
Effects of Throughfall Exclusion on Photosynthetic Traits in Mature Japanese Cedar (Cryptomeria japonica (L. f.) D. Don.)

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
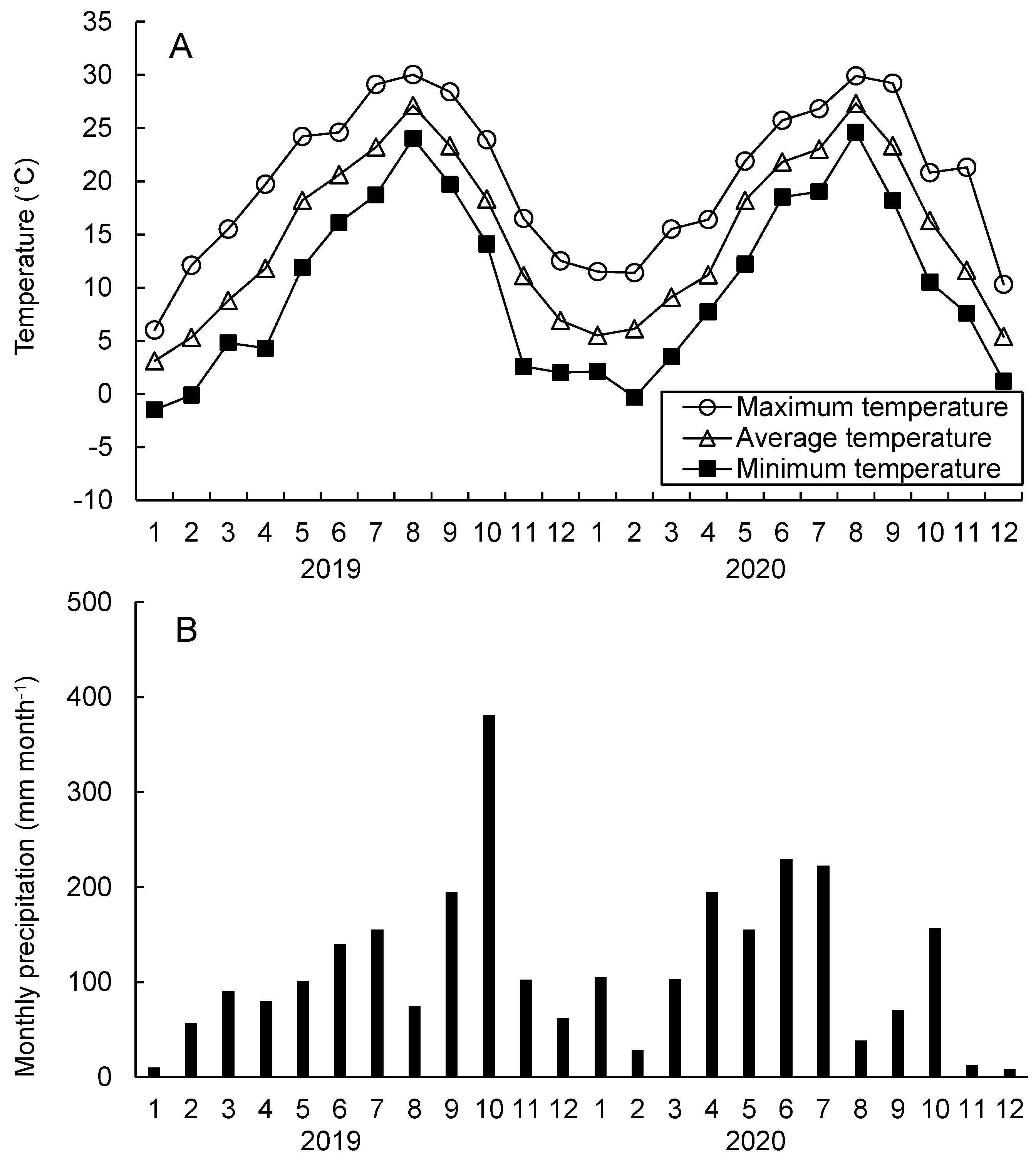
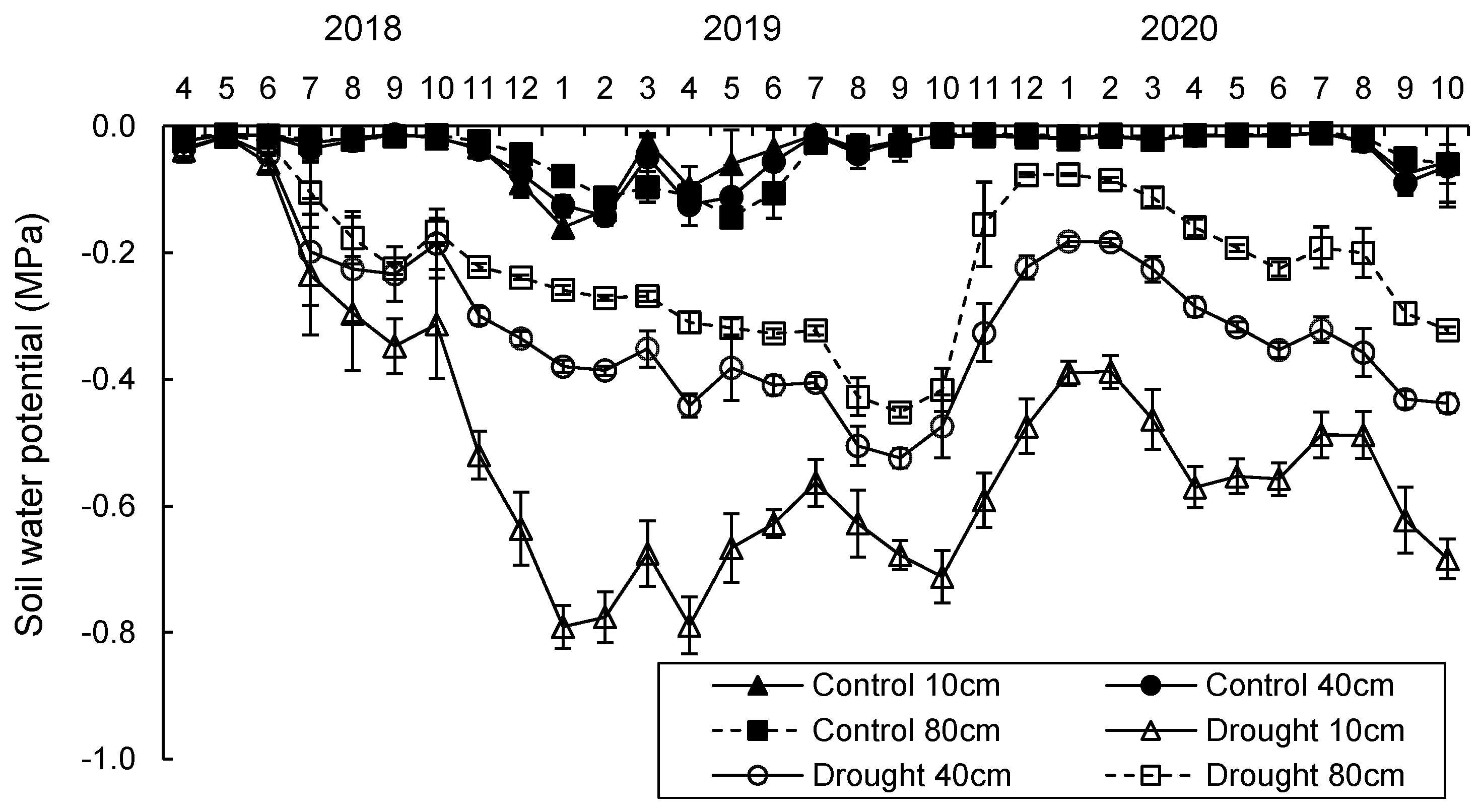
2.1. Study Site and Climate Conditions
2.2. Plot Establishment and Throughfall Exclusion
2.3. Foliar Ecophysiological Measurements
2.4. Foliar area and Nitrogen Measurements
2.5. Statistical Analyses
3. Results
3.1. Temperature, Precipitation, and Soil Water Potential
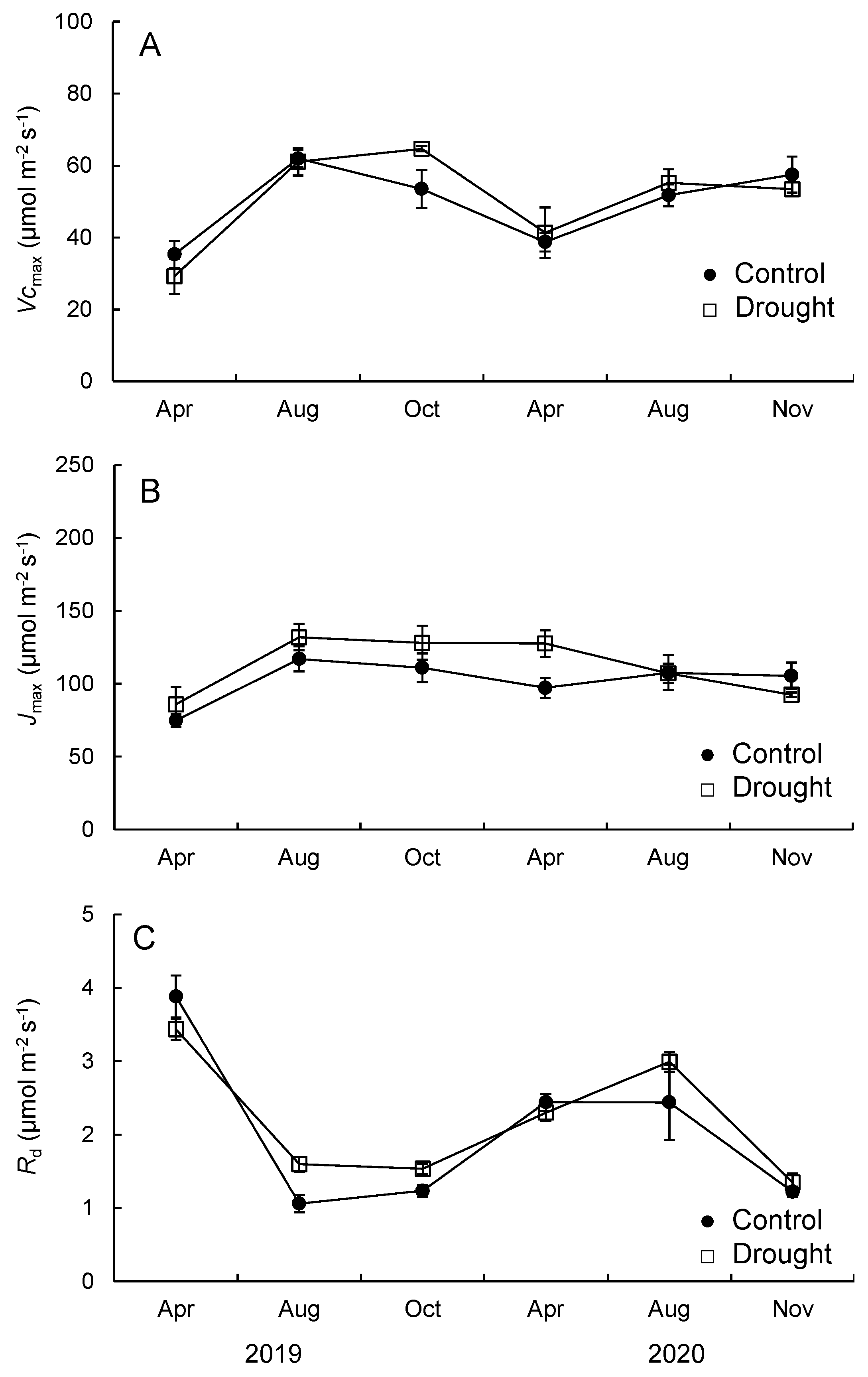
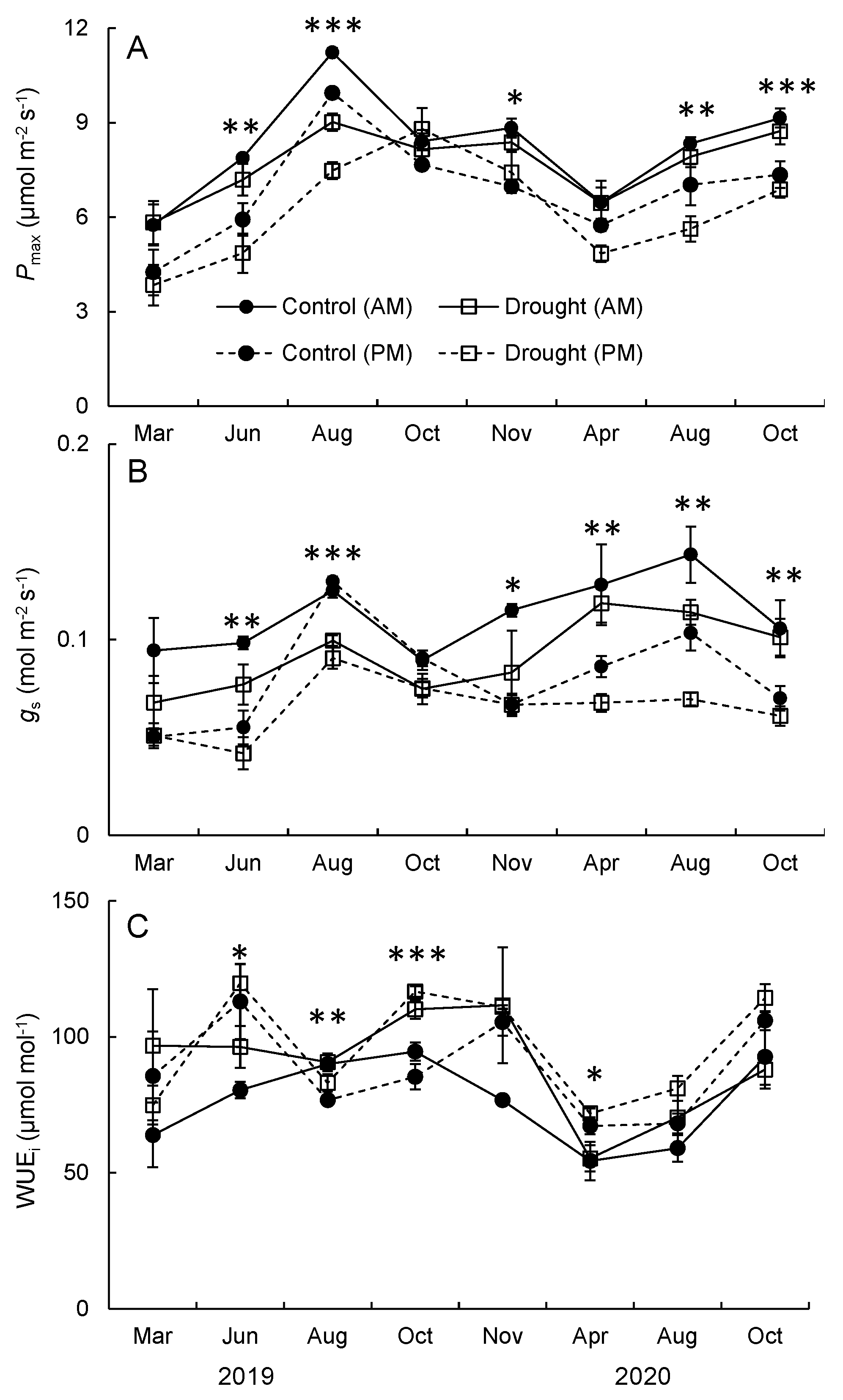
3.2. Photosynthetic Capacity, Stomatal Conductance, and Water Use Efficiency
3.3. LMA and Nitrogen Content
3.4. Foliar Traits and Pmax
4. Discussion
4.1. Photosynthetic Capacity and Dark Respiration Rate
4.2. Effects of Leaf Morphology and Nitrogen Concentration
4.3. Foliar Gas Exchange and Drought Response
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Han, A.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Matsumoto, Y.; Shigenaga, H.; Miura, S.; Nagakura, J.; Taoda, H. Mapping of Japanese cedar (Cryptomeria japonica) forests vulnerable to global warming in Japan. Glob. Environ. Res. 2006, 10, 181–188. [Google Scholar]
- Lambers, H.; Chapin, F.S., III; Pons, T.L. Plant Physiological Ecology; Springer: New York, NY, USA, 1998. [Google Scholar]
- Niinemets, Ü. Responses of forest trees to single and multiple environmental stresses from seedlings to mature plants: Past stress history, stress interactions, tolerance and acclimation. For. Ecol. Manag. 2010, 260, 1623–1639. [Google Scholar] [CrossRef]
- Misson, L.; Limousin, J.M.; Rodriguez, R.; Letts, M.G. Leaf physiological responses to extreme droughts in Mediterranean Quercus ilex forest. Plant Cell Environ. 2010, 33, 1898–1910. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Guo, W.; Liu, J.; Du, N.; Wang, R. Increased nitrogen deposition alleviated the adverse effects of drought stress on Quercus variabilis and Quercus mongolica seedlings. Acta Physiol. Plant. 2015, 37, 107. [Google Scholar] [CrossRef]
- Maggard, A.O.; Will, R.E.; Wilson, D.S.; Meek, C.R.; Vogel, J.G. Fertilization reduced stomatal conductance but not photosynthesis of Pinus taeda which compensated for lower water availability in regards to growth. For. Ecol. Manag. 2016, 381, 37–47. [Google Scholar] [CrossRef] [Green Version]
- Ogaya, R.; Peñuelas, J. Comparative field study of Quercus ilex and Phillyrea latifolia: Photosynthetic response to experimental drought conditions. Environ. Exp. Bot. 2003, 50, 137–148. [Google Scholar] [CrossRef]
- Ambrose, A.R.; Baxter, W.L.; Wong, C.S.; Næsborg, R.R.; Williams, C.B.; Dawson, T.E. Contrasting drought-response strategies in California redwoods. Tree Physiol. 2015, 35, 453–469. [Google Scholar] [CrossRef] [Green Version]
- Inoue, Y.; Ichie, T.; Kenzo, T.; Yoneyama, A.; Kumagai, T.O.; Nakashizuka, T. Effects of rainfall exclusion on leaf gas exchange traits and osmotic adjustment in mature canopy trees of Dryobalanops aromatica (Dipterocarpaceae) in a Malaysian tropical rain forest. Tree Physiol. 2017, 37, 1301–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nepstad, D.C.; Moutinho, P.; Dias-Filho, M.B.; Davidson, E.; Cardinot, G.; Markewitz, D.; Schwalbe, K. The effects of partial throughfall exclusion on canopy processes, aboveground production, and biogeochemistry of an Amazon forest. J. Geophys. Res. Atmos. 2002, 107, LBA-53. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; O’Grady, A.P.; Duursma, R.A.; Choat, B.; Huang, G.; Smith, R.A.; Jiang, Y.; Tissue, D.T. Drought responses of two gymnosperm species with contrasting stomatal regulation strategies under elevated [CO2] and temperature. Tree Physiol. 2015, 35, 756–770. [Google Scholar] [CrossRef] [Green Version]
- Limousin, J.M.; Bickford, C.P.; Dickman, L.T.; Pangle, R.E.; Hudson, P.J.; Boutz, A.L.; Gehres, N.; Osuna, J.L.; Pockman, W.T.; McDowell, N.G. Regulation and acclimation of leaf gas exchange in a piñon–juniper woodland exposed to three different precipitation regimes. Plant Cell Environ. 2013, 36, 1812–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenzo, T.; Ichie, T.; Watanabe, Y.; Yoneda, R.; Ninomiya, I.; Koike, T. Changes in photosynthesis and leaf characteristics with tree height in five dipterocarp species in a tropical rain forest. Tree Physiol. 2006, 26, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Tange, T. Ecophysiological study on the growth of Cryptomeria japonica planted trees. Bull. Tokyo Univ. For. 1995, 93, 65–145. [Google Scholar]
- Osone, Y.; Hashimoto, S.; Kenzo, T.; Araki, M.G.; Inoue, Y.; Shichi, K.; Toriyama, J.; Yamashita, N.; Tsuruta, K.; Ishizuka, S.; et al. Plant trait database for Cryptomeria japonica and Chamaecyparis obtusa (SugiHinoki DB): Their physiology, morphology, anatomy and biochemistry. Ecol. Res. 2020, 35, 274–275. [Google Scholar] [CrossRef]
- Sakaguchi, K. All of the Sugi (Cryptomeria japonica); Zenrinkyo: Tokyo, Japan, 1983. (In Japanese) [Google Scholar]
- Suzuki, E.; Tsukahara, J. Age structure and regeneration of old growth Cryptomeria japonica forests on Yakushima Island. Bot. Mag. Tokyo 1987, 100, 223–241. [Google Scholar] [CrossRef]
- Tsumura, Y.; Uchiyama, K.; Moriguchi, Y.; Kimura, M.K.; Ueno, S.; Ujino-Ihara, T. Genetic differentiation and evolutionary adaptation in Cryptomeria japonica. Genes Genomes Genet. 2014, 4, 2389–2402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoo, T. Drought resistance of some conifers at the first summer after their emergence. Bull. Tokyo Univ. For. 1956, 51, 1–108, (In Japanese with English summary). [Google Scholar]
- Negisi, K. Photosynthesis, respiration and growth in 1-year-old seedlings of Pinus densiflora, Cryptomeria japonica and Chamaecyparis obtusa. Bull. Tokyo Univ. For. 1996, 62, 1–115. [Google Scholar]
- Nagakura, J.; Shigenaga, H.; Akama, A.; Takahashi, M. Growth and transpiration of Japanese cedar (Cryptomeria japonica) and Hinoki cypress (Chamaecyparis obtusa) seedlings in response to soil water content. Tree Physiol. 2004, 24, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Sakata, M. Evaluation of possible causes for the decline of Japanese cedar (Cryptomeria japonica) based on elemental composition and δ13C of needles. Environ. Sci. Technol. 1996, 30, 2376–2381. [Google Scholar] [CrossRef]
- Shigenaga, H.; Matsumoto, Y.; Taoda, H.; Takahashi, M. The potential effect of climate change on the transpiration of Sugi (Cryptomeria japonica D. Don) plantations in Japan. J. Agric. Meteorol. 2005, 60, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Tange, T.; Someya, M.; Norisada, M.; Masumori, M. Photosynthetic limitation of similar-height Cryptomeria japonica trees growing at different rates. Photosynthetica 2013, 51, 158–160. [Google Scholar] [CrossRef]
- Tange, T.; Ge, F. Topographic factors and tree heights of aged Cryptomeria japonica plantations in the Boso Peninsula, Japan. Forests 2020, 11, 771. [Google Scholar] [CrossRef]
- Niinemets, Ü. Global-scale climatic controls of leaf dry mass per area, density, and thickness in trees and shrubs. Ecology 2001, 82, 453–469. [Google Scholar] [CrossRef]
- Ryan, M.G.; Phillips, N.; Bond, B.J. The hydraulic limitation hypothesis revisited. Plant Cell Environ. 2006, 29, 367–381. [Google Scholar] [CrossRef]
- Thomas, S.C. Photosynthetic capacity peaks at intermediate size in temperate deciduous trees. Tree Physiol. 2010, 30, 555–573. [Google Scholar] [CrossRef] [Green Version]
- Kenzo, T.; Inoue, Y.; Yoshimura, M.; Yamashita, M.; Tanaka-Oda, A.; Ichie, T. Height-related changes in leaf photosynthetic traits in diverse Bornean tropical rain forest trees. Oecologia 2015, 177, 191–202. [Google Scholar] [CrossRef]
- Ishii, H.T.; Jennings, G.M.; Sillett, S.C.; Koch, G.W. Hydrostatic constraints on morphological exploitation of light in tall Sequoia sempervirens trees. Oecologia 2008, 156, 751–763. [Google Scholar] [CrossRef] [PubMed]
- Kenzo, T.; Yoneda, R.; Sano, M.; Araki, M.; Shimizu, A.; Tanaka-Oda, A.; Chann, S. Variations in leaf photosynthetic and morphological traits with tree height in various tree species in Cambodian tropical dry evergreen forest. Jpn. Agric. Res. Q. JARQ 2012, 46, 167–180. [Google Scholar] [CrossRef] [Green Version]
- Koch, G.W.; Sillett, S.C.; Jennings, G.M.; Davis, S.D. The limits to tree height. Nature 2004, 428, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Negisi, K.; Satoo, T. Influence of soil moisture on photosynthesis and respiration of seedlings of Akamatu (Pinus densiflora Sieb. et Zucc.) and Sugi (Cryptomeria japonica D. Don.). J. Jpn. For. Soc. 1954, 36, 113–117. [Google Scholar]
- Negisi, K.; Satoo, T. Soil moisture in relation to apparent photosynthesis and respiration of Akamatu and Sugi seedlings. J. Jpn. For. Soc. 1955, 37, 100–103. [Google Scholar]
- Nagakura, J.; Kaneko, S.; Takahashi, M.; Tange, T. Nitrogen promotes water consumption in seedlings of Cryptomeria japonica but not in Chamaecyparis obtusa. For. Ecol. Manag. 2008, 255, 2533–2541. [Google Scholar] [CrossRef]
- Dietrich, L.; Hoch, G.; Kahmen, A.; Körner, C. Losing half the conductive area hardly impacts the water status of mature trees. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Leuzinger, S.; Körner, C. Tree species diversity affects canopy leaf temperatures in a mature temperate forest. Agric. For. Meteorol. 2007, 146, 29–37. [Google Scholar] [CrossRef]
- Tang, Z.; Sayer, M.A.S.; Chambers, J.L.; Barnett, J.P. Interactive effects of fertilization and throughfall exclusion on the physiological responses and whole-tree carbon uptake of mature loblolly pine. Can. J. Bot. 2004, 82, 850–861. [Google Scholar] [CrossRef] [Green Version]
- Gaul, D.; Hertel, D.; Borken, W.; Matzner, E.; Leuschner, C. Effects of experimental drought on the fine root system of mature Norway spruce. For. Ecol. Manag. 2008, 256, 1151–1159. [Google Scholar] [CrossRef]
- Limousin, J.M.; Misson, L.; Lavoir, A.V.; Martin, N.K.; Rambal, S. Do photosynthetic limitations of evergreen Quercus ilex leaves change with long-term increased drought severity? Plant Cell Environ. 2010, 33, 863–875. [Google Scholar]
- Misson, L.; Degueldre, D.; Collin, C.; Rodriguez, R.; Rocheteau, A.; Ourcival, J.-M.; Rambal, S. Phenological responses to extreme droughts in a Mediterranean forest. Glob. Chang. Biol. 2010, 17, 1036–1048. [Google Scholar] [CrossRef]
- Grams, T.E.; Hesse, B.D.; Gebhardt, T.; Weikl, F.; Rötzer, T.; Kovacs, B.; Pritsch, K. The KROOF experiment: Realization and efficacy of a recurrent drought experiment plus recovery in a beech/spruce forest. Ecosphere 2021, 12, e03399. [Google Scholar] [CrossRef]
- Rowland, L.; Lobo-do-Vale, R.L.; Christoffersen, B.O.; Melém, E.A.; Kruijt, B.; Vasconcelos, S.S.; Meir, P. After more than a decade of soil moisture deficit, tropical rainforest trees maintain photosynthetic capacity, despite increased leaf respiration. Glob. Chang. Biol. 2015, 21, 4662–4672. [Google Scholar] [CrossRef] [PubMed]
- Tomasella, M.; Beikircher, B.; Häberle, K.H.; Hesse, B.; Kallenbach, C.; Matyssek, R.; Mayr, S. Acclimation of branch and leaf hydraulics in adult Fagus sylvatica and Picea abies in a forest through-fall exclusion experiment. Tree Physiol. 2018, 38, 198–211. [Google Scholar] [CrossRef]
- Pretzsch, H.; Grams, T.; Häberle, K.H.; Pritsch, K.; Bauerle, T.; Rötzer, T. Growth and mortality of Norway spruce and European beech in monospecific and mixed-species stands under natural episodic and experimentally extended drought. Results of the KROOF throughfall exclusion experiment. Trees 2020, 34, 957–970. [Google Scholar] [CrossRef]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef]
- Chapin, F.S., III; Matson, P.A.; Vitousek, P. Principles of Terrestrial Ecosystem Ecology; Springer Science & Business Media: New York, NY, USA, 2011. [Google Scholar]
- Forest Soil Division. Classification of forest soils in Japan 1975. Bull. Gov. For. Exp. Stn. 1976, 280, 1–28, (In Japanese with English summary). [Google Scholar]
- Inoue, Y.; Kitaoka, K.; Araki, M.; Kenzo, T.; Saito, S. Seasonal changes in leaf water potential, photosynthetic and transpiration rates in upper canopy needles in Cryptomeria japonica. Kanto J. For. Res. 2018, 69, 19–22, (In Japanese with English summary). [Google Scholar]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Harley, P.C.; Thomas, R.B.; Reynolds, J.F.; Strain, B.R. Modeling photosynthesis of cotton in elevated CO2. Plant Cell Environ. 1992, 15, 271–282. [Google Scholar] [CrossRef]
- Wullschleger, S.D. Biochemical limitation to carbon assimiration in C3 plants—A retrospective analysis of the A/Ci curves from 109 species. J. Exp. Bot. 1993, 44, 907–920. [Google Scholar] [CrossRef]
- Inoue, Y.; Araki, M.; Kitaoka, S.; Kenzo, T.; Saito, S. Relationship between projected shoot area and projected needle area in Cryptomeria japonica D. Don trees. Nihon Shinrin Gakkaishi 2020, 102, 7–14. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry. The Principles and Practice of Statistics in Biological Research, 3rd ed.; W. H. Freeman and Company: New York, NY, USA, 1995. [Google Scholar]
- Zhou, L.; Wang, S.; Chi, Y.; Li, Q.; Huang, K.; Yu, Q. Responses of photosynthetic parameters to drought in subtropical forest ecosystem of China. Sci. Rep. 2015, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wilson, K.B.; Baldocchi, D.D.; Hanson, P.J. Spatial and seasonal variability of photosynthetic parameters and their relationship to leaf nitrogen in a deciduous forest. Tree Physiol. 2000, 20, 565–578. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.R.; Dreyer, E.; Adams, M.A. Photosynthesis-Rubisco relationships in foliage of Pinus sylvestris in response to nitrogen supply and the proposed role of Rubisco and amino acids as nitrogen stores. Trees 2003, 17, 359–366. [Google Scholar] [CrossRef]
- Ripullone, F.; Grassi, G.; Lauteri, M.; Borghetti, M. Photosynthesis–nitrogen relationships: Interpretation of different patterns between Pseudotsuga menziesii and Populus×euroamericana in a mini-stand experiment. Tree Physiol. 2003, 23, 137–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitaoka, S.; Laiye, Q.; Watanabe, Y.; Watanabe, M.; Watanabe, T.; Koike, T. Heterophyllous shoots of Japanese larch trees: The seasonal and yearly variation in CO2 assimilation capacity of the canopy top with changing environment. Plants 2020, 9, 1278. [Google Scholar] [CrossRef]
- Walker, A.P.; Beckerman, A.P.; Gu, L.; Kattge, J.; Cernusak, L.A.; Domingues, T.F.; Woodward, F.I. The relationship of leaf photosynthetic traits–Vcmax and Jmax–to leaf nitrogen, leaf phosphorus, and specific leaf area: A meta-analysis and modeling study. Ecol. Evol. 2014, 4, 3218–3235. [Google Scholar] [CrossRef] [Green Version]
- Varone, L.; Gratani, L. Leaf respiration responsiveness to induced water stress in Mediterranean species. Environ. Exp. Bot. 2015, 109, 141–150. [Google Scholar] [CrossRef]
- Amthor, J.S. Respiration and Crop Productivity; Springer Science & Business Media: New York, NY, USA, 1989. [Google Scholar]
- Kramer, P.J.; Boyer, J.S. Water Relations of Plants and Soils; Academic Press: San Diego, CA, USA, 1995. [Google Scholar]
- Poorter, H.; Niinemets, Ü.; Poorter, L.; Wright, I.J.; Villar, R. Causes and consequences of variation in leaf mass per area (LMA): A meta-analysis. New Phytol. 2009, 182, 565–588. [Google Scholar] [CrossRef]
- Ichie, T.; Yoneyama, A.; Hashimoto, T.; Tanaka-Oda, A.; Kusin, K.; Kenzo, T. Drainage effects on leaf traits of trees in tropical peat swamp forests in Central Kalimantan, Indonesia. Tropics 2019, 28, 1–11. [Google Scholar] [CrossRef]
- Azuma, W.; Ishii, H.R.; Kuroda, K.; Kuroda, K. Function and structure of leaves contributing to increasing water storage with height in the tallest Cryptomeria japonica trees of Japan. Trees 2016, 30, 141–152. [Google Scholar] [CrossRef]
- Niinemets, Ü. Stomatal conductance alone does not explain the decline in foliar photosynthetic rates with increasing tree age and size in Picea abies and Pinus sylvestris. Tree Physiol. 2002, 22, 515–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helms, J.A. Diurnal and seasonal patterns of net assimilation in Douglas-Fir, Pseudotsuga menziesii (Mirb). Franco, as influenced by environment. Ecology 1965, 46, 698–708. [Google Scholar] [CrossRef]
- Kenzo, T.; Yoneda, R.; Ninomiya, I. Seasonal changes in photosynthesis and starch content in Japanese fir (Abies firma Sieb. et Zucc.) saplings under different levels of irradiance. Trees 2018, 32, 429–439. [Google Scholar] [CrossRef]
- Saiki, S.T.; Ando, Y.; Yazaki, K.; Tobita, H. Drought hardening contributes to the maintenance of proportions of non-embolized xylem and cambium status during consecutive dry treatment in container-grown seedling of Japanese cedar (Cryptomeria japonica). Forests 2020, 11, 441. [Google Scholar] [CrossRef]
- Bacelar, E.A.; Moutinho-Pereira, J.M.; Gonçalves, B.C.; Ferreira, H.F.; Correia, C.M. Changes in growth, gas exchange, xylem hydraulic properties and water use efficiency of three olive cultivars under contrasting water availability regimes. Environ. Exp. Bot. 2007, 60, 183–192. [Google Scholar] [CrossRef]
- Samuelson, L.J.; Kane, M.B.; Markewitz, D.; Teskey, R.O.; Akers, M.K.; Stokes, T.A.; Qi, J. Fertilization increased leaf water use efficiency and growth of Pinus taeda subjected to five years of throughfall reduction. Can. J. For. Res. 2018, 48, 227–236. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vcmax | Jmax | Rd | LMA | Narea | Nmass | |
|---|---|---|---|---|---|---|
| Fixed Effect | F-value, p | F-value, p | F-value, p | F-value, p | F-value, p | F-value, p |
| Season | 30.5, <0.0001 | 3.0, ns | 51.6, <0.0001 | 200.9, <0.0001 | 142.7, <0.0001 | 9.1, <0.001 |
| Treatment | 0.9, ns | 1.3, ns | 0.0, ns | 0.2, ns | 0.8, ns | 0.2, ns |
| Season × treatment | 1.0, ns | 1.6, ns | 1.7, ns | 1.5, ns | 2.6, ns | 0.4, ns |
| Pmax | gs | WUEi | |
|---|---|---|---|
| Fixed effect | d.f.,F-value, p | d.f.,F-value, p | d.f.,F-value, p |
| Season | 2, 50.1, <0.0001 | 2, 4.7, <0.05 | 2, 26.9, <0.0001 |
| Treatment | 1, 7.6, <0.01 | 1, 17.6, <0.0001 | 1, 6.9, <0.05 |
| AM/PM | 1, 44.3, <0.0001 | 1, 53.0, <0.0001 | 1, 8.9, <0.01 |
| Season × treatment | 2, 4.1, ns | 2, 1.4, ns | 2, 0.5, ns |
| Season × AM/PM | 2, 0.8, ns | 2, 1.3, ns | 2, 0.1, ns |
| Treatment × AM/PM | 1, 0.1, ns | 1, 0.2, ns | 1, 0.6, ns |
| Season × treatment × AM/PM | 2, 1.0, ns | 2, 0.2, ns | 2, 0.9, ns |
| Treatment | Control | Drought | ||
|---|---|---|---|---|
| Measurement Time | Morning | Afternoon | Morning | Afternoon |
| Β-value, p | β-value, p | β-value, p | β-value, p | |
| gs | 0.24, ns | 0.79, <0.0001 | 0.39, <0.05 | 0.67, <0.0001 |
| LMA | −0.72, <0.001 | −0.31, <0.01 | −0.55, <0.01 | −0.31, <0.05 |
| Nmass | −0.17, ns | 0.07, ns | −0.26, ns | −0.08, ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kenzo, T.; Inoue, Y.; Araki, M.G.; Kawasaki, T.; Kitaoka, S.; Tsurita, T.; Sakata, T.; Saito, S. Effects of Throughfall Exclusion on Photosynthetic Traits in Mature Japanese Cedar (Cryptomeria japonica (L. f.) D. Don.). Forests 2021, 12, 971. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080971
Kenzo T, Inoue Y, Araki MG, Kawasaki T, Kitaoka S, Tsurita T, Sakata T, Saito S. Effects of Throughfall Exclusion on Photosynthetic Traits in Mature Japanese Cedar (Cryptomeria japonica (L. f.) D. Don.). Forests. 2021; 12(8):971. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080971
Chicago/Turabian StyleKenzo, Tanaka, Yuta Inoue, Masatake G. Araki, Tatsuro Kawasaki, Satoshi Kitaoka, Tatsuya Tsurita, Tadashi Sakata, and Satoshi Saito. 2021. "Effects of Throughfall Exclusion on Photosynthetic Traits in Mature Japanese Cedar (Cryptomeria japonica (L. f.) D. Don.)" Forests 12, no. 8: 971. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080971