The Strong and the Stronger: The Effects of Increasing Ozone and Nitrogen Dioxide Concentrations in Pollen of Different Forest Species

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pollen Sampling
2.2. Exposure Experiments of Pollen to Pollutant Gasses
2.3. Pollen Viability
2.4. Pollen Protein Extraction and Quantification
2.5. Pollen Oxidative Stress Analysis
2.5.1. Detection of Reactive Oxygen Species (ROS)
2.5.2. NADPH Oxidase Enzymatic Activity
2.5.3. Western Blotting Analysis of Superoxide Dismutase (SOD)
2.6. Raman Microspectroscopy
2.7. Statistical Analysis
3. Results
3.1. Pollen Viability
3.2. Pollen Total Soluble Protein Content
3.3. Pollen Oxidative Stress
3.3.1. Detection of Reactive Oxygen Species (ROS)
3.3.2. NADPH Oxidase Enzymatic Activity
3.3.3. Superoxide Dismutase (SOD) Expression
3.4. Raman Microspectroscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dobbs, C.; Escobedo, F.J.; Zipperer, W.C. A framework for developing urban forest ecosystem services and goods indicators. Landsc. Urban Plan. 2011, 99, 196–206. [Google Scholar] [CrossRef]
- Ghazoul, J. Forests: A Very Short Introduction; Oxford University Press: Oxford, UK, 2015. [Google Scholar]
- Rajoo, K.S.; Karam, D.S.; Abdullah, M.Z. The physiological and psychosocial effects of forest therapy: A systematic review. Urban For. Urban Green. 2020, 54, 126744. [Google Scholar] [CrossRef]
- Ostoić, S.K.; Salbitano, F.; Borelli, S.; Verlič, A. Urban forest research in the Mediterranean: A systematic review. Urban For. Urban Green. 2018, 31, 185–196. [Google Scholar] [CrossRef]
- Vujcic, M.; Tomicevic-Dubljevic, J.; Živojinović, I.; Tošković, O. Connection between urban green areas and visitors’ physical and mental well-being. Urban For. Urban Green. 2019, 40, 299–307. [Google Scholar] [CrossRef]
- Berland, A.; Shiflett, S.A.; Shuster, W.D.; Garmestani, A.; Goddard, H.; Herrmann, D.L.; Hopton, M.E. The role of trees in urban stormwater management. Landsc. Urban Plan. 2017, 162, 167–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Dong, L.; Yu, C.; Zhang, Y.; Cheng, B. Can forest city construction affect urban air quality? The evidence from the Beijing-Tianjin-Hebei urban agglomeration of China. J. Clean. Prod. 2020, 264, 121607. [Google Scholar] [CrossRef]
- Nowak, D.J.; Hirabayashi, S.; Bodine, A.; Greenfield, E. Tree and forest effects on air quality and human health in the United States. Environ. Pollut. 2014, 193, 119–129. [Google Scholar] [CrossRef] [Green Version]
- Nowak, D.J.; Crane, D.E. Carbon storage and sequestration by urban trees in the USA. Environ. Pollut. 2002, 116, 381–389. [Google Scholar] [CrossRef]
- Douglas, A.N.; Irga, P.J.; Torpy, F.R. Determining broad scale associations between air pollutants and urban forestry: A novel multifaceted methodological approach. Environ. Pollut. 2019, 247, 474–481. [Google Scholar] [CrossRef]
- Shiraiwa, M.; Selzle, K.; Pöschl, U. Hazardous components and health effects of atmospheric aerosol particles: Reactive oxygen species, soot, polycyclic aromatic compounds and allergenic proteins. Free. Radic. Res. 2012, 46, 927–939. [Google Scholar] [CrossRef]
- Smith, W.H. Forests, Crops, and Air Quality. BioScience 1990, 40, 781–782. [Google Scholar] [CrossRef]
- Wielgosinski, G. Pollutant Formation in Combustion Processes. In Advances in Chemical Engineering; Nawaz, Z., Naveed, S., Eds.; IntechOpen: London, UK, 2012; p. 30. [Google Scholar]
- Najjar, Y.S.H. Gaseous Pollutants Formation and Their Harmful Effects on Health and Environment. Innov. Energy Policies 2011, 1, 1–9. [Google Scholar] [CrossRef]
- Mijnsbrugge, K.V.; Turcsán, A.; Moreels, S.; Van Goethem, M.; Meeus, S.; Van Der Aa, B. Does Drought Stress on Seedlings Have Longer Term Effects on Sapling Phenology, Reshooting, Growth and Plant Architecture in Quercus robur, Q. petraea and Their Morphological Intermediates? Forests 2019, 10, 1012. [Google Scholar] [CrossRef] [Green Version]
- Sgrigna, G.; Baldacchini, C.; Dreveck, S.; Cheng, Z.; Calfapietra, C. Relationships between air particulate matter capture efficiency and leaf traits in twelve tree species from an Italian urban-industrial environment. Sci. Total. Environ. 2020, 718, 137310. [Google Scholar] [CrossRef] [PubMed]
- Hoshika, Y.; Carrari, E.; Mariotti, B.; Martini, S.; De Marco, A.; Sicard, P.; Paoletti, E. Flux-Based Ozone Risk Assessment for a Plant Injury Index (PII) in Three European Cool-Temperate Deciduous Tree Species. Forests 2020, 11, 82. [Google Scholar] [CrossRef] [Green Version]
- Lucas, J.A.; Gutierrez-Albanchez, E.; Alfaya, T.; Brito, F.F.; Gutierrez-Mañero, F.J. Oxidative stress in ryegrass growing under different air pollution levels and its likely effects on pollen allergenicity. Plant Physiol. Biochem. 2019, 135, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Plant Physiology and Biochemistry Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Kinose, Y.; Matsumura, H.; Izuta, T. Evaluation of O3 Effects on Cumulative Photosynthetic CO2 Uptake in Seedlings of Four Japanese Deciduous Broad-Leaved Forest Tree Species Based on Stomatal O3 Uptake. Forests 2019, 10, 556. [Google Scholar] [CrossRef] [Green Version]
- Leghari, S.K.; Saeed, S.; Asrar, M.; Ahmed, A.; Tariq, I.; Marri, A.A.; Sadiq, N.; Baloch, A.; Latif, A.; Shawani, N.A. Response of sweet cherry (Prunus avium L.) pollen grains to vehicular exhaust pollution at quetta, balochistan, pakistan. Appl. Ecol. Environ. Res. 2018, 16, 4387–4399. [Google Scholar] [CrossRef]
- Fröhlich-Nowoisky, J.; Kampf, C.J.; Weber, B.; Huffman, J.A.; Pöhlker, C.; Andreae, M.O.; Lang-Yona, N.; Burrows, S.M.; Gunthe, S.S.; Elbert, W.; et al. Bioaerosols in the Earth system: Climate, health, and ecosystem interactions. Atmos. Res. 2016, 182, 346–376. [Google Scholar] [CrossRef] [Green Version]
- Colls, J. Air Pollution, 2nd ed.; Spon Press: London, UK, 2002. [Google Scholar]
- Duque, L.; Guimarães, F.M.G.; Ribeiro, H.; Sousa, R.; Abreu, I. Elemental characterization of the airborne pollen surface using Electron Probe Microanalysis (EPMA). Atmos. Environ. 2013, 75, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Oduber, F.; Calvo, A.; Blanco-Alegre, C.; Castro, A.; Vega-Maray, A.; Valencia-Barrera, R.; Fernández-González, D.; Fraile, R. Links between recent trends in airborne pollen concentration, meteorological parameters and air pollutants. Agric. For. Meteorol. 2019, 264, 16–26. [Google Scholar] [CrossRef]
- Sénéchal, H.; Visez, N.; Charpin, D.; Shahali, Y.; Peltre, G.; Biolley, J.-P.; Lhuissier, F.; Couderc, R.; Yamada, O.; Malrat-Domenge, A.; et al. A Review of the Effects of Major Atmospheric Pollutants on Pollen Grains, Pollen Content, and Allergenicity. Sci. World J. 2015, 2015, 1–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, H.; Costa, C.; Abreu, I.; Da Silva, J.C.E. Effect of O3 and NO2 atmospheric pollutants on Platanus x acerifolia pollen: Immunochemical and spectroscopic analysis. Sci. Total. Environ. 2017, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Pasqualini, S.; Tedeschini, E.; Frenguelli, G.; Wopfner, N.; Ferreira, F.; D’Amato, G.; Ederli, L. Ozone affects pollen viability and NAD(P)H oxidase release from Ambrosia artemisiifolia pollen. Environ. Pollut. 2011, 159, 2823–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sousa, R.; Duque, L.; Duarte, A.J.; Gomes, C.R.; Ribeiro, H.; Cruz, A.; Da Silva, J.C.G.E.; Abreu, I. In Vitro Exposure of Acer negundo Pollen to Atmospheric Levels of SO2 and NO2: Effects on Allergenicity and Germination. Environ. Sci. Technol. 2012, 46, 2406–2412. [Google Scholar] [CrossRef] [Green Version]
- Lu, S.; Ren, J.; Hao, X.; Liu, D.; Zhang, R.; Wu, M.; Yi, F.; Lin, J.; Shinich, Y.; Wang, Q. Characterization of protein expression of Platanus pollen following exposure to gaseous pollutants and vehicle exhaust particles. Aerobiologia 2014, 30, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Ribeiro, H.; Oliveira, M.; Cruz, A.; Da Silva, J.C.G.E. Effects of CO2 on Acer negundo pollen fertility, protein content, allergenic properties, and carbohydrates. Environ. Sci. Pollut. Res. 2014, 22, 6904–6911. [Google Scholar] [CrossRef]
- Cuinica, L.G.; Abreu, I.; Gomes, C.R.; Da Silva, J.C.G.E. Exposure of Betula pendula Roth pollen to atmospheric pollutants CO, O3 and SO2. Grana 2013, 52, 299–304. [Google Scholar] [CrossRef]
- Chassard, G.; Choël, M.; Gosselin, S.; Vorng, H.; Petitprez, D.; Shahali, Y.; Tsicopoulos, A.; Visez, N. Kinetic of NO2 uptake by Phleum pratense pollen: Chemical and allergenic implications. Environ. Pollut. 2015, 196, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Farah, J.; Choël, M.; De Nadaï, P.; Gosselin, S.; Petitprez, D.; Baroudi, M.; Visez, N. Extractable lipids from Phleum pratense pollen grains and their modifications by ozone exposure. Aerobiologia 2019, 36, 171–182. [Google Scholar] [CrossRef]
- Zhu, C.; Farah, J.; Choël, M.; Gosselin, S.; Baroudi, M.; Petitprez, D.; Visez, N. Uptake of ozone and modification of lipids in Betula Pendula pollen. Environ. Pollut. 2018, 242, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Heslop-Harrison, J.; Heslop-Harrison, Y.; Shivanna, K.R. The evaluation of pollen quality, and a further appraisal of the fluorochromatic (FCR) test procedure. Theor. Appl. Genet. 1984, 67, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Zafra, A.; Castro, A.J.; Alché, J.D.D. Identification of novel superoxide dismutase isoenzymes in the olive (Olea europaea L.) pollen. BMC Plant Biol. 2018, 18, 114. [Google Scholar] [CrossRef] [PubMed]
- Luria, G.; Rutley, N.; Lazar, I.; Harper, J.F.; Miller, G. Direct analysis of pollen fitness by flow cytometry: Implications for pollen response to stress. Plant J. 2019, 98, 942–952. [Google Scholar] [CrossRef]
- Marino, D.; Dunand, C.; Puppo, A.; Pauly, N. A burst of plant NADPH oxidases. Trends Plant Sci. 2012, 17, 9–15. [Google Scholar] [CrossRef]
- Ren, R.; Li, Z.; Zhang, L.; Zhou, H.; Jiang, X.; Liu, Y. Enzymatic and nonenzymatic antioxidant systems impact the via-bility of cryopreserved Paeonia suffruticosa pollen. Plant Cell Tissue Organ Cult. 2020. [Google Scholar] [CrossRef]
- Uka, U.N.; Hogarh, J.; Belford, E.J.D. Morpho-Anatomical and Biochemical Responses of Plants to Air Pollution. Int. J. Mod. Bot. 2017, 7, 1–11. [Google Scholar]
- Rao, M.; Dubey, P. Biochemical aspects (antioxidants) for development of tolerance in plants growing at different low levels of ambient air pollutants. Environ. Pollut. 1990, 64, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Diehn, S.; Zimmermann, B.; Tafintseva, V.; Seifert, S.; Bağcıoğlu, M.; Ohlson, M.; Weidner, S.; Fjellheim, S.; Kohler, A.; Kneipp, J. Combining Chemical Information from Grass Pollen in Multimodal Characterization. Front. Plant Sci. 2020, 10, 1788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, A.; Ribeiro, H.; Fernández-González, M.; Aira, M.; Abreu, I. Pollen Raman spectra database: Application to the identification of airborne pollen. Talanta 2014, 119, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Ivleva, N.P.; Niessner, R.; Panne, U. Characterization and discrimination of pollen by Raman microscopy. Anal. Bioanal. Chem. 2005, 381, 261–267. [Google Scholar] [CrossRef] [PubMed]
- Kanter, U.; Heller, W.; Durner, J.; Winkler, J.B.; Engel, M.; Behrendt, H.; Holzinger, A.; Braun, P.; Hauser, M.; Ferreira, F.; et al. Molecular and Immunological Characterization of Ragweed (Ambrosia artemisiifolia L.) Pollen after Exposure of the Plants to Elevated Ozone over a Whole Growing Season. PLoS ONE 2013, 8, e61518. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Elkelsih, A.; Durner, J.; Lindermayr, C.; Winkler, J.B.; Ruëff, F.; Behrendt, H.; Traidl-Hoffmann, C.; Holzinger, A.; Kofler, W.; et al. Common ragweed (Ambrosia artemisiifolia L.): Allergenicity and molecular characterization of pollen after plant exposure to elevated NO2. Plant Cell Environ. 2015, 39, 147–164. [Google Scholar] [CrossRef] [PubMed]
- Møller, I.M.; Jensen, P.E.; Hansson, A. Oxidative Modifications to Cellular Components in Plants. Annu. Rev. Plant Biol. 2007, 58, 459–481. [Google Scholar] [CrossRef] [Green Version]
- Gupta, M.K.; Uhm, S.J.; Lee, H.T. Effect of vitrification and beta-mercaptoethanol on reactive oxygen species activity and in vitro development of oocytes vitrified before or after in vitro fertilization. Fertil. Steril. 2010, 93, 2602–2607. [Google Scholar] [CrossRef]
- Depciuch, J.; Kasprzyk, I.; Sadik, O.; Parlińska-Wojtan, M. FTIR analysis of molecular composition changes in hazel pollen from unpolluted and urbanized areas. Aerobiologia 2017, 33, 1–12. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
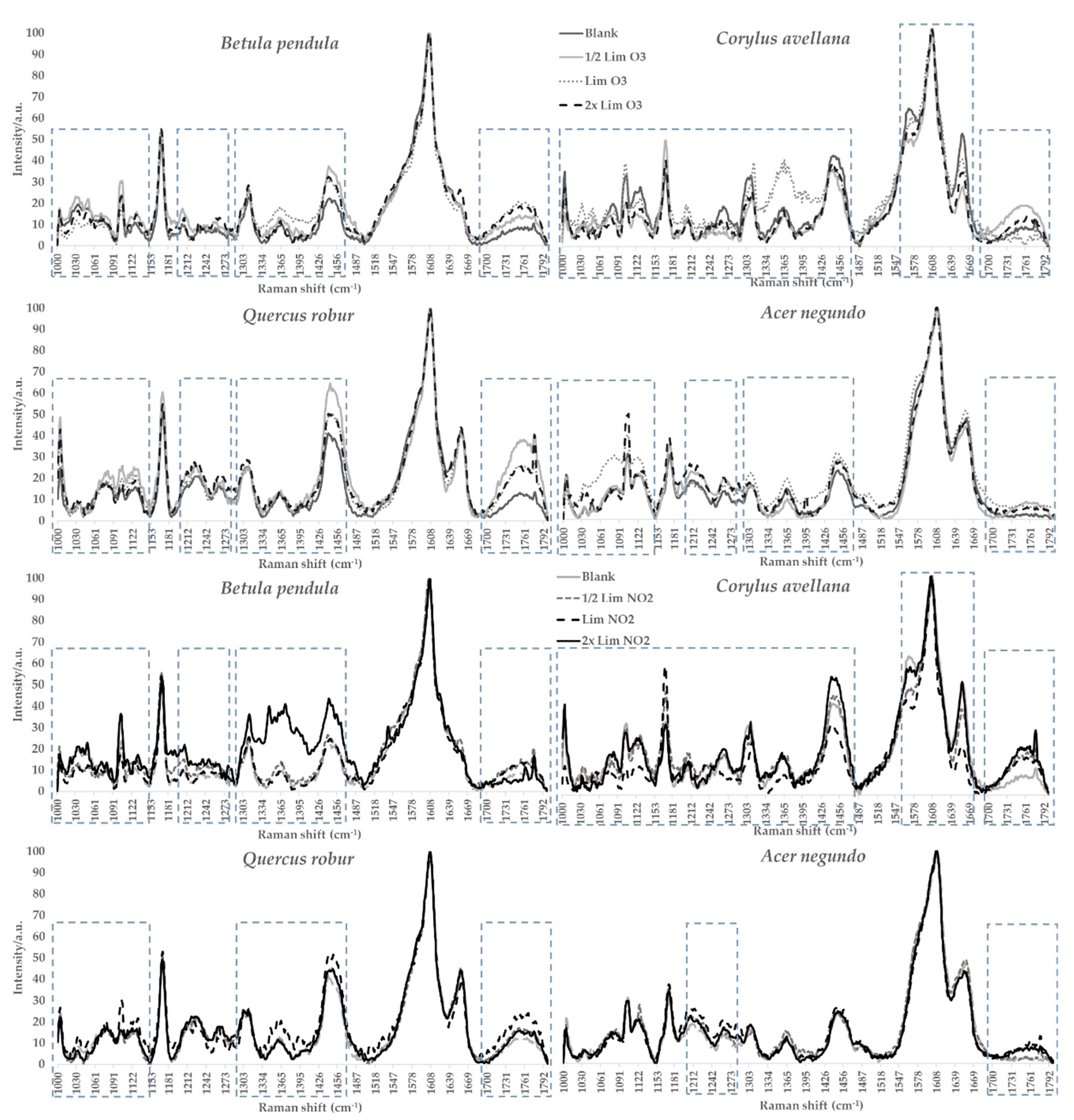{kind=link}
| Blank | Ozone | NO2 | |||||
|---|---|---|---|---|---|---|---|
| ½ Lim | Lim | 2 × Lim | ½ Lim | Lim | 2 × Lim | ||
| Acer negundo | |||||||
| Δ T (°C) | 24.5 ± 0.391 | 25.4 ± 1.554 | 25.2 ± 0.236 | 23.7 ± 0.905 | 25.3 ± 0.424 | 24.6 ± 0.645 | 24.2 ± 0.259 |
| Δ RH (%) | 59.1 ± 0.654 | 61.2 ± 1.641 | 61.9 ± 0.968 | 61.4 ± 1.274 | 62.3 ± 0.241 | 61.2 ± 1.238 | 60.2 ± 0.427 |
| (Gas) (ppm) | --- | 0.031 ± 0.002 | 0.064 ± 0.014 | 0.125 ± 0.016 | 0.051 ± 0.010 | 0.113 ± 0.037 | 0.197 ± 0.039 |
| Betula pendula | |||||||
| Δ T (°C) | 25.6 ± 0.880 | 25.6 ± 0.975 | 21.5 ±1.182 | 24.6 ± 1.080 | 28.4 ± 0.706 | 25.6 ± 0.680 | 25.4 ±0.866 |
| Δ RH (%) | 59.6 ± 0.991 | 60.7 ± 0.932 | 59.1 ± 1.398 | 60.9 ± 0.944 | 58.2 ± 1.098 | 60.3 ± 1.310 | 59.8 ± 1.285 |
| (Gas) (ppm) | --- | 0.030 ± 0.001 | 0.061 ± 0.015 | 0.120 ± 0.030 | 0.055 ± 0.021 | 0.104 ± 0.017 | 0.195 ± 0.036 |
| Corylus avellana | |||||||
| Δ T (°C) | 24.4 ± 0.643 | 25.3 ± 0.899 | 24.8 ±0.714 | 24.6 ± 0.628 | 25.0 ± 0.868 | 25.3 ± 0.645 | 24.0 ± 0.882 |
| Δ RH (%) | 59.9 ± 0.925 | 56.2 ± 1.802 | 60.9 ± 2.010 | 59.9 ± 1.000 | 57.4 ± 0.377 | 57.4 ± 1.527 | 63.2 ± 0.575 |
| (Gas) (ppm) | --- | 0.030 ± 0.005 | 0.059 ± 0.013 | 0.119 ± 0.090 | 0.057 ± 0.022 | 0.105 ± 0.044 | 0.203 ± 0.060 |
| Quercus robur | |||||||
| Δ T (°C) | 24.0 ± 0.979 | 24.9 ± 1.529 | 25.5 ± 1.103 | 25.0 ± 1.577 | 24.6 ± 0.923 | 24.6 ± 0.903 | 24.9 ± 0.557 |
| Δ RH (%) | 60.7 ± 2.218 | 61.4 ± 1.407 | 58.5 ±1.568 | 62.1 ± 1.670 | 59.0 ± 1.189 | 61.2 ± 1.572 | 61.0 ± 1.157 |
| (Gas) (ppm) | --- | 0.029 ± 0.010 | 0.060 ± 0.017 | 0.120 ± 0.015 | 0.059 ± 0.018 | 0.112 ± 0.027 | 0.220 ± 0.046 |
| Ozone | NO2 | |||||
|---|---|---|---|---|---|---|
| ½ Lim | Lim | 2 × Lim | ½ Lim | Lim | 2 × Lim | |
| Viability | ||||||
| Acer negundo | ≈ | ≈ | ≈ | ≈ | 7%↓ | 9%↓ |
| Betula pendula | ≈ | 9%↓ | 8%↓ | ≈ | 8%↓ | 17%↓ |
| Corylus avellana | 12% ↓ | 21%↓ | 13%↓ | 5%↓ | 13%↓ | 16%↓ |
| Quercus robur | 4% ↓ | 5%↓ | 7%↓ | 3%↓ | 7%↓ | 12%↓ |
| Total Soluble Protein content | ||||||
| Acer negundo | 13%↓ | 18%↓ | 22%↓ | 19%↓ | 6%↓ | 19%↓ |
| Betula pendula | ≈ | 22%↑ | ≈ | ≈ | ≈ | ≈ |
| Corylus avellana | 20%↓ | 10%↑ | ≈ | ≈ | 16%↑ | 28%↑ |
| Quercus robur | ≈ | 19%↓ | 13%↓ | 29%↓ | ≈ | 53%↓ |
| ROS content | ||||||
| Acer negundo | 12%↑ | 13%↑ | 12%↑ | 12%↑ | 12%↑ | 23%↑ |
| Betula pendula | ≈ | 6%↑ | 3%↑ | ≈ | 2%↑ | 6%↑ |
| Corylus avellana | ≈ | 12%↑ | ≈ | ≈ | ≈ | 7%↑ |
| Quercus robur | 8%↑ | 12%↑ | 11%↑ | ≈ | 12%↑ | 10%↑ |
| NADPH oxidase enzymatic activity | ||||||
| Acer negundo | ≈ | ≈ | ≈ | ≈ | ≈ | 28%↑ |
| Betula pendula | 15%↑ | ≈ | ≈ | ≈ | 15%↑ | 14%↑ |
| Corylus avellana | ≈ | ≈ | ≈ | ≈ | ≈ | ≈ |
| Quercus robur | 21%↑ | ≈ | ≈ | ≈ | ≈ | 20%↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pereira, S.; Fernández-González, M.; Guedes, A.; Abreu, I.; Ribeiro, H. The Strong and the Stronger: The Effects of Increasing Ozone and Nitrogen Dioxide Concentrations in Pollen of Different Forest Species. Forests 2021, 12, 88. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010088
Pereira S, Fernández-González M, Guedes A, Abreu I, Ribeiro H. The Strong and the Stronger: The Effects of Increasing Ozone and Nitrogen Dioxide Concentrations in Pollen of Different Forest Species. Forests. 2021; 12(1):88. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010088
Chicago/Turabian StylePereira, Sónia, Maria Fernández-González, Alexandra Guedes, Ilda Abreu, and Helena Ribeiro. 2021. "The Strong and the Stronger: The Effects of Increasing Ozone and Nitrogen Dioxide Concentrations in Pollen of Different Forest Species" Forests 12, no. 1: 88. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010088