
An Analysis of Atlantic White Cedar (Chamaecyparis thyoides (L.) B.S.P.) Tree Rings as Indicators of Ghost Forest Development in a Globally Threatened Ecosystem


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Wood Sample Collection and Preparation
2.3. Cross-Dating and Chronology Development
2.4. Climatological Data
2.5. Data Analysis
3. Results
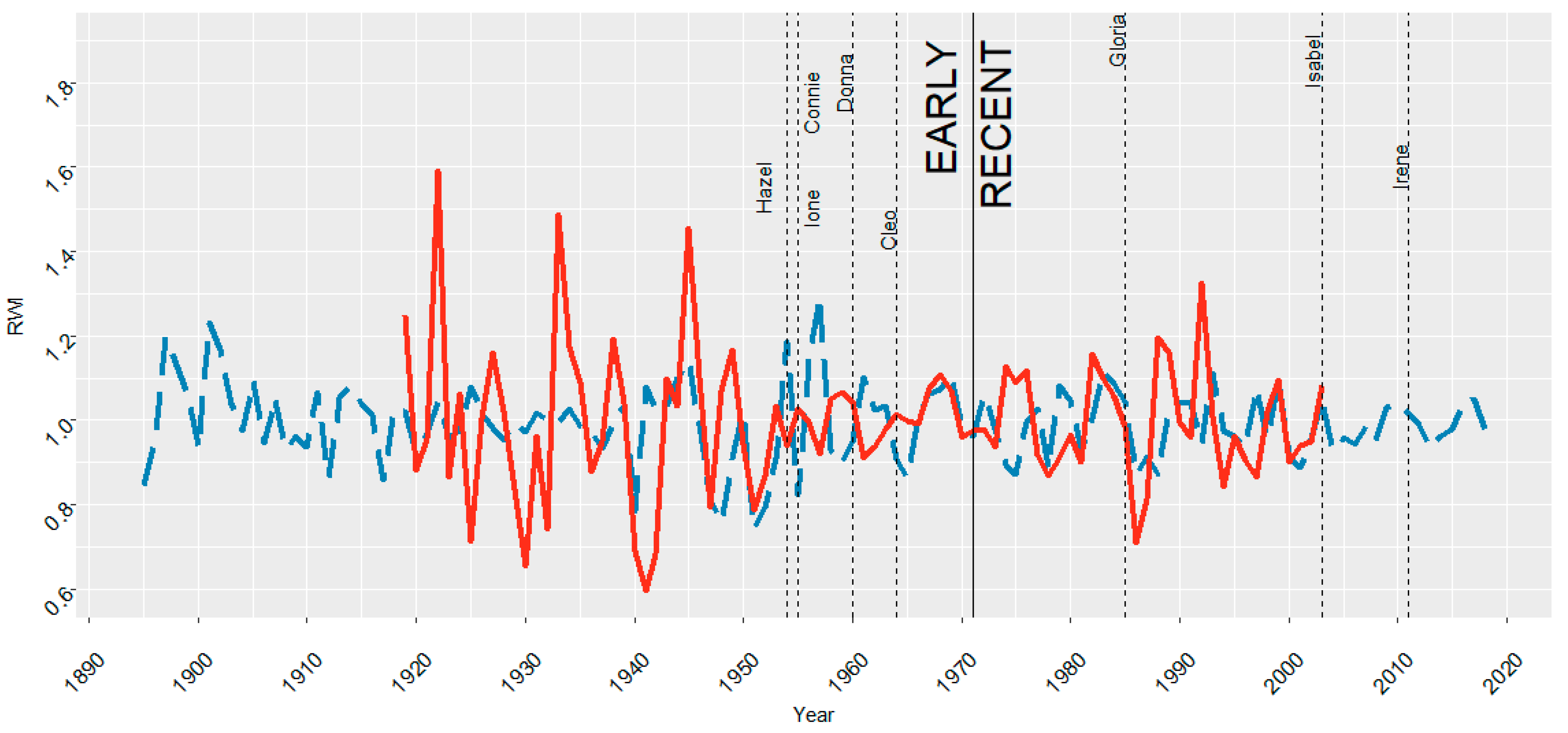
3.1. ARNWR and GDSNWR Ring Width Index Results for the Full Chronologies
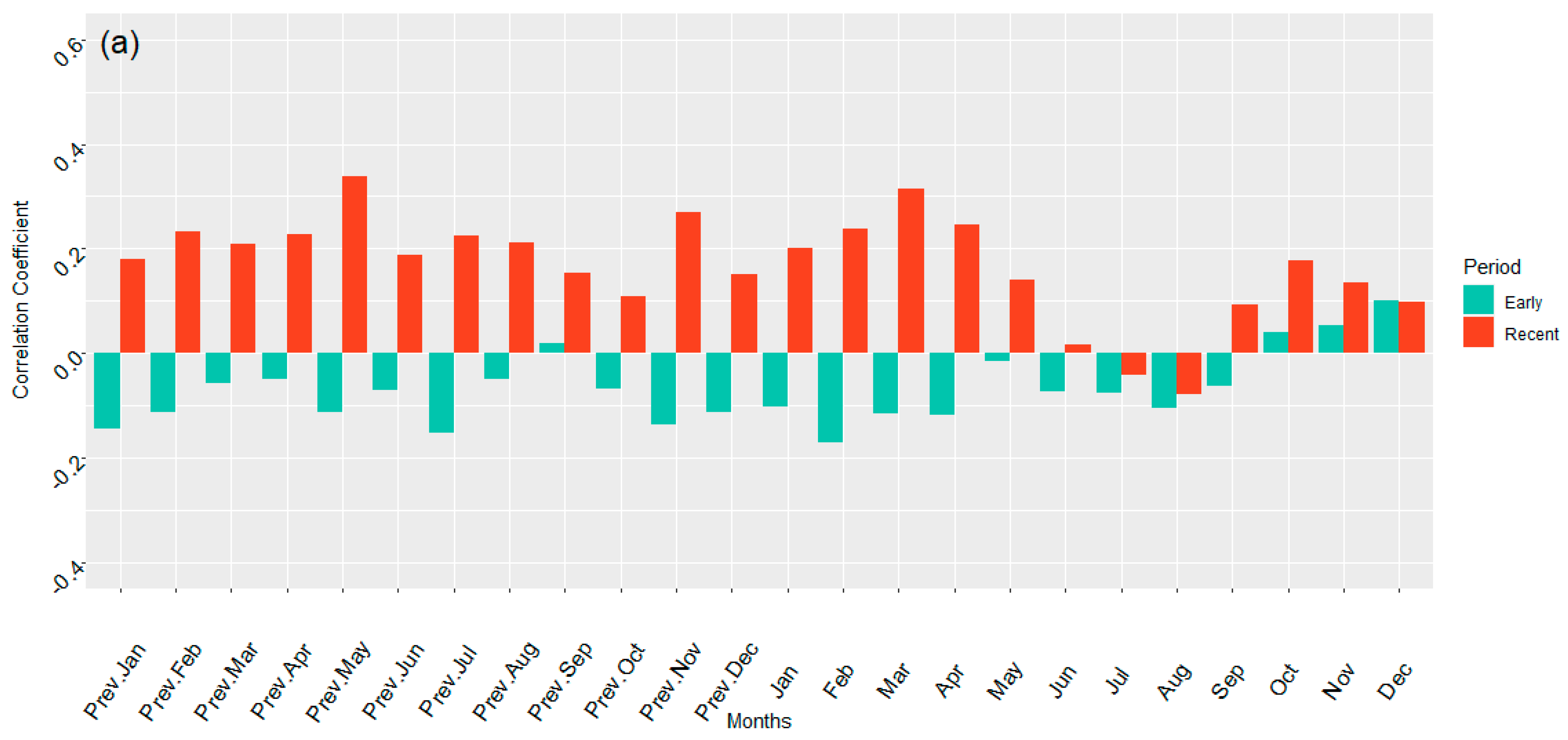
3.2. Correlations of AWC Growth and PDSI at ARNWR and GDSNWR for Divided (Early and Recent) Chronologies
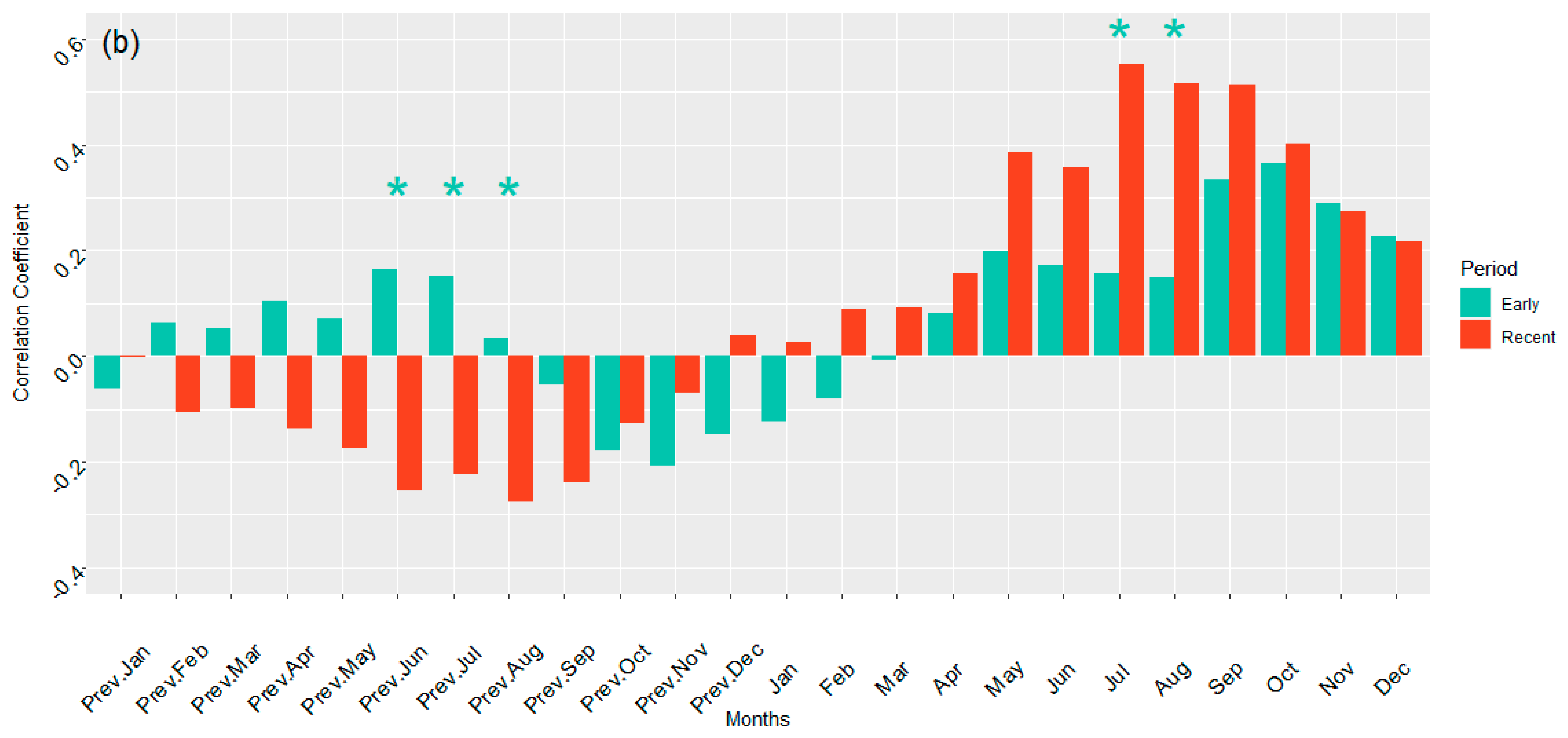
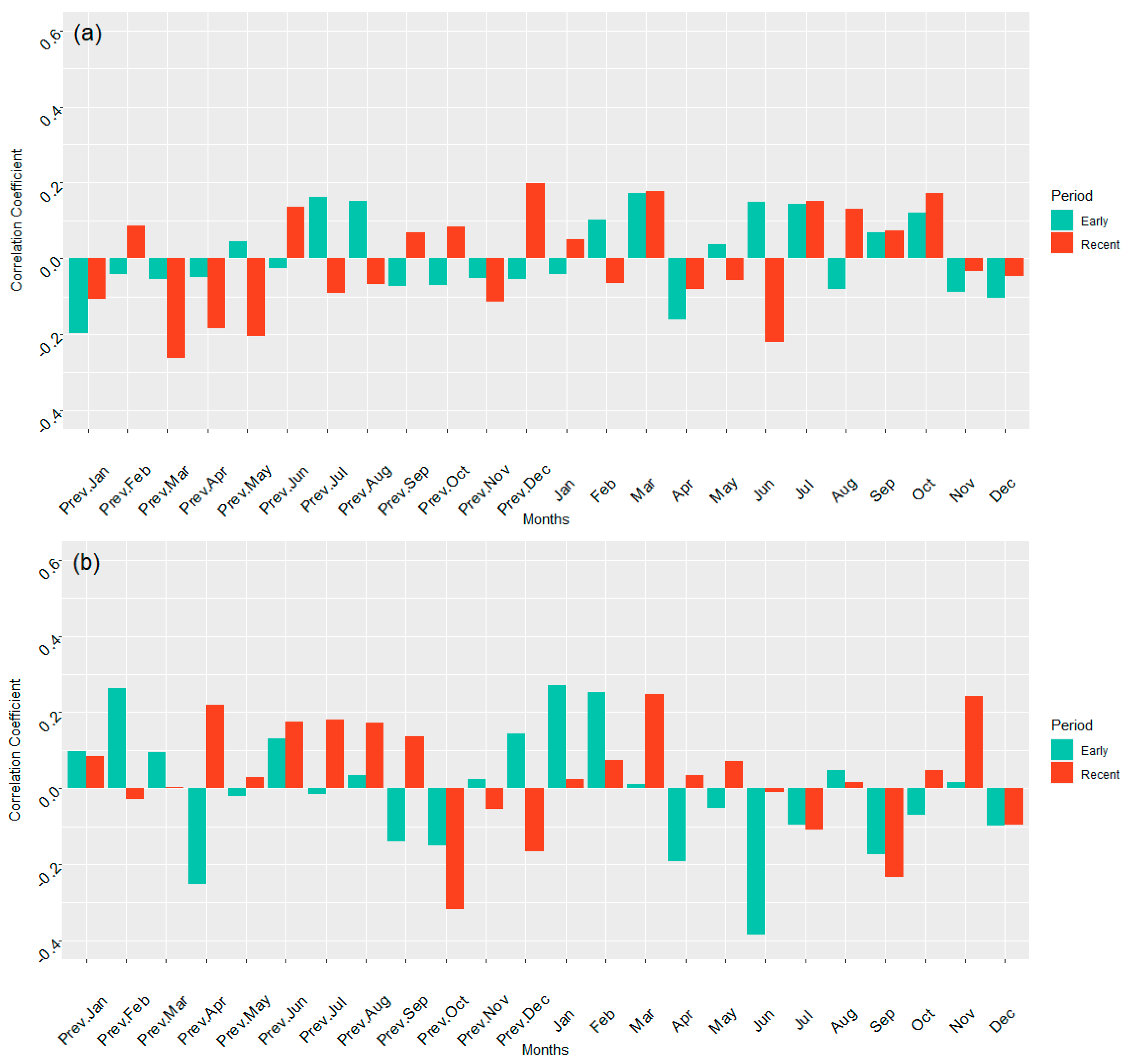
3.3. Temperature Correlation with Growth in ARNWR and GDSNWR
4. Discussion
4.1. Climate-Growth Relationships among Study Periods and Reference Sites
4.1.1. PDSI and Growth
4.1.2. Temperature and Growth
4.2. Hurricane Responses for Both Periods in the Study Site
4.3. Self-Maintenance Implications of Water Level and Salinity
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Manda, A.K.; Giuliano, A.S.; Allen, T.R. Influence of artificial channels on the source and extent of saline water intrusion in the wind tide dominated wetlands of the southern Albemarle estuarine system (USA). Env. Earth Sci. 2014, 71, 4409–4419. [Google Scholar] [CrossRef]
- Church, J.A.; Clark, P.U.; Cazenave, A.; Gregory, J.M.; Jevrejeva, S.; Levermann, A.; Merrifield, M.A.; Milne, G.A.; Nerem, R.S.; Nunn, P.D.; et al. Sea level change. In Climate Change 2013: The Physical Science Basis; Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2013; pp. 1–1535. [Google Scholar]
- Magolan, J.; Halls, J. A multi-decadal investigation of tidal creek wetland changes, water level rise, and ghost forests. Remote Sens. 2020, 12, 1141. [Google Scholar] [CrossRef] [Green Version]
- Craft, C.; Clough, J.; Ehman, J.; Joye, S.; Park, R.; Pennings, S.; Guo, H.; Machmuller, M. Forecasting the effects of accelerated sea-level rise on tidal marsh ecosystem services. Front. Ecol. Environ. 2009, 7, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Conner, W.H.; Day, J.W. The ecology of forested wetlands in the southeastern United States. In Wetlands Ecology and Management; National Institute of Ecology and International Scientific Publications: Jaipur, India, 1982; pp. 69–87. [Google Scholar]
- Brinson, M.M. A Hydrogeomorphic Classification of Wetlands; U.S. Army Engineer Waterways Experiment Station: Vicksburg, MS, USA, 1993; pp. 1–103. [Google Scholar]
- Kirwan, M.; Gedan, K. Sea-level driven land conversion and the formation of ghost forests. Nat. Clim. Chang. 2019, 9, 450–457. [Google Scholar] [CrossRef] [Green Version]
- Schieder, N.W.; Kirwan, M.L. Sea-level driven acceleration in coastal forest retreat. Geology 2019, 47, 1151–1155. [Google Scholar] [CrossRef]
- Laderman, A.D. Why does the Freshwater Genus Chamaecyparis Hug Marine Coasts? In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 1–30. [Google Scholar]
- Kalm, P.; Forster, J.R.; Joseph Meredith Toner Collection. Travels into North America; Containing Its Natural History, and a Circumstantial Account of Its Plantations and Agriculture in General, with the Civil, Ecclesiastical and Commercial State of the Country, the Manners of the Inhabitants, and Several Curious and Important Remarks on Various Subjects, 2nd ed.; Longman, Hurst, Rees, and Orme; Biodiversity Heritage Library: London, UK, 1812; Volume 1, pp. 1–452. [Google Scholar]
- USDA Plants Database. Available online: https://plants.usda.gov/home (accessed on 19 June 2021).
- Thompson, G.; Belcher, R.T.; Atkinson, R.B. Soil biogeochemistry in Virginia and North Carolina Atlantic white cedar swamps. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 113–124. [Google Scholar]
- DeBerry, J.W.; Atkinson, R.B. Aboveground forest biomass and litter production patterns in Atlantic white cedar swamps of differing hydroperiods. Southeast. Nat. 2014, 13, 673–690. [Google Scholar] [CrossRef]
- Weiser, J.L. The Effect of Water Levels on Carbon Dioxide Emissions from Soil Microcosms in the Great Dismal Swamp. Master’s Thesis, Christopher Newport University, Newport News, VA, USA, 2014. [Google Scholar]
- Duttry, P.M.; Thompson, G.; Belcher, R.T.; Atkinson, R.B. Soil respiration response to fluctuating water levels in laboratory columns of soils from AWC peatlands and mitigation sites in Virginia and North Carolina. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 165–174. [Google Scholar]
- Crawford, E.R.; Day, F.P.; Atkinson, R.B. Decomposition dynamics in an Atlantic white cedar restoration site. In Proceedings of the Arlington Echo Symposium, Asheville, NC, USA, 2–4 June 2003; Burke, M.K., Sheridan, P., Eds.; Department of Agriculture Forest Service: Asheville, NC, USA, 2005; pp. 11–16. [Google Scholar]
- Korstian, C.F.; Brush, W.D. Southern White Cedar, 1st ed.; U.S. Department of Agriculture: Washington, DC, USA, 1931; pp. 1–1485.
- Little, S., Jr. Ecology and Silviculture of Whitecedar and Associated Hardwoods in Southern New Jersey; Yale University School of Forestry Bulletin: New Haven, CT, USA, 1950; pp. 1–134. [Google Scholar]
- Laderman, A.D. The Ecology of Atlantic White Cedar Wetlands: A Community Profile; U.S. Fish and Wildlife Service: Washington, DC, USA, 1989; Volume 85, pp. 1–125.
- Ponnamperuma, F.N. The chemistry of submerged soils. Adv. Agron. 1972, 24, 29–92. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 5th ed.; Wiley: Hoboken, NJ, USA, 1993; p. 736. [Google Scholar]
- Tiner, R.W. Wetland Indicators: A Guide to Wetland Formation, Identification, Delineation, Classification, and Mapping, 2nd ed.; CRC Press: New York, NY, USA, 2017; pp. 1–631. [Google Scholar]
- Kozlowski, T.T. Responses of woody plants to flooding and salinity. Tree Physiol. 1997, 17, 490. [Google Scholar] [CrossRef]
- Conner, W.; Song, B.; Williams, T.; Vernon, J. Long-term tree productivity of a South Carolina coastal plain forest across a hydrology gradient. J. Plant Ecol. 2011, 4, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Shaler, N.S. General Account of the Fresh-Water Morasses of the United States; with a Description of Virginia and North. Carolina, 10th Annual Report for 1888–1889; U.S. Government Printing Office: Washington, DC, USA, 1890; pp. 1–124.
- Lilly, J.P. The Blackland Soils of North Carolina, Their Characteristics and Management for Agriculture; N.C. Agricultural Research Service technical bulletin no. 270; North Carolina State University: Raliegh NC, USA, 1981; pp. 1–70. [Google Scholar]
- Megonigal, J.P.; Day, F.P. Effects of flooding on root and shoot production of bald cypress in large experimental enclosures. Ecology 1992, 73, 1182–1193. [Google Scholar] [CrossRef]
- Conner, W.H.; Day, J.W. Productivity and composition of a bald cypress-water tupelo site and a bottomland hardwood site in a Louisiana swamp. Am. J. Bot. 1976, 63, 1354–1364. [Google Scholar] [CrossRef]
- Conner, W.H.; Gosselink, J.G.; Parrondo, R.T. Comparison of the vegetation of three Louisiana swamp sites with different flooding regimes. Am. J. Bot. 1981, 68, 320–331. [Google Scholar] [CrossRef]
- Ehrenfeld, J.G. Microtopography and vegetation in Atlantic white cedar swamps; the effects of natural disturbances. Can. J. Bot. 1995, 73, 474–484. [Google Scholar] [CrossRef]
- Atkinson, R.B.; Loomis, D.T.; DeBerry, J.W.; Crawford, E.R.; Belcher, R.T. Water tables in Atlantic white cedar swamps: Implications for restoration. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 137–150. [Google Scholar]
- Rodgers, H.L.; Day, F.P.; Atkinson, R.B. Fine root dynamics in two Atlantic white-cedar wetlands with contrasting hydroperiods. Wetlands 2003, 23, 941–949. [Google Scholar] [CrossRef]
- Brown, D.A.; Atkinson, R.B. Remote sensing interpretation of twenty-five years of Atlantic white cedar clearcutting in North Carolina. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 55–66. [Google Scholar]
- Harrison, J.M.; DeBerry, J.W.; Belcher, R.T.; Loomis, D.T.; Atkinson, R.B. Effects of water table on survival and growth of Atlantic white cedar in two young planted sites. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 181–196. [Google Scholar]
- Cook, J.W.B.; Foster, E.M.; Atkinson, R.B. Effect of hydrology on post-fire survival and growth of containerized seedlings, rooted cuttings, and naturally-regenerated Atlantic white cedar in the Great Dismal Swamp National Wildlife Refuge. In Proceedings of the Restoration and Management of Atlantic White Cedar Swamps Symposium, Suffolk, VA, USA, 12–14 June 2012; Belcher, R.T., Hudson, H.W., Eds.; Great Dismal Swamp National Wildlife Refuge: Suffolk, VA, USA, 2015; pp. 1–15. [Google Scholar]
- Foster, E.M.; Cook, J.W.B.; Atkinson, R.B. Post-fire survival and growth of containerized seedlings, rooted cuttings, and natural regenerants of Atlantic white cedar (Chamaecyparis thyoides) in the Great Dismal Swamp National Wildlife Refuge. In Proceedings of the Restoration and Management of Atlantic White Cedar Swamps Symposium, Suffolk, VA, USA, 12–14 June 2012; Belcher, R.T., Hudson, H.W., Eds.; Great Dismal Swamp National Wildlife Refuge: Suffolk, VA, USA, 2015; pp. 16–27. [Google Scholar]
- Giese, G.L.; Wilder, H.B.; Parker, G.G., Jr. Hydrology of Major Estuaries and Sounds of North Carolina; U.S. Government Printing Office: Washington, DC, USA, 1985; pp. 1–119.
- Anderson, R.R.; Brown, R.G.; Rappleye, R.D. Water quality and plant distribution along the upper Patuxent River, Maryland. Chesap. Sci. 1968, 9, 145–156. [Google Scholar] [CrossRef]
- Atkinson, R.B.; Bodkin, N.L.; Perry, J.E. New county records collected in tidal wetlands of four coastal plain counties along the James River, Virginia. Castanea 1990, 55, 55–64. [Google Scholar]
- Perry, J.E.; Atkinson, R.B. York River Tidal Marshes. J. Coast. Res. 2009, 10057, 40–49. [Google Scholar] [CrossRef]
- Kirwan, M.; Kirwan, J.; Copenheaver, C. Dynamics of an estuarine forest and its response to rising sea level. J. Coast. Res. 2007, 23, 457–463. [Google Scholar] [CrossRef]
- Poulter, B.; Christensen, N.L.; Qian, S.S. Tolerance of Pinus taeda and Pinus serotina to low salinity and flooding: Implications for equilibrium vegetation dynamics. J. Veg. Sci. 2008, 19, 15–22. [Google Scholar] [CrossRef]
- Bowen, B.D. Dendroecological Assessment of Loblolly Pine (Pinus taeda) along a Salinity Gradient within the York River Subestuary in Virginia. Master’s Thesis, Christopher Newport University, Newport News, VA, USA, 2016. [Google Scholar]
- Robichaud, A.; Bégin, Y. The effects of storms and sea-level rise on a coastal forest margin in New Brunswick, Eastern Canada. J. Coast. Res. 1997, 13, 429–439. [Google Scholar]
- Weakley, A.S.; Ludwig, J.C.; Townsend, J.F. Flora of Virginia; Crowder, B., Ed.; Botanical Research Institute of Texas Press: Fort Worth, TX, USA, 2012. [Google Scholar]
- Yanosky, T.; Hupp, C.; Hackney, C. Chloride concentrations in growth rings of Taxodium distichum in a saltwater-intruded estuary. Ecol. Appl. 1995, 5, 785–792. [Google Scholar] [CrossRef]
- Krauss, K.W.; Duberstein, J.A.; Doyle, T.W.; Conner, W.H.; Day, R.H.; Inabinette, L.W.; Whitbeck, J.L. Site condition, structure, and growth of Bald cypress along tidal/non-tidal salinity gradients. Wetlands 2009, 29, 505–519. [Google Scholar] [CrossRef]
- Hinesley, E.; North Carolina State University, Raleigh, NC, USA. Personal communication, 2021.
- Stotts, S.; Callahan, J.; Gulledge, O. Impact of channel dredging and straightening in an Atlantic white cedar (Chamaecyparis thyoides L. (B.S.P.)) freshwater tidal wetland. J. Coast. Res. 2021, 1, 1–14. [Google Scholar]
- Williams, B.; Pine Creek Forestry, Laurel Springs, NJ, USA. Personal communication, 2020.
- Reddy, A.; Hawbaker, T.; Wurster, F.; Zhu, Z.; Ward, S.; Newcomb, D.; Murray, R. Quantifying soil carbon loss and uncertainty from a peatland wildfire using multi-temporal LiDAR. Remote Sens. Environ. 2015, 170, 306–316. [Google Scholar] [CrossRef] [Green Version]
- Trettin, C.; Jurgensen, M.F.; Grigal, D.F.; Gale, M.R.; Jeglum, J.K. Northern Forested Wetlands: Ecology and Management, 1st ed.; Lewis Publishers: New York, NY, USA, 1997; pp. 1–512. [Google Scholar]
- Keeland, B.D.; Sharitz, R.R. Seasonal growth patterns of Nyssa sylvatica var. Biflora, Nyssa aquatica, and Taxodium distichum as affected by hydrologic regime. Can. J. For. Res. 1995, 25, 1084–1096. [Google Scholar] [CrossRef]
- Atkinson, R.B. Growth rates of Atlantic white cedar depend on hydrologic regimes at two time scales. Wetlands 2020, 40, 1–12. [Google Scholar] [CrossRef]
- Heddinghaus, T.R.; Sabol, P.A. Review of the Palmer drought severity index and where do we go from here? In Proceedings of the 7th conference on Applied Climatology, Salt Lake City, UT, USA, 10–13 September 1991; pp. 242–246. [Google Scholar]
- Ury, E.A.; Yang, X.; Wright, J.P.; Bernhardt, E.S. Rapid deforestation of a coastal landscape driven by sea-level rise and extreme events. Ecol. Appl. 2021, 31, e2339. [Google Scholar] [CrossRef] [PubMed]
- Ferrell, G.M.; Strickland, A.G.; Spruill, T.B. Effects of canals and roads on hydrologic conditions and health of Atlantic white cedar at the Emily and Richardson Preyer Buckridge Coastal Reserve, North Carolina, 2003–2006. Geol. Surv. 2007, 1–188. [Google Scholar] [CrossRef]
- Stewart, D.; Alligator River National Wildlife Refuge, Manteo, NC, USA. Personal communication, 2003.
- Alligator River National Wildlife Refuge Annual Narrative Report; U.S. Fish and Wildlife Service: Washington, DC, USA, 2007; pp. 1–110.
- Alligator River National Wildlife Refuge Comprehensive Conservation Plan; U.S. Fish and Wildlife Service: Washington, DC, USA, 2008; pp. 1–260.
- Official Series Description—PUNGO Series. Available online: https://soilseries.sc.egov.usda.gov/OSD_Docs/P/PUNGO.html (accessed on 20 June 2021).
- Shacochis, K.M.; DeBerry, J.W.; Loomis, D.T.; Belcher, R.T.; Atkinson, R.B. Vegetation importance values and weighted averages of Atlantic white cedar stands in Great Dismal Swamp and Alligator River National Wildlife Refuges. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 227–234. [Google Scholar]
- Great Dismal Swamp—U.S. Fish and Wildlife Service. Available online: https://www.fws.gov/refuge/great_dismal_swamp/ (accessed on 21 June 2021).
- Carter, A.R. Cedar Restoration in the Dismal Swamp of Virginia and North. Carolina; Laderman, A.D., Ed.; Westview Press: Boulder, CO, USA, 1987; pp. 323–325. [Google Scholar]
- Atkinson, R.B.; Morgan, T.E.; Belcher, R.T.; Brown, D.A. The role of historical inquiry in the restoration of Atlantic white cedar swamps. In Proceedings of the Atlantic White Cedar Restoration Ecology and Management Symposium, Christopher Newport University, Newport News, VA, USA, 31 May–2 June 2000; Atkinson, R.B., Belcher, R.T., Brown, D.A., Perry, J.E., Eds.; VIMS Publication Center: Gloucester Point, VA, USA, 2003; pp. 43–54. [Google Scholar]
- Duberstein, J.; Krauss, K.W. Forest Community Biomass and Growth in Great Dismal Swamp, Virginia and North Carolina, USA [Data Set]; U.S. Geological Survey: Reston, VA, USA, 2018.
- Day, F.P. Litter accumulation in four plant communities in the Dismal Swamp, Virginia. Am. Midl. Nat. 1979, 102, 281–289. [Google Scholar] [CrossRef]
- Great Dismal Swamp National Wildlife Refuge and Nansemond National Wildlife Refuge Final Comprehensive Conservation Plan; U.S. Fish and Wildlife Service: Washington, DC, USA, 2006; pp. 1–272.
- Patterson, C.L.; Atkinson, R.B. Radial growth of peatland Atlantic white cedar (Chamaecyparis thyoides) in Great Dismal Swamp National Wildlife Refuge and its association with Lake Drummond water levels. In Proceedings of the Restoration and Management of Atlantic White Cedar Swamps Symposium, Suffolk, VA, USA, 12–14 June 2012; Belcher, R.T., Hudson, H.W., Eds.; Great Dismal Swamp National Wildlife Refuge: Suffolk, VA, USA, 2015; p. 102. [Google Scholar]
- Merry, S.D. Factors Affecting Tree Ring Width in Atlantic White Cedar, Chamaecyparis thyoides (L.) B.S.P., within Great Dismal Swamp National Wildlife Refuge and Alligator River National Wildlife Refuges. Master’s Thesis, Christopher Newport University, Newport News, VA, USA, 2005. [Google Scholar]
- Stokes, M.A.; Smiley, T.C. An Introduction to Tree-Ring Dating; The University of Arizona Press: Tucson, AZ, USA, 1996; pp. 1–73. [Google Scholar]
- Copenheaver, C.A.; Kyle, K.H.; Stevens, G.N.; Kamp, M.H. Comparing Juniperus virginiana tree-ring chronologies from forest edge vs. forest interior positions in the Cedars Natural Area Preserve in Virginia, USA. Dendrochronologia 2005, 23, 39–45. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C.; Buras, A.; Cecile, J.; Mudelsee, M.; et al. DplR: Dendrochronology Program Library in R; R package version 1.7.2. 2021. Available online: https://cran.r-project.org/web/packages/dplR/index.html (accessed on 21 June 2021).
- The R Project for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 21 June 2021).
- RStudio: Integrated Development Environment for R. RStudio. Available online: http://www.rstudio.com/.http://www.rstudio.com (accessed on 21 June 2021).
- Cook, E.; Pederson, N. Uncertainty, emergence, and statistics in dendrochronology. Dendroclimatol. Prog. Prospect. 2011, 11, 77–112. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Divisional Data Select. Available online: https://www7.ncdc.noaa.gov/CDO/CDODivisionalSelect.jsp# (accessed on 21 June 2021).
- Historical Hurricane Tracks. Available online: https://coast.noaa.gov/hurricanes/#map=4/32/-80 (accessed on 21 June 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Royal Stat. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- EnviroAtlas. Available online: https://enviroatlas.epa.gov/enviroatlas/interactivemap/ (accessed on 18 June 2021).
- Hackney, C.T.; Avery, G.B. Tidal wetland community response to varying levels of flooding by saline water. Wetlands 2015, 35, 227–236. [Google Scholar] [CrossRef]
- Wetlands Subcommittee. Classification of Wetlands and Deepwater Habitats of the United States; Federal Geographic Data Committee: Reston, VA, USA, 2013; pp. 1–91.
- Reddy, K.R.; Patrick, W.H. Effect of alternate aerobic and anaerobic conditions on redox potential, organic matter decomposition and nitrogen loss in a flooded soil. Soil Biol. Biochem. 1975, 7, 87–94. [Google Scholar] [CrossRef]
- Moore, T.R.; Dalva, M. Methane and carbon dioxide exchange potentials of peat soils in aerobic and anaerobic laboratory incubations. Soil Biol. Biochem. 1997, 29, 1157–1164. [Google Scholar] [CrossRef]
- Vompersky, S.E.; Sirin, A.J. Hydrology of drained forested wetlands. In Northern Forested Wetlands Ecology and Management, 1st ed.; Trettin, C.C., Jurgensen, M.F., Grigal, D.F., Gale, M.R., Jeglum, J.K., Eds.; CRC Press: Boca Raton, FL, USA, 1996; pp. 189–211. [Google Scholar]
- Mirda, C.G. The Effects of Water Table on Soil Carbon Dioxide Emissions in Cavalier Wildlife Management Area: The Contributions of Root and Microbial Respiration. Master’s Thesis, Christopher Newport University, Newport News, VA, USA, 2018. [Google Scholar]
- Kalnins, M. Empirically Derived Estimates of Gaseous Carbon Loss from Peatlands in North-Central Alberta, Canada, and Northeastern North Carolina, USA. Master’s Thesis, Christopher Newport University, Newport News, VA, USA, 2000. [Google Scholar]
- Kelsey, R.G.; Joseph, G.; McWilliams, M.G. Ethanol synthesis by anoxic root segments from five cedar species relates to their habitat attributes but not their known differences in vulnerability to Phytophthora lateralis root disease. Can. J. For. Res. 2011, 41, 1202–1211. [Google Scholar] [CrossRef]
- Wurster, F.; Great Dismal Swamp National Wildlife Refuge, Suffolk, VA, USA. Personal communication, 2021.
- Seim, A.M. The Effect of Climate on the Growth of Red Maple (Acer rubrum L.) and Atlantic White Cedar (Chamaecyparis thyoides (L.) B.S.P.) in the Great Dismal Swamp National Wildlife Refuge. Master’s Thesis, Christopher Newport University, Newport News, VA, USA, 2005. [Google Scholar]
- Pearl, J.K.; Anchukaitis, K.J.; Pederson, N.; Donnelly, J.P. Reconstructing northeastern United States temperatures using Atlantic white cedar tree rings. Environ. Res. Lett. 2017, 12, 1–10. [Google Scholar] [CrossRef]
- Hwang, T.; Martin, K.L.; Vose, J.M.; Wear, D.; Miles, B.; Kim, Y.; Band, L.E. Nonstationary hydrologic behavior in forested watersheds is mediated by climate-induced changes in growing season length and subsequent vegetation growth. Water Resour. Res. 2018, 54, 5359–5375. [Google Scholar] [CrossRef]
- Williams, K.; MacDonald, M.; Sternberg, L.D. Interactions of storm, drought, and sea-level rise on coastal forest: A case study. J. Coast. Res. 2003, 19, 1116–1121. [Google Scholar]
- Zimmermann, G.; Stockton University, Galloway, NY, USA. Personal communication, 2020.
- Conner, W.H.; Day, J.W. Variations in vertical accretion in a Louisiana Swamp. J. Coast. Res. 1991, 7, 617–622. [Google Scholar]
- Sutter, L.A.; Chambers, R.M.; Perry, J.E. Seawater intrusion mediates species transition in low salinity, tidal marsh vegetation. Aquat. Bot. 2015, 122, 32–39. [Google Scholar] [CrossRef]
- Friedrichs, C.T.; Perry, J.E. Tidal salt marsh morphodynamics: A synthesis. J. Coast. Res. 2001, 27, 7–37. [Google Scholar]
- Waldron, M.C.B.; Carter, G.A.; Biber, P.D. Using aerial imagery to determine the effects of sea-level rise on fluvial marshes at the mouth of the Pascagoula River (Mississippi, USA). J. Coast. Res. 2021, 37, 389–407. [Google Scholar]
- Whitehead, R.D. Developmental and environmental history of the Dismal Swamp. Ecol. Monogr. 1972, 42, 301–315. [Google Scholar] [CrossRef]
- Watershed Report. Available online: https://watersgeo.epa.gov/watershedreport/?comid=10484491#streamcat-data (accessed on 23 June 2021).
- CRC Science Panel. North Carolina Sea Level Rise Assessment Report; North Carolina Department of Natural and Cultural Resources: Raleigh, NC, USA, 2015; pp. 1–43.






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Hurricane Name | Max Wind Speed in AOI (kt) | 10-Year Running Average RWI | Percent Change from 10-Year Average | |||
|---|---|---|---|---|---|---|---|
| +0 | +1 | +2 | +3 | ||||
| 1954 | Hazel | 80 | 0.926 | 28.269 | −11.089 | 22.449 | 39.807 |
| 1955 | Connie, Ione | 85, 65 | 0.984 | −7.929 | 26.801 | 44.776 | 3.944 |
| 1960 | Donna | 85 | 0.966 | −1.622 | 13.918 | 6.107 | 6.929 |
| 1964 | Cleo | 40 | 1.010 | −10.034 | −14.378 | 0.434 | 5.218 |
| 1985 | Gloria | 90 | 1.021 | 2.273 | −14.681 | −10.487 | −14.670 |
| 2003 | Isabel | 85 | 0.981 | 7.507 | −5.390 | −2.230 | −4.045 |
| 2011 | Irene | 65 | 0.986 | 3.337 | 0.366 | −5.243 | −2.396 |
| ARNWR | GDSNWR | |||||
|---|---|---|---|---|---|---|
| Full | Early | Recent | Full | Early | Recent | |
| Number of Trees (number of time series) | 103 (420) | 102 (416) | 103 (419) | 42 stem cuts (≥168 radii) | 42 stem cuts (≥168 radii) | 42 stem cuts (≥168 radii) |
| Chronology Length (years) | 1895–2018 (123) | 1895–1971 (76) | 1972–2018 (46) | 1919–2003 (84) | 1919–1971 (52) | 1972–2003 (31) |
| Mean Age | 62.25 | 28.88 | 33.69 | 81.26 | 49.26 | 32 |
| Median Age | 61 | 30 | 32 | 81 | 49 | 32 |
| Signal-to-Noise Ratio (SNR) | 31.147 | 38.57 | 28.341 | 43.888 | 64.379 | 28.565 |
| Mean Subsample Signal Strength (SSS) | 0.8175 | 0.7859 | 0.9659 | 0.9943 | 0.9935 | 1 |
| Expressed Population Signal (EPS) | 0.969 | 0.975 | 0.966 | 0.978 | 0.985 | 0.966 |
| Effective Average Correlation Between Series (RBAR.eff) | 0.184 | 0.273 | 0.126 | 0.522 | 0.623 | 0.405 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Doyle, J.M.; Earley, K.E.; Atkinson, R.B. An Analysis of Atlantic White Cedar (Chamaecyparis thyoides (L.) B.S.P.) Tree Rings as Indicators of Ghost Forest Development in a Globally Threatened Ecosystem. Forests 2021, 12, 973. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080973
Doyle JM, Earley KE, Atkinson RB. An Analysis of Atlantic White Cedar (Chamaecyparis thyoides (L.) B.S.P.) Tree Rings as Indicators of Ghost Forest Development in a Globally Threatened Ecosystem. Forests. 2021; 12(8):973. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080973
Chicago/Turabian StyleDoyle, James M., Kendal E. Earley, and Robert B. Atkinson. 2021. "An Analysis of Atlantic White Cedar (Chamaecyparis thyoides (L.) B.S.P.) Tree Rings as Indicators of Ghost Forest Development in a Globally Threatened Ecosystem" Forests 12, no. 8: 973. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080973