
Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region


1
Instituto Interuniversitario de Investigación del Sistema Tierra en Andalucía (IISTA), Universidad de Granada, Avda. del Mediterráneo s/n, E-18006 Granada, Spain
2
Grupo de Ecología Terrestre, Departamento de Ecología, Universidad de Granada, Avda. Fuentenueva s/n, E-18071 Granada, Spain
3
Servicio de Evaluación, Restauración y Protección de Agrosistemas Mediterráneos, Estación Experimental del Zaidín (CSIC), C/Profesor Albareda 1, E-18008 Granada, Spain
4
Departamento de Botánica, Ecología y Fisiología Vegetal, Universidad de Córdoba, Edificio Celestino Mutis, 1° Planta, Campus de Rabanales, E-14014 Córdoba, Spain
*
Author to whom correspondence should be addressed.
†
Current address: Affiliation 3.
Forests 2021, 12(11), 1584; https://0-doi-org.brum.beds.ac.uk/10.3390/f12111584
Submission received: 22 September 2021
/
Revised: 25 October 2021
/
Accepted: 15 November 2021
/
Published: 17 November 2021
(This article belongs to the Section Forest Ecology and Management)
Abstract
:Land abandonment is a major global change driver in the Mediterranean region, where anthropic activity has played an important role in shaping landscape configuration. Understanding the woodland expansion towards abandoned croplands is critical to develop effective management strategies. In this study, we analyze the colonization pattern of abandoned croplands by Quercus pyrenaica in the Sierra Nevada mountain range (southern Spain). We aimed to assess differences among populations within the rear edge of the Q. pyrenaica distribution. For this purpose, we characterized (i) the colonization pattern of Q. pyrenaica, (ii) the structure of the seed source (surrounding forests), and (iii) the abundance of the main seed disperser (Eurasian jay, Garrulus glandarius). The study was conducted in five abandoned croplands located in two representative populations of Q. pyrenaica located on contrasting slopes. Vegetation plots within three habitat types (mature forest, edge-forest and abandoned cropland) were established to compute the abundance of oak juveniles. The abundance of European jay was determined using data of bird censuses (covering 7 years). Our results indicate that a natural recolonization of abandoned croplands by Q. pyrenaica is occurring in the rear edge of the distribution of this oak species. Oak juvenile abundance varied between study sites. Neither the surrounding-forest structure nor the abundance of jays varied significantly between study sites. The differences in the recolonization patterns seem to be related to differences in the previous- and post-abandonment management.
1. Introduction
Land-use change is considered the main global change driver worldwide [1,2], affecting biodiversity [3], modifying ecological processes [4], and altering the provision of ecosystem services [5]. Croplands abandonment and afforestation are the main processes of land-use change in the Northern hemisphere [2,6]. In the Mediterranean region, where anthropic activity has played an important role in shaping landscape configuration, cropland abandonment has been widespread during the second half of the last century [7,8,9]. Land-use change models predict an increase in this trend in the future [10,11]. The abandonment of traditional activities has left many Mediterranean landscapes in almost barren states, with poor vegetation cover [6,12]. Consequently, a natural vegetation regeneration process started, with a spontaneous recovery of abandoned croplands [13,14,15,16,17].
Thus, the abandonment of traditional uses since the middle of the last century [18] has caused a decrease in anthropogenic pressure on Mediterranean forest ecosystems [8], being particularly important for mountain areas [7,15,16,19]. The dramatic rural exodus occurred in mountain areas due to changes in socio-economic conditions [20], resulting in the abandonment of traditional activities and significant environmental changes [15,16,17,18,21,22]. Moreover the abandonment of mountain agricultural areas is causing an increase in forest expansion via the spontaneous recovery of vegetation [15,17], which can causes a homogenization of the landscape [23] with several ecological consequences [22].
Quercus pyrenaica Willd. woodlands, like other forest formations in the Mediterranean region, have been subjected to intense anthropogenic pressures over time [24,25], which have led to the reduction in their distribution area, as well as in modifications of their floristic and structural patterns [26,27,28]. Historically, the woodlands of Q. pyrenaica have been exploited mainly for firewood, charcoal, and tannins [29,30]. Some areas were also burned and thinned to create pastures with low densities of mature trees that provide acorns, firewood, and large areas for grazing [31,32,33]. All these anthropogenic processes have fundamentally transformed the oak woodlands so that it is difficult to find stands that can be considered as natural forests [29]. Nonetheless, the decrease in anthropogenic pressure has paradoxically caused many of the Q. pyrenaica oak stands to present a state of advanced degradation, showing growth stagnation, a lack of fruiting, and also symptoms of branch dieback [34,35,36,37,38].
Therefore, understanding the woodland expansion towards abandoned croplands is critical to develop effective management strategies, particularly for populations at the rear edge of their distribution [39], which deserve particular attention due to their high conservation value [40]. The study of ecological dynamics within the rear-edge populations is considered essential to establish proper management guidelines under current climate uncertainties [40,41]. Rear-edge populations are often adapted to local environmental conditions at the limit of the species ecological amplitude, and often show a long-term persistence [39]. Local responses to environmental changes may differ from the species’ mean response [42,43,44], and such differences may either promote or hamper the survival of edge populations under global change [40,41]. In this work, we analyze the colonization pattern of abandoned croplands by Quercus pyrenaica in the Sierra Nevada mountain region. This mountain has undergone significant land-use changes in the last 50 years [45] with increases in forest densities [19], and tree growth [46]. We are interested in exploring the colonization pattern of abandoned croplands by Q. pyrenaica on both slopes of Sierra Nevada. We aimed to assess differences among populations in the rear edge of its distribution. Our specific goals are: (i) to analyze the colonization pattern of abandoned cropland by Q. pyrenaica and its relationship with time after abandonment; (ii) to explore differences in the structures of the seed source (mature woodlands); and (iii) to compare the abundance of the main seed disperser (Eurasian jay, Garrulus glandarius).
2. Materials and Methods
2.1. Sampling Description
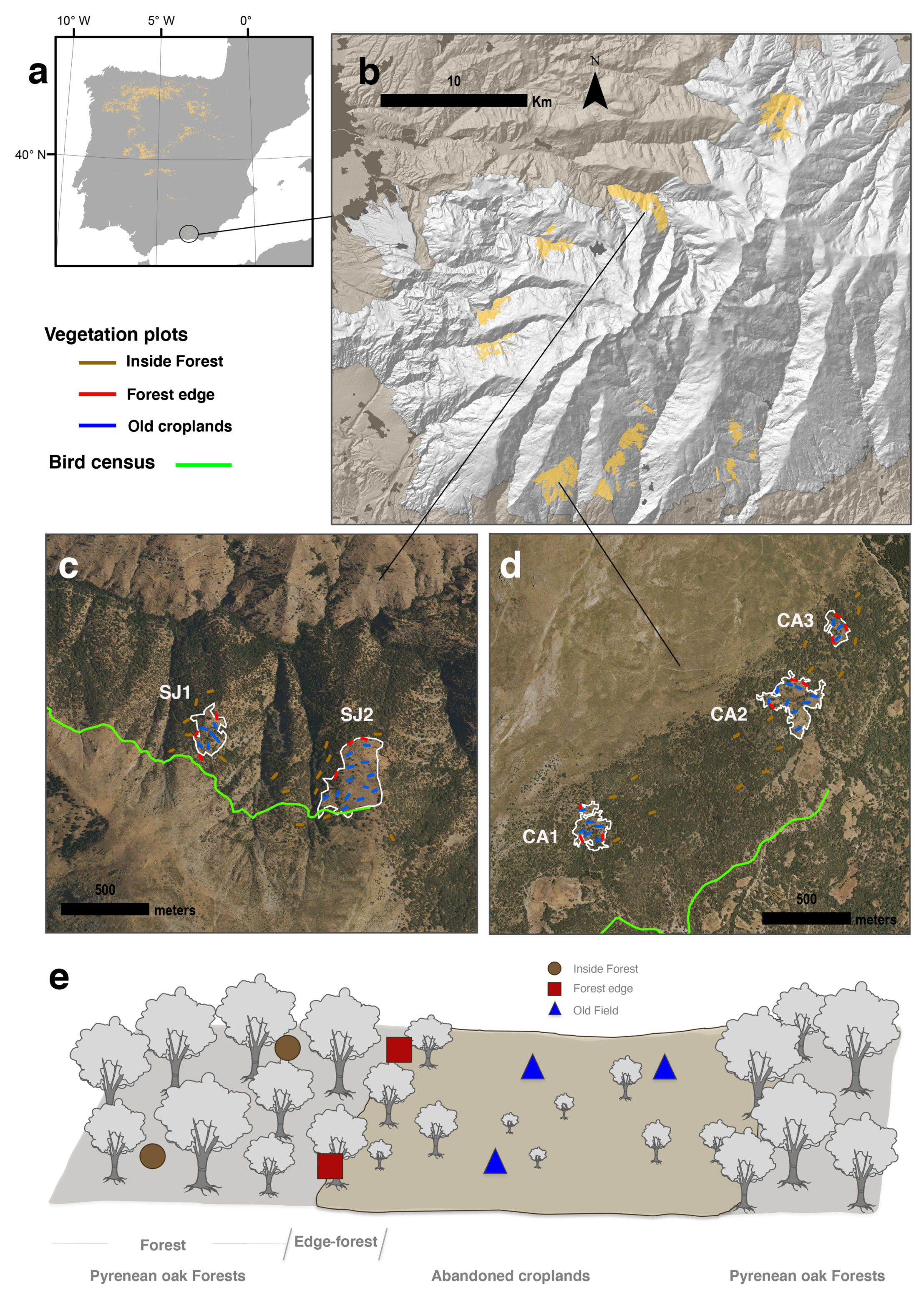
We sampled 5 abandonment croplands located at two Pyrenean oak forests in contrasting slopes of Sierra Nevada (southern Spain): Robledal de San Juan (SJ), located at the northern aspect (37°7′29.63″ N, 3°21′54.60″ W; Güejar-Sierra, Granada, Spain); and Robledal de Cáñar (CA), located at the southern aspect (37°57′28.04″ N, 3°25′57.1″ W; Cáñar, Granada). We selected oak populations located at contrasting slopes since differences in environmental variables have been reported for these oak woodlands [47]. Each cropland was delimited using land-use and land-cover maps of Andalusia for 1956 [48], combined with a detailed photographic interpretation of the black and white 1956 orthophotos (1 m spatial resolution) (see the work of Navarro-González et al. [49] for more details). The estimation of the age abandonment for each cropland was performed combining the interpretation of orthophotographies with information from local neighbors. We compiled all the available aerial ortophotographies of the study areas from Fototeca Digital of the Spanish National Geographic Institute (http://fototeca.cnig.es/, accessed on 12 August 2018). These dates were verified using information about past land use, compiled from local neighbours (by local workshops and interviews with retired elders: farmers, shepherds and loggers; see details in [50,51]). The estimated rank of ages could be considered accurate (see Table 1).
For each abandonment cropland, vegetation plots (30 m × 10 m) were randomly distributed in the old field; at the forest edges; and inside the surrounding forests (Figure 1). The number of plots within the old fields and surrounding forests were proportional to the size of the abandonment cropland (Table 1). A total of 83 vegetation plots were sampled in autumn 2012. In each vegetation plot, all tree species were recorded, and tree height and diameter were measured. For each plot, we computed the juvenile abundance as the number of individuals smaller than 150 cm on height. We did not distinguish the reproductive and vegetative origins of young oaks, since it is difficult due to the resprouting trait of this species. In addition to the juvenile abundance, we explored differences between several recruitment stages based on individual size (see [52]). We considered five size categories based on height (every 30 cm). All data were properly documented and published in an international repository (see [53] for a detailed description of the dataset).
To explore the main bird disperser in our study sites, we used bird censuses carried out by the Sierra Nevada Global Change Observatory (https://obsnev.es/, accessed on 12 August 2014). This dataset contains bird censuses at different ecosystems types of Sierra Nevada, starting from 2008 (see [54,55] for more details). We only used data for the Eurasian jay (Garrulus glandarius), since it is the main disperser of Quercus pyrenaica [56]. We assumed that jays can move acorns into abandoned croplands based on the habitat preferences to cache acorns reported by Pons and Pausas [57], and also considering preliminary data on jays flights carried out in our study sites by Zamora et al. [58], who found a high proportion of flights of this bird at the same elevation, and around 40% of open areas as arrival habitat. Since we were interested in the comparison of the Eurasian jay populations between the two study sites, we computed the annual bird abundances (in terms of birds/10 ha) for each site over a 7-year period (2008–2013). The sampling procedure was the line-transect method with a bandwith of 50 m (25 m on each side). Transects length were 2.80 km for Cáñar site (CA), and 3.22 km for San Juan site (SJ). Sight and sound records within the sample area were accepted as contacts. All transects were sampled in the early morning. Eurasian jay abundance was calculated in terms of birds/10 ha. All counts in one month were averaged, and the yearly result was obtained from the average of all the months studied. For more details about bird censuses, see [54,59].
2.2. Data Analysis
We used the vegetation plots carried out inside the forest (habitat type = forest) to analyze the structure of the seed source (surrounding forest). Several parameters related to forest structure and functioning were computed: tree density, juvenile abundance, tree species composition, tree size related statistics (i.e., mean, median, maximum, 75 and 90 percentiles of tree-height), and basal area (BA). Differences between sites were assessed using the non-parametric Mann–Whitney U-test, since data did not meet normality either homocedasticity assumptions. We also compared whether there was variation within plots belonging to the same sites. ANOVA analyses were performed to explore the differences of bird disperser abundance (G. glandarius) between sites and across years.
The variation of the juvenile abundance between study sites, habitat type, and their interaction (site-habitat type), was analyzed using generalized linear models with a Tweedie distribution with a log link [60]. Study sites and habitat type were the explanatory variables. Prior to the analysis, data exploration was applied following protocols described by [61,62]. As the dataset comprised count data, we initially used the Poisson and the negative binomial distribution. However, these models were overdispersed. A variance power parameter of 1.28 (1.20–1.40, 95% confidence interval) was used in the Tweedie GLM model. This parameter was estimated using the tweedie.profile function of the tweedie R package [63,64]. Model comparison (univariate models) was carried out using the Akaike’s information criterion (AIC) [65]. The model accuracy was tested by Nagelkerke’s pseudo-, used as a measure of goodness of fit. The significance of the explanatory variables in the selected model was tested using the likelihood ratio tests (LRT). Wald z-tests and Tukey’s HSD-corrected post hoc comparisons were used to test for differences in juvenile abundance among sites and habitat type.
3. Results
The forest structure of Quercus pyrenaica woodlands did not show significant differences for the forest attributes between study sites (Table 2). Quercus pyrenaica woodlands of southern site (CA) showed higher tree density but smaller tree heights (mean, median and percentiles) than those on the northern site (SJ) (Table 2). In addition, higher abundance of juveniles was found for CA site, which also showed greater basal area than SJ site (Table 2).
Regarding the abundance of Garrulus glandarius, no differences between study sites were found ( = 2.387; p = 0.126; CA = 1.69 ± 0.21 and SJ = 1.33 ± 0.22 birds/10 ha), neither across years in the studied period (2008–2014) ( = 1.234; p = 0.297). The interaction term was also not significant ( = 1.26; p = 0.284).
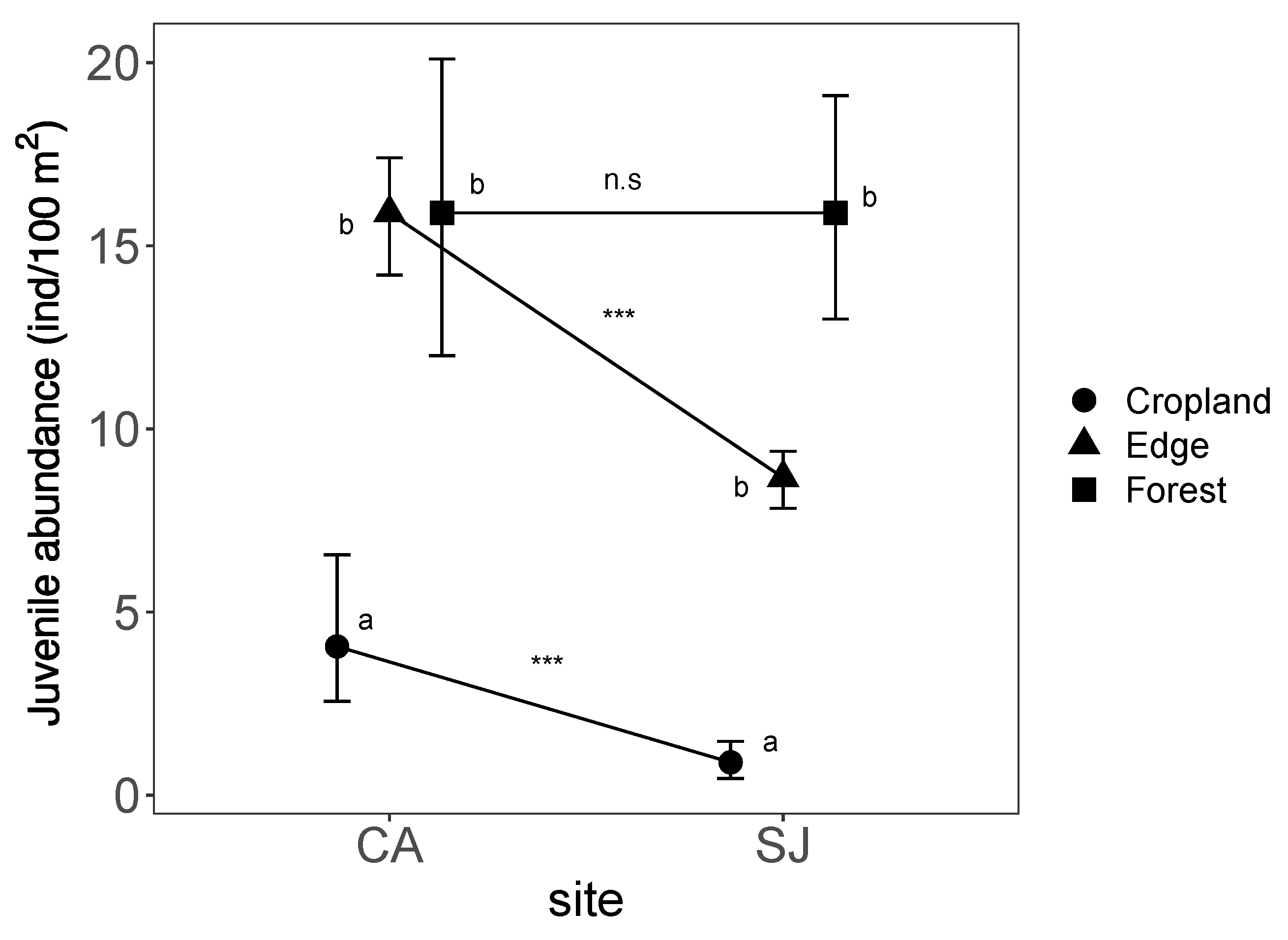
The juvenile oak abundance model including all terms (i.e., full model) showed a higher strength of empirical support than did models for each of the independent variables (i.e., univariate models) (Table 3). Oak juvenile abundance differed among habitat types ( = 72.95; p < 0.0001), and between study sites ( = 8.16; p = 0.0054; Table 4).
A decreasing gradient of oak-juvenile abundance was found across habitat type, from higher values in forest type (15.90 ± 1.30 juvenile/100 m) to lower values inside the old croplands (2.43 ± 0.55 juvenile/100 m; Figure 2). The abundance of oak juveniles in the old croplands of the southern site (CA) was significantly higher (4.06 ± 0.98 juvenile/100 m) than in those of the northern site (SJ; 0.90 ± 0.26 juvenile/100 m).
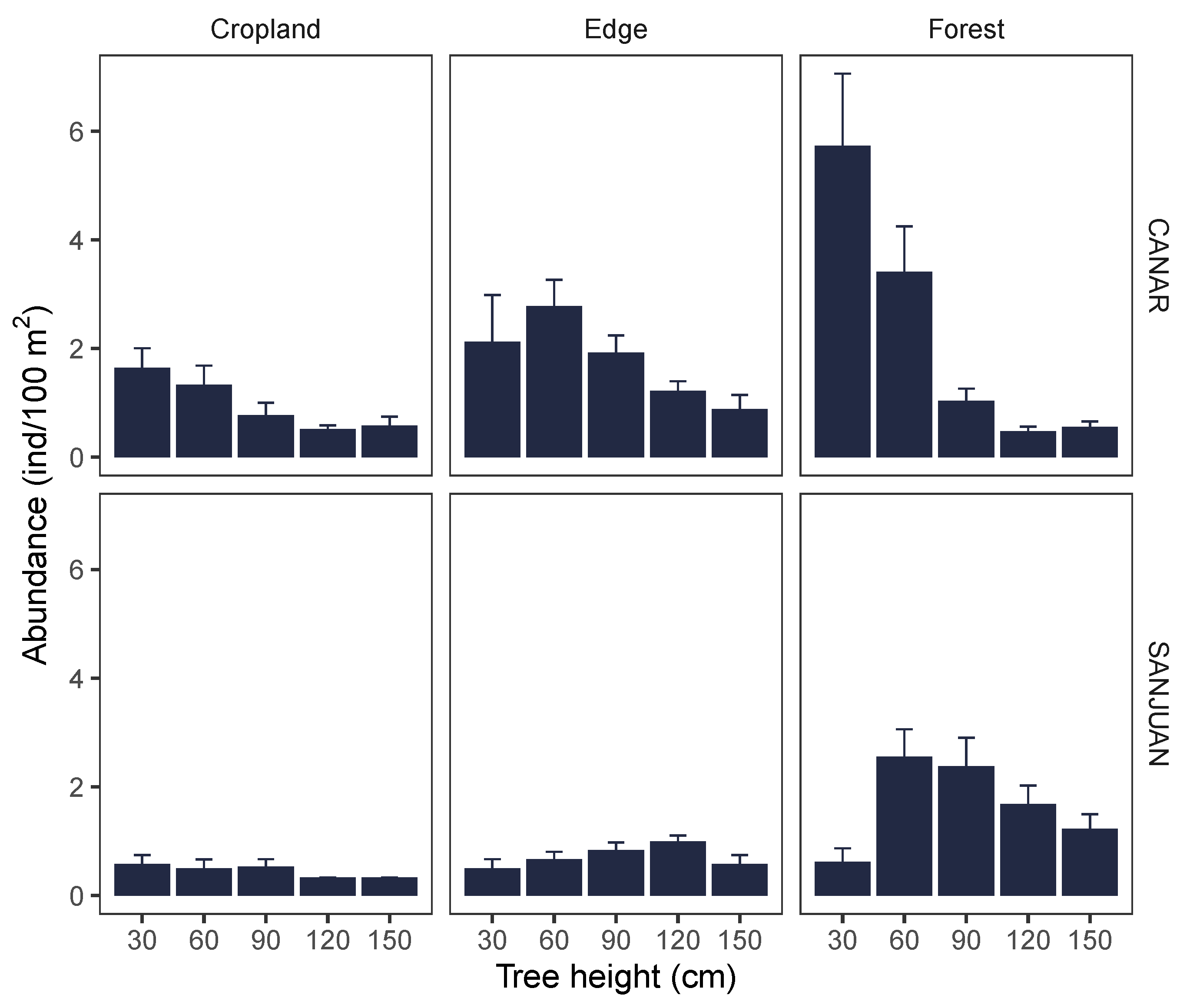
The size distribution of juveniles was also different among the study sites (Figure 3). An even size distribution of the oak juveniles was observed in the old croplands of northern site. Conversely, higher contribution of small oak juveniles (<30 cm) were found at old croplands of the southern site (Figure 3).
A positive relation was found between the oak juvenile abundance and the estimate age of crop abandonment (Figure 4) with higher oak juvenile abundances in the earlier abandoned croplands.
4. Discussion
We observed a colonization process of Quercus pyrenaica into abandoned croplands in this mountain region despite the strong recruitment constraints described for this species [35,66,67]. Forest expansion towards abandoned croplands has been recorded in several marginal habitats in other European mountainous regions [15,16,17,68,69,70,71,72], as consequence mainly of rural depopulation and decrease in herbivores pressure [18,73]. Our results show a relationship between juvenile oak abundance and the estimated age of crop abandonment (Figure 4). As the time after crop abandonment increases, species heterogeneity and functional diversity of the ecosystem increases [74,75], boosting the multifuntionality of the ecosystems [76].
It has been also reported that both age of the surrounding forests and previous land use could influence the colonization pattern [77]. In this sense, it is expected that the colonization process will continue, since the forests surrounding our study areas are relatively young. Dendrochronological estimates of the forest age for our study woodlands, ranged approximately 90–100 and 200 years for northern and southern oak populations, respectively [46,78], which is younger than the estimated ages for these forests along their distribution range [78]. Cruz-Alonso et al. [76], in a study of the recovery of multifunctionality in Mediterranean forests, reported for this oak species, a minimum of 80 years after the abandonment for the recovery of the reference multifunctionality.
The colonization pattern of abandoned croplands varies within the rear edge of the Q. pyrenaica. Our results showed different abundances of oak juveniles between sites, with higher abundances at the southern sites (CA) (Table 4; Figure 2). It is known that the distance to the source and the structure of the seed source influenced the propagule input into new habitats [79,80,81]. In our study sites, all the abandoned croplands are surrounded by native forests. Thus, the observed differences in oak juvenile abundance would not seem to be explained by the distance to the seed source (Table 1). Regarding the structure of the seed source, we found no differences between the study sites (Table 2). Forest attributes potentially related to the acorn production (e.g., tree density, basal area) [82] are not different for the surrounding forests between study sites (Table 2). Although in our work we have not carried out an estimation of acorn production that would allow us to compare between study sites, previous studies analyzing the variation in reproductive parameters and comparing the acorn production across oak woodlands of Sierra Nevada, found slight differences among oak populations, with a higher acorn yield for southern oak woodlands than northern one [83]. However, the data series used by Leal [83] was very short (only two years) and Quercus pyrenaica have a marked mast-seeding behaviour [35,84]. In addition to this, there do not seem to be marked differences in the current seed source that could explain our observed differences in the abundance of juveniles in the abandoned crops.
Another key aspect for the tree colonization of abandoned croplands is the dispersion vectors. Q. pyrenaica acorn are mainly dispersed by woodmouse (Apodemus sylvaticus) and Eurasian jay (Garrulus glandarius) [66,67]. We used a time series (a 7-year period) to analyze the abundance of jays at each study site. The aim of using this time series was to explore whether there were differences between study sites with respect to the abundance of this bird acorn disperser. We are aware of the limitation of using these data, i.e., we do not know if the abundance of jays at both sites has followed the same temporal trajectory, but we have used the longest time series we have for both sites, to at least find out if there are differences between the sites in recent years. Our results showed similar abundances of this acorn disperser between the two study sites. Therefore, the observed differences in oak juvenile abundance do not appear to be explained by differences in abundance of this acorn disperser. Having observed no differences in the seed source neither in the main dispersal vector, a logical next step would be to explore differences in the seed arrival site.
For the recolonization of abandoned croplands, in addition to the importance of factors related to dispersal in time (seed bank related) and space (distance related), the previous use to which the crop field has been subjected is a key factor determining the abundance of native tree species [75,85]. The colonization pattern of woody species is affected by fine-scale variations in abiotic factors [86,87], but it has been observed that land-use history mainly controlled the forest expansion rates [15,88]. The forest history of our study sites, inferred from several compiling studies [45,46,50,51,89,90], indicated that both sites were subjected to intense anthropic uses in the past. At the northern site (SJ), uplands areas were dedicated to grazing, and in the forest areas there were also some croplands with grazing. In addition, timber extraction for mining was recorded at this site. The southern site (CA) has been exploited for firewood, charcoal and acorns, with less presence of livestock use. Although we could not estimate the intensity of use to which both zones have been subjected before the abandonment of crops, the northern zone seems to have had a management history with a higher grazing intensity than the southern site [50,51,91]. In addition to the land-use legacies before the cropland abandonment, another relevant question would be the management history after the crop cessation, focusing on the livestock pressure, since herbivory imposes severe constraints to the establishment and regeneration of this oak species [66,67]. There are no data available on the temporal evolution of grazing pressure in detail in our study sites. Only Robles-Cruz [92] showed a general quantification for several ecosystems of Sierra Nevada. Notwithstanding, several studies and reports, combining interviews with shepherds and reviews of historical documents, have determined the recent livestock history in several oak woodlands of the Sierra Nevada [50,51,91]. For our study sites, both higher numbers of herds and sheperds, and more livestock density in the northern site (SJ) than in the southern one (CA), have been observed, which could be translated into a greater herbivore pressure that would explain our observed differences in juvenile oak abundance within the abandoned croplands. This distinct herbivory pressure in the two study sites could explain the differences observed in the juvenile oak abundance not only at cropland habitat type, either both at edge and forest habitat type. The exploration of abundance juveniles by size categories between habitats type for each study site showed lower abundance values for the northern site (Figure 3). The size-age structure is also more homogeneous at the northern site, reinforcing the hypothesis of higher herbivore pressure. Gómez et al. [66] found that herbivory rather than abiotic factors are the main cause of seedling mortality of Q. pyrenaica in Sierra Nevada. Thus, herbivores killed most of the seedlings, although the way in which they severed the seedlings was very diverse [66]. They found that trampling by livestock and acorn predation by wild boars and hares was the main cause of the mortality observed, but the browsing by livestock is marginal (only 1% of the mortality) [66]. Another factor to consider is the presence of shrubs which can act as nurse plants. Facilitation by nurse plants has been reported as an essential process for the regeneration of some tree species [93,94]. The survival of Q. pyrenaica substantially increases when it is under individual pioneer shrubs [93,95]. Shrubs may protect Q. pyrenaica seedlings from the browsing and trampling of vertebrate herbivores. They also offer safe sites that reduce the high mortality rates during the summer in the early stages of recruitment [93,96]. Although we did not estimate the cover and diversity of shrub species in our studied abandoned crops, previous studies in the surrounding forests of the same localities did not find differences in the diversity and richness of shrubs within oak forests [97].
5. Concluding Remarks
The results of our study show that, even in the current increasingly dry climatic conditions, Q. pyrenaica woodlands are able to recover the abandoned, formerly arable fields at the same attitudinal level where oak woodland can potentially exist. Thus, the Pyrenean oak woodland is clearly expanding in the Sierra Nevada mountain range, a rear edge of the distribution of this oak species. This natural process can provide solutions for conserving biodiversity, and enhances the mitigation of and adaptation to climate change (see [98] and references therein). In addition to this, active restoration could aid in the recovery of the oak forest’s multifunctionality [76] in the abandoned croplands that are being colonized by tree species.
The differences in the recolonization patterns within the rear edge seem be related to differences in the management prior to and after the abandonment of mountain croplands. A higher herbivory pressure after cropland abandonment seems to limit the forest expansion towards marginal habitats. Related to this, and in order to improve the forest expansion, it would be recommended to take advantage of the presence of native shrubs that offer safe sites. These safe sites could aid in the reduction in the mortality of Q. pyrenaica seedlings, and therefore increase the chances of establishing this oak species. This would also aid in increasing heterogeneity within the ongoing secondary forest, which also increases the resilience to perturbations and the recovery of the ecosystem’s multifunctionality [76,99]. On the other hand, attention also needs to be paid to maintaining healthy seed sources (surroundings forests) and a stable seed disperser community, particularly the Eurasian jay, since acorn dispersion by this bird species is considered a key process in the regeneration of Quercus forests after land abandonment [100].
Author Contributions
A.J.P.-L.: Conceptualization, Methodology, Software, Formal analysis, Data curation, Writing—original draft, Writing—review and editing, and Visualization; F.J.B.-G.: Conceptualization, Investigation, Writing—review and editing, and Funding acquisition; R.Z.: Conceptualization, Methodology, Investigation, Writing—review and editing, and Funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was partially funded by the LIFE-ADAPTAMED (LIFE14CCA/ES/000612) project, by MIGRAME Project (Excellence Research Group Programme of the Andalusian Government (RNM 6734), and by eLTER H2020 project.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data used in this work have been properly documented and uploaded to the GBIF repository. They are available at https://0-doi-org.brum.beds.ac.uk/10.15470/orboj4 and https://0-doi-org.brum.beds.ac.uk/10.15468/ow9noo (accessed on 7 September 2021). Descriptions of those datasets are availables in Pérez-Luque et al. [53] and Pérez-Luque et al. [55], respectively.
Acknowledgments
This research work was conducted in the collaborative framework of the “Sierra Nevada Global Change Observatory” monitoring program (https://obsnev.es/, accessed on 7 September 2021). We would like to thank to I. Navarro, B. Benito, R. Pérez-Pérez, L. Matías, R. Ruiz-Puche, F. Suzart, R. Moreno-Llorca and J. Rodríguez-Infante for their help in the field work. We thank the LIFE-ADAPTAMED (LIFE14 CCA/ES/000612): Protection of key ecosystem services by adaptive management of Climate Change endangered Mediterranean socioecosystems for the funding support. The projects MIGRAME “Global Change, Migration and colonization of habitats altitudinal gradients on Mediterranean mountains” (RNM 6734) from the Excellence Research Group Programme of the Andalusian Government, and the H2020 project European Long-Term Ecosystem and socio-ecological Research Infrastructure (eLTER) provided partial funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global Biodiversity: Indicators of Recent Declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- Winkler, K.; Fuchs, R.; Rounsevell, M.; Herold, M. Global land use changes are four times greater than previously estimated. Nat. Commun. 2021, 12, 2501. [Google Scholar] [CrossRef]
- Sala, O.E. Global Biodiversity Scenarios for the Year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Lindenmayer, D.; Cunningham, S.; Young, A. Land Use Intensification: Effects on Agriculture, Biodiversity and Ecological Processes; CSIRO Publishing: Melbourne, Australia, 2012. [Google Scholar]
- Hasan, S.S.; Zhen, L.; Miah, M.G.; Ahamed, T.; Samie, A. Impact of land use change on ecosystem services: A review. Environ. Dev. 2020, 34, 100527. [Google Scholar] [CrossRef]
- Rey Benayas, J.M. Abandonment of agricultural land: An overview of drivers and consequences. CAB Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 2007, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Pías, B.; Escribano-Avila, G.; Virgós, E.; Sanz-Pérez, V.; Escudero, A.; Valladares, F. The colonization of abandoned land by Spanish juniper: Linking biotic and abiotic factors at different spatial scales. For. Ecol. Manag. 2014, 329, 186–194. [Google Scholar] [CrossRef]
- Valbuena-Carabaña, M.; de Heredia, U.L.; Fuentes-Utrilla, P.; González-Doncel, I.; Gil, L. Historical and recent changes in the Spanish forests: A socio-economic process. Rev. Palaeobot. Palynol. 2010, 162, 492–506. [Google Scholar] [CrossRef]
- Martínez-Fernández, J.; Ruiz-Benito, P.; Zavala, M.A. Recent land cover changes in Spain across biogeographical regions and protection levels: Implications for conservation policies. Land Use Policy 2015, 44, 62–75. [Google Scholar] [CrossRef]
- Rounsevell, M.; Reginster, I.; Araújo, M.; Carter, T.; Dendoncker, N.; Ewert, F.; House, J.; Kankaanpää, S.; Leemans, R.; Metzger, M.; et al. A coherent set of future land use change scenarios for Europe. Agric. Ecosyst. Environ. 2006, 114, 57–68. [Google Scholar] [CrossRef]
- Perpiña Castillo, C.; Jacobs-Crisioni, C.; Diogo, V.; Lavalle, C. Modelling agricultural land abandonment in a fine spatial resolution multi-level land-use model: An application for the EU. Environ. Model. Softw. 2021, 136, 104946. [Google Scholar] [CrossRef]
- Sheffer, E. A review of the development of Mediterranean pine–oak ecosystems after land abandonment and afforestation: Are they novel ecosystems? Ann. For. Sci. 2012, 69, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Debussche, M.; Lepart, J.; Dervieux, A. Mediterranean landscape changes: Evidence from old postcards. Glob. Ecol. Biogeogr. 1999, 8, 3–15. [Google Scholar] [CrossRef]
- Peñuelas, J.; Boada, M. A global change-induced biome shift in the Montseny mountains (NE Spain): A global change-induced biome shift. Glob. Chang. Biol. 2003, 9, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Martínez, J.M.; Suárez-Seoane, S.; Stoorvogel, J.J.; de Luis Calabuig, E. Influence of land use and climate on recent forest expansion: A case study in the Eurosiberian—Mediterranean limit of north-west Spain. J. Ecol. 2014, 102, 905–919. [Google Scholar] [CrossRef] [Green Version]
- de Natale, F.; Gasparini, P.; Carriero, A. A study on tree colonization of abandoned land in the Italian Alps: Extent and some characteristics of new forest stands in Trentino. In Sustainable Forestry: From Monitoring and Modelling to Knowledge Management and Policy Science; Reynolds, K.M., Thomson, A.J., Köhl, M., Shannon, M.A., Ray, D., Rennolls, K., Eds.; CABI: Wallingford, UK, 2007; pp. 269–284. [Google Scholar]
- Piussi, P. Expansion of European mountain forests. In Forests in Sustainable Mountain Development: A State of Knowledge Report for 2000. Task Force on Forests in Sustainable Mountain Development; CABI: Wallingford, UK, 2000; pp. 19–28. [Google Scholar]
- MacDonald, D.; Crabtree, J.; Wiesinger, G.; Dax, T.; Stamou, N.; Fleury, P.; Gutierrez Lazpita, J.; Gibon, A. Agricultural abandonment in mountain areas of Europe: Environmental consequences and policy response. J. Environ. Manag. 2000, 59, 47–69. [Google Scholar] [CrossRef]
- Jiménez-Olivencia, Y.; Porcel, L.; Caballero, A. Medio siglo en la evolución de los paisajes naturales y agrarios de Sierra Nevada (España). Boletín de la Asociación de Geógrafos Españoles 2015, 68, 205–232. [Google Scholar] [CrossRef] [Green Version]
- European Environment Agency. Europe’s Ecological Backbone: Recognising the True Value of Our Mountains; Publications Office: København, Denmark, 2010. [Google Scholar]
- Rutherford, G.N.; Bebi, P.; Edwards, P.J.; Zimmermann, N.E. Assessing land-use statistics to model land cover change in a mountainous landscape in the European Alps. Ecol. Model. 2008, 212, 460–471. [Google Scholar] [CrossRef]
- Zimmermann, P.; Tasser, E.; Leitinger, G.; Tappeiner, U. Effects of land-use and land-cover pattern on landscape-scale biodiversity in the European Alps. Agric. Ecosyst. Environ. 2010, 139, 13–22. [Google Scholar] [CrossRef]
- Mietkiewicz, N.; Kulakowski, D.; Rogan, J.; Bebi, P. Long-term change in sub-alpine forest cover, tree line and species composition in the Swiss Alps. J. Veg. Sci. 2017, 28, 951–964. [Google Scholar] [CrossRef]
- García, I.; Jiménez, P. 9230 Robledales de Quercus pyrenaica y robledales de Quercus robur y Quercus pyrenaica del noroeste ibérico. In Bases Ecológicas Preliminares para la Conservación de los Tipos de Hábitat de Interés Comunitario en España; VV.AA., Ed.; Ministerio de Medio Ambiente, y Medio Rural y Marino: Madrid, Spain, 2009; pp. 1–66. [Google Scholar]
- Alba-Sánchez, F.; Abel-Schaad, D.; López-Sáez, J.A.; Sabariego-Ruiz, S.; Pérez-Díaz, S.; Luelmo-Lautenschlaeger, R.; Garrido-García, J.A. Early anthropogenic change in western Mediterranean mountains (Sierra Nevada, SE Spain). Anthropocene 2021, 33, 100278. [Google Scholar] [CrossRef]
- Gavilán, R.G.; Escudero, A.; Rubio, A. Effects of disturbance on floristic patterns of Quercus pyrenaica forests in Central Spain. In Vegetation Science in Retrospect and Perspective—Proceedings 41st IAVS Symposium; International Association for Vegetation Science: Montpellier, France, 2000; pp. 226–229. [Google Scholar]
- Calvo, L.; Tárrega, R.; de Luis, E. Post-fire succession in two Quercus pyrenaica communities with different disturbance histories. Ann. For. Sci. 1999, 56, 441–447. [Google Scholar] [CrossRef] [Green Version]
- Tárrega, R.; Calvo, L.; Marcos, E.; Taboada, A. Forest structure and understory diversity in Quercus pyrenaica communities with different human uses and disturbances. For. Ecol. Manag. 2006, 227, 50–58. [Google Scholar] [CrossRef]
- Ruiz de la Torre, J. Flora Mayor; Organismo Autónomo Parques Nacionales: Madrid, Spain, 2006. [Google Scholar]
- Sánchez Palomares, O.; Roig, S.; del Río, M.; Rubio-Sánchez, A.; Gandullo, J.M. Las Estaciones Ecológicas Actuales y Potenciales de los Rebollares Españoles; Number nº 17 in Monografias INIA, Serie Forestal; Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria: Madrid, Spain, 2008. [Google Scholar]
- Herrera Calvo, P.M. El uso pastoral como alternativa de gestión de los hábitats vinculados a los rebollares ibéricos. Rev. Pastos 2016, 46, 6–23. [Google Scholar]
- Álvarez, R.; García-Duro, J.; Muñoz, J.; Pesqueira, X.; Rivas, M.; Reyes, O.; Casal, M. Cambios en la estructura y diversidad, y en los modelos de combustible del bosque de Quercus pyrenaica tras fuego. In Proceedings of the V Congreso Forestal Español, Ávila, Spain, 21–25 September 2009. [Google Scholar]
- Valbuena-Carabaña, M.; Gil, L. Centenary coppicing maintains high levels of genetic diversity in a root resprouting oak (Quercus pyrenaica Willd.). Tree Genet. Genomes 2017, 13, 28. [Google Scholar] [CrossRef]
- Cañellas, I.; Del-Río, M.; Roig, S.; Montero, G. Growth response to thinning in Quercus pyrenaica Willd. coppice stands in Spanish central mountain. Ann. For. Sci. 2004, 61, 243–250. [Google Scholar] [CrossRef] [Green Version]
- Bravo, J.A.; Roig, S.; Serrada, R. Selvicultura en montes bajos y medios de Quercus ilex L., Q. pyrenaica Willd. y Q. faginea Lam. In Compendio de Selvicultura Aplicada en España; Serrada, R., Montero, G., Reque, J.A., Eds.; INIA: Madrid, Spain, 2008; pp. 657–744. [Google Scholar]
- Valbuena-Carabaña, M.; Gil, L. Efectos de la gestión selvícola pasada y presente sobre la diversidad genética actual y futura de Quercus pyrenaica Willd. en Sierra Nevada. Ecosistemas 2014, 23, 90–98. [Google Scholar]
- Piqué, M.; Vericat, P. Evolution and perspectives of coppice forests in European countries and South Africa. In FPS COST Action FP1301: Innovative Management and Multifunctional Utilization of Traditional Coppice Forests? An Answer to Future Ecological, Economic and Social Challenges in the European Forestry Sector (EuroCoppice); Bartlett, D., Nicolescu, V.N., Pyttel, P., Eds.; 2015; pp. 38–40. Available online: https://gala.gre.ac.uk/id/eprint/14009/ (accessed on 1 May 2018).
- Piqué, M.; Laina, R.; Vericat Grau, P.; Beltrán, M.; Busquets, E.; Tolosana, E. Spain. In Coppice Forests in Europe; Unrau, A., Becker, G., Spinelli, R., Lazdina, D., Magagnotti, N., Nicolescu, V., Buckley, P., Barlett, D., Kofman, P., Eds.; Albert Ludwig University of Freiburg: Freiburg im Breisgau, Germany, 2018; pp. 331–336. [Google Scholar]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef] [Green Version]
- Fady, B.; Aravanopoulos, F.A.; Alizoti, P.; Mátyás, C.; von Wühlisch, G.; Westergren, M.; Belletti, P.; Cvjetkovic, B.; Ducci, F.; Huber, G.; et al. Evolution-based approach needed for the conservation and silviculture of peripheral forest tree populations. For. Ecol. Manag. 2016, 375, 66–75. [Google Scholar] [CrossRef] [Green Version]
- Jump, A.S.; Cavin, L.; Hunter, P.D. Monitoring and managing responses to climate change at the retreating range edge of forest trees. J. Environ. Monit. 2010, 12, 1791–1798. [Google Scholar] [CrossRef]
- Benavides, R.; Rabasa, S.G.; Granda, E.; Escudero, A.; Hódar, J.A.; Martínez-Vilalta, J.; Rincón, A.M.; Zamora, R.; Valladares, F. Direct and indirect effects of climate on demography and early growth of Pinus sylvestris at the rear edge: Changing roles of biotic and abiotic factors. PLoS ONE 2013, 8, e59824. [Google Scholar] [CrossRef] [Green Version]
- Matías, L.; Linares, J.C.; Sánchez-Miranda, A.; Jump, A.S. Contrasting growth forecasts across the geographical range of Scots pine due to altitudinal and latitudinal differences in climatic sensitivity. Glob. Chang. Biol. 2017, 23, 4106–4116. [Google Scholar] [CrossRef] [PubMed]
- Castro, J.; Zamora, R.; Hódar, J.A.; Gómez, J.M. Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: Consequences of being in a marginal Mediterranean habitat. J. Ecol. 2004, 92, 266–277. [Google Scholar] [CrossRef]
- Jiménez-Olivencia, Y.; Porcel-Rodríguez, L.; Caballero-Calvo, A.; Bonet, F.J. Evolución de los usos del suelo en Sierra Nevada en los últimos 50 años y cambios en el paisaje. In La Huella del Cambio Global en Sierra Nevada: Retos Para la Conservación; Zamora, R., Pérez-Luque, A.J., Bonet, F., Barea-Azcón, J., Aspizua, R., Eds.; Junta de Andalucía, Consejería de Medio Ambiente y Ordenación del Territorio: Seville, Spain, 2015; pp. 54–56. [Google Scholar]
- Pérez-Luque, A.; Gea-Izquierdo, G.; Zamora, R. Land-use legacies and climate change as a double challenge to oak forest resilience: Mismatches of geographical and ecological rear edges. Ecosystems 2021, 24, 755–773. [Google Scholar] [CrossRef]
- Pérez-Luque, A.; Benito, B.M.; Bonet-García, F.J.; Zamora, R. Ecological Diversity within Rear-Edge: A Case Study from Mediterranean Quercus pyrenaica Willd. Forests 2021, 12, 10. [Google Scholar] [CrossRef]
- CMA. Mapa de usos y Coberturas de Andalucía 1956–1999–2003, 1:25,000; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2007. [Google Scholar]
- Navarro-González, I.; Bonet-García, F.; Pérez-Luque, A. Cartografía histórica de la vegetación mediante ortofotos. In Observatorio de Cambio Global de Sierra Nevada. Metodologías de Seguimiento; Aspizua, R., Barea-Azcón, J., Bonet, F., Pérez-Luque, A., Zamora, R., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2012; pp. 16–17. [Google Scholar]
- Moreno-Llorca, R.A.; Pérez-Luque, A.J.; Bonet, F.J.; Pérez-Pérez, R.; Zamora, R. Caracterización de fuentes de información para la reconstrucción histórica de la vegetación. Un caso de estudio en Sierra Nevada. In Proceedings of the XII Congreso Nacional de Medio Ambiente (CONAMA 2014), Madrid, Spain, 24–27 November 2014. [Google Scholar]
- Moreno-Llorca, R.; Pérez-Luque, A.; Bonet, F.; Zamora, R. Historical analysis of socio-ecological changes in the municipality of Cáñar (Alpujarra, Sierra Nevada) over the last 5 centuries. In Global Change Impacts in Sierra Nevada: Challenges for Conservation; Zamora, R., Pérez-Luque, A., Bonet, F., Barea-Azcón, J., Aspizua, R., Eds.; Consejería de Medio Ambiente y Ordenación del Territorio, Junta de Andalucía: Sevilla, Spain, 2016; pp. 59–62. [Google Scholar]
- Plieninger, T.; Rolo, V.; Moreno, G. Large-Scale Patterns of Quercus ilex, Quercus suber, and Quercus pyrenaica Regeneration in Central-Western Spain. Ecosystems 2010, 13, 644–660. [Google Scholar] [CrossRef]
- Pérez-Luque, A.J.; Zamora, R.; Bonet, F.J.; Pérez-Pérez, R. Dataset of MIGRAME project (global change, altitudinal range shift and colonization of degraded habitats in Mediterranean mountains). PhytoKeys 2015, 56, 61–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barea-Azcón, J.; Martín-Jaramillo, J.; López-Sanjuan, R. Passeriformes y otras aves. In Observatorio de Cambio Global de Sierra Nevada. Metodologías de Seguimiento; Aspizua, R., Barea-Azcón, J., Bonet, F., Pérez-Luque, A., Zamora, R., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2012; pp. 86–87. [Google Scholar]
- Pérez-Luque, A.J.; Barea-Azcón, J.M.; Álvarez-Ruiz, L.; Bonet-García, F.J.; Zamora, R. Dataset of Passerine bird communities in a Mediterranean high mountain (Sierra Nevada, Spain). ZooKeys 2016, 552, 137–154. [Google Scholar] [CrossRef] [Green Version]
- Gómez, J.M. Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography 2003, 26, 573–584. [Google Scholar] [CrossRef] [Green Version]
- Pons, J.; Pausas, J.G. Acorn dispersal estimated by radio-tracking. Oecologia 2007, 153, 903–911. [Google Scholar] [CrossRef]
- Zamora, R.; Pérez-Luque, A.J.; Benito, B.M.; Bonet-García, F.J.; Navarro, I.; Pérez-Pérez, R.; Hódar, J.A.; Matías, L. Cambio Global, Migración Altitudinal y Colonización de Hábitats Degradados en Montañas Mediterráneas (MIGRAME); Invitación a la Ecología; Asociación Española de Ecología Terrestre: Pamplona, Spain, 2013. [Google Scholar]
- Zamora, R.; Barea-Azcón, J.M. Long-term changes in mountain passerine bird communities in the Sierra Nevada (southern Spain): A 30-year case study. Ardeola 2015, 62, 3–18. [Google Scholar] [CrossRef]
- Dunn, P.K.; Smyth, G.K. Tweedie GLMs. In Generalized Linear Models With Examples in R; Springer Texts in Statistics; Springer: New York, NY, USA, 2018; pp. 457–490. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Ieno, E.N.; Zuur, A.F. A Beginner’S Guide to Data Exploration and Visualization with R; Highland Statistics Ltd.: Newburgh, UK, 2015. [Google Scholar]
- Dunn, P.K.; Smyth, G.K. Series evaluation of Tweedie exponential dispersion models. Stat. Comput. 2005, 15, 267–280. [Google Scholar] [CrossRef] [Green Version]
- Dunn, P.K. Tweedie: Evaluation of Tweedie Exponential Family Models, R package version 2.3.3; 2017. Available online: https://cran.r-project.org/web/packages/tweedie/tweedie.pdf (accessed on 7 September 2021).
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd ed.; Springer: New York, NY, USA, 2010. [Google Scholar]
- Gómez, J.; García, D.; Zamora, R. Impact of vertebrate acorn- and seedling-predators on a Mediterranean Quercus pyrenaica forest. For. Ecol. Manag. 2003, 180, 125–134. [Google Scholar] [CrossRef]
- Perea, R.; San Miguel, A.; Gil, L. Interacciones planta-animal en la regeneración de Quercus pyrenaica: Ecología y gestión. Ecosistemas 2014, 23, 18–26. [Google Scholar]
- Ameztegui, A.; Coll, L.; Brotons, L.; Ninot, J.M. Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees. Glob. Ecol. Biogeogr. 2016, 25, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Améztegui, A.; Brotons, L.; Coll, L. Land-use changes as major drivers of mountain pine (Pinus uncinata Ram.) expansion in the Pyrenees: Land-use changes drive Pinus uncinata expansion. Glob. Ecol. Biogeogr. 2010, 19, 632–641. [Google Scholar] [CrossRef] [Green Version]
- Lasanta-Martínez, T.; Vicente-Serrano, S.M.; Cuadrat-Prats, J.M. Mountain Mediterranean landscape evolution caused by the abandonment of traditional primary activities: A study of the Spanish Central Pyrenees. Appl. Geogr. 2005, 25, 47–65. [Google Scholar] [CrossRef]
- Kozak, J. Forest cover change in the western carpathians in the past 180 years: A case study in the Orawa region in Poland. Mt. Res. Dev. 2003, 23, 369–375. [Google Scholar] [CrossRef] [Green Version]
- Vicente-Serrano, S.M.; Lasanta, T.; Romo, A. Analysis of spatial and temporal evolution of vegetation cover in the spanish central pyrenees: Role of human management. Environ. Manag. 2004, 34, 802–818. [Google Scholar] [CrossRef]
- European Environment Agency. European Forest Ecosystems State and Trends; EUR-OP: København, Denmark, 2016. [Google Scholar]
- Puerta-Piñero, C.; Espelta, J.M.; Sánchez-Humanes, B.; Rodrigo, A.; Coll, L.; Brotons, L. History matters: Previous land use changes determine post-fire vegetation recovery in forested Mediterranean landscapes. For. Ecol. Manag. 2012, 279, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Hermy, M.; Verheyen, K. Legacies of the past in the present-day forest biodiversity: A review of past land-use effects on forest plant species composition and diversity. Ecol. Res. 2007, 22, 361–371. [Google Scholar] [CrossRef]
- Cruz-Alonso, V.; Ruiz-Benito, P.; Villar-Salvador, P.; Rey-Benayas, J.M. Long-term recovery of multifunctionality in Mediterranean forests depends on restoration strategy and forest type. J. Appl. Ecol. 2019, 56, 745–757. [Google Scholar] [CrossRef]
- Minotta, G.; Degioanni, D. Naturally regenerated English oak (Quercus robur L.) stands on abandoned agricultural lands in Rilate valley (Piedmont Region, NW Italy). IForest Biogeosci. For. 2011, 4, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Gea-Izquierdo, G.; Cañellas, I. Local climate forces instability in long-term productivity of a Mediterranean oak along climatic gradients. Ecosystems 2014, 17, 228–241. [Google Scholar] [CrossRef]
- Nathan, R. Long-distance dispersal of plants. Science 2006, 313, 786–788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hewitt, N.; Kellman, M. Tree seed dispersal among forest fragments: II. Dispersal abilities and biogeographical controls. J. Biogeogr. 2002, 29, 351–363. [Google Scholar] [CrossRef]
- Kurek, P.a.; Dobrowolska, D.; Wiatrowska, B. Dispersal distance and burial mode of acorns in eurasian jays garrulus glandarius in european temperate forests. Acta Ornithol. 2019, 53, 155–162. [Google Scholar] [CrossRef]
- Gea-Izquierdo, G.; Cañellas, I.; Montero, G. Acorn production in Spanish holm oak woodlands. Investig. Agrar. Sist. y Recur. For. 2006, 15, 339–354. [Google Scholar] [CrossRef] [Green Version]
- Leal, J. Análisis del Crecimiento Vegetativo e Inversión Reproductiva de Quercus pyrenaica Willd. en tres localidades de Sierra Nevada. Master’s Thesis, Universidad de Granada, Granada, Spain, 2013. [Google Scholar]
- Gómez, J.; Gómez-Aparicio, L.; Zamora, R.; Montes, J. Problemas de Regeneración de Especies Forestales Autóctonas en el Espacio Natural Protegido de Sierra Nevada. In Proceedings of the III Congreso Forestal Español, Granada, Spain, 25–28 September 2001. [Google Scholar]
- Navarro-González, I.; Pérez-Luque, A.J.; Bonet, F.J.; Zamora, R. The weight of the past: Land-use legacies and recolonization of pine plantations by oak trees. Ecol. Appl. 2013, 23, 1267–1276. [Google Scholar] [CrossRef]
- Milder, A.I.; Fernández-Santos, B.; Martínez-Ruiz, C. Colonization patterns of woody species on lands mined for coal in Spain: Preliminary insights for forest expansion. Land Degrad. Dev. 2013, 24, 39–46. [Google Scholar] [CrossRef]
- Leverkus, A.B.; Rey Benayas, J.M.; Castro, J. Shifting demographic conflicts across recruitment cohorts in a dynamic post-disturbance landscape. Ecology 2016, 97, 2628–2639. [Google Scholar] [CrossRef] [PubMed]
- Perring, M.P.; De Frenne, P.; Baeten, L.; Maes, S.L.; Depauw, L.; Blondeel, H.; Carón, M.M.; Verheyen, K. Global environmental change effects on ecosystems: The importance of land-use legacies. Glob. Chang. Biol. 2016, 22, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Titos, M. (Ed.) Las minas de la Estrella. In La Aventura de Sierra-Nevada 1717–1915; Editorial Universidad de Granada: Granada, Spain, 1990; pp. 226–236. [Google Scholar]
- Mesa-Torres, M. Cáñar: Balcón de la Alpujarra; Fundación Caja General de Ahorros de Granada: Granada, Spain, 2009. [Google Scholar]
- Moreno-Llorca, R.; Zamora, R. Caracterización de la Carga Ganadera en Sierra Nevada en la Segunda Mitad del Siglo XX. Proyecto MIGRAME; Project Report; IISTA-CEAMA; University of Granada: Granada, Spain, 2012. [Google Scholar]
- Robles-Cruz, A.B. “En el conjunto de las Sierras Béticas”: Pastos, producción, diversidad y cambio global. In Pastos, Clave en la Gestión de los Territorios: Integrando Disciplinas; Fernández Rebollo, P., Gómez Cabrera, A., Guerrero, J.E., Garrido Varo, A., Calzado Martínez, C., García Moreno, A.M., Carbonero Muñoz, M.D., Blázquez Carrasco, A., Escuín Royo, S., Castillo Carrión, S., Eds.; Junta de Andalucía, Consejería de Agricultura y Pesca: Sevilla, Spain, 2008; pp. 30–51. [Google Scholar]
- Castro, J.; Zamora, R.; Hódar, J.A. Restoring Quercus pyrenaica forests using pioneer shrubs as nurse plants. Appl. Veg. Sci. 2006, 9, 137–142. [Google Scholar] [CrossRef]
- Gómez-Aparicio, L.; Zamora, R.; Gómez, J.M.; Hódar, J.A.; Castro, J.; Baraza, E. Applying plant facilitation to forest restoration: A meta-analysis of the use of shrubs as nurse plants. Ecol. Appl. 2004, 14, 1128–1138. [Google Scholar] [CrossRef] [Green Version]
- Costa, A.; Villa, S.; Alonso, P.; García-Rodríguez, J.; Martín, F.; Martínez-Ruiz, C.; Fernández-Santos, B. Can native shrubs facilitate the early establishment of contrasted co-occurring oaks in Mediterranean grazed areas? J. Veg. Sci. 2017, 28, 1047–1056. [Google Scholar] [CrossRef]
- Baraza, E.; Gómez, J.M.; Hódar, J.A.; Zamora, R. Herbivory has a greater impact in shade than in sun: Response of Quercus pyrenaica seedlings to multifactorial environmental variation. Can. J. Bot. 2004, 82, 357–364. [Google Scholar] [CrossRef]
- Muñoz, J. Bosques autóctonos y matorrales de media y alta montaña. In Observatorio de Cambio Global de Sierra Nevada: Metodologías de Seguimiento; Aspizua, R., Barea-Azcón, J., Bonet, F., Pérez-Luque, A., Zamora, R., Eds.; Consejería de Medio Ambiente, Junta de Andalucía: Sevilla, Spain, 2014; pp. 74–75. [Google Scholar]
- Chazdon, R.L.; Lindenmayer, D.; Guariguata, M.R.; Crouzeilles, R.; Rey Benayas, J.M.; Lazos Chavero, E. Fostering natural forest regeneration on former agricultural land through economic and policy interventions. Environ. Res. Lett. 2020, 15, 043002. [Google Scholar] [CrossRef]
- Stritih, A.; Senf, C.; Seidl, R.; Grêt-Regamey, A.; Bebi, P. The impact of land-use legacies and recent management on natural disturbance susceptibility in mountain forests. For. Ecol. Manag. 2021, 484, 118950. [Google Scholar] [CrossRef]
- Pausas, J.; Ribeiro, E.; Dias, S.; Pons, J.; Beseler, C. Regeneration of a marginal Quercus suber forest in the eastern Iberian Peninsula. J. Veg. Sci. 2006, 17, 729–738. [Google Scholar] [CrossRef]
Figure 1.
Distribution of Quercus pyrenaica forests in the Iberian Peninsula (a), and in Sierra Nevada mountain range (b). Location of the old croplands (white lines) in the study sites: Robledal de San Juan (SJ) (c), and Robledal de Cáñar (CA) (d). Names of the old croplands as in Table 1. Sampling design (e). Vegetation plots were randomly placed in the abandoned croplands (triangles), and inside the forest (circles) surrounding the old field. Several plots were also located at forest-old field edge (squares).
Figure 1.
Distribution of Quercus pyrenaica forests in the Iberian Peninsula (a), and in Sierra Nevada mountain range (b). Location of the old croplands (white lines) in the study sites: Robledal de San Juan (SJ) (c), and Robledal de Cáñar (CA) (d). Names of the old croplands as in Table 1. Sampling design (e). Vegetation plots were randomly placed in the abandoned croplands (triangles), and inside the forest (circles) surrounding the old field. Several plots were also located at forest-old field edge (squares).

Figure 2.
Interaction plot for the oak juvenile abundance. Habitat type differences within each site were indicated with different letters. Differences between sites for each habitat type were indicated with asterisk. CA: Robledal de Cáñar site (southern slopes); SJ: Robledal de San Juan site (northern slopes).
Figure 2.
Interaction plot for the oak juvenile abundance. Habitat type differences within each site were indicated with different letters. Differences between sites for each habitat type were indicated with asterisk. CA: Robledal de Cáñar site (southern slopes); SJ: Robledal de San Juan site (northern slopes).

Figure 3.
Juvenile abundance classified by tree-size (see Materials and Methods) by habitat type in the two study sites. Mean and standard error are shown.
Figure 3.
Juvenile abundance classified by tree-size (see Materials and Methods) by habitat type in the two study sites. Mean and standard error are shown.

Figure 4.
Relation of the juvenile oak abundance with the estimated age of crop abandonment.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Main characteristics of the abandoned croplands studied.
| Site | Cropland | Elevation (m) | Area (ha) | Number of Plots | |||
|---|---|---|---|---|---|---|---|
| Code | Abandonment Age (Years) | Cropland | Edge | Forest | |||
| Robledal de Cáñar | CA1 | >60 | 1796–1866 | 3.29 | 6 | 3 | 4 |
| CA2 | <30 | 1789–1858 | 5.80 | 9 | 3 | 7 | |
| CA3 | 40–60 | 1851–1892 | 1.56 | 3 | 3 | 4 | |
| Robledal de San Juan | SJ1 | 40–60 | 1507–1674 | 3.47 | 6 | 3 | 6 |
| SJ2 | 30–40 | 1575–1746 | 10.36 | 13 | 3 | 10 | |
Table 2.
Forest attributes of northern (SJ) and southern (CA) sites. U Mann–Whitney statistics with significance at 0.05 level. Mean and SE are shown.
Table 2.
Forest attributes of northern (SJ) and southern (CA) sites. U Mann–Whitney statistics with significance at 0.05 level. Mean and SE are shown.
| Variable | Southern Site (CA) | Northern Site (SJ) | U Statistic | p Value |
|---|---|---|---|---|
| % of Q. pyrenaica | 96.11 ± 1.28 | 100 ± 0 | 3.688 | 0.0001 |
| Tree density (ind/ha) | 1671.11 ± 229.21 | 1587.5 ± 161.67 | 4.808 | 0.9369 |
| Juvenile abundance (ind/ha) | 1004.44 ± 195.72 | 883.33 ± 127.18 | 4.852 | 0.7667 |
| Adult abundance (ind/ha) | 584.44 ± 80.47 | 704.17 ± 63.31 | 4.448 | 0.1780 |
| Maximum tree height (m) | 13.93 ± 0.65 | 13.75 ± 0.71 | 4.824 | 0.8736 |
| Tree height mean (m) | 4.32 ± 0.6 | 5.09 ± 0.37 | 4.330 | 0.0855 |
| Tree height median (m) | 3.19 ± 0.83 | 3.57 ± 0.66 | 4.564 | 0.3527 |
| Tree height 75 percentile (m) | 5.73 ± 1.02 | 8.29 ± 0.6 | 4.343 | 0.0922 |
| Tree height 90 percentile (m) | 10.07 ± 0.95 | 11.22 ± 0.54 | 4.605 | 0.4399 |
| Basal Area (m/ha) | 37.56 ± 4.23 | 33.58 ± 3.6 | 4.912 | 0.5400 |
Table 3.
Model selection for the oak juvenile abundance, sorted by minimum AICc value.
| Model | df | logLik | AICc | AICc | Nagelkerke R2 |
|---|---|---|---|---|---|
| Habitat type + Site + Habitat type × Site | 6 | −221.89 | 457.78 | 0 | 0.970 |
| Habitat type + Site | 4 | −231.83 | 473.65 | 15.87 | 0.955 |
| Habitat type | 3 | −236.21 | 480.42 | 22.64 | 0.946 |
| Site | 2 | −291.06 | 588.12 | 130.34 | 0.116 |
| null model | 1 | −293.09 | 590.19 | 132.41 | 0 |
Table 4.
ANOVA table of the selected GLM model for the abundance of Quercus pyrenaica juvenile across study sites and habitat types. F-value and p-values are displayed.
Table 4.
ANOVA table of the selected GLM model for the abundance of Quercus pyrenaica juvenile across study sites and habitat types. F-value and p-values are displayed.
| Variable | SS | df | F | p-Value |
|---|---|---|---|---|
| Habitat type | 233.89 | 2 | 72.95 | 0.0001 |
| Site | 13.09 | 1 | 8.16 | 0.0054 |
| Habitat type × Site | 27.19 | 2 | 8.48 | 0.0004 |
| Residuals | 123.43 | 77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pérez-Luque, A.J.; Bonet-García, F.J.; Zamora, R. Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region. Forests 2021, 12, 1584. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111584
AMA Style
Pérez-Luque AJ, Bonet-García FJ, Zamora R. Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region. Forests. 2021; 12(11):1584. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111584
Chicago/Turabian StylePérez-Luque, Antonio J., Francisco J. Bonet-García, and Regino Zamora. 2021. "Colonization Pattern of Abandoned Croplands by Quercus pyrenaica in a Mediterranean Mountain Region" Forests 12, no. 11: 1584. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111584
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.