ab | 4.6 ± 2.0 b |
| Q. robur | S | F | S × F |
|---|---|---|---|
| Sb | <0.0001 | <0.0001 | 0.6355 |
| MRb | 0.9205 | 0.0610 | 0.7401 |
| FOLRb <1 | 0.0212 | 0.5548 | 0.9441 |
| FOLRb 1–5 | <0.0001 | <0.0001 | 0.1665 |
| FOLRb >5 | 0.8056 | 0.2704 | 0.6460 |
| Rb | 0.0454 | 0.0001 | 0.7101 |
| Sb + Rb | 0.0001 | <0.0001 | 0.9119 |
| Q. pubescens | |||
| Sb | <0.0001 | 0.0002 | 0.2060 |
| MRb | <0.0001 | 0.1275 | 0.2324 |
| FOLRb <1 | 0.0081 | 0.1637 | 0.0127 |
| FOLRb 1–5 | <0.0001 | 0.0075 | 0.1767 |
| FOLRb >5 | 0.8859 | 0.3703 | 0.4351 |
| Rb | <0.0001 | 0.3417 | 0.4148 |
| Sb + Rb | <0.0001 | 0.0438 | 0.4563 |
| Q. ilex | |||
| Sb | <0.0001 | <0.0001 | 0.1204 |
| MRb | <0.0001 | 0.0005 | 0.0230 |
| FOLRb <1 | 0.0059 | 0.5604 | 0.0674 |
| FOLRb 1–5 | <0.0001 | 0.0013 | 0.1812 |
| FOLRb >5 | - | - | - |
| Rb | <0.0001 | 0.0001 | 0.0389 |
| Sb + Rb | <0.0001 | <0.0001 | 0.0424 |
| Q. robur | Q. pubescens | Q. ilex | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| S | F | S × F | S | F | S × F | S | F | S × F | ||
| N | Leaves | 0.1398 | 0.8712 | 0.3971 | 0.2220 | 0.2340 | 0.2340 | <0.0001 | 0.0067 | 0.3531 |
| Shoot | 0.3956 | 0.1357 | 0.1760 | 0.0922 | 0.0681 | 0.0681 | <0.0001 | 0.0027 | 0.1779 | |
| Root | 0.0711 | 0.0002 | 0.9724 | 0.5779 | 0.6927 | 0.6927 | <0.0001 | 0.0017 | 0.2128 | |
| P | Leaves | 0.0036 | 0.0317 | 0.4204 | 0.0002 | 0.0001 | 0.0001 | 0.0020 | 0.3830 | 0.4956 |
| Shoot | 0.4027 | 0.0013 | 0.0439 | 0.3254 | 0.0001 | 0.0001 | 0.1794 | 0.0923 | 0.9658 | |
| Root | 0.0088 | <0.0001 | 0.0208 | 0.0034 | 0.0024 | 0.0024 | 0.7227 | 0.0381 | 0.5865 | |
| K | Leaves | 0.1448 | 0.5164 | 0.1323 | 0.0469 | 0.4150 | 0.4150 | 0.0751 | 0.7980 | 0.2119 |
| Shoot | 0.4515 | 0.7074 | 0.1073 | 0.2170 | 0.5737 | 0.5737 | 0.8051 | 0.0423 | 0.6514 | |
| Root | 0.4930 | 0.6484 | 0.1861 | 0.4115 | 0.3866 | 0.3866 | 0.0188 | 0.3926 | 0.1645 | |
| Q. robur | S | F | S × F | Pe | Co | St | P | K |
|---|---|---|---|---|---|---|---|---|
| Nitrate N | 0.0160 | 0.0004 | 0.5459 | 1.32 ± 0.49 B | 0.86 ± 0.66 A | 1.11 ± 0.36 ab | 1.65 ± 0.47 b | 0.52 ± 0.39 a |
| Ammonium N | 0.9211 | 0.0029 | 0.2031 | 1.02 ± 0.34 | 1.01 ± 0.54 | 0.92 ± 0.26 ab | 1.44 ± 0.37 b | 0.69 ± 0.29 a |
| P | 0.2960 | 0.9878 | 0.3515 | 0.26 ± 0.08 | 0.22 ± 0.06 | 0.24 ±0.06 | 0.23 ±0.05 | 0.24 ± 0.11 |
| K | 0.3836 | 0.0821 | 0.9781 | 0.89 ± 0.24 | 0.79 ± 0.25 | 0.66 ± 0.13 | 0.88 ± 0.26 | 0.98 ± 0.23 |
| Q. pubescens | ||||||||
| Nitrate N | 0.0612 | 0.0020 | 0.3632 | 1.22 ± 0.70 B | 0.83 ± 0.48 A | 0.84 ± 0.42 a | 1.64 ± 0.54 b | 0.61 ± 0.35 a |
| Ammonium N | 0.7522 | 0.0036 | 0.6327 | 0.96 ± 0.55 | 0.91 ± 0.37 | 0.81 ± 0.26 ab | 1.39 ± 0.38 b | 0.60 ± 0.28 a |
| P | 0.9340 | 0.0222 | 0.6351 | 0.19 ± 0.07 | 0.19 ± 0.04 | 0.18 ± 0.04 ab | 0.24 ± 0.05 b | 0.15 ± 0.04 a |
| K | 0.5296 | 0.1437 | 0.5819 | 0.73 ± 0.25 | 0.87 ± 0.64 | 0.50 ± 0.13 | 1.07 ± 0.70 | 0.83 ± 0.26 |
| Q. ilex | ||||||||
| Nitrate N | 0.7839 | 0.0212 | 0.4642 | 0.65 ± 0.51 | 0.71 ± 0.51 | 0.77 ± 0.35 ab | 1.02 ± 0.55 b | 0.25 ± 0.20 a |
| Ammonium N | 0.0299 | 0.0260 | 0.3370 | 0.44 ± 0.36 A | 0.83 ± 0.46 B | 0.65 ± 0.39 ab | 0.93 ± 0.53 b | 0.32 ± 0.14 a |
| P | 0.0824 | 0.3603 | 0.8507 | 0.15 ± 0.06 | 0.21 ± 0.08 | 0.17 ± 0.05 | 0.22 ± 0.11 | 0.16 ± 0.05 |
| K | 0.0295 | 0.0005 | 0.7127 | 0.68 ± 0.34 A | 0.90 ± 0.28 B | 0.59 ± 0.23 a | 0.65 ± 0.25 a | 1.14 ± 0.14 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mariotti, B.; Martini, S.; Raddi, S.; Tani, A.; Jacobs, D.F.; Oliet, J.A.; Maltoni, A. Coconut Coir as a Sustainable Nursery Growing Media for Seedling Production of the Ecologically Diverse Quercus Species. Forests 2020, 11, 522. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050522
Mariotti B, Martini S, Raddi S, Tani A, Jacobs DF, Oliet JA, Maltoni A. Coconut Coir as a Sustainable Nursery Growing Media for Seedling Production of the Ecologically Diverse Quercus Species. Forests. 2020; 11(5):522. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050522
Chicago/Turabian StyleMariotti, Barbara, Sofia Martini, Sabrina Raddi, Andrea Tani, Douglass F. Jacobs, Juan A. Oliet, and Alberto Maltoni. 2020. "Coconut Coir as a Sustainable Nursery Growing Media for Seedling Production of the Ecologically Diverse Quercus Species" Forests 11, no. 5: 522. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050522