
Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy



1
Biotechnical Faculty, University of Ljubljana, Jamnikarjeva 101, 1000 Ljubljana, Slovenia
2
Department of Agricultural Sciences, University of Naples Federico II, Via Università 100, I-80055 Portici, Naples, Italy
*
Author to whom correspondence should be addressed.
Forests 2021, 12(2), 188; https://0-doi-org.brum.beds.ac.uk/10.3390/f12020188
Submission received: 19 January 2021
/
Revised: 1 February 2021
/
Accepted: 3 February 2021
/
Published: 7 February 2021
(This article belongs to the Special Issue Wood Formation and Environmental Constraints: Multiscale Approach)
Abstract
:Quercus ilex L. dieback has been recently reported at numerous Mediterranean sites. Wood and phloem formation dynamics and tree-ring series of anatomical traits can be used to evaluate growth conditions of trees. We monitored cambial activity in Q. ilex trees growing at a site in southern Italy in order to assess how xylem and phloem production are affected by harsh seasonal climatic variation during a dry year. We followed xylogenesis by counting the number of cambial cells and detecting the occurrence of post-cambial cells throughout the year. As phloem did not show clear growth rings and boundaries between them, we followed the development of phloem fibres—their morphological traits during development and the distance from the cambium served as a reference point to evaluate the phloem production during the year. We detected a multimodal pattern in cambial activity, with wood production in three periods of the year and consequent formation of intra-annual density fluctuations (IADFs). The lowest production of xylem cells was observed in the dry late spring and summer period (likely due to the low water availability), while the highest occurred in autumn (the wettest period). Although we could not differentiate between early and late phloem, the analysis of phloem traits was useful to follow the dynamics of phloem production, which is generally difficult in Mediterranean tree species. We found cambial production of phloem throughout the year, even in the periods without xylem production. The results showed that if tree growth was constrained by environmental limitations, the ratio between xylem to phloem cells decreased and, in the most severely affected trees, more cells were formed preferentially in the phloem compared to xylem. We also briefly report the way in which to solve technical problems with tissue preparation due to extreme hardness and to the peculiar structure of Q. ilex wood and outer bark.

1. Introduction
Ongoing climate change is having a profound impact on forest growth and productivity, especially in the Mediterranean region where trees are suffering due to the general increase in temperature and in the frequency of extreme events [1], as severe and prolonged droughts and heat waves often are followed by insect outbreaks. Such events, together with extremes such as fires, are causing dieback phenomena affecting Mediterranean forest in its structure and services [1,2]. Here, the currently dominant species are being increasingly replaced by shrub and invasive species which are better acclimated and/or adapted to the changing climate [2]. For example, in the case of forests in southern Italy, invasive species such as Robinia pseudoacacia L. are spreading, competing with endemic species such as holm oak (Quercus ilex L.) [3].
Q. ilex is an evergreen species, shade-tolerant, with deep roots, which can maintain high stomata conductivity during dry periods [4]. Although it is considered to be a drought-resistant Mediterranean tree species [5,6,7], under extreme water stress conditions, Q. ilex can suffer high losses of xylem conductivity, being also susceptible to embolisms [8,9,10]. Recently, Q. ilex dieback has been reported at numerous locations in its natural sites, suggesting that weather extremes, such as extremely hot and dry summer conditions, might be among the reasons for the decline [2,11,12,13,14]. Q. ilex shows higher vulnerability to drought compared to other co-occurring species, such as Phyllirea latifolia L. and Erica arborea L. [15,16,17,18]. In the future, drier conditions could lead to higher mortality of Q. ilex, considered a key element of Mediterranean forests, as it plays a very important role in the functioning of these ecosystems [19]. The ability of this species to suspend vegetative growth during periods of drought can lead to irregular cambial production, which is probably responsible for the formation of more than one annual ring within a calendar year.
Dendrochronology is widely used to establish health condition of trees. Tree-ring width trends and the relations between anatomical traits and climatic factors can reveal the duration and nature of decline [20,21]. In Q. ilex, the classical dendrochronological approach often cannot be directly applied since it is difficult to recognise the boundaries between tree-rings. As opposed to most Quercus species which are ring porous, Q. ilex wood shows diffuse porosity, with radially arranged large vessels of more or less uniform size. It often forms intra-annual density fluctuations (IADFs) which additionally hamper the recognition of tree rings [11,22,23,24,25,26,27,28,29]. Indeed, recent studies showed that if the weather conditions are favourable, most Mediterranean species may continue to grow even during winter; as a consequence, if the cambium does not interrupt wood production at the end of the calendar year, tree rings cannot be exactly ascribed to a calendar year [30,31]. On the other side, hot and dry summer conditions possibly interrupt cambial production leading to IADFs [32,33,34,35]. IADFs in Q. ilex are described as an “extra growth band” of wood at the end of the true ring, with smaller vessels than in true earlywood, and often with a higher abundance of fibres and banded axial parenchyma [25,29]. The occurrence of IADFs in Q. ilex wood has been associated with the alternation of drought in summer, followed by wet periods [22,25,36]. For Q. ilex growing in southern Italy, it has been recently reported that the IADF frequency is associated with higher efficiency of water use regardless of the different xylem growth [37], and that in a scenario of increasing temperature IADF occurrence in Q. ilex would be decreased compared to other species [29]. In Q. ilex growing in Spain, a relationship has been reported between the width of IADFs and the amount of precipitation in the second part of the growing season, suggesting that IADFs in this species can be indicators of phenotypic plasticity [25]. Wood formation studies are of great help to obtain information on when and how a tree-ring is formed, to ascribe the IADFs to the period of their formation and to exactly define tree rings [23]. Moreover, thanks to the recently acknowledged relevance of the xylogenesis studies for dendroecology, wood anatomy and the study of intra-annual dynamics of wood formation have become more and more relevant, especially over the last two decades. Research on cambial activity and xylem formation has become an invaluable piece in the mosaic of multidisciplinary and multi-scale studies to understand the response of trees and forests to climate change [23,38,39,40,41,42,43].
Since the vascular cambium is a bi-facial meristem which produces xylem and phloem, it is crucial to consider not only wood formation but also the seasonal dynamics of phloem formation to show the entire process of cambial cell productivity during the year or growing season [44]. Xylem and phloem formation processes are not fully synchronous [39,44] and show different dynamics, as internal and external influences have different effects on the mechanisms of their formation. The structure of the non-collapsed phloem and the dynamics of its formation can help assess tree vitality in temperate species where early and late phloem can be defined in phloem rings which are clearly delineated by boundaries [44,45,46]. In Mediterranean tree species, phloem annual rings cannot be recognised, and therefore it is difficult to study phloemogenesis. Phloem annual rings are not clearly demarcated and do not show clear early and late phloem [47,48,49]. As we lack reference points in phloem anatomy, it is usually not possible to define how much phloem has been formed between two tissue samplings. Recently, it was shown that quantitative anatomy used to study phloem of Pinus pinea L. and Pinus halepensis Mill. from drought-prone environments helps obtaining information on phloem formation during the growth season [48]. In this case, quantitative anatomy was used to measure the variations in the size of phloem sieve cells in non-collapsed phloem in the radial direction, the width of non-collapsed phloem, and the trend in the number of axial parenchyma cells throughout the non-collapsed phloem. This methodology proved to be suitable to obtain information about intra-annual variations of phloem traits in Mediterranean conifers. To the best of our knowledge, in Mediterranean hardwoods, the dynamics of phloem formation has not been investigated yet. It is known that the phloem of the genus Quercus contains groups of phloem fibres which in temperate Quercus species demarcate the boundary between early and late phloem [50,51]. If Mediterranean oaks also contain groups of fibres, they could serve as reference points when studying the dynamics of phloem formation. To the best of our knowledge, xylogenesis, phloemogenesis, and timing of IADF formation in wood of Q. ilex have not been monitored yet.
In this study, we monitored the cambium production in Q. ilex growing in southern Italy in the old-growth forest of “Parco Gussone”, which is part of the Royal Palace of Portici (Naples, Italy), during the particularly dry year 2017. We aimed to (a) assess the cambial activity and analyse the relations with the current climatic parameters, (b) assess whether annual xylem and phloem rings with clear boundaries were formed during the year, (c) detect the possible occurrence of IADFs in the xylem, (d) use phloem fibres as an indicator of the seasonal variation in phloem genesis, and (e) assess whether the processes of xylem and phloem formation respond in the same way to the intra-seasonal variations in climate. Our hypothesises were (1) the cambial activity would stop in summer and winter, leading to the formation of IADFs in the xylem, and (b) the phloem would not form a clear annual ring. We also discuss the limitations of the methodology and possible bias due to the hardness of wood and the fragility of the cambial zone.
2. Materials and Methods
2.1. Study Site
The study was conducted in a Quercus ilex L. forest located at “Parco Gussone-Bosco Superiore” (40°48′44.1972″, 14°20′50.7408″, 50 m above sea level, m a.s.l.), surrounding the Royal Palace of Portici (Naples), in southern Italy in the year 2017. The establishment of the Parco Gussone started in 1738 when Q. ilex seedlings from Sicily were used for the afforestation of the area. The forest of Parco Gussone extends over 16.4 ha and represents a real pool of floristic biodiversity in the metropolitan area of Naples [52], where forest dynamics and functional processes, such as natural regeneration and nutrient cycling, resemble those of a natural forest ecosystem [53].
The site is an evergreen Q. ilex woodland with the scattered occurrence of other deciduous species such as Fraxinus ornus L., Quercus pubescens Willd., Ulmus campestris L., Celtis australis L., and Mediterranean conifers [54]. The undergrowth is rich in shrub and climbing species [52]. Some of the older Q. ilex trees show phytosanitary problems due to inoculation with fungi with occurrence of fruiting bodies and hyphae of basidiomycetes fungi [52].
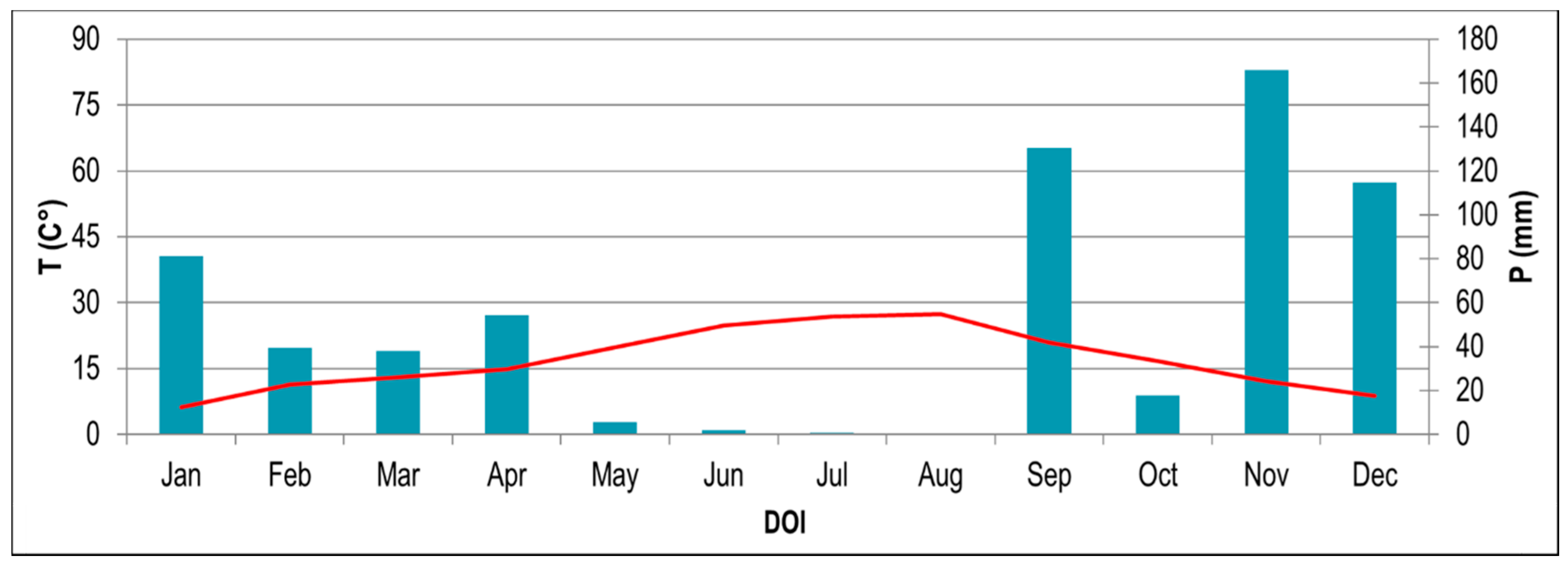
The climate is the Mediterranean humid-type, with a cumulative mean annual rainfall of ≈1000 mm, which is irregularly distributed. In 2017, the annual mean temperature was 16.98 °C. The hottest period occurred between June and August (monthly average mean temperature of 27.37 °C) and the coldest month was January (monthly average mean temperature of 8.74 °C) (Figure 1). The cumulative annual precipitation in 2017 was 650 mm; the wettest month was November, with cumulative monthly precipitation of 165.8 mm, while the lowest value was reached in August (cumulative monthly precipitation of 0 mm). Data are from the Sant’ Anastasia meteorological station, 40°53′25.61″ N, 14°21′28.02″ E, 25 m a.s.l, 10 km away from the sampling site.
2.2. Sampling, Microscopy, and Quantitative Wood Anatomy
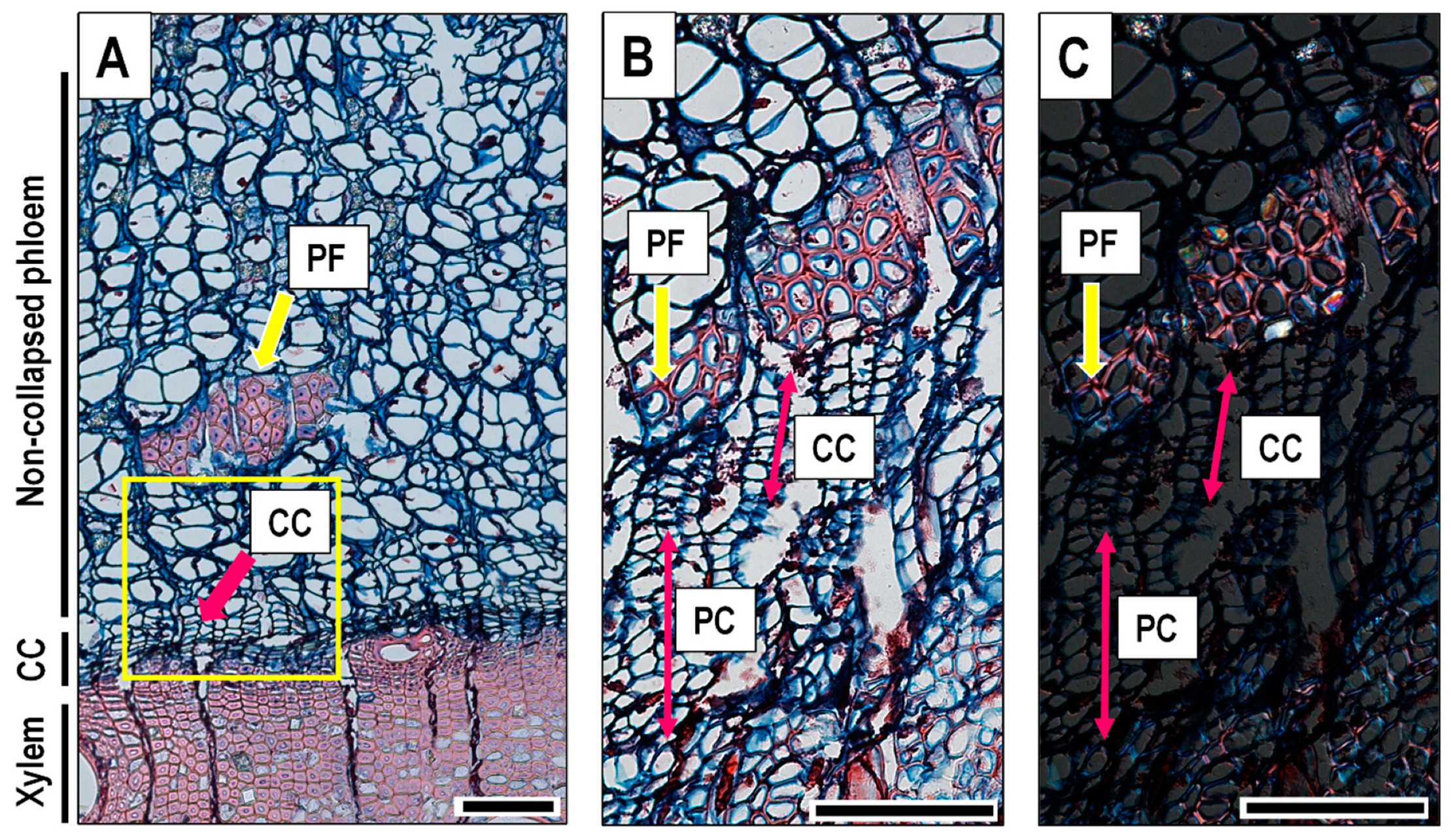
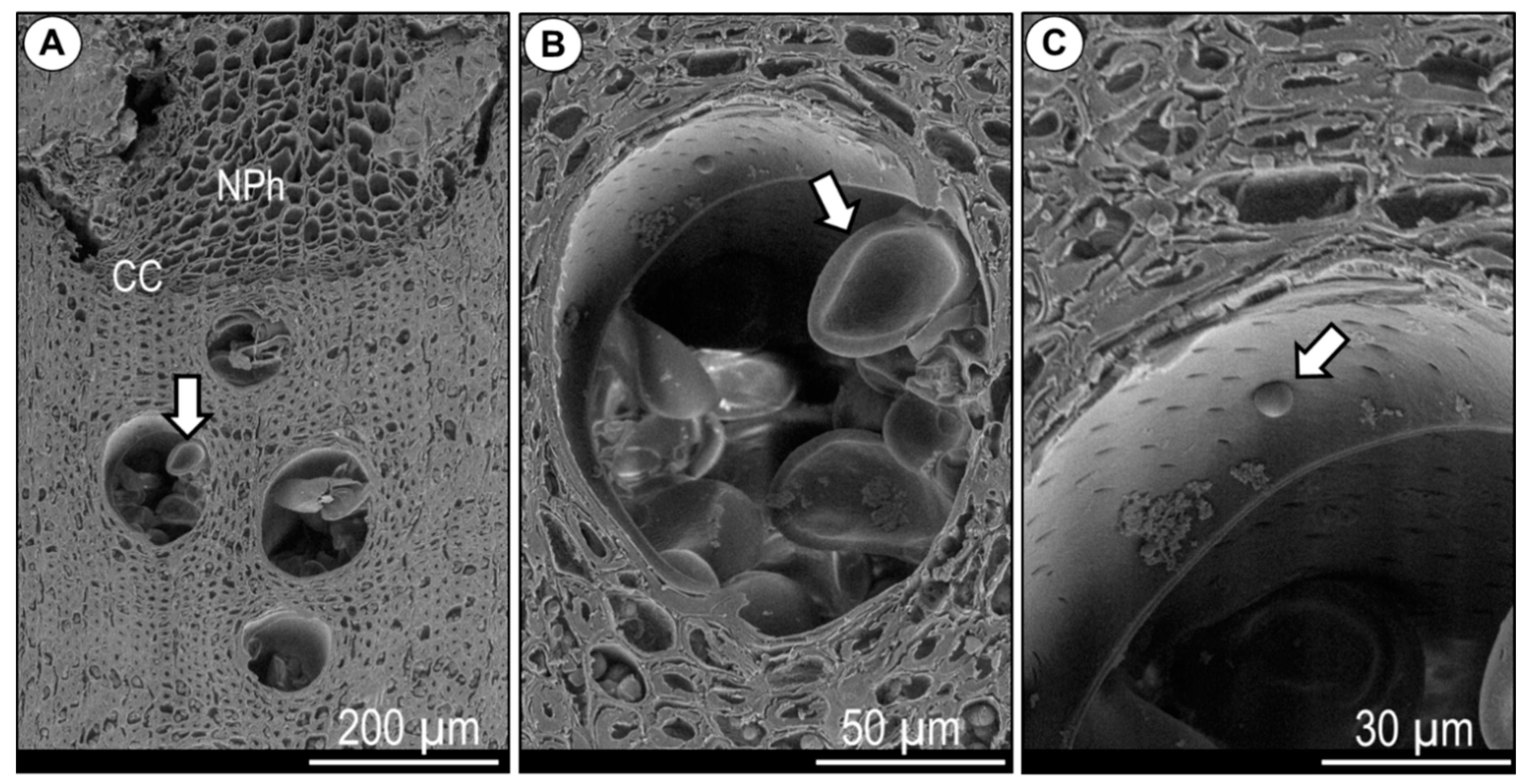
Microcores (1.8 mm in diameter) were collected every 2 weeks using the Trephor tool [55] from 8 Q. ilex dominant trees showing no signs of decay. To avoid wound effects, we extracted the microcores, collected from the trunk at breast height, by following a spiral with a distance of 4–6 cm between consecutive samples [30]. The microcores containing phloem, cambium, and a few millimetres of xylem were immediately fixed in 70% ethanol where they were kept for 1 week. Samples were then fixed in F.A.A. (38% formaldehyde, glacial acetic acid, 50% ethanol—5/5/90 by volume) for a week. Microcores were subsequently dehydrated in a graded ethanol series (70, 90, 95, and 100%), infiltrated with bio-clear (D-limonene), and embedded in paraffin blocks using a Leica TP1020-1 (Nussloch, Germany) tissue processor. Cross-sections (9 μm thick) were obtained with a semi-automatic rotary microtome RM 2245 (Leica, Nussloch) and stained with a safranin (0.04%) and astra blue (0.15%) water solution [56]. The sections were observed under a Nikon Eclipse 800 light microscope, and microphotographs were taken using a DS-Fi1 digital camera with the NIS-Elements BR 3 image analysis system (Melville, NY, USA). The sections were analysed to identify cambial cells and differentiating xylem and phloem cells using visual criteria of proportional dimensions and wall thickness [32,34,38,57]. In detail, we counted the cambial cells (CC) which appeared radially flattened (Figure 2A). Then, we identified the period when post-cambial cells (PC) (Figure 2B,C) were formed on the xylem side. PC are cells in radial enlargement, showing only primary walls that do not shine under polarised light compared with cells with secondary walls [30,58].
We also detected the time of likely transition from early to late phloem [59], identified by the appearance of small, tangentially orientated groups of phloem fibres (PF) (Figure 2A,B) separating the 2 parts [50]. Such fibres could be detected as long slender cells developing from fusiform initials of the cambium and reaching maturity in the non-collapsed phloem [60]. Mature phloem fibres had thick lignified cell walls that stained red with safranin, exhibited birefringence in polarised light, and were considered a part of the late phloem (Figure 2C) [51,60]. Moreover, we applied quantitative anatomy to measure the distance between the cambium and the nearest phloem fibres cluster (PF) to use it as a reference point to estimate the time and phases of phloem formation. Finally, we defined the increments of xylem and non-collapsed phloem (Figure 2A) on the last sampling date at the end of 2017. The whole portion of xylem produced in the year 2017 was classified for the presence of IADFs on the basis of the occurrence of 1 (ring without IADF) or more (ring with IADFs, namely, with an extra growth band) xylem increments [25].
After cutting the slides for light microscopy, we prepared the remaining parts of the microcores for observation with a scanning electron microscope (SEM). To this purpose, they were infiltrated with bio-clear for 1 night and washed with alcohol to remove the paraffine. Samples were then mounted on stubs with a conductive carbon adhesive and sputter-coated with Au/Pd (Q150R ES Coating System; Quorum technologies, Laughton, UK) for 30 s with a constant current of 20 mA. SEM micrographs were taken at voltages between 5 kV and low vacuum conditions with an Everhart–Thornley detector (ETD) in a FEI Quanta 250 SEM microscope (FEI Company, Hillsboro, OR, USA) at a working distance of 9–10 mm [61].
3. Results
3.1. Xylem Anatomical Features
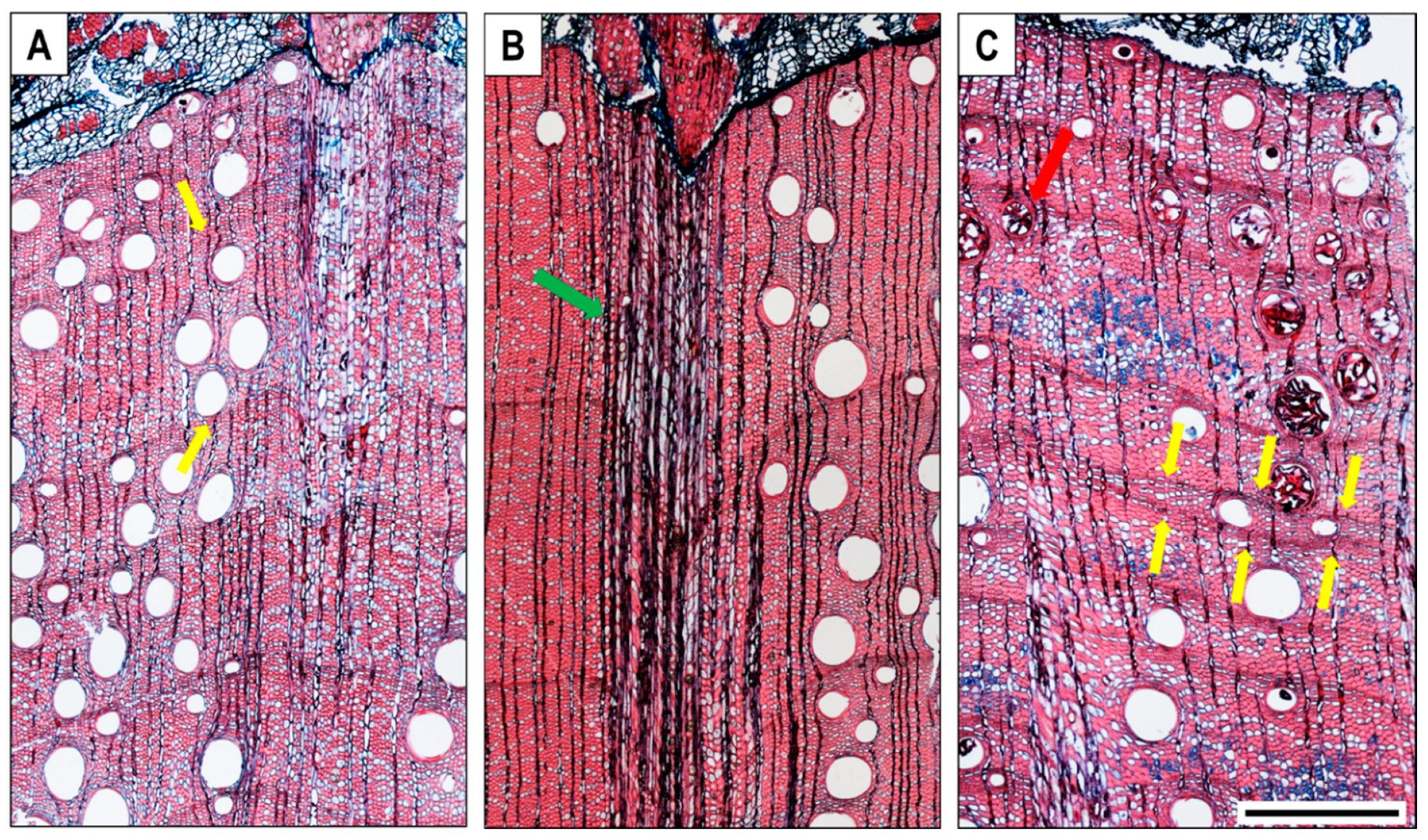
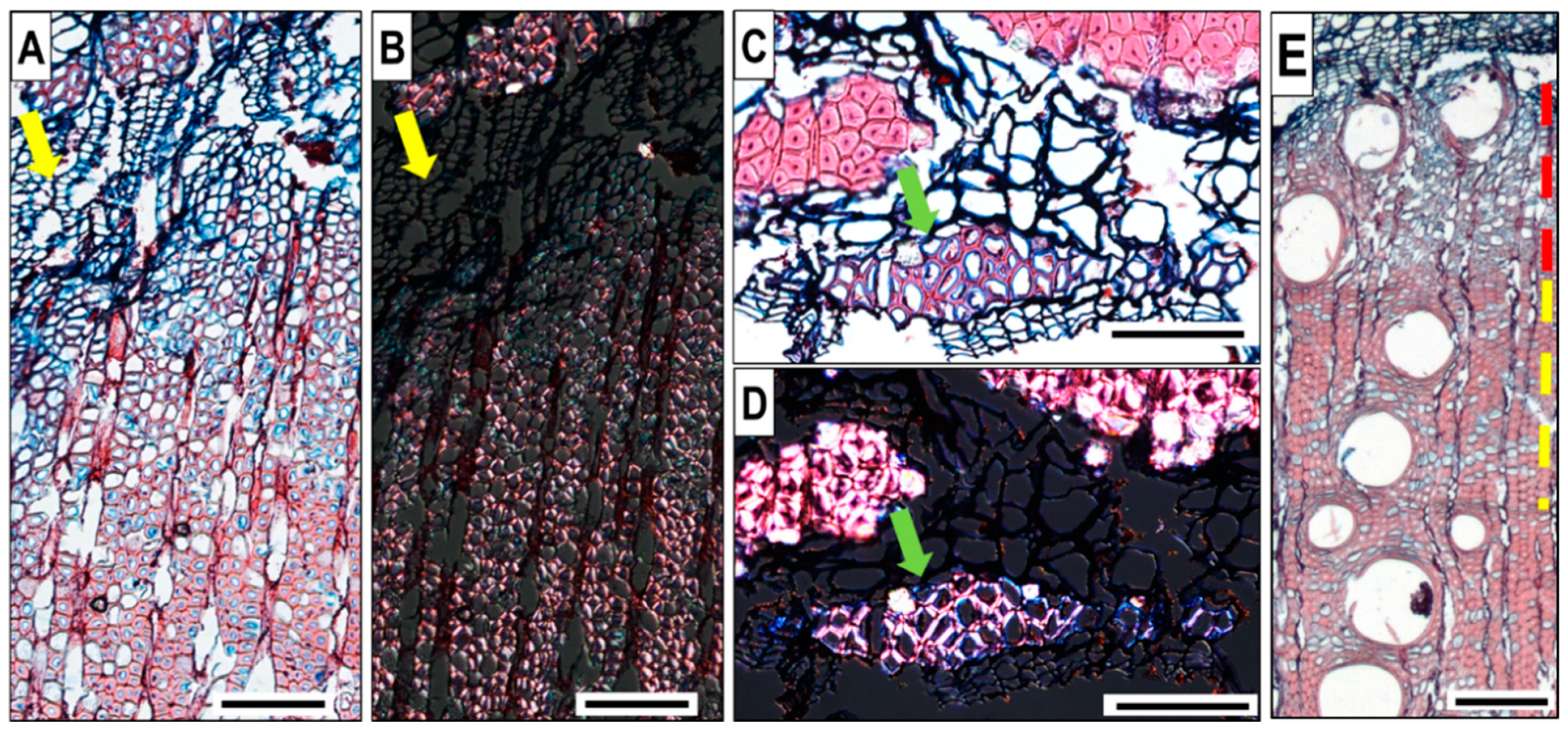
The cross-sections of Q. ilex microcores contained several growth layers of mature wood of the previous years. Microcores taken from different trees on the same date showed high variability in the tree-ring structure (Figure 3). The analysis of growth increments deviated from the normal structure of a ‘‘diffuse to semi-ring-porous’’ wood in so far as the transition from earlywood to latewood was not evident; wood increments mostly held large vessels not always decreasing from the beginning towards the end of the ring (Figure 3A yellow arrows), and the boundary of a growth increment was marked only by flattened fibres. Large wide multiseriate rays (Figure 3B green arrow), which were fewer in number than the uniseriate ones, sometimes covered a substantial portion of the transverse section of a microcore, thus hampering the observation. The presence of solitary prismatic crystals was variable, and they occurred in both ray and axial parenchyma cells, sometimes occurring in a very large portion of the axial parenchyma cells. Discontinuous rings (wedging rings) (Figure 3C yellow arrow) were commonly found, which made the tree-ring boundary identification difficult. We also detected the formation of tyloses in vessels, even in recently formed wood (Figure 3C red arrow, Figure 4).
3.2. Cambial Activity and Wood Increments Formed Throughout the Year
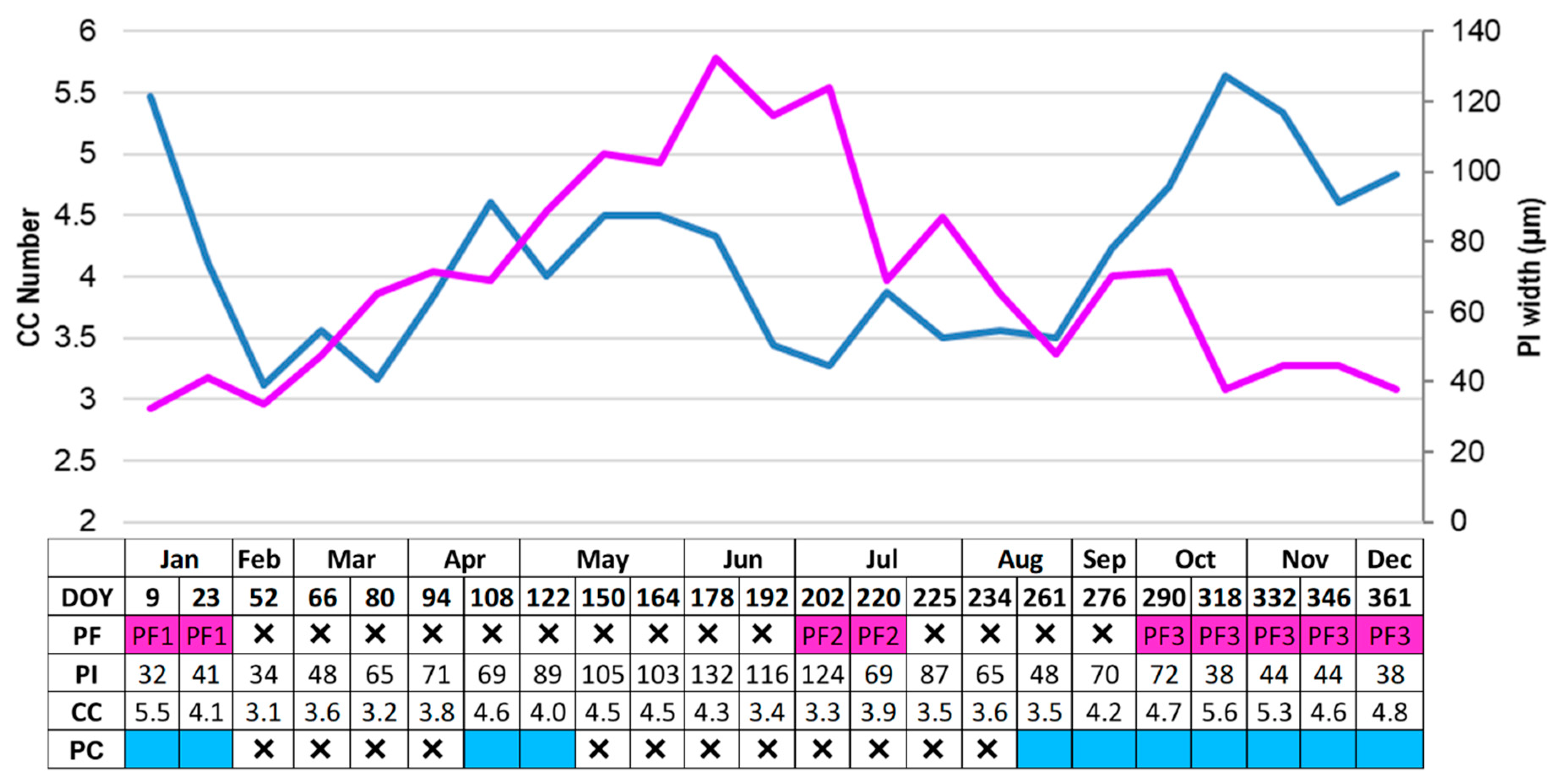
At the beginning of the sampling in January 2017 (9 DOY, day of the year), the cambium was active, as indicated by the number of CC (cambial cells) (5.5 ± 1.2), with the presence of PC (post-cambial) cells indicating that cambium was producing xylem cells (Figure 5). Throughout the sampling period, the number of CC varied in a multimodal pattern, with minimum and maximum number of 3.1 and 5.6 cells, respectively. CC number decreased to a minimum at 52 DOY, then increased reaching a maximum in April (108 DOY, 4.6 ± 1.0) and remained stable until June (178 DOY) (Figure 5). After a decrease in July (202 DOY), CC number started to increase again, reaching a maximum value in November (318 DOY); afterwards, it decreased and raised again at the end of the year (361 DOY). PC, consisting of xylem cells with primary wall and indicating that cambium is active at the xylem side, were detected in three periods of the year. PC were detected between 9 and 16 DOY, but after these two weeks, no PC cells were detected until the middle of April (108 DOY). After the first week of May, with the driest months July and August (Figure 1), no PC cells were formed until the end of August (Figure 5).
At the beginning of September, PC were again evident and their production lasted until December (346 DOY) (Figure 5 and Figure 6A,B, showing PC in October). PC production was thus detected in three periods of the year, namely, January, April, and October–December when the highest number occurred. In the other periods, despite the occurrence of CC, the absence of PC led to the formation of IADFs. The IADF structure differed from the structure usually described as “extra growth band”, but appeared as a second growth band accounting for approximately at 50% of the total wood increment, in which mostly large earlywood vessels appeared, such as in the first growth increment (Figure 6E). Therefore, the cross-sections of the microcores sampled in December showed two wood increments formed during the year (Figure 6E). The first, formed in April, appeared narrower than the second, formed from October until December. On the last sampling date, most of the samples showed relatively large total xylem increments with identifiable boundaries, while others showed very narrow growth layers with no clear boundaries or no xylem formed at all (Table 1). The trees not forming xylem were not characterised by any external sign of decline or wounding which could have been interpreted as indicators of a peculiar condition of stress. An IADF was formed in each total xylem increment (Table 1).
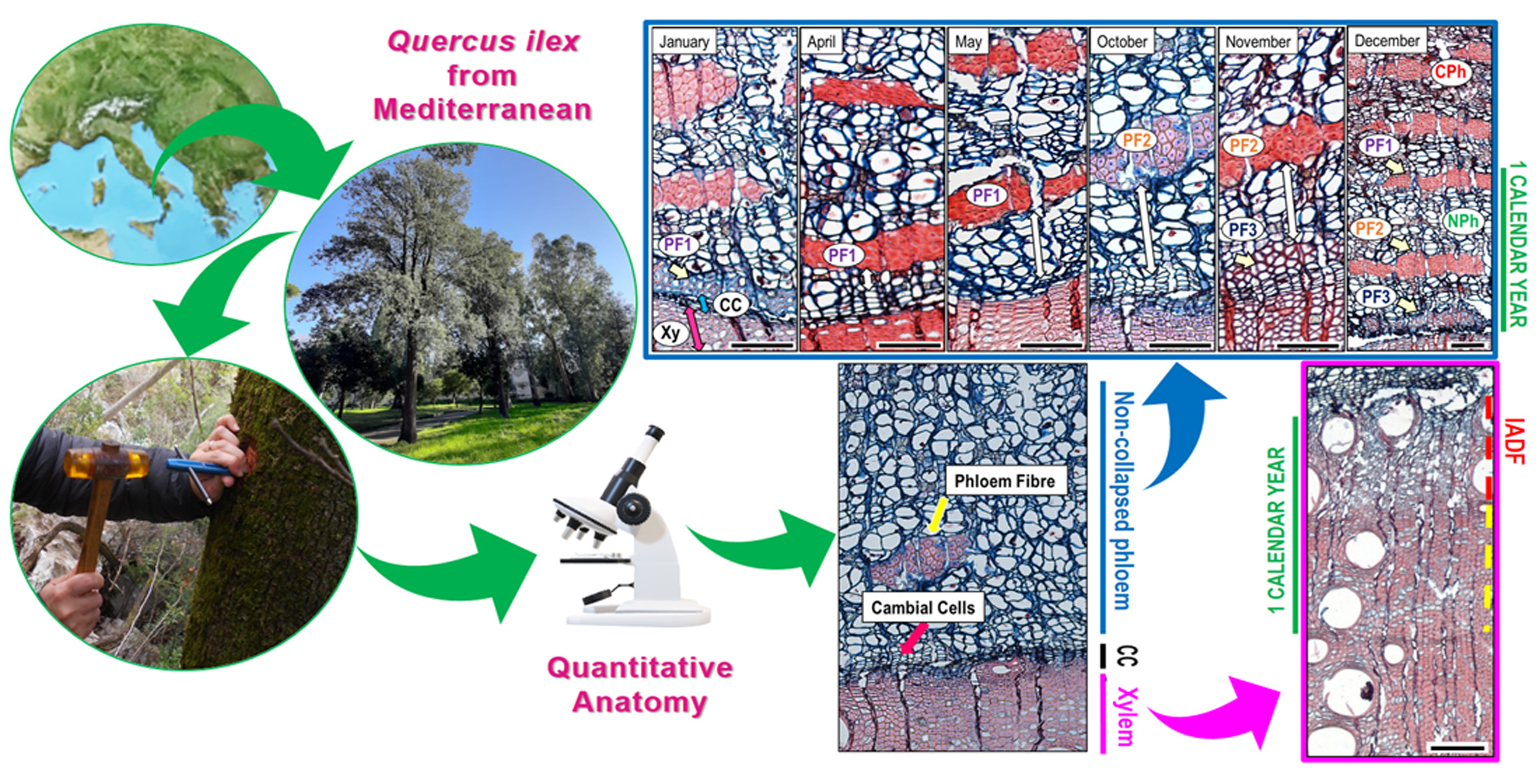
The development of phloem fibres (PF), in ring porous oaks, normally indicating the transition from early to late phloem, was detected in three periods of the year: in January (PF1) (9–23 DOY), between the end of June and July (PF2) (202–220 DOY), and between the end of October and December (PF3) (290–361 DOY) (Figure 5 and Figure 6c,d). At the beginning, the distance between PF1 and cambium was the lowest (Figure 5 and Figure 7). The distance began to increase in April and especially May, until the maximum at the beginning of June (Figure 7) when the second PF2 group appeared (Figure 5) and at the end of September/beginning of October (Figure 7), before the third group of PF3 was formed (Figure 5). In samples collected at the end of the sampling year, the width of non-collapsed phloem increment was similar in all sampled trees, as well as in the two trees where no xylem increment was observed (Table 1).
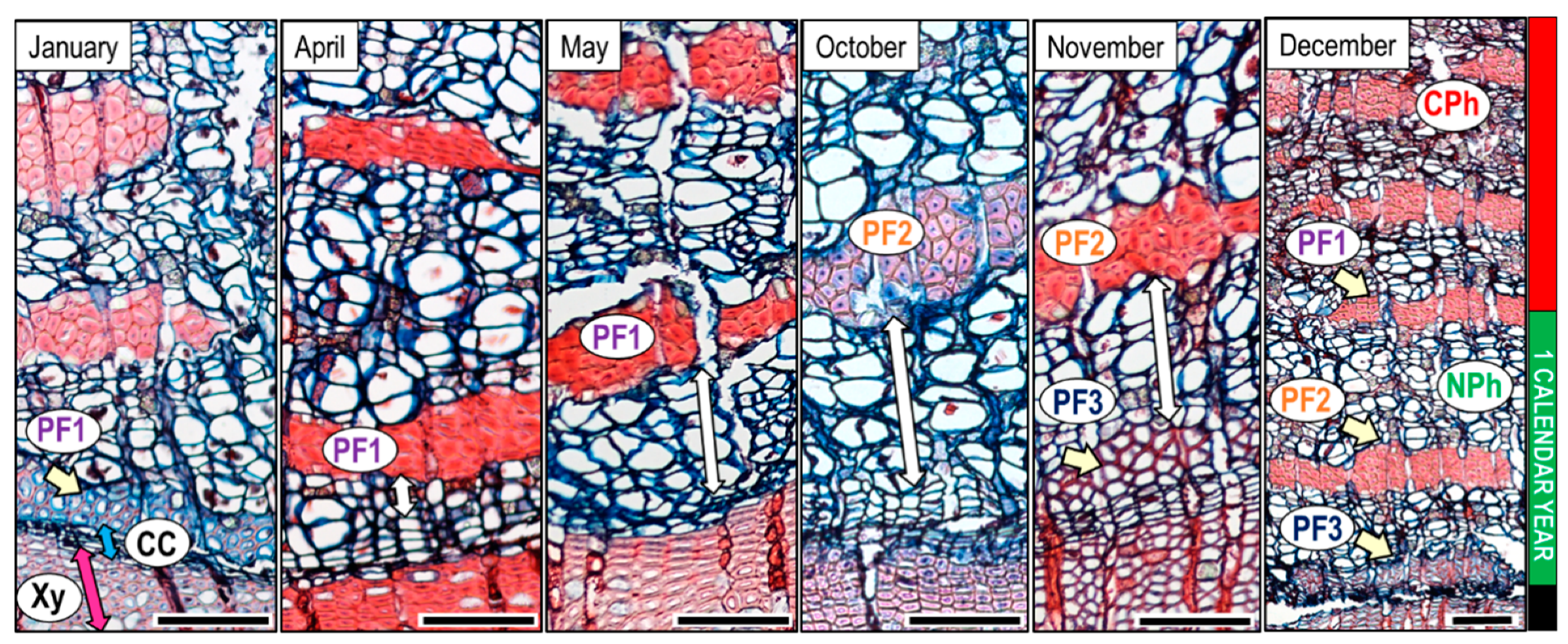
In the samples collected in December, we could observe the three groups of PF (PF1, PF2, PF3) formed throughout the calendar year, which indicate the total phloem annual increment. The annual increment mostly consisted of non-collapsed phloem; indeed collapsed phloem is only present in proximity of PF1 formed at the beginning of January (Figure 7).
Figure 7 shows the phloem genesis of tree number 7. The first PF group (PF1) was located in immediate vicinity of the cambium in January, with fibres in the phase of lignification (Figure 7). Until April and especially in May, PF1 appeared with cell walls completely lignified. Due to the phloem formed between January and the sampling date, PF1 was located at larger distances from the cambium. In October, the second group PF2 appeared, whose cells had still thin, non-lignified cell-walls. In November, the PF2 cell walls were thicker and lignified and PF2 were farther from the cambium, while PF3 in early stage of development appeared close to the cambium. The described processes indicate dynamics of phloem formation and the distances shown by arrows with double head in Figure 7 indicate how much phloem was formed by the given date. As phloem in December contained non-collapsed phloem (NPh) with three groups of fibres formed throughout the calendar year of 2017, and the phloem formed before PF1 started to collapse, we suppose that the width of the phloem formed throughout the calendar year approximately agrees with the width of the non-collapsed phloem. In five out of eight trees, the ratio between xylem and phloem production in the year 2017 was around 2:1, as wood production was almost double that of phloem production (NPh) (Table 1).
4. Discussion
4.1. Cambial Activity and Xylem Increment
Q. ilex, as opposed to most European oaks, is not ring porous. Therefore, it is often difficult to define its tree-rings and tree-ring boundaries. Dendrochronological studies have previously reported on IADFs in Q. ilex wood described as “extra growth bands” located at the end of the ring and formed due to bimodal cambial activity [25,29,35]. In our samples of Q. ilex from southern Italy, we detected a multimodal pattern of cambial activity. The active production of xylem by the cambium was indicated by an increase in the number of cells (and width) of CC and the presence of wood cells in PC phase. We observed the highest peak of cambial activity during the autumn months and not in spring. This finding is not in line with the literature, which mainly reports that spring is the most active period for growth in Q. ilex, while generally the trees enter dormancy in winter and summer [11,22,27,62,63]. This peculiar pattern is likely due to the excessively dry spring/summer and wet and mild winter during the year 2017, which triggered a slow-down or even a stop in xylogenesis in spring/summer. This would also explain why two of eight trees did not produce any xylem at all during the year 2017. The variability in the xylem production found among Q. ilex trees is in line with the high variability in tree-ring width and IADF occurrence in Mediterranean softwood and hardwood species [22,34]. Indeed, such variability can be ascribed to the different sensitivity of the single trees to environmental constraints, also due to their physiological status that influences the onset and duration of the different phases of xylogenesis [30].
In general, the observed variation of CC number seems to follow the trend in precipitation, suggesting a very prompt response of the cambium (activating and deactivating several times during the year) to the rain input. Such plasticity allows the trees to produce biomass even in stressful environments in a sort of prompt start-and-stop cambium production of different intensity following the environmental fluctuations. Indeed, the highest peak of CC number was detected in November, the wettest month of the year 2017 with warm temperatures. As a result of the activation of CC in autumn, the majority of the investigated trees formed IADFs. The type of IADFs detected had different structure from the usual structure described in Q. ilex [25,29,35,64]. It appeared indeed very similar to the first growth increment with mostly large vessels and no intra-seasonal diffuse parenchyma. By definition of IADF, we should consider the IADF as the transition zone between the first and second increment. However, for simplicity and for coherence with previous classifications of IADFs in Q. ilex, we will refer to the second growth increment as IADF, although in our case the anatomical traits differ from previous findings. The absence of a true latewood (with smaller vessels) in the first growth increment and in IADF is likely the result of sudden cessation of wood production due to the beginning of the drought period and the lowering of temperatures, respectively.
It has already been reported that in other species growing at dry sites, the water availability at the growth site has the highest influence on growth rates and cessation of wood formation [65]. A previous tree-ring study in Q. ilex growing at a nearby site in southern Italy showed that the occurrence of IADFs was linked to rain events following periods of summer drought [35]. In our case, the IADFs in Q. ilex would be similar to L-type IADFs found in many Mediterranean pines and possibly ascribable to a second flush of growth after the restoration of favourable conditions [23,66]. Besides IADFs, we also detected wedging rings, whose development is probably the result of locally absent cambial production at a certain part of the stem [67].
Extreme drought stress could also provoke the occurrence of tyloses in the vessels of the latest formed xylem, very soon after vessel formation. Indeed, tyloses in sapwood can be linked to vessel embolism or can be a consequence of a traumatic event [68,69,70]. Tyloses block vessels in the outer sapwood crucial for water conduction, and therefore they might affect tree survival. In our case, trees showed no sign of mechanical stress, and thus we suppose that tyloses were formed as a response to the severe drought stress in the 2017 year. We also detected the presence of starch in xylem parenchyma of the last xylem increment on all sampling dates. Starch grains in the storage system of the xylem are needed to recover from embolism thanks to osmotic regulation mechanisms. Indeed, the ability to recover from embolism is species-specific and correlated with the concentration of soluble carbohydrates accumulated at the stem level [71,72]. Q. ilex’s high capacity for osmotic adjustment is recognised [24] as a mechanism possibly working in the case of short-term drought events, but not in case of drought stress protracted in time, which would lead to tree decline [73,74]. The occurrence of tyloses and abundance of starch may also suggest the beginning of a fungal infection, to which stressed plants are most vulnerable.
4.2. Phloem Formation
To our knowledge, phloem formation processes in Q. ilex, and generally in Mediterranean hardwoods, has not been investigated yet. Although it has been shown that in Mediterranean conifers [47,48,49] it is not possible to identify phloem annual rings with early and late phloem, and it is not possible to follow the amount of phloem formed between two samplings, in Q. ilex, the occurrence of phloem fibres can help estimate the amount of phloem formed between two samplings. In Castanea sativa growing at a site in central Italy, with similar phloem structure as oaks, the transition from early to late phloem was marked by the occurrence of PF; late phloem was formed from the end of June until the end of September [59]. However, in Q. ilex, we detected the development of PF in three periods of the calendar year 2017: in January, between June and July, and between November and December. In the samples collected in January, the PF1 (where cell-wall thickening and lignification were not completed yet) was located in immediate vicinity of the cambium and was probably formed in the previous autumn/winter 2016. Assuming that the PF are part of the late phloem of the previous year [50,59], we presume that phloem formed between January and June may be ascribed to early phloem and the appearance of PF2 between June and July likely indicates the transition between early and late phloem. Nevertheless, to confirm this distinction between potential early and late phloem, we needed to test additional measurements of phloem cell size in Q. ilex. Using PF (PF1, PF2, PF3) as reference points, we were able to follow phloem formation during the year to evaluate the phloem increment during time. The width of annual phloem increment formed between January and December agreed with the width of non-collapsed phloem, suggesting that the phloem remains non-collapsed and possibly active for approximately one year in Q. ilex. This finding agrees with the results of a detailed study in beech conducted by Prislan et al. [21].
Our study proved that even during the period when xylem was not produced, phloem development was still ongoing and lasted the entire year, even in the trees not forming any xylem at all. The width of non-collapsed phloem measured on the last sampling date turned out to be less variable among the trees than xylem width, suggesting that, under stressful conditions, phloem production is either favoured or less affected compared to xylem, in agreement with observations in temperate and Mediterranean conifers [46,48,49].
4.3. Technical Notes to Improve Sample Preparation of Q. ilex
Q. ilex has a wood of very high density and hardness, and very hard bark. Sampling of cores and microcores is therefore technically very demanding. Furthermore, the tissues are anatomically very heterogenous with great differences between very hard wood, with great portion of very thick-walled fibres, followed by soft, extremely thin-walled cambium, thin-walled non-collapsed phloem alternating with groups of very hard thick-walled fibres, and hard collapsed outer bark. Therefore, it is extremely difficult to produce thin sections without failures such as torn and collapsed parts. Due to very heterogeneous tissues, the microcore often breaks in the region of cambium which is then destroyed. In addition, Q. ilex has, like all oaks, very wide (>10 cells) massive rays which potentially become sclerified in the bark. In the worst cases, the microcore can mostly consist of one big ray. In the vicinity of the ray, de-orientations of the tissue are frequent, and the measurement of cell dimensions is not possible. All this hampers the estimation of the number of cells the cambium is producing on both xylem and phloem sides.
We also observed high variability in the anatomical structure along the circumference of the stem, where irregular occurrence of wedging (incomplete) rings and IADFs constrained the measurements and observations. This high variability along the stem circumference is also typical of Mediterranean hardwoods, as already shown in Arbutus unedo L. [34]. Although, despite all these constraints, we were able to reconstruct the pattern of cambial activity with xylem and phloem formation in Q. ilex. There are some measures that can help avoiding the bias. A bigger Trephor (cutter diameter > 2 mm) [55] for microcoring of Q. ilex would help obtaining wider tissue portion to perform the analysis, even in the cases in which the wide rays occur in the sample. The use of very sharp Trephor is needed. Perfectly sharp blades for sectioning are also essential; it is often required that the position of the blade is changed at every cut. Embedding in hard-grade resins could also be very useful.
5. Conclusions
Q. ilex trees growing at a dry site in southern Italy showed a cambium production throughout the year with interruptions in wood production in some periods of the year, while the production of phloem was uninterrupted. The multimodal pattern in cambial production on the xylem side seemed to be linked with the precipitation pattern and lead to the formation of IADFs. Xylem production was scanty in the first half of the year, while more pronounced between October/December, likely due to the extremely dry spring/summer and wet and mild winter of 2017. In the outer part of the xylem, the occurrence of tyloses in the vessels and starch accumulation in parenchyma cells suggested possible stress due to drought and/or possible fungal infection favoured by the harsh climatic conditions. Appling quantitative anatomy to measure the cells and tissues in phloem and using groups of phloem fibres as orientation points to detect phloem formation between the sampling dates, we showed that phloem was formed throughout the entire year, even in the periods with no xylem production and in trees not producing wood increments.
Q. ilex forests subjected to stressful conditions due to abiotic and biotic factors represent a useful model for studying cambial activity and to better understand adaptation ability of this species in a scenario of climate change.
The study of xylem and phloem genesis in Q. ilex allowed us to identify the growth increments throughout a calendar year, also showing the occurrence of IADFs in wood with different anatomical traits compared to the typical IADFs described for this species, suggesting that their genesis and possible functional meaning are similar to L-type IADFs found in Mediterranean pines.
Author Contributions
Conceptualisation, A.B., K.Č., and V.D.M.; methodology, A.B., K.Č., and V.D.M.; formal analysis, A.B. and V.D.M.; investigation, A.B. and V.D.M.; resources, A.B., V.D.M., and K.Č.; data curation, A.B., V.D.M., and K.Č.; writing—original draft preparation, A.B., V.D.M., and K.Č.; writing—review and editing, A.B., V.D.M., and K.Č.; visualisation, A.B., K.Č., and V.D.M.; supervision, V.D.M.; project administration, K.Č.; funding acquisition, K.Č. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Slovenian Research Agency (ARRS), program P4-0015.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Masson-Delmotte, V.; Zhai, P.; Pörtner, H.O.; Roberts, D.; Skea, J.; Shukla, P.R.; Pirani, A.; Moufouma-Okia, W.; Péan, C.; Pidcock, R.; et al. Global warming of 1.5 C. IPCC Spec. Rep. Impacts Glob. Warm. 1 2018, 1, 1–9. [Google Scholar]
- Ogaya, R.; Liu, D.; Barbeta, A.; Peñuelas, J. Stem mortality and forest dieback in a 20-year experimental drought in a Mediterranean holm oak forest. Front. For. Glob. Chang. 2019, 2, 89. [Google Scholar] [CrossRef]
- De Marco, A.; Arena, C.; Giordano, M.; De Santo, A.V. Impact of the invasive tree black locust on soil properties of Mediterranean stone pine-holm oak forests. Plant Soil 2013, 372, 473–486. [Google Scholar] [CrossRef]
- Barbero, M.; Loisel, R.; Quézel, P. Biogeography, ecology and history of Mediterranean Quercus ilex ecosystems. In Quercus ilex L. Ecosystems: Function, Dynamics and Management; Springer: Dordrecht, The Netherlands, 1992; pp. 19–34. [Google Scholar]
- Canadell, J.; Roda, F. Root biomass of Quercus ilex in a montane Mediterranean forest. Can. J. For. Res. 1991, 21, 1771–1778. [Google Scholar] [CrossRef] [Green Version]
- Savé, R.; Castell, C.; Terradas, J. Gas exchange and water relations. In Ecology of Mediterranean Evergreen Oak Forests; Springer: Berlin/Heidelberg, Germany, 1999; pp. 135–147. [Google Scholar]
- Infante, J.M.; Domingo, F.; Alés, R.F.; Joffre, R.; Rambal, S. Quercus ilex transpiration as affected by a prolonged drought period. Biol. Plant 2003, 46, 49–55. [Google Scholar] [CrossRef]
- Tognetti, R.; Longobucco, A.; Raschi, A. Vulnerability of xylem to embolism in relation to plant hydraulic resistance in Quercus pubescens and Quercus ilex co-occurring in a Mediterranean coppice stand in central Italy. New Phytol. 1998, 139, 437–447. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Mangirón, M.; Ogaya, R.; Sauret, M.; Serrano, L.; Peñuelas, J.; Piñol, J. Sap flow of three co-occurring Mediterranean woody species under varying atmospheric and soil water conditions. Tree Physiol. 2003, 23, 747–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sade, N.; Gebremedhin, A.; Moshellon, M. Risk taking plants. Anisohydric behaviour as a stress resistance trait. Plant Signal Behav. 2012, 7, 767–770. [Google Scholar] [CrossRef] [Green Version]
- Corcuera, L.; Camarero, J.J.; Gil-Pelegrín, E. Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 2004, 18, 83–92. [Google Scholar] [CrossRef]
- Ogaya, R.; Peñuelas, J. Species-specific drought effects on flower and fruit production in a Mediterranean holm oak forest. Forestry 2007, 80, 351–357. [Google Scholar] [CrossRef] [Green Version]
- Barbeta, A.; Ogaya, R.; Peñuelas, J. Dampening effects of long-term experimental drought on growth and mortality rates of a Holm oak forest. Glob. Chang. Biol. 2013, 19, 3133–3144. [Google Scholar] [CrossRef]
- Liu, D.; Ogaya, R.; Barbeta, A.; Yang, X.; Peñuelas, J. Long-term experimental drought combined with natural extremes accelerate vegetation shift in a Mediterranean holm oak forest. Environ. Exp. Bot. 2018, 151, 1–11. [Google Scholar] [CrossRef]
- Lloret, F.; Siscart, D. Drought demographic effects in Holm Oak populations. Cuad. Soc. Esp. Cien. For. 1995, 2, 77–81. [Google Scholar]
- Peñuelas, J.; Filella, I.; Lloret, F.; Piñol, J.; Siscart, D. Effects of a severe drought on water and nitrogen use by Quercus ilex and Phyllirea latifolia. Biol. Plant 2000, 43, 47–53. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Piñol, J.; Beven, K. A hydraulic model to predict drought-induced mortality in woody plants: An application to climate change in the Mediterranean. Ecol. Model. 2002, 155, 127–147. [Google Scholar] [CrossRef]
- Lorenz, M.; Fischer, R.; Becher, G.; Mues, V.; Seidling, W.; Kraft, P.; Nagel, H.D. Forest Condition in Europe: 2006 Technical Report of ICP Forests (No. 2006/1); Work Report of the Institute for World Forestry: Hamburg, Germany, 2006; Available online: http://hdl.handle.net/10419/96611 (accessed on 1 September 2006).
- Bonal, R.; Muñoz, A. Multi-trophic effects of ungulate intraguild predation on acorn weevils. Oecologia 2007, 152, 533–540. [Google Scholar] [CrossRef] [PubMed]
- De Micco, V.; Carrer, M.; Rathgeber, C.B.K.; Camarero, J.J.; Voltas, J.; Cherubini, P.; Battipaglia, G. From xylogenesis to tree rings: Wood traits to investigate tree response to environmental changes. IAWA J. 2019, 40, 155–182. [Google Scholar] [CrossRef] [Green Version]
- Prislan, P.; Čufar, K.; De Luis, M.; Gričar, J. Precipitation is not limiting for xylem formation dynamics and vessel development in European beech from two temperate forest sites. Tree Physiol. 2018, 38, 186–197. [Google Scholar] [CrossRef] [Green Version]
- Cherubini, P.; Gartner, B.L.; Tognetti, R.; Braeker, O.U.; Schoch, W.; Innes, J.L. Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol. Rev. 2003, 78, 119–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Micco, V.; Campelo, F.; De Luis, M.; Bräuning, A.; Grabner, M.; Battipaglia, G.; Cherubini, P. Intra-annual density fluctuations in tree rings: How, when, where, and why? IAWA J. 2016, 37, 232–259. [Google Scholar] [CrossRef]
- Terradas, J.; Savé, R. The influence of summer and winter stress and water relationships on the distribution of Quercus ilex L. In Quercus ilex L. Ecosystems: Function, Dynamics and Management; Springer: Dordrecht, The Netherlands, 1992; pp. 137–145. [Google Scholar]
- Campelo, F.; Gutiérrez, E.; Ribas, M.; Nabais, C.; Freitas, H. Relationships between climate and double rings in Quercus ilex from northeast Spain. Can. J. For. Res. 2007, 37, 1915–1923. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Martín-Benito, D.; Cherubini, P.; Isabel, C. Climate-growth variability in Quercus ilex L. west Iberian open woodlands of different stand density. Ann. For. Sci. 2009, 66, 802. [Google Scholar] [CrossRef] [Green Version]
- Gea-Izquierdo, G.; Cherubini, P.; Cañellas, I. Tree-rings reflect the impact of climate change on Quercus ilex L. along a temperature gradient in Spain over the last 100 years. For. Ecol. Manag. 2011, 262, 1807–1816. [Google Scholar] [CrossRef]
- Gutiérrez, E.; Campelo, F.; Camarero, J.J.; Ribas, M.; Muntán, E.; Nabais, C.; Freitas, H. Climate controls act at different scales on the seasonal pattern of Quercus ilex L. stem radial increments in NE Spain. Trees 2011, 25, 637–646. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Contrasting physiological responses to Mediterranean climate variability are revealed by intra-annual density fluctuations in tree rings of Quercus ilex L. and Pinus pinea L. Tree Physiol. 2018, 38, 1213–1224. [Google Scholar] [CrossRef] [PubMed]
- Balzano, A.; Čufar, K.; Battipaglia, G.; Merela, M.; Prislan, P.; Aronne, G.; De Micco, V. Xylogenesis reveals the genesis and ecological signal of IADFs in Pinus pinea L. and Arbutus unedo L. Ann. Bot. 2018, 121, 1231–1242. [Google Scholar] [CrossRef]
- Balzano, A.; De Micco, V.; Merela, M.; Čufar, K. Tree-rings in Mediterranean pines–can we ascribe them to calendar years? Les/Wood 2019, 68, 5–14. [Google Scholar] [CrossRef] [Green Version]
- De Luis, M.; Gričar, J.; Čufar, K.; Raventós, J. Seasonal dynamics of wood formation in Pinus halepensis from dry and semi-arid ecosystems in Spain. IAWA J. 2007, 28, 389–404. [Google Scholar] [CrossRef] [Green Version]
- Novak, K.; De Luis, M.; Gričar, J.; Prislan, P.; Merela, M.; Smith, K.T.; Čufar, K. Missing and dark rings associated with drought in Pinus halepensis. IAWA J. 2016, 37, 260–274. [Google Scholar] [CrossRef]
- De Micco, V.; Balzano, A.; Čufar, K.; Aronne, G.; Gričar, J.; Merela, M.; Battipaglia, G. Timing of false ring formation in Pinus halepensis and Arbutus unedo in Southern Italy: Outlook from an analysis of xylogenesis and tree-ring chronologies. Front. Plant Sci. 2016, 7, 705. [Google Scholar] [CrossRef] [Green Version]
- Balzano, A.; Battipaglia, G.; Cherubini, P.; De Micco, V. Xylem Plasticity in Pinus pinaster and Quercus ilex Growing at Sites with Different Water Availability in the Mediterranean Region: Relations between Intra-Annual Density Fluctuations and Environmental Conditions. Forests 2020, 11, 379. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.H.; Romane, F. Diameter growth of Quercus ilex L. and the interannual variability of climatic characteristics. Ann. Sci. For. 1991, 48, 225–234. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; De Micco, V. Site conditions influence the climate signal of intra-annual density fluctuations in tree rings of Q. ilex L. Ann. For. Sci. 2018, 75, 68. [Google Scholar] [CrossRef] [Green Version]
- Čufar, K.; Prislan, P.; De Luis, M.; Gričar, J. Tree-ring variation, wood formation and phenology of beech (Fagus sylvatica) from a representative site in Slovenia, SE Central Europe. Trees 2008, 22, 749–758. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; De Luis, M.; Smith, K.T.; Čufar, K. Phenological variation in xylem and phloem formation in Fagus sylvatica from two contrasting sites. Agric. For. Metereol. 2013, 180, 142–151. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; De Luis, M.; Novak, K.; Martinez del Castillo, E.; Schmitt, U.; Koch, G.; Štrus, J.; Mrak, P.; Žnidarič, M.T.; et al. Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front. Plant Sci. 2016, 7, 1923. [Google Scholar] [CrossRef] [Green Version]
- Prislan, P.; Gričar, J.; Čufar, K.; De Luis, M.; Merela, M.; Rossi, S. Growing season and radial growth predicted for Fagus sylvatica under climate change. Clim. Chang. 2019, 153, 181–197. [Google Scholar] [CrossRef]
- Gričar, J.; Prislan, P.; De Luis, M.; Gryc, V.; Hacurová, J.; Vavrčík, H.; Čufar, K. Plasticity in variation of xylem and phloem cell characteristics of Norway spruce under different local conditions. Front. Plant Sci. 2015, 6, 730. [Google Scholar] [CrossRef] [Green Version]
- Del Castillo, E.M.; Prislan, P.; Gričar, J.; Gryc, V.; Merela, M.; Giagli, K.; De Luis, M.; Vavrčík, H.; Čufar, K. Challenges for growth of beech and co-occurring conifers in a changing climate context. Dendrochronologia 2018, 52, 1–10. [Google Scholar] [CrossRef]
- Gričar, J. Cambial cell production and structure of xylem and phloem as an indicator of tree vitality: A review. In Sustainable Forest Management-Current Research; IntechOpen: Rijeka, Hrvatska, 2012; pp. 111–134. [Google Scholar]
- Gričar, J.; Jagodic, Š.; Šefc, B.; Trajković, J.; Eler, K. Can the structure of dormant cambium and the widths of phloem and xylem increments be used as indicators for tree vitality? Eur. J. For. Res. 2014, 133, 551–562. [Google Scholar] [CrossRef] [Green Version]
- Gričar, J.; Krže, L.; Čufar, K. Number of cells in xylem, phloem and dormant cambium in silver fir (Abies alba), in trees of different vitality. IAWA J. 2009, 30, 121–133. [Google Scholar] [CrossRef] [Green Version]
- Gričar, J.; Prislan, P.; De Luis, M.; Novak, K.; Longares, L.A.; del Castillo, E.M.; Čufar, K. Lack of annual periodicity in cambial production of phloem in trees from Mediterranean areas. IAWA J. 2016, 37, 349–364. [Google Scholar] [CrossRef] [Green Version]
- Balzano, A.; De Micco, V.; Čufar, K.; De Luis, M.; Gričar, J. Intra-seasonal trends in phloem traits in Pinus spp. from drought-prone environments. IAWA J. 2020, 41, 219–235. [Google Scholar] [CrossRef]
- Sökücü, A.; Güney, A. Vitality versus cambium, xylem, and phloem characteristics in Cedrus libani. IAWA J. 2020, 1, 1–15. [Google Scholar] [CrossRef]
- Gričar, J. Xylem and phloem formation in sessile oak from Slovenia in 2007. Wood Res. 2007, 55, 15–22. [Google Scholar]
- Gričar, J.; Jagodic, Š.; Prislan, P. Structure and subsequent seasonal changes in the bark of sessile oak (Quercus petraea). Trees 2015, 29, 747–757. [Google Scholar] [CrossRef]
- Teobaldelli, M.; Cona, F.; Stinca, A.; Saulino, L.; Anzano, E.; Giordano, D.; Migliozzi, A.; Bonanomi, G.; D’Urso, G.; Mazzoleni, S.; et al. Improving resilience of an old-growth urban forest in Southern Italy: Lesson (s) from a stand-replacing windstorm. Urban For. Urban Green. 2020, 47, 126521. [Google Scholar] [CrossRef]
- Ricciardi, M.; Mazzoleni, S.; Pizzolongo, P. Il bosco del parco Gussone in Portici. Aspetti naturalistici e problemi di conservazione. In Parchi e Giardini Storici, Parchi Letterari. Conoscenza, tutela e valorizzazione. Atti III Convegno (primo internazionale)“Paesaggi e Giardini del Mediterraneo”. 2-La valorizzazione; Ministero per i Beni Culturali e Ambientali. Comitato Nazionale per lo studio e la conservazione dei Giardini Storici. Soprintendenza Archeologica di Pompei, Tipolitografia GRG: Salerno, Italy, 1993. [Google Scholar]
- Stinca, A.; Motti, R. The vascular flora of the Royal Park of Portici (Naples, Italy). Webbia 2009, 64, 235–266. [Google Scholar] [CrossRef]
- Rossi, S.; Anfodillo, T.; Menardi, R. Trephor: A new tool for sampling microcores from tree stems. IAWA J. 2006, 27, 89–97. [Google Scholar] [CrossRef] [Green Version]
- Van der Werf, G.W.; Sass-Klaassen, U.G.; Mohren GM, J. The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 2007, 25, 103–112. [Google Scholar] [CrossRef]
- De Luis, M.; Novak, K.; Raventós, J.; Gričar, J.; Prislan, P.; Čufar, K. Climate factors promoting intra-annual density fluctuations in Aleppo pine (Pinus halepensis) from semiarid sites. Dendrochronologia 2011, 29, 163–169. [Google Scholar] [CrossRef]
- Rossi, S.; Deslauriers, A.; Anfodillo, T.; Morin, H.; Saracino, A.; Motta, R.; Borghetti, M. Conifers in cold environments synchronize maximum growth rate of tree-ring formation with day length. New Phytol. 2006, 170, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čufar, K.; Cherubini, M.; Gričar, J.; Prislan, P.; Spina, S.; Romagnoli, M. Xylem and phloem formation in chestnut (Castanea sativa Mill.) during the 2008 growing season. Dendrochronologia 2011, 29, 127–134. [Google Scholar] [CrossRef]
- Trockenbrodt, M. Survey and discussion of the terminology used in bark anatomy. IAWA J. 1990, 11, 141–166. [Google Scholar] [CrossRef]
- Merela, M.; Thaler, N.; Balzano, A.; Plavčak, D. Optimal Surface Preparation for Wood Anatomy Research of Invasive Species by Scanning Electron Microscopy. Drv. Ind. 2020, 71, 117–127. [Google Scholar] [CrossRef]
- Allard, V.; Ourcival, J.M.; Rambal, S.; Joffre, R.; Rocheteau, A. Seasonal and annual variation of carbon exchange in an evergreen Mediterranean forest in southern France. Glob. Chang. Biol. 2008, 14, 714–725. [Google Scholar] [CrossRef]
- Montagnoli, A.; Dumroese, R.K.; Terzaghi, M.; Onelli, E.; Scippa, G.S.; Chiatante, D. Seasonality of fine root dynamics and activity of root and shoot vascular cambium in a Quercus ilex L. forest (Italy). For. Ecol. Manag. 2019, 431, 26–34. [Google Scholar] [CrossRef]
- Zalloni, E.; Battipaglia, G.; Cherubini, P.; Saurer, M.; De Micco, V. Wood growth in pure and mixed Quercus ilex L. forests: Drought influence depends on site conditions. Front. Plant Sci. 2019, 10, 397. [Google Scholar] [CrossRef]
- Güney, A.; Kerr, D.; Sökücü, A.; Zimmermann, R.; Küppers, M. Cambial activity and xylogenesis in stems of Cedrus libani A. Rich at different altitudes. Bot. Stud. 2015, 56, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalloni, E.; de Luis, M.; Campelo, F.; Novak, K.; De Micco, V.; Di Filippo, A.; Vieira, J.; Nabais, C.; Rozas, V.; Battipaglia, G. Climatic signals from intra-annual density fluctuation frequency in Mediterranean pines at a regional scale. Front. Plant Sci. 2016, 7, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worbes, M. Wood anatomy and tree-ring structure and their importance for tropical dendrochronology. In Amazonian Floodplain Forests; Springer: Dordrecht, The Netherlands, 2010; pp. 329–346. [Google Scholar] [CrossRef]
- De Micco, V.; Balzano, A.; Wheeler, E.A.; Baas, P. Tyloses and gums: A review of structure, function and occurrence of vessel occlusions. IAWA J. 2016, 37, 186–205. [Google Scholar] [CrossRef] [Green Version]
- Oven, P.; Merela, M.; Mikac, U.; Serša, I. 3D magnetic resonance microscopy of a wounded beech branch. Holzforschung 2008, 62, 322–328. [Google Scholar] [CrossRef]
- Oven, P.; Merela, M.; Mikac, U.; Serša, I. Application of 3D magnetic resonance microscopy to the anatomy of woody tissues. IAWA J. 2011, 32, 401–414. [Google Scholar] [CrossRef] [Green Version]
- Savi, T.; Casolo, V.; Luglio, J.; Bertuzzi, S.; Gullo, M.A.L.; Nardini, A. Species-specific reversal of stem xylem embolism after a prolonged drought correlates to endpoint concentration of soluble sugars. Plant Physiol. Biochem. 2016, 106, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Pagliarani, C.; Casolo, V.; Ashofteh Beiragi, M.; Cavalletto, S.; Siciliano, I.; Schubert, A.; Gullino, M.L.; Zwieniecki, M.A.; Secchi, F. Priming xylem for stress recovery depends on coordinated activity of sugar metabolic pathways and changes in xylem sap pH. Plant Cell Environ. 2019, 42, 1775–1787. [Google Scholar] [CrossRef] [Green Version]
- Dwyer, J.P.; Cutter, B.E.; Wetteroff, J.J. A dendrochronological study of black and scarlet oak decline in the Missouri Ozarks. For. Ecol. Manag. 1995, 75, 69–75. [Google Scholar] [CrossRef]
- Voelker, S.L.; Muzika, R.M.; Guyette, R.P. Individual tree and stand level influences on the growth, vigor, and decline of red oaks in the Ozarks. For. Sci. 2008, 54, 8–20. [Google Scholar] [CrossRef]
Figure 1.
Climatic diagram of monthly mean temperatures (T, red line) and total precipitation (P, blue bars) for 2017 according to data recorded at the Sant’Anastasia meteorological station.
Figure 1.
Climatic diagram of monthly mean temperatures (T, red line) and total precipitation (P, blue bars) for 2017 according to data recorded at the Sant’Anastasia meteorological station.

Figure 2.
Cross section of Quercus ilex. (A) Non-collapsed phloem, xylem, and cambial zone; the pink arrow indicates the cambial cells (CC), the yellow arrow points towards the group of phloem fibres (PF). (B) The pink arrows indicate the CC and post-cambial cells (PC), the yellow arrow shows the developing phloem fibre (PF). (C) The same section zone as picture (B) but viewed under polarised light. Scale bars = 100 µm.
Figure 2.
Cross section of Quercus ilex. (A) Non-collapsed phloem, xylem, and cambial zone; the pink arrow indicates the cambial cells (CC), the yellow arrow points towards the group of phloem fibres (PF). (B) The pink arrows indicate the CC and post-cambial cells (PC), the yellow arrow shows the developing phloem fibre (PF). (C) The same section zone as picture (B) but viewed under polarised light. Scale bars = 100 µm.

Figure 3.
Cross-section of microcores from three different Q. ilex trees showing (A) tree-ring structure without clear latewood (yellow arrows), (B) multiseriate ray (green arrow), and (C) wedging ring (yellow arrows) and tyloses (red arrows). All images are of the same magnification; scale bar = 500 µm.
Figure 3.
Cross-section of microcores from three different Q. ilex trees showing (A) tree-ring structure without clear latewood (yellow arrows), (B) multiseriate ray (green arrow), and (C) wedging ring (yellow arrows) and tyloses (red arrows). All images are of the same magnification; scale bar = 500 µm.

Figure 4.
Cross-section of Q. ilex wood: (A) vessels with tyloses (arrow) in the last formed xylem increment below the cambium (CC) and non-collapsed phloem (NPh), and (B) detailed view of a vessel with tyloses from (A), and visible starch grains in parenchyma cells; (C) the beginning of tylosis formation with proliferation of a parenchyma cell into a vessel lumen through the vessel pit.
Figure 4.
Cross-section of Q. ilex wood: (A) vessels with tyloses (arrow) in the last formed xylem increment below the cambium (CC) and non-collapsed phloem (NPh), and (B) detailed view of a vessel with tyloses from (A), and visible starch grains in parenchyma cells; (C) the beginning of tylosis formation with proliferation of a parenchyma cell into a vessel lumen through the vessel pit.

Figure 5.
The graph shows the number of cambial cells (CC, blue line) and phloem increments distance (PI, violet line) between CC and the nearest phloem fibres cluster (PF) in Q. ilex. The scheme below the X-axis indicates the occurrence timing of new formed PF groups (PF1, PF2, PF3 in violet square) and PC (cells with primary wall in blue square) in different periods of the year. X× = no PF or PC (post-cambial) cells detected. DOY, day of the year from January to December 2017.
Figure 5.
The graph shows the number of cambial cells (CC, blue line) and phloem increments distance (PI, violet line) between CC and the nearest phloem fibres cluster (PF) in Q. ilex. The scheme below the X-axis indicates the occurrence timing of new formed PF groups (PF1, PF2, PF3 in violet square) and PC (cells with primary wall in blue square) in different periods of the year. X× = no PF or PC (post-cambial) cells detected. DOY, day of the year from January to December 2017.

Figure 6.
Q. ilex cross-sections with (A) development of PC cells stained with astra blue in October; (B) the same section region as in (A) but observed in polarised light; (C) formation of developing phloem fibres PF stained with safranin and astra blue in November; (D) the same wood region as in (C) but under polarised light. (E) Total wood increment (yellow dashes) with IADF (red dashes) formed during the year 2017. Scale bars = 100 µm.
Figure 6.
Q. ilex cross-sections with (A) development of PC cells stained with astra blue in October; (B) the same section region as in (A) but observed in polarised light; (C) formation of developing phloem fibres PF stained with safranin and astra blue in November; (D) the same wood region as in (C) but under polarised light. (E) Total wood increment (yellow dashes) with IADF (red dashes) formed during the year 2017. Scale bars = 100 µm.

Figure 7.
Wood, cambium, and phloem with fibre groups (PF) in January, April, May, October, November, and December in tree number 7. PF1, PF2, and PF3 serve as an orientation to evaluate the amount of phloem formed from January by the sampling date (white arrows with double heads). The sample taken in December (right image) contains non-collapsed phloem (NPh) with three groups of fibres which corresponds with the amount of phloem formed in one calendar year, 2017; the phloem formed in the previous year is already collapsed (CPh). Scale bar = 100 µm.
Figure 7.
Wood, cambium, and phloem with fibre groups (PF) in January, April, May, October, November, and December in tree number 7. PF1, PF2, and PF3 serve as an orientation to evaluate the amount of phloem formed from January by the sampling date (white arrows with double heads). The sample taken in December (right image) contains non-collapsed phloem (NPh) with three groups of fibres which corresponds with the amount of phloem formed in one calendar year, 2017; the phloem formed in the previous year is already collapsed (CPh). Scale bar = 100 µm.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Width of total growth increments in the wood and non-collapsed phloem, their ratio, and occurrence of IADFs on the last sampling date at the end of 2017.
Table 1.
Width of total growth increments in the wood and non-collapsed phloem, their ratio, and occurrence of IADFs on the last sampling date at the end of 2017.
| Tree | Total Xylem | Non-Collapsed | Phloem/Xylem | IADF |
|---|---|---|---|---|
| Increment | Phloem Increment | Increment Ratio | Occurrence | |
| Number | µm | µm | Number | |
| 1 | 808.9 | 314.4 | 0.389 | 1 |
| 2 | 682.0 | 260.0 | 0.381 | 1 |
| 3 | 188.0 | 230.0 | 1.223 | 1 |
| 4 | 0.0 | 204.3 | -- | - |
| 5 | 0.0 | 408.0 | -- | - |
| 6 | 682.0 | 394.0 | 0.578 | 1 |
| 7 | 632.0 | 420.0 | 0.665 | 1 |
| 8 | 803.0 | 362.0 | 0.451 | 1 |
| Average | 474.5 | 324.1 | 0.614 | 0.4 |
| STD | 351.1 | 84.5 | 0.318 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Balzano, A.; Čufar, K.; De Micco, V. Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy. Forests 2021, 12, 188. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020188
AMA Style
Balzano A, Čufar K, De Micco V. Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy. Forests. 2021; 12(2):188. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020188
Chicago/Turabian StyleBalzano, Angela, Katarina Čufar, and Veronica De Micco. 2021. "Xylem and Phloem Formation Dynamics in Quercus ilex L. at a Dry Site in Southern Italy" Forests 12, no. 2: 188. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020188
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.