
Light Spectra during Somatic Embryogenesis of Norway Spruce—Impact on Growth, Embryo Productivity, and Embling Survival


Abstract
:1. Introduction
2. Materials and Methods
2.1. Origin of Embryogenic Lines
2.2. SE Development, Germination, and Plant Production
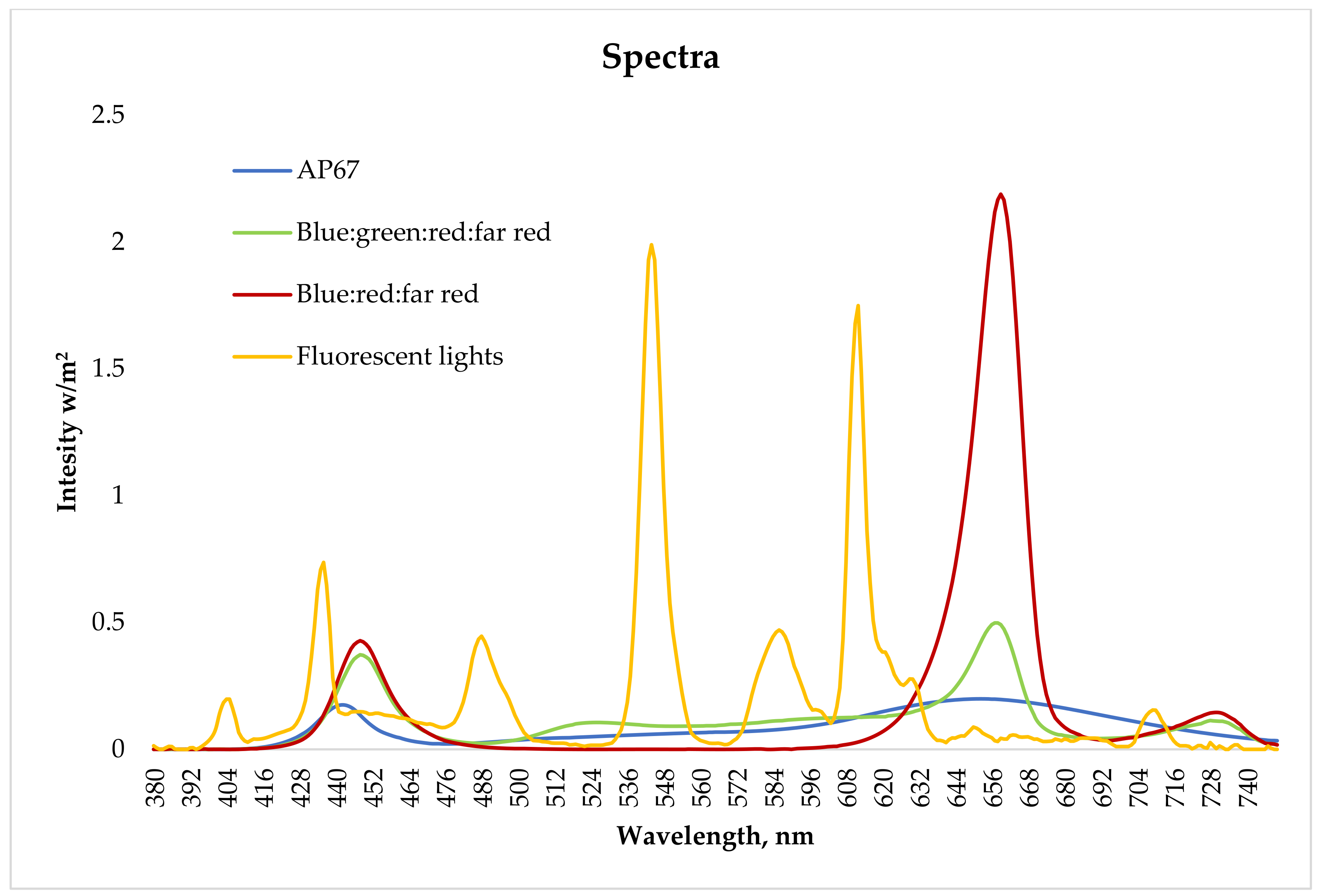
2.3. Experiment I: Proliferation in Different Spectra
2.4. Experiment II: Maturation of Embryos in Different Spectra
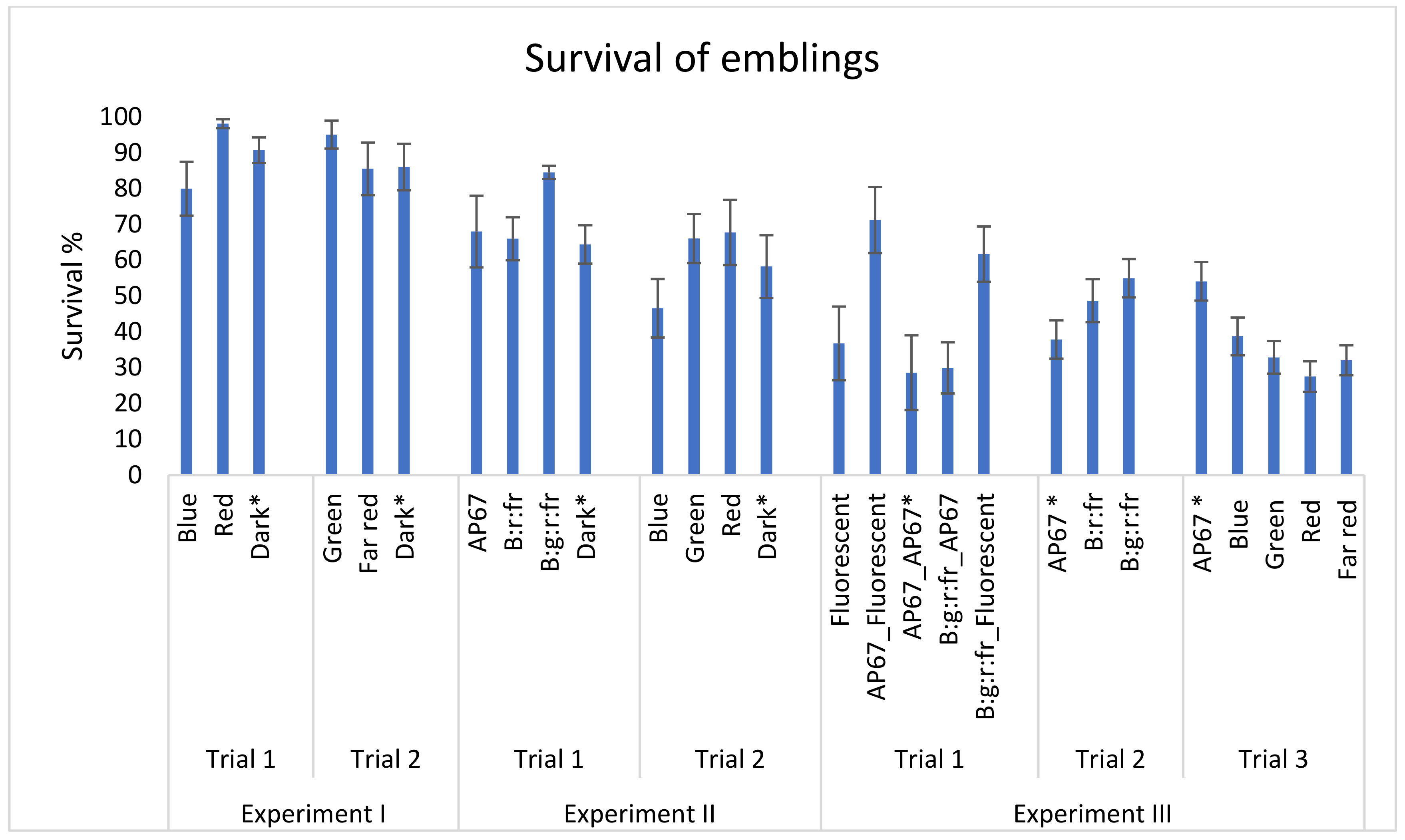
2.5. Experiment III: Germination of Embryos in Different Spectra
2.6. Measurements and Data Analysis
3. Results
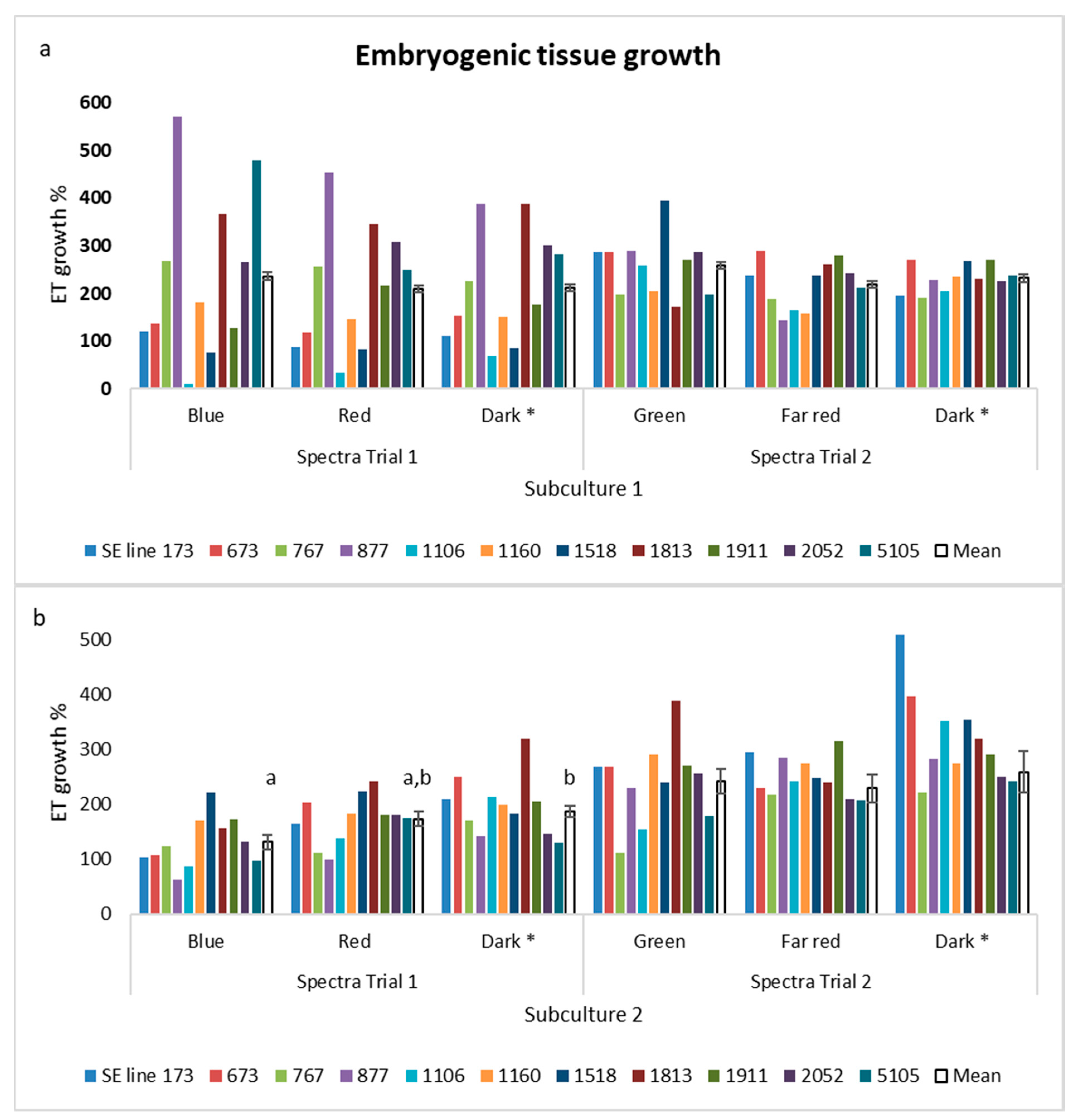
3.1. Experiment I: Proliferation in Different Spectra
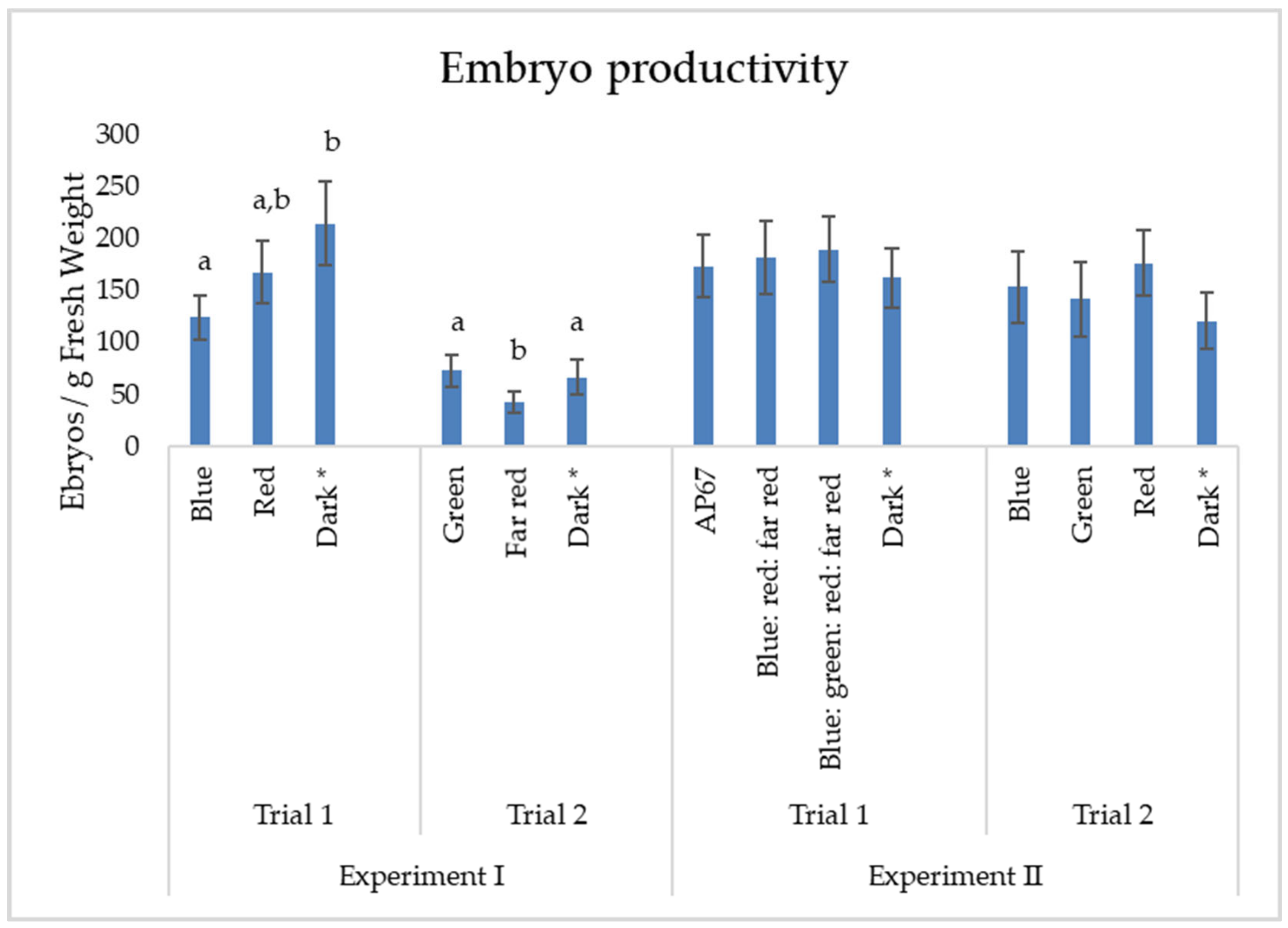
3.2. Experiment II: Maturation of Embryos in Different Light Spectra
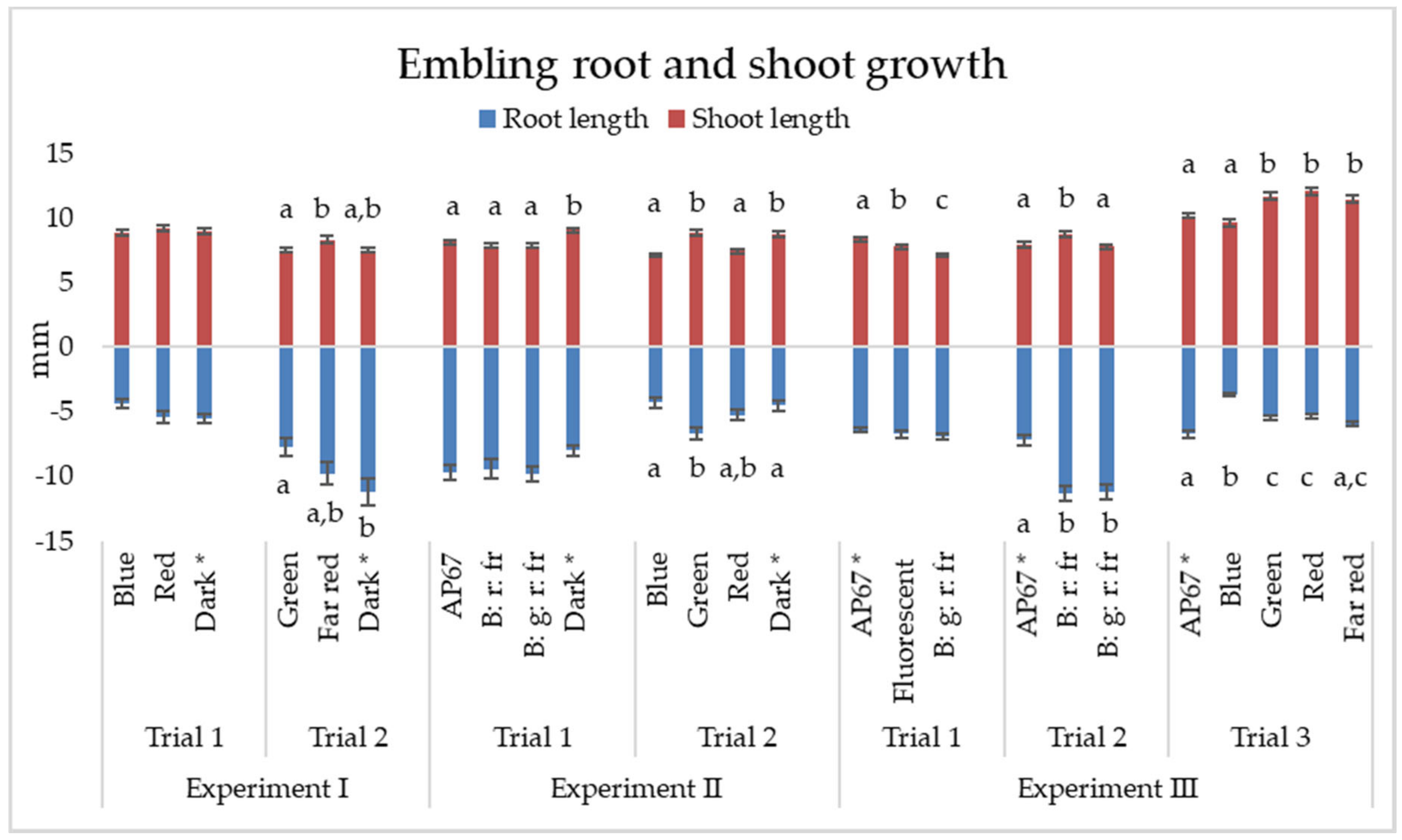
3.3. Experiment III: Germination of Embryos in Different Light Spectra
4. Discussion
4.1. The Effect of Light on the Proliferation of ET and Maturation of Somatic Embryos
4.2. Embryo Germination and Embling Growth under Different Lights
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Haapanen, M. Performance of Genetically Improved Norway Spruce in One-Third Rotation-Aged Progeny Trials in Southern Finland. Scand. J. For. Res. 2020, 35, 1–6. [Google Scholar] [CrossRef]
- Sutton, B. Commercial Delivery of Genetic Improvement to Conifer Plantations using Somatic Embryogenesis. Ann. For. Sci. 2002, 59, 657–661. [Google Scholar] [CrossRef]
- Lelu-Walter, M.; Thompson, D.; Harvengt, L.; Sanchez, L.; Toribio, M.; Pâques, L.E. Somatic Embryogenesis in Forestry with a Focus on Europe: State-of-the-Art, Benefits, Challenges and Future Direction. Tree Genet. Genomes 2013, 9, 883–899. [Google Scholar] [CrossRef]
- Högberg, K.; Varis, S. Vegetative Propagation of Norway Spruce: Experiences and Present Situation in Sweden and Finland. In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 538–550. [Google Scholar]
- Adams, G.W.; Kunze, H.A.; McCartney, A.; Millican, S.; Park, Y.S. An Industrial Perspective on the use of Advanced Reforestation Stock Technologies. In Vegetative Propagation of Forest Trees; Park, Y.-S., Bonga, J.M., Moon, H.-K., Eds.; National Institute of Forest Science: Seoul, Korea, 2016; pp. 323–334. [Google Scholar]
- Chalupa, V. Somatic Embryogenesis and Plantlet Regeneration from Cultured Immature and Mature Embryos of Picea Abies (L.). Karst. Commun. Inst. For. Cech. 1985, 14, 57–63. [Google Scholar]
- Hakman, I.; Von Arnold, S. Plantlet Regeneration through Somatic Embryogenesis in Picea Abies (Norway Spruce). J. Plant Physiol. 1985, 121, 149–158. [Google Scholar] [CrossRef]
- Jain, S.M.; Newton, R.J.; Soltes, E.J. Enhancement of Somatic Embryogenesis in Norway Spruce (Picea Abies L.). Theor. Appl. Genet. 1988, 76, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Vagner, M.; Fischerova, L.; Spackova, J.; Vondrakova, Z. Somatic embryogenesis in Norway spruce. In Protocol for Somatic Embryogenesis in Woody Plant; Jain, S.M., Gupta, P., Eds.; Springer: Dordrecht, The Netherlands, 2005; Volume 77, pp. 141–155. [Google Scholar]
- Varis, S. Norway spruce Picea abies (L.) Karst. In Step Wise Protocols for Somatic Embryogenesis of Important Woody Plants; Jain, S.M., Gupta, P., Eds.; Springer: Dordrecht, The Netherlands, 2018; Volume 84, pp. 255–268. [Google Scholar] [CrossRef]
- Tikkinen, M. Improved propagation efficiency in a laboratory–nursery interface for somatic embryogenesis in Norway spruce. Diss. For. 2018, 265, 35. [Google Scholar] [CrossRef]
- Thompson, D. Challenges for the Large-Scale Propagation of Forest Trees by Somatic Embryogenesis–a Review. In Proceedings of the Third International Conference of the IUFRO Unit 2.09.02 on Woody Plant Production Integrating Genetic and Vegetative Propagation Technologies, Vitoria-Gasteiz, Spain, 18–12 September 2014; pp. 81–91. [Google Scholar]
- Singh, H. Embryology of Gymnosperms; Gebrüder Borntraeger: Berlin, Germany, 1978. [Google Scholar]
- Contreras, S.; Bennett, M.A.; Metzger, J.D.; Tay, D. Maternal Light Environment during Seed Development Affects Lettuce Seed Weight, Germinability, and Storability. HortScience 2008, 43, 845–852. [Google Scholar] [CrossRef] [Green Version]
- von Arnold, S.; Hakman, I. Plantlet regeneration in vitro via adventitious buds and somatic embryos in Norway spruce (Picea abies). In Genetic Manipulation of Woody Plants; Hanover, J.W., Keatly, D.E., Eds.; Springer: Boston, MA, USA, 1988; Volume 44, pp. 199–215. [Google Scholar]
- Kvaalen, H.; Appelgren, M. Light Quality Influences Germination, Root Growth and Hypocotyl Elongation in Somatic Embryos but Not in Seedlings of Norway Spruce. Vitr. Cell. Dev. Biol. Plant 1999, 35, 437–441. [Google Scholar] [CrossRef]
- Merkle, S.A.; Montello, P.M.; Xia, X.; Upchurch, B.L.; Smith, D.R. Light Quality Treatments Enhance Somatic Seedling Production in Three Southern Pine Species. Tree Physiol. 2005, 26, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riikonen, J.; Kettunen, N.; Gritsevich, M.; Hakala, T.; Särkkä, L.; Tahvonen, R. Growth and Development of Norway Spruce and Scots Pine Seedlings Under Different Light Spectra. Environ. Exp. Bot. 2016, 121, 112–120. [Google Scholar] [CrossRef]
- Fløistad, I.S.; Patil, G.G. Growth and Terminal Bud Formation in Picea Abies Seedlings Grown with Alternating Diurnal Temperature and Different Light Qualities. Scand. J. For. Res. 2002, 17, 15–27. [Google Scholar] [CrossRef]
- Latkowska, M.J.; Kvaalen, H.; Appelgren, M. Genotype Dependent Blue and Red Light Inhibition of the Proliferation of the Embryogenic Tissue of Norway Spruce. Vitr. Cell. Dev. Biol. Plant 2000, 36, 57–60. [Google Scholar] [CrossRef]
- Hakman, I.; Fowke, L.C.; Von Arnold, S.; Eriksson, T. The Development of Somatic Embryos in Tissue Cultures Initiated from Immature Embryos of Picea abies (Norway Spruce). Plant Sci. 1985, 38, 53–59. [Google Scholar] [CrossRef]
- Loo, K.H.; Lun, W.; Tan, S.; Lai, Y.M.; Chi, K.T. On Driving Techniques for LEDs: Toward a Generalized Methodology. IEEE Trans. Power Electron. 2009, 24, 2967–2976. [Google Scholar] [CrossRef]
- Shimada, A.; Taniguchi, Y. Red and Blue Pulse Timing Control for Pulse Width Modulation Light Dimming of Light Emitting Diodes for Plant Cultivation. J. Photochem. Photobiol. B Biol. 2011, 104, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Klimaszewska, K.; Lachance, D.; Pelletier, G.; Lelu, M.; Séguin, A. Regeneration of Transgenic Picea Glauca, P. Mariana, and P. Abies After Cocultivation of Embryogenic Tissue with Agrobacterium Tumefaciens. Vitr. Cell. Dev. Biol. Plant 2001, 37, 748–755. [Google Scholar] [CrossRef]
- Varis, S.; Ahola, S.; Jaakola, L.; Aronen, T. Reliable and Practical Methods for Cryopreservation of Embryogenic Cultures and Cold Storage of Somatic Embryos of Norway Spruce. Cryobiology 2017, 76, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Litvay, J.D.; Verma, D.C.; Johnson, M.A. Influence of a Loblolly Pine (Pinus Taeda L.). Culture Medium and its Components on Growth and Somatic Embryogenesis of the Wild Carrot (Daucus Carota L.). Plant Cell Rep. 1985, 4, 325–328. [Google Scholar] [CrossRef]
- Tikkinen, M.; Varis, S.; Aronen, T. Development of Somatic Embryo Maturation and Growing Techniques of Norway Spruce Emblings Towards Large-Scale Field Testing. Forests 2018, 9, 325. [Google Scholar] [CrossRef] [Green Version]
- Tikkinen, M.; Varis, S.; Peltola, H.; Aronen, T. Improved Germination Conditions for Norway Spruce Somatic Cotyledonary Embryos Increased Survival and Height Growth of Emblings. Trees 2018, 32, 1489–1504. [Google Scholar] [CrossRef]
- Aphalo, P.J. R Packages for Photobiology. 2015. Available online: http://www.r4photobiology.info/ (accessed on 4 March 2021).
- Svobodová, H.; Albrechtová, J.; Kumstýřová, L.; Lipavská, H.; Vágner, M.; Vondráková, Z. Somatic Embryogenesis in Norway Spruce: Anatomical Study of Embryo Development and Influence of Polyethylene Glycol on Maturation Process. Plant Physiol. Biochem. 1999, 37, 209–221. [Google Scholar] [CrossRef]
- Von Aderkas, P.; Teyssier, C.; Charpentier, J.; Gutmann, M.; Pâques, L.; Le Metté, C.; Ader, K.; Label, P.; Kong, L.; Lelu-Walter, M. Effect of Light Conditions on Anatomical and Biochemical Aspects of Somatic and Zygotic Embryos of Hybrid Larch (Larix× Marschlinsii). Ann. Bot. 2015, 115, 605–615. [Google Scholar] [CrossRef]
- Folta, K.M.; Maruhnich, S.A. Green Light: A Signal to Slow Down Or Stop. J. Exp. Bot. 2007, 58, 3099–3111. [Google Scholar] [CrossRef] [PubMed]
- Johnsen, Ø.; Dæhlen, O.G.; Østreng, G.; Skrøppa, T. Daylength and Temperature during Seed Production Interactively Affect Adaptive Performance of Picea Abies Progenies. New Phytol. 2005, 168, 589–596. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.; Ribeyre, Z.; Vigneaud, J.; Sow, M.D.; Fichot, R.; Messier, C.; Pinto, G.; Nolet, P.; Maury, S. Advances and promises of epigenetics for forest trees. Forests 2020, 11, 976. [Google Scholar] [CrossRef]
- Breton, D.; Harvengt, L.; Trontin, J.-F.; Bouvet, A.; Favre, J.-M. Long-term subculture randomly affects morphology and subsequent maturation of early somatic embryos in maritime pine. Plant Cell Tiss. Organ Cult. 2006, 87, 95–108. [Google Scholar] [CrossRef]
- Klimaszewska, K.; Noceda, C.; Pelletier, G.; Label, P.; Rodriguez, R.; Lelu-Walter, M.A. Biological characterization of young and aged embryogenic cultures of Pinus pinaster (Ait.). Vitr. Cell. Dev. Biol. Plant 2009, 45, 20–33. [Google Scholar] [CrossRef]
- von Arnold, S.; Hakman, I. Regulation of Somatic Embryo Development in Picea Abies by Abscisic Acid (ABA). J. Plant Physiol. 1988, 132, 164–169. [Google Scholar] [CrossRef]
- Senol, R.; Kilic, S.; Tasdelen, K. Pulse Timing Control for LED Plant Growth Unit and Effects on Carnation. Comput. Electron. Agric. 2016, 123, 125–134. [Google Scholar] [CrossRef]
- Hernandez Velasco, M.; Mattsson, A. Light Quality and Intensity of Light-Emitting Diodes during Pre-Cultivation of Picea Abies (L.) Karst. and Pinus Sylvestris L. Seedlings–impact on Growth Performance, Seedling Quality and Energy Consumption. Scand. J. For. Res. 2019, 34, 159–177. [Google Scholar] [CrossRef] [Green Version]
- Landis, T.; Dumroese, R.; Haase, D. The Container Tree Nursery Manual: Seedling Processing, Storage, and Outplanting; U.S. Department of Agriculture, Forest Service: Washington, DC, USA, 2010; Volume 7, p. 200.
- Rikala, R. Metsäpuiden Paakkutaimien Kasvatusopas (Manual for Container Seedlings of Forest Trees); Metsäntutkimuslaitos (Finnish Forest Research Institute): Suonenjoki, Finland, 2012; p. 247. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Trial | Proliferation Spectrum | Maturation Spectrum | Germination Spectrum | Growth Spectrum |
|---|---|---|---|---|---|
| Experiment I | 1 | Blue | Dark | AP67 1 | AP67 |
| Red | Dark | AP67 1 | AP67 | ||
| Dark | Dark | AP67 1 | AP67 | ||
| 2 | Green | Dark | AP67 1 | AP67 | |
| Far red | Dark | AP67 1 | AP67 | ||
| Dark | Dark | AP67 1 | AP67 | ||
| Experiment II | 1 | Dark | AP67 | AP67 1 | AP67 |
| Dark | Blue, red, far red | AP67 1 | AP67 | ||
| Dark | Blue, green, red, far red | AP67 1 | AP67 | ||
| Dark | Dark | AP67 1 | AP67 | ||
| 2 | Dark | Blue | AP67 1 | AP67 | |
| Dark | Green | AP67 1 | AP67 | ||
| Dark | Red | AP67 1 | AP67 | ||
| Dark | Dark | AP67 1 | AP67 | ||
| Experiment III | 1 | Dark | Dark | Fluorescent 2 | Fluorescent |
| Dark | Dark | AP67 3 | Fluorescent | ||
| Dark | Dark | AP67 3 | AP67 | ||
| Dark | Dark | Blue, green, red, far red 3 | Fluorescent | ||
| Dark | Dark | Blue, green, red, far red 3 | AP67 | ||
| 2 | Dark | Dark | AP67 4 | AP67 | |
| Dark | Dark | Blue, red, far red 4 | AP67 | ||
| Dark | Dark | Blue, green, red, far red 4 | AP67 | ||
| 3 | Dark | Dark | AP67 4 | AP67 | |
| Dark | Dark | Blue 4 | AP67 | ||
| Dark | Dark | Green 4 | AP67 | ||
| Dark | Dark | Red 4 | AP67 | ||
| Dark | Dark | Far red 4 | AP67 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varis, S.; Tikkinen, M.; Välimäki, S.; Aronen, T. Light Spectra during Somatic Embryogenesis of Norway Spruce—Impact on Growth, Embryo Productivity, and Embling Survival. Forests 2021, 12, 301. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030301
Varis S, Tikkinen M, Välimäki S, Aronen T. Light Spectra during Somatic Embryogenesis of Norway Spruce—Impact on Growth, Embryo Productivity, and Embling Survival. Forests. 2021; 12(3):301. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030301
Chicago/Turabian StyleVaris, Saila, Mikko Tikkinen, Sakari Välimäki, and Tuija Aronen. 2021. "Light Spectra during Somatic Embryogenesis of Norway Spruce—Impact on Growth, Embryo Productivity, and Embling Survival" Forests 12, no. 3: 301. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030301