
Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer


1
Department of Forest Resources, Kangwon National University, Chuncheon 24341, Korea
2
Department of Environmental Science and Ecological Engineering, Korea University, Seoul 02841, Korea
3
Forest Technology and Management Research Center, National Institute of Forest Science, Pochoen 11186, Korea
*
Author to whom correspondence should be addressed.
Forests 2021, 12(11), 1595; https://0-doi-org.brum.beds.ac.uk/10.3390/f12111595
Submission received: 7 October 2021
/
Revised: 1 November 2021
/
Accepted: 17 November 2021
/
Published: 19 November 2021
(This article belongs to the Special Issue Impact of Extreme Weather Events on Forest Functions)
Abstract
:Extreme climate events such as heat waves, drought, and heavy rainfall are occurring more frequently and are more intense due to ongoing climate change. This study evaluated the early growth performance of one-year-old Larix kaempferi (Lamb.) Carr. seedlings under open-field extreme climate conditions including experimental warming and different precipitation regimes. We recorded the survival rate, root collar diameter, height, biomass, shoot-to-root ratio, and seedling quality index using nine treatments (three temperature levels, i.e., control, warming by 3 °C and by 6 °C, × three precipitation levels, i.e., control, drought, and heavy rainfall) in July and August 2020. The survival rate of seedlings did not differ between treatments, showing high values exceeding 94% across treatments. The measured shoot height was largest under warming by 3 °C and high rainfall, indicating that moderate warming increased seedling height growth in a moist environment. Heavy rainfall decreased stem volume by 21% and 25% under control and warming by 6 °C treatments, respectively. However, drought manipulation using rain-out shelters did not decrease the growth performance. Overall, extreme climate events did not affect the survival rate, biomass, shoot-to-root ratio, and seedling quality index of L. kaempferi. We thus conclude that, regarding growth responses, L. kaempferi seedlings may be resistant to short-term extreme warming and drought events during summer.
1. Introduction
Climate change results in an increased frequency and higher intensity of extreme weather events such as heat waves, drought, and heavy rainfall [1,2,3]. Persistent global warming has also caused extremely high temperatures, excessive precipitation, and severe drought events [3,4,5]. Climate models report that the global mean temperature rise could exceed 4 °C in the 21st century and the extreme climates associated with different global warming scenarios have been extensively investigated [3]. Extreme climatic events have been defined in different ways, based on statistical quantification of climatic variables or synthetic extremes of both driving and response variables [6,7]. The Intergovernmental Panel on Climate Change defined an extreme climatic event as an event being rarer than the 10th or 90th percentile of climate events within its statistical frequency distribution at a particular place over a certain period of time [1,7]. Such unprecedented climate events and uncertain future climate projections may affect tree growth, mortality, and forest structure and functioning [6,7,8,9,10]. Given the effects of extreme climate events on plants, these processes may reduce forest productivity, so it is crucial to predict the responses of plants to extreme climate events.
Many plants are resilient to heat stress and moisture deficit to some extent; however, extreme climate events irreversibly damage various physiological traits and mechanisms of plants [11,12]. Extreme heat deteriorates the photosynthetic processes, chlorophyll functioning, and biomass production of woody plants [11,13], and extreme drought markedly affects physiological traits and reduces tree biomass, which, however, depends on external conditions throughout the year [14,15,16,17]. Moreover, excessive rainfall may also decrease plant productivity under extreme rainfall due to reduced photosynthetic and stomatal conductance under waterlogged conditions [18]. However, it is unclear whether tree mortality occurs only due to an excessively wet environment [19]. The vulnerability of plants may increase when such extreme events occur simultaneously. For instance, extreme heat in combination with drought increases tree mortality [20]. Therefore, multifactor experiments are crucial for determining the interaction effects of simultaneous extreme climate events [15].
The early growth performance of small or young seedlings planted in open fields can be particularly affected by extreme temperatures and precipitation [21,22]. Various adverse effects of unpredictable climate factors on seedling quality, which can be assessed by measuring numerous morphological and physiological traits, have been observed in many nursery and silvicultural operations [23]. Furthermore, seedling growth depends on seedling quality [24], and whether seedlings can survive and thrive under extreme climatic conditions in nurseries is a key factor in determining the supply of seedlings and, thus, the success of plantations and restorations [17,25,26]. Despite substantial information on the production of high-quality seedlings, the growth responses of seedlings to extreme climate events in nurseries and at planting sites under changing climates are not yet comprehensively understood [17,27,28,29].
In this study, we used the Japanese larch, Larix kaempferi (Lamb.) Carr., which is one of the most popular plantation tree species due to its high economic value, wood quality, and restoration potential in Asia [17,30]. The plantation areas of L. kaempferi gradually increased approximately 10-fold over the past decade across South Korea, and the planted area in 2019 accounted for approximately 20% (4559 ha) of the total plantation area [31]. Larix kaempferi is an important fast-growing deciduous coniferous species encircling the Northern Hemisphere, due to its disease and cold resistance compared to other species [17,30,31,32,33,34]. In general, drought stress is considered a main factor causing a decline in larch tree species [10,32,33], whereas saturated soil water conditions may also adversely affect photosynthetic activity and stomatal conductance [18]. A few previous studies suggested a high risk of growth cessation in similar larch species such as Larix principis-rupprechtii Mayr planted across large areas in north-central China during extreme drought [10], and quantitative changes in the distribution and productivity of L. kaempferi under future climate change were predicted [34]. However, the growth responses of L. kaempferi seedlings to multiple factors of extreme climate have not been examined, even though this would be required to understand how climate change will affect successful planting in the future.
Our previous studies showed that physiological traits such as photosynthetic activity, stomatal conductance, and the transpiration rate of two-year-old L. kaempferi were reduced due to warming by 3 °C in the second year of the warming treatment [35], and warming by 3 °C combined with reduced precipitation increased the mortality of one-year-old L. kaempferi seedlings due to drought-induced heat stress [26]. However, the impact of multiple factors of extreme climate events on the early growth performance, biomass allocation, and seedling quality of L. kaempferi has not yet been documented. Therefore, the objective of this study was to determine the survival rate and growth performance of larch seedlings under experimental warming and precipitation conditions. We hypothesized that moderate warming would increase early growth performance regarding height, root collar diameter (RCD), biomass accumulation, and survival rate, whereas extreme drought, excessive precipitation, and extremely high temperatures should decrease the growth performances and hence seedling quality index and survival rate.
2. Materials and Methods
2.1. Experimental Design
This study was conducted at an experimental tree nursery located in the Forest Technology and Management Research Center, Pocheon, Korea (37°45′39″ N, 127°10′13″ E, 106 m a.s.l.). The annual mean air temperature and precipitation in this area are typically 10.2 °C and 1365 mm, respectively (1997–2019) [36]. In April 2020, 27 experimental plots of 1.5 × 1.0 m were established (Figure 1a). In total, 88 one-year-old L. kaempferi seedlings were planted with 11-cm intervals between seedlings in each plot containing a homogeneous sandy loam soil (70% sand, 20% silt, and 10% clay), following the guidelines for nursery practices (Figure 1b and Figure S1b) [37]. The seeds obtained from a seed orchard located in Anmyeondo (36°29′ N, 126°23′ E, 40–50 m a.s.l.) of the National Forest Seed Variety Center were grown in the container tree nursery at the Forest Technology and Management Research Center. The annual mean air temperature and precipitation in the seed orchard are 13 °C and 1380 mm, respectively [36]. There was no difference in the initial RCD and height of the planted seedlings between pretreatment plots (p > 0.05, Table S1). The plots were arrayed with buffer zones of 0.5 m to the external boundary and 1.5 m between plots to prevent treatments from affecting each other (Figure 1).
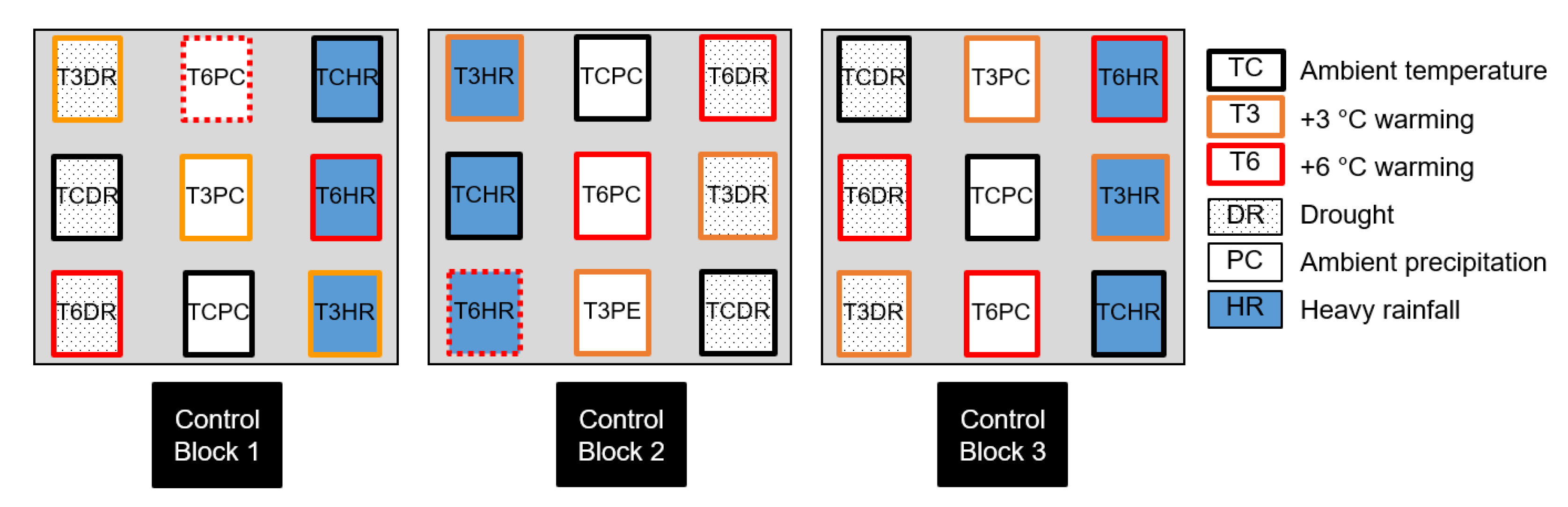
In July and August 2020, the experimental plots were subjected to nine treatments (three temperature levels [TC: ambient, T3: warming by 3 °C, T6: warming by 6 °C] × three precipitation levels [DR: drought, PC: ambient, HR: heavy rainfall]), using three replicates per treatment (Figure 2). The target temperatures for T3 and T6 were manipulated based on the 90th and 99th percentiles of the daily maximum temperature at the study site during the reference period (1961–2019) [38]. For the DR treatment, we considered the longest consecutive days with rainfall <1 mm during the reference period [39], and the HR treatment was produced using the 95th percentile of daily precipitation [40,41]. Consequently, the DR treatment lasted for nine days during the reference period, and the threshold of HR treatment was 113 mm per day [42].
Based on the analysis of extreme climate scenarios, we elevated the soil surface temperature by 3 °C and 6 °C, respectively, for seven days, compared to the ambient temperature to produce T3 and T6 treatments. We used infrared heaters (FT-1000, Mor Electronic Heating Assoc., Comstock Park, MI, USA) mounted approximately 30 cm above the canopy of the seedlings and thermometers (SI-111, Apogee Instruments, Logan, UT, USA) connected to data loggers (CR1000X, Campbell Scientific, Inc., Logan, UT, USA) and relays (SDM-CD-16AC, Campbell Scientific, Inc.). We then excluded ambient rainfall for nine days to simulate drought, and HR plots were irrigated with 113 mm water per day every three days during the DR treatment period. To manipulate the DR plots, we used automatic rainout shelters coupled with rain detectors (Figure 1c). To simulate heavy rain, we simultaneously applied 170 L of stored water for 1.4 h per day at a pressure of 1.0 bar to each plot using rainfall simulators consisting of two spraying nozzles (Unijet D5-35, Spraying Systems Co., Wheaton, IL, USA) per plot. These manipulations were repeated twice during the experimental period (day of year 195–233).
We measured the soil temperature (°C) and volumetric soil water content (vol %) at 5 cm soil depth at the center of each plot every 30 min using soil sensor probes (CS655, Campbell Scientific, Inc., Logan, UT, USA) connected to the data loggers. An automatic weather station recorded the precipitation every hour at the study site.
2.2. Seedling Measurements
The survival of all seedlings was monitored during the experimental period. Measurements of RCD (in mm, at ground level) and shoot length (height, H, in mm) were made using digital calipers and folding rulers, respectively. For growth measurements, 30 of 88 seedlings per plot were selected from near the center (n = 90 per treatment) as the 30 seedlings were relatively well warmed by the infrared heating lamps compared to the outer seedlings (Figure S1). The measurements were made before (mid-May) and after (early October) the treatments. In October, three of the surviving seedlings per plot, grown from near the center of the plot, were harvested to measure the biomass accumulation and allocation (n = 9 per treatment). The harvested seedlings were divided into shoots (leaf and stem) and roots, and the samples were weighed after oven-drying at 65 °C for 96 h.
Height and RCD measurements were used to calculate the height-to-RCD ratio (H/D) and stem volume (V, cm3) of seedlings, which were approximated using the equation for an elliptical cone [43]:
V = π × RCD2 × H/6.
2.3. Data Analyses
A two-way analysis of variance was used to examine the effects of temperature and precipitation and their interactions on soil temperature and moisture, and on the survival rate, RCD, height, volume, biomass, H/D and R/S ratios, and SQI of seedlings. We excluded one replicate from the T6PC and T6HR treatments as two plots among the 27 randomized complete block-designed plots were partly malfunctional. Consequently, we analyzed the data from each treatment as replicates (n = 60 or 90 per treatment for RCD and height; n = 9 per treatment for biomass). When significant results were observed, Tukey’s HSD test was used to determine significant differences in mean values between treatments. To determine the effect of precipitation treatment on tree height growth with the exclusion of the influence of RCD, a linear mixed-effects model was fitted using the ‘lme’ function of the ‘nlme’ R package [46]. The precipitation treatment was considered a fixed factor, the seedlings within blocks were a random factor, and the RCD was a covariate. Pearson correlations between height and RCD were analyzed using the ‘stat_corr’ function of the ‘ggpubr’ R package. Statistical significance is reported at α = 0.05. All statistical analyses were performed with the available variables measured in May and October using R version 4.1.1 [47].
3. Results
3.1. Effects of Temperature and Precipitation Treatments on Seedling Development
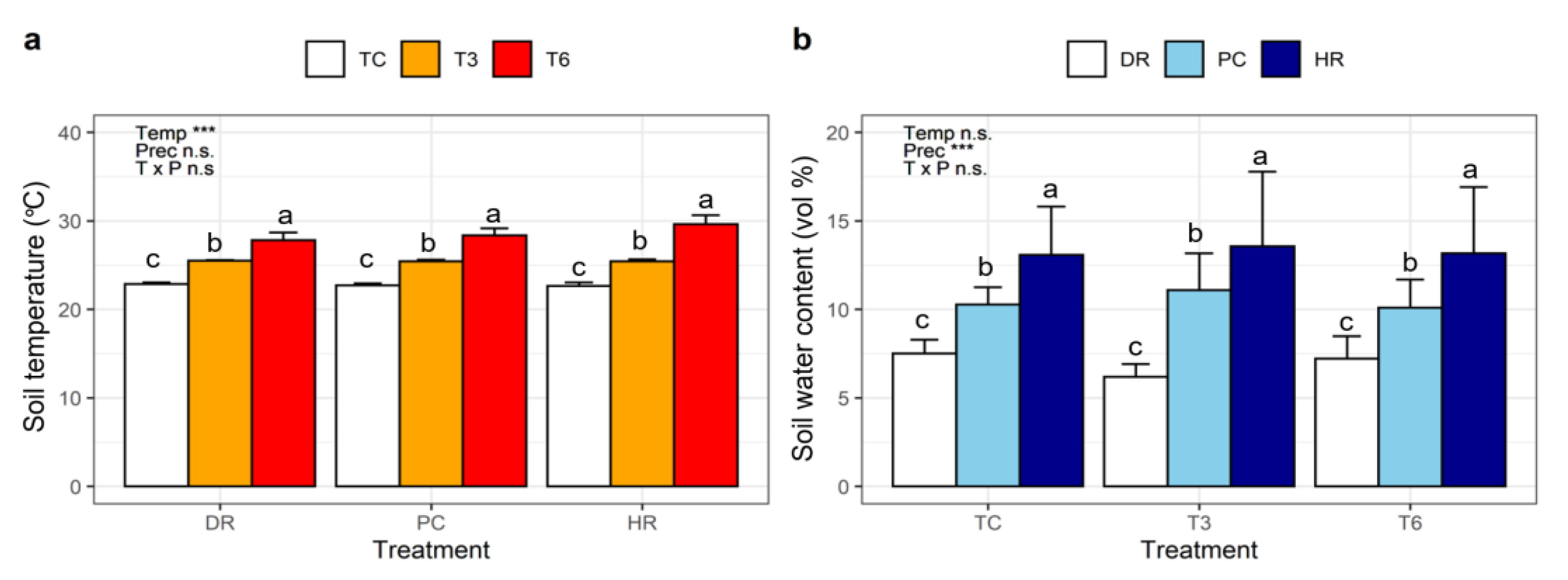
Temperature (T) treatment significantly affected the soil temperature (p < 0.001), but did not affect the soil water content (Table 1). Precipitation (P) treatment also significantly affected the soil water content (p < 0.001), but did not affect soil temperature (Table 1). There was no interaction effect of temperature and precipitation treatments on soil temperature and water content. Mean temperatures were 22.7 ± 0.3 °C, 25.5 ± 0.2 °C, and 28.6 ± 1.1 °C in TC, T3, and T6, respectively, and mean soil moisture content was 7.0% ± 1.0%, 10.5% ± 1.5%, and 13.3% ± 3.1% in DR, PC, and HR, respectively (Figure 3).
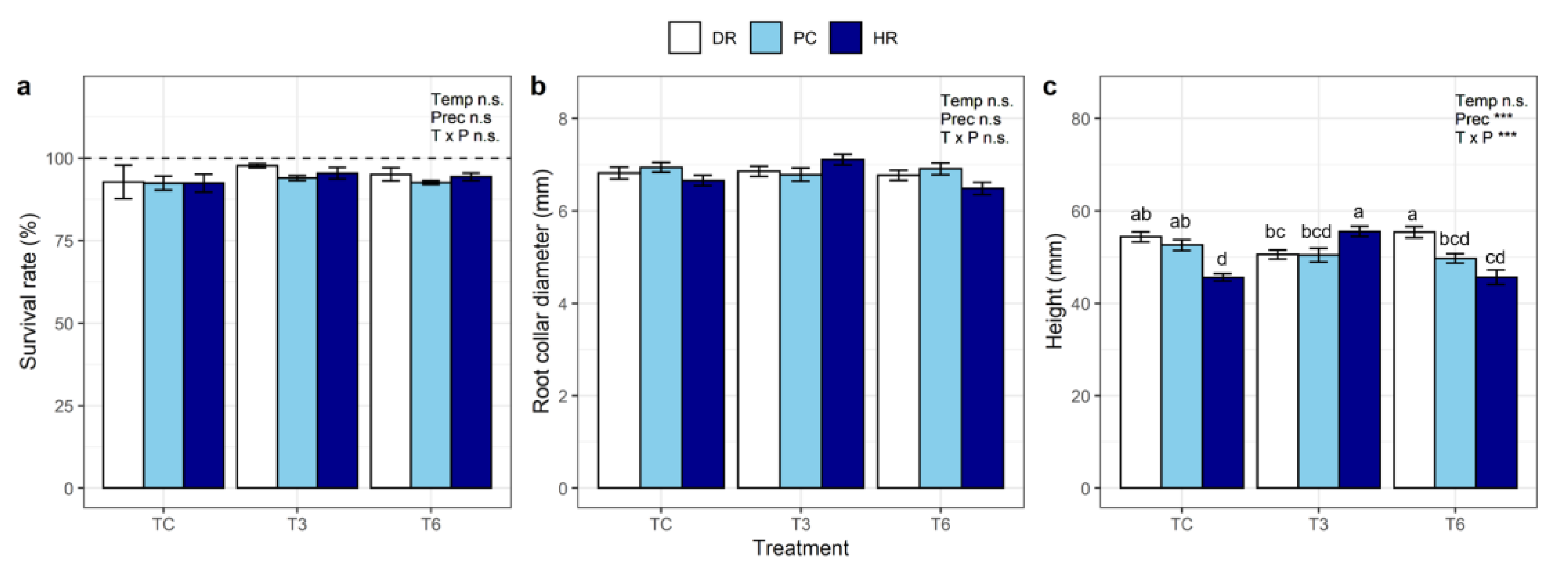
The survival rate of seedlings did not differ between treatments (Table 1) and was generally high, with values exceeding 94% across treatments (Figure 4a, Table S1). There was no significant difference in the RCD and height measured in mid-May between pretreatments (Table S1). There was no significant effect of the treatments on RCD, which showed a narrow range of 6.4 mm (T6HR) to 7.1 mm (T3HR) at the end of the experiment (Figure 4b).
The warming treatments did not affect seedling height, but an effect of precipitation and interaction effects on height were observed (p < 0.001, Table 1). The height of the seedlings was largest in T3HR (57.5 cm) and T6DR (56.0 cm) and smallest in TCHR (45.9 cm) and T6HR (48.6 cm; p < 0.05, Figure 4c).
Temperature and precipitation treatments did not affect the estimated stem volume of seedlings, but significant interaction effects on the volume were observed (p < 0.001, Table 1). The stem volume of seedlings was highest in T3HR (15.4 cm3 seedling–1) and lowest in TCHR and T6HR (11.0 and 10.4 cm3 seedling–1, respectively; p < 0.05). The seedling biomass (root, stem, leaf, and total biomass) did not differ between treatments. The total biomass of seedlings ranged from 13.24 to 15.88 g seedling–1 (Table 2). The small sample size may result in no significant difference in biomass accumulation between the treatments; however, stem biomass was directly correlated with stem volume across the treatments (p < 0.001, Figure S2).
3.2. S/R Ratios and SQI
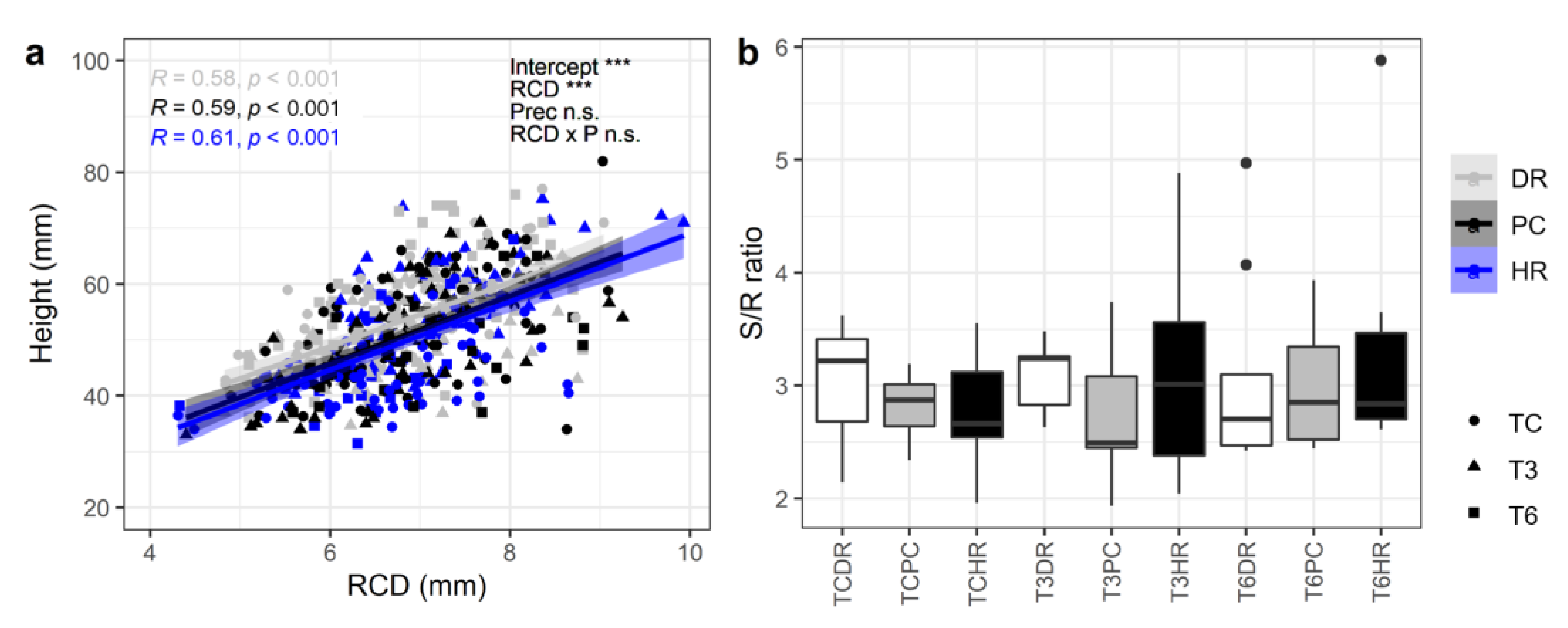
Height was strongly correlated with RCD, but the treatments did not affect the relationship between RCD and height (Figure 5a). The shoot-to-root ratio was not significantly affected by the treatments, with a range from 2.75 to 3.42 (Figure 5b). The SQI was also not affected by the treatments (Table 2).
4. Discussion
4.1. Survival
We hypothesized that extreme warming, drought, and heavy rainfall would decrease survival rates. In general, warming-induced heat stress can decrease tree growth and thus frequently decreases the survival rate [9]. In temperate regions, extensive drought can induce large-scale tree population declines [48,49,50]; however, some field studies reported that severe drought treatments did not significantly alter primary production [7]. Our study showed that the experimental climate events did not affect seedling survival at the end of the experimental period. In our open-field experiment, we used one-year-old container seedlings, which can be advantageous for drought avoidance potential, resulting in higher levels of field survival when compared to bare-root seedlings in terms of planting stress and early field performance [51,52]. As we applied the treatments in July, two months after the planting of the seedlings, the seedlings had sufficient time to stabilize and adapt to the field environments, compared to other experiments conducted during different periods including the early growing season [16,53]. Therefore, such manipulation experiments should be executed over multiple years, rather than within a single year, particularly with respect to variation in air humidity [15]. Furthermore, long-term field experiments are required to determine the impact of drought on tree mortality because plants may show a time lag regarding the effects of drought on productivity and mortality [7,54].
4.2. Plant Growth
We expected that moderate warming would increase seedling growth in terms of RCD, height, and biomass, whereas extreme warming should reduce growth. In general, increasing temperatures stimulate plant biomass accumulation, but temperature-induced heat stress and water deficit reduce plant growth, thereby offsetting the stimulation effect [7,55,56,57,58]. The temperature treatments (T3 and T6) of the current study did not significantly affect the overall growth performance; however, seedling height in the HR treatments was significantly larger in T3HR than in TCHR and T6HR (Figure 4c), which showed consistent patterns regarding stem volume following the order T3HR (40.3% increase) > TCHR > T6HR (5.2% decrease; Table S1). These results suggest that extreme temperatures within a certain range of water availability decrease plant growth [9].
In our study, seedling height was highest under experimental warming by 3 °C and heavy rainfall treatment (Figure 4c), showing that optimum levels of warming without drought stress can stimulate plant growth. However, the seedling height in T3HR was not significantly higher than that in TCPC, which is the ambient condition. Numerous studies on experimental warming have reported that increasing the temperature stimulates plant growth [59,60], whereas extreme climate conditions typically reduce plant growth and primary production [6,61]. A meta-analysis showed that warming increased plant carbon pools by 6.8% and 7.0% regarding above- and belowground parts, respectively, with significantly higher responses in forests, especially in tree seedlings and saplings, than in other ecosystem types [60]. In general, warming by 3 °C and 6 °C increased emergence, development, and shoot length in 15 North American tree species, and the seedlings showed increased growth in response to warming, whereas exceedingly dry or wet conditions limited this positive response [9].
We also hypothesized that drought and heavy rainfall would decrease seedling growth performance. Precipitation manipulation did not affect the RCD of the seedlings; however, we found a significant precipitation effect on seedling height and a significant interaction effect of precipitation and temperature on seedling height and stem volume, which is a function of both shoot height and RCD. For instance, heavy rainfall decreased seedling height and stem volume under TC and T6 treatments, but increased height under the T3 treatment. Heavy rainfall results in water logging, which impedes gas exchange and limits oxygen availability, thus restricting plant photosynthesis and production [7,62]. Our previous study on L. kaempferi seedlings observed that a higher precipitation-induced increase by 32.5% in soil water content decreased the total chlorophyll content and net photosynthetic rate by 8.24% and 4.55%, respectively [63]. Another previous study reported that short-term waterlogging strongly induced stomatal closure and reduced net photosynthesis in L. kaempferi seedlings [18]. The irrigation system in our well-drained soil did not result in water logging; however, soil moisture in HR treatments reached 31 vol %. Manipulated amounts of rainfall produced significant differences in mean soil moisture content at the surface soil layer (5 cm soil depth) between treatments; however, we did not monitor soil moisture at deeper soil layers. Thus, such measurements in deeper layers including the rooting zone are required to validate the effects on soil moisture conditions, and additional experiments under drier conditions are needed to observe more conclusive effects regarding available water content and drought stress.
In the current study, drought did not affect seedling height in the TC and T3 treatments. The seedling height under the T6 treatment was greater in the DR, rather than reduced, in the PC and HR, but did not differ from that in the TCPC treatment (Figure 4c). A previous study on the effect of drought on L. kaempferi seedlings highlighted that lower precipitation increased the total chlorophyll content and net photosynthetic rate by 6.40% and 4.32%, respectively, indicating that this species can have positive physiological responses to moderate drought [63]. In addition, higher accumulated precipitation during the experimental period (July to August 2020) may reduce water stress and heat stress in the seedlings. The number of precipitation days (28 days) and the accumulated precipitation (832 mm) during the experimental period was 30% and 20% higher, respectively, than the average values over the past 30 years [36]. Although tolerance to drought depends on species, soil type, and drought intensity and duration [27,64,65], our results suggest that short-term drought events in summer may not constitute extreme climatic events for L. kaempferi seedlings, whereas excessive rainfall during short periods may affect plant growth.
4.3. Biomass Allocation and Seedling Quality
In general, moderate warming enhances plant growth due to the increased allocation of carbon to aboveground rather than belowground parts, thereby increasing the shoot-to-root ratio or total biomass-to-root biomass ratio [66,67,68]. By contrast, drought-induced stress increases the root-to-shoot ratio by increasing the proportion of soluble sugars and starch in roots [16,69,70]. Thus, these climatic factors may exert interaction effects on carbon allocation and metabolic activity [67,71]. In our study, similar to the lack of remarkable responses of height growth to soil drought, no significant effect of drought on S/R ratios was observed (Figure 5b). The responses of carbon allocation to heat and drought stress can also vary according to species-specific water use strategy, season, and development stage [9,56,66,71,72,73]. Taeger et al. [56] showed that experimental drought increased taproot length and root-to-shoot ratios of Pinus sylvestris L. seedlings. Arend et al. [67] showed that the ratio of root length to shoot height increased in response to drought in Quercus species, but decreased in response to air warming, indicating provenance-specific sensitivity to drought and air warming. A previous study reported that an 80% reduction in precipitation led to a reduction in carbon isotopic composition for L. kaempferi seedlings, but did not affect stomatal conductance, predawn leaf water potential, and transpiration [17]. However, there has still been no comparable research in this species on the response of biomass allocation to warming and drought. In our current study, short-term drought stress in summer may not increase the root to shoot ratio for L. kaempferi seedlings as drought could alter the root structures by allocating more carbon to 1st- and 2nd-order fine roots than coarse roots [74]. Thus, further studies on drought responses to root morphological and physiological traits such as mean root diameter, specific root length, and respiration will be prerequisites to understanding biomass allocation for various species in a changing climate.
Regarding seedling quality at the nursery stage, we expected that extreme warming and heavy rainfall would reduce seedling quality. Practical guidelines for seedling management propose grading criteria for characterizing seedling quality, and nurseries commonly rely on morphological traits such as height and RCD as indicators of seedling quality [23,43]. According to the Korean practical nursery guidelines, the standard measurements of good-quality 1-1 L. kaempferi seedlings are 35–60 cm height, >6 mm RCD, and a H/D ratio <90 at a height exceeding 60 cm [37]. In our study, the measurements of a few seedlings per treatment were outside these ranges, but the mean values of all treatments were within the ranges of good-quality seedlings. Significant differences in the H/D ratio were mainly due to significant variations in height between treatments (Table S1). These results indicate that our short-term climate treatments altered the H/D ratio of 1-1 L. kaempferi seedlings, but they did not critically reduce seedling quality. The SQI, of which higher values are preferable, reflects changes in biomass, H/D, and S/R ratios, and thus appears to be a good index for describing seedling quality [23,45]. The SQI of germinated seedlings of L. kaempferi decreased as the temperature or precipitation increased [26]. In our study, there was no significant difference in SQI between the treatments (Table 2). This suggests that short-term extreme climate events during humid summers do not significantly affect the quality of L. kaempferi seedlings.
5. Conclusions
Overall, seedling biomass, biomass allocation and mortality were not significantly affected by climate manipulation, indicating tolerance to short-term extreme climate events in summer. By contrast, moderate warming with increased precipitation (e.g., T3HR) in well-drained soils may constitute suitable nursery conditions for L. kaempferi seedlings. However, more frequent heat stress and a high soil water content due to heavy rainfall reduce seedling quality, as evidenced by the reduced height, volume, and SQI in T6HR compared to other treatments. Our results highlight that, regarding viability and early growth, L. kaempferi seedlings may tolerate short-term extreme warming events in summer, supporting the idea that L. kaempferi is beneficial to reforestation and restoration in arid woodlands and in the context of climate change. However, to elucidate the potential impact on tree growth performance, further studies are needed to examine the effects of higher frequencies and longer periods of extreme climate events on various species regarding growth and other physiological responses.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f12111595/s1, Table S1: Survival rate, root collar diameter, height, stem volume, H/D, S/R, and T/R ratios of Larix kaempferi seedlings under extreme climate events by temperature and precipitation manipulations. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C extreme warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Values are means ± S.D. (n = 3 for survival rate, n = 90 for RCD, height, stem volume and H/D ratio, n = 9 for S/R and T/R ratios. Values with different letters in a column indicate statistical differences between treatments within a species at 5% levels by Tukey HSD test, Figure S1: The descriptions of (a) warming treatment system and (b) areas warmed by infrared heating lamps and locations of the sampled seedlings (30 samples for root collar diameter and height growth measurements and three samples for biomass measurement. (modified from Kim et al., under review), Figure S2: The relationship between stem volume and stem biomass of Larix kaempferi seedlings across the temperature and precipitation manipulations. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C extreme warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Values are means ± S.D. (n = 9 for stem biomass, n = 60 or 90 for stem volume per treatment). Results from the Pearson correlation analysis are shown in the figure.
Author Contributions
Conceptualization, N.-J.N. and Y.S.; methodology, N.-J.N. and Y.S.; formal analysis, N.-J.N.; investigation, N.-J.N., G.-J.K., and M.-S.C.; resources, N.-J.N.; data curation, N.-J.N.; writing—original draft preparation, N.-J.N.; writing—review and editing, N.-J.N., G.-J.K., Y.S., and M.-S.C.; visualization, N.-J.N.; supervision, M.-S.C.; project administration, Y.S. and M.-S.C.; funding acquisition, Y.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Korea Forest Service, grant number 2020181A00-2122-BB01.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This work was carried out at the Forest Technology and Management Research Center. We thank Won-Geuk Kim for nursery settings for the experiment.
Conflicts of Interest
The authors declare no conflict of interest.
References
- IPCC. Climate change 2013: The physical science basis. In Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Stocker, T.F., Ed.; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Mazdiyasni, O.; AghaKouchak, A. Substantial increase in concurrent droughts and heatwaves in the United States. Proc. Natl. Acad. Sci. USA 2015, 37, 11484–11489. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Jiang, D.; Lang, X. Future extreme climate changes linked to global warming Intensity. Sci. Bull. 2017, 62, 1673–1680. [Google Scholar] [CrossRef] [Green Version]
- Orlowsky, B.; Seneviratne, S.I. Global changes in extreme events: Regional and seasonal Dimension. Clim. Chang. 2012, 110, 669–696. [Google Scholar] [CrossRef] [Green Version]
- Myhre, G.; Alterskjær, K.; Stjern, C.W.; Hodnebrog, Ø.; Marelle, L.; Samset, B.H.; Sillmann, J.; Schaller, N.; Fischer, E.; Schulz, M.; et al. Frequency of extreme precipitation increases extensively with event rareness under global warming. Sci. Rep. 2019, 9, 16063. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Bahn, M.; Ciais, P.; Frank, D.; Mahecha, M.D.; Seneviratne, S.I.; Zscheischler, J.; Beer, C.; Buchmann, N.; Frank, D.C.; et al. Climate extremes and the carbon cycle. Nature 2013, 500, 287–295. [Google Scholar] [CrossRef] [PubMed]
- Niu, S.; Luo, Y.; Li, D.; Cao, S.; Xia, J.; Li, J.; Smith, M.D. Plant growth and mortality under climate extremes: An overview. Environ. Exp. Bot. 2014, 98, 13–19. [Google Scholar] [CrossRef]
- Reyer, C.P.O.; Leuzinger, S.; Rammig, A.; Wolf, A.; Bartholomeus, R.P.; Bonfante, A.; de Lorenzi, F.; Dury, M.; Gloning, P.; Abou Jaoude, R.; et al. A plant’s perspective of extremes: Terrestrial plant responses to changing climatic variability. Glob. Chang. Biol. 2013, 19, 75–89. [Google Scholar] [CrossRef]
- Fisichelli, N.; Wright, A.; Rice, K.; Mau, A.; Buschena, C.; Reich, P.B. First-year seedlings and climate change: Species-specific responses of 15 North American tree species. Oikos 2014, 123, 1331–1340. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Manzanedo, R.D.; D’Orangeville, L.; Lv, P.; Wang, C.; Xu, C.; Hou, M.; Huang, X.; Rademacher, T. High risk of growth cessation of planted larch under extreme drought. Environ. Res. Lett. 2021, 16, 014040. [Google Scholar] [CrossRef]
- Teskey, R.; Wertin, T.; Bauweraerts, I.; Ameye, M.; McGuire, M.A. Responses of tree species to heat waves and extreme heat events. Plant Cell Environ. 2015, 38, 1699–1712. [Google Scholar] [CrossRef]
- Ruehr, N.K.; Grote, R.; Mayr, S.; Arneth, A. Beyond the extreme: Recovery of carbon and water relations in woody plants following heat and drought stress. Tree Physiol. 2019, 39, 1285–1299. [Google Scholar] [CrossRef] [PubMed]
- Bauweraerts, I.; Wertin, T.M.; Ameye, M.; McGuire, M.A.; Teskey, R.O.; Steppe, K. The Effect of heat waves, elevated [CO2] and low soil water availability on northern red oak (Quercus rubra L.) seedlings. Glob. Change. Biol. 2013, 19, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Kuster, T.M.; Arend, M.; Bleuler, P.; Günthardt-Goerg, M.S.; Schulin, R. Water regime and growth of young oak stands subjected to air-warming and drought on two different forest soils in a model ecosystem experiment. Plant Biol. 2012, 15, 138–147. [Google Scholar] [CrossRef] [PubMed]
- Kreyling, J.; Beier, C. Complexity in climate change manipulation experiments. Bioscience 2013, 63, 763–767. [Google Scholar] [CrossRef] [Green Version]
- Matías, L.; Castro, J.; Villar-Salvador, P.; Quero, J.L.; Jump, A.S. Differential impact of hotter drought on seedling performance of five ecologically distinct pine species. Plant Ecol. 2017, 218, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Bhusal, N.; Lee, M.; Han, A.R.; Han, A.; Kim, H.S. Responses to drought stress in Prunus sargentii and Larix kaempferi seedlings using morphological and physiological parameters. For. Ecol. Manag. 2020, 465, 118099. [Google Scholar] [CrossRef]
- Terazawa, K.; Maruyama, Y.; Morikawa, Y. Photosynthetic and stomatal responses of larix kaempferi seedlings to short-term waterlogging. Ecol. Res. 1992, 7, 193–197. [Google Scholar] [CrossRef]
- Tei, S.; Sugimoto, A.; Yonenobu, H.; Kotani, A.; Maximov, T.C. Effects of extreme drought and wet events for tree mortality: Insights from tree-ring width and carbon isotope ratio in a siberian larch forest. Echohydrology 2019, 12, e2143. [Google Scholar] [CrossRef]
- Allen, C.D.; Breshears, D.D.; McDowell, N.G. On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the anthropocene. Ecosphere 2015, 6, 129. [Google Scholar] [CrossRef]
- Saccone, P.; Sylvain, D.; Pagés, J.-P.; Brun, J.-J.; Michalet, R. The role of biotic interactions in altering tree seedling responses to an extreme climate event. J. Veg. Sci. 2009, 20, 403–414. [Google Scholar] [CrossRef]
- Andivia, E.; Villar-Salvador, P.; Oliet, J.A.; Puértolas, J.; Dumroese, R.K.; Ivetić, V.; Molina-Venegas, R.; Arellano, E.C.; Li, G.; Ovalle, J.F. Climate and species stress resistance modulate the higher survival of large seedlings in forest restorations worldwide. Ecol. Appl. 2021, 31, e02394. [Google Scholar] [CrossRef] [PubMed]
- Grossnickle, S.C.; MacDonald, J.E. Seedling quality: History, application, and plant attributes. Forests 2018, 9, 283. [Google Scholar] [CrossRef] [Green Version]
- Riikonen, J.; Luoranen, J. Seedling production and the field performance of seedlings. Forests 2018, 9, 740. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.S.; Hwang, J.; Yang, A.; Han, S.; Son, Y. Seed germination and seedling survival rate of pinus densiflora and abies holophylla in open-field experimental warming using the infrared lamp. J. Kor. For. Soc. 2014, 103, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Kwon, B.; Cho, M.S.; Yang, A.-R.; Chang, H.; An, J.; Son, Y. The early growth performances of pinus densiflora and larix kaempferi seedlings under open-field experimental warming and precipitation manipulation. J. Korean Soc. For. Sci. 2020, 109, 31–40. [Google Scholar]
- Corcobado, T.; Cubera, E.; Juarez, E.; Moreno, G.; Solla, A. Drought events determine performance of quercus ilex seedlings and increase their susceptibility to phytophthora cinnamomic. Agiric. For. Meteorol. 2014, 192, 1–8. [Google Scholar]
- Hou, Q.Q.; Chen, B.M.; Peng, S.L.; Chen, L.Y. Effects of extreme temperature on seedling establishment of nonnative invasive plants. Biol. Invasions 2014, 16, 2049–2061. [Google Scholar] [CrossRef]
- Sukhbaatar, G.; Ganbaatar, B.; Jamsran, T.; Purevragchaa, B.; Nachin, B.; Gradel, A. Assessment of early survival and growth of planted scots pine (Pinus sylvestris) seedlings under extreme continental climate conditions of northern mongolia. J. For. Res. 2020, 31, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Takata, K.; Kurinobu, K.; Koisumi, A.; Yasue, K.; Tamai, Y.; Kisanuki, M. Bibliography on Japanese Larch (Larix kaempferi (Lamb.) Carr.). Eurasian J. For. Res. 2005, 8, 111–126. [Google Scholar]
- Korea Forest Service. Statistical Yearbook of Forestry; Korea Forest Service: Daejeon, Korea, 2020. (In Korean) [Google Scholar]
- Dulamsuren, C.; Hauck, M.; Leuschner, C. Recent drought stress leads to growth reductions in Larix sibirica in the Western Khentey, Mongolia. Glob. Change Biol. 2010, 16, 3024–3035. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway Spruce and European larch. Glob. Change Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Chen, D.; Shen, J.; Sun, X.; Zhang, S. Estimating the distribution and productivity characters of larix kaempferi in response to climate change. J. Environ. Manag. 2021, 280, 111633. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Chang, H.; Park, M.J.; Han, S.H.; Hwang, J.H.; Cho, M.S.; Son, Y. Effect of experimental warming on physiological and growth responses of larix kaempferi seedlings. J. Clim. Chang. Res. 2016, 7, 77–84. [Google Scholar] [CrossRef]
- Korea Meteorological Administration. Climate Data Open Portal. 2021. Available online: Data.kma.go.kr/cmmn/main.do (accessed on 30 September 2021).
- Korea Forest Service. Guidelines for Seed and Seedlings Management; Korea Forest Service: Daejeon, Korea, 2020. [Google Scholar]
- Alexander, L.V.; Zhang, X.; Peterson, T.C.; Caesar, J.; Gleason, B.; Klein Tank, A.M.G.; Haylock, M.; Collins, D.; Trewin, B.; Rahimzadeh, R.; et al. Global Observed changes in daily climate extremes of temperature and precipitation. J. Geophys. Res. 2006, 111, D05109. [Google Scholar] [CrossRef] [Green Version]
- Grant, K.; Kreyling, J.; Heilmeier, H.; Beierkuhnlein, C.; Jentsch, C. Extreme weather events and plant-plant interactions: Shifts between competition and facilitation among grassland species in the face of drought and heavy rainfall. Ecol. Res. 2014, 29, 991–1001. [Google Scholar] [CrossRef]
- Zakaria, R.; Radi, N.F.A.; Satari, S.Z. Extraction method of extreme rainfall Data. J. Phys. Conf. Ser. 2017, 890, 012154. [Google Scholar] [CrossRef]
- Pendergrass, A.G. What precipitation is extreme? Science 2018, 360, 1072–1073. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.J.; Jo, H.; Kim, H.; Cho, M.S.; Noh, N.J.; Chang, H.; Kim, H.S.; Son, Y. Open-field temperature and precipitation manipulations system design to simulate extreme climate events for plants and soils. For. Sci. Tech. 2021. under review. [Google Scholar]
- Pinto, J.R.; Marshall, J.D.; Dumroese, R.K.; Davis, A.S.; Cobos, D.R. Establishment and growth of container seedlings for reforestation: A function of stocktype and edaphic conditions. For. Ecol. Manag. 2011, 261, 1876–1884. [Google Scholar] [CrossRef]
- Deans, J.; Mason, W.; Cannell, M.; Sharpe, A.; Sheppard, L. Growing regimes for bare-root stock of sitka spruce, douglas-fir and scots pine. 1. morphology at the end of the nursery phase. Forestry 1989, 62, 53–60. [Google Scholar]
- Cho, M.S.; Lee, S.W.; Park, B.B. Effects of fertilization methods on the growth and physiological characteristics of larix kaempferi seedlings in the container nursery system. J. Bio-Environ. Con. 2012, 21, 57–65. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D.; R Core Team. nlme: Linear and Nonlinear Mixed Effects models. R Package Version 3, 2018, pp 1–137. Available online: http://CRAN.R-project.org/package=nlme (accessed on 1 September 2021).
- R Development Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2018. Available online: http://www.R-project.org (accessed on 1 September 2021).
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate forest trees and stands under severe drought: A review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef] [Green Version]
- Weemstra, M.; Eilmann, B.; Sass-Klaassen, U.G.W.; Sterch, F.J. Summer droughts limit tree growth across 10 temperate species on a productive forest site. For. Ecol. Manag. 2013, 306, 142–149. [Google Scholar] [CrossRef]
- Gazol, A.; Ribas, M.; Gutiérrez, E.; Camarero, J.J. Aleppo pine forests from across Spain show drought-induced growth decline partial recovery. Agric. For. Meteorol. 2017, 232, 186–194. [Google Scholar] [CrossRef]
- South, D.B.; Harris, S.W.; Barnett, J.P.; Hainds, M.J.; Gjerstad, D.H. Effect of container type and seedling size on survival and early height growth of pinus palustris seedlings in Alabama, U.S.A. For. Ecol. Manag. 2005, 204, 385–398. [Google Scholar] [CrossRef]
- Grossnickle, S.C.; El-Kassaby, Y.A. Bareroot versus container stocktypes: A performance comparison. New For. 2016, 47, 1–51. [Google Scholar] [CrossRef]
- Adams, H.D.; Collins, A.D.; Briggs, S.P.; Vennetier, M.; Dickman, L.T.; Sevanto, S.A.; Garcia-Forner, N.; Powers, H.H.; Mcdowell, N.G. Experimental drought and heat can delay phenological development and reduce foliar and shoot growth in semiarid trees. Glob. Chang. Biol. 2015, 21, 4210–4220. [Google Scholar] [CrossRef] [PubMed]
- Bigler, C.; Gavin, D.G.; Gunning, C.; Veblen, T.T. Drought induces lagged tree mortality in a subalpine forest in the rocky mountains. Oikos 2007, 116, 1983–1994. [Google Scholar] [CrossRef]
- Hoeppner, S.S.; Dukes, J.S. Interactive responses of open-field plant growth and composition to warming and precipitation. Glob. Chang. Biol. 2012, 18, 1754–1786. [Google Scholar] [CrossRef]
- Taeger, S.; Spark, T.H.; Menzel, A. Effects of temperature and drought manipulations on seedlings of scots pine provenances. Plant Biol. 2015, 17, 361–372. [Google Scholar] [CrossRef]
- Harter, D.E.; Nagy, L.; Backhaus, S.; Beierkuhnlein, C.; Fuss, B.; Huber, G.; Jentsch, A.; Konnert, M.; Thiel, D.; Kreyling, J.A. Comparison of genetic diversity and phenotypic plasticity among european beech (Fagus sylvatica L.) populations from Bulgaria and Germany under drought and temperature manipulation. Int. J. Plant Sci. 2015, 176, 232–244. [Google Scholar] [CrossRef]
- Noh, N.J.; Lee, S.J.; Jo, W.; Han, S.; Yoon, T.K.; Chung, H.; Muraoka, H.; Son, Y. Effects of experimental warming on soil respiration and biomass in Quercus variabilis blume and Pinus densiflora zieb. and zucc. seedling. Ann. For. Sic. 2016, 73, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Rustad, L.E.; Campbell, J.L.; Marion, G.M.; Norby, R.J.; Mitchell, M.J.; Hartley, A.E.; Cornelissen, J.H.C.; Gurevitch, J. A Meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 2001, 126, 543–562. [Google Scholar] [CrossRef]
- Lu, M.; Zhou, X.; Yang, Q.; Li, H.; Luo, Y.; Fang, C.; Chen, J.; Yang, X.; Li, B. Responses of ecosystem carbon cycle to experimental warming: A meta-analysis. Ecology 2013, 94, 726–738. [Google Scholar] [CrossRef] [PubMed]
- Ciais, P.; Reichstein, M.; Viovy, N.; Granier, A.; Ogee, J.; Allard, V.; Aubinet, M.; Buchmann, N.; Bernhofer, C.; Carrara, A.; et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 2003, 437, 529–533. [Google Scholar] [CrossRef]
- Voesenek, L.A.C.J.; Colmer, T.D.; Pierik, R.; Millenaar, F.F.; Peeters, A.J.M. How plants cope with complete submergence. N. Phytol. 2006, 170, 213–226. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, H.; Chang, H.; An, J.; Cho, M.S.; Son, Y. Species specific physiological responses of pinus densiflora and larix kaempferi seedlings to open-field experimental warming and precipitation manipulation. For. Sci. Tech. 2019, 15, 44–50. [Google Scholar] [CrossRef] [Green Version]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Levesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Matisons, R.; Krišāns, O.; Jansons, Ā.; Kondratovičs, T.; Elferts, D.; Ievinsh, G. Norway spruce seedlings from an Eastern Baltic provenance show tolerance to simulated drought. Forests 2021, 12, 82. [Google Scholar] [CrossRef]
- Way, D.A.; Oren, R. Differential Responses to Changes in Growth Temperature between trees from different functional groups and biomes: A review and synthesis of data. Tree Physiol. 2010, 30, 669–688. [Google Scholar] [CrossRef] [Green Version]
- Arend, M.; Kuster, T.; Günthardt-Goerg, M.S.; Dobbertin, M. Provenance-specific growth responses to drought and air warming in three european oak species (Quercus robur, Q. pertaea and Q. pubescens). Tree Physiol. 2011, 31, 287–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noh, N.J.; Crous, K.Y.; Li, J.; Choury, Z.; Barton, C.V.M.; Arndt, S.K.; Reich, P.B.; Tjoelker, M.G.; Pendall, E. Does root respiration in australian rainforest tree seedlings acclimate to experimental warming? Tree Physiol. 2020, 40, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Cui, K.; Xu, A.; Nie, L.; Huang, J.; Peng, S. Drought stress condition increases root to shoot ratio via alteration of carbohydrate partitioning and enzymatic activity. Acta Physiol. Plant 2015, 37, 9. [Google Scholar] [CrossRef]
- Aaltonen, H.; Lindén, A.; Heinonsalo, J.; Biasi, C.; Pumpanen, J. Effect of prolonged drought stress on scots pine seedling carbon allocation. Tree Physiol. 2016, 37, 418–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargallo-Garriga, A.; Sardans, J.; Pérez-Trujillo, M.; Oravec, M.; Urban, O.; Jentsch, A.; Kreyling, J.; Beierkuhnlein, C.; Parella, T.; Peñuelas, J. Warming differentially influences the effects of drought on stoichiometry and metabolomics in shoots and roots. New Phytol. 2015, 207, 591–603. [Google Scholar] [CrossRef]
- Granda, E.; Camarero, J.J.; Gimeno, T.E.; Martínez-Fernández, J.; Valladares, F. Intensity and timing of warming and drought differentially affect growth patterns of co-occuring mediterranean tree species. Eur. J. For. Res. 2013, 132, 469–480. [Google Scholar] [CrossRef]
- Grossiord, C.; Sevanto, S.; Dawson, T.E.; Adams, H.D.; Collins, A.D.; Dickman, L.T.; Newman, B.D.; Stockton, E.A.; McDowell, N.G. Warming combined with more extreme precipitation regimes modifies the water sources used by trees. New Phytol. 2017, 213, 584–596. [Google Scholar] [CrossRef]
- Liu, Y.; Li, P.; Xiao, L.; Wang, W.; Yu, K.; Shi, P. Heterogeneity in short-term allocation of carbon to roots of Pinus tabuliformis seedlings and root respiration under drought stress. Plant Soil 2020, 452, 359–378. [Google Scholar] [CrossRef]
Figure 1.
(a) Overview of the open-field experiment of extreme climate event manipulation; (b) one-year-old Larix kaempferi (Lamb.) Carr. seedlings (n = 88) planted in a plot of 1.5 × 1.0 m; and (c) climate manipulation systems using infrared heating lamps for warming, rain shelters for drought, and irrigation spraying for heavy rainfall treatments.
Figure 1.
(a) Overview of the open-field experiment of extreme climate event manipulation; (b) one-year-old Larix kaempferi (Lamb.) Carr. seedlings (n = 88) planted in a plot of 1.5 × 1.0 m; and (c) climate manipulation systems using infrared heating lamps for warming, rain shelters for drought, and irrigation spraying for heavy rainfall treatments.

Figure 2.
Open-field experimental design of extreme climate events regarding temperature and precipitation using L. kaempferi seedlings.
Figure 2.
Open-field experimental design of extreme climate events regarding temperature and precipitation using L. kaempferi seedlings.

Figure 3.
(a) Mean soil temperature (°C) and (b) mean soil water content (vol %) during the experimental period of extreme climate events. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C extreme warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Results of a two-way ANOVA are shown in each panel. Asterisks indicate significant differences between treatments; n.s.: not significant. Different letters indicate significant differences between treatment within each group factor. Error bars indicate the standard deviation of the mean (n = 3).
Figure 3.
(a) Mean soil temperature (°C) and (b) mean soil water content (vol %) during the experimental period of extreme climate events. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C extreme warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Results of a two-way ANOVA are shown in each panel. Asterisks indicate significant differences between treatments; n.s.: not significant. Different letters indicate significant differences between treatment within each group factor. Error bars indicate the standard deviation of the mean (n = 3).

Figure 4.
(a) Survival rate (%), (b) root collar diameter (mm), and (c) height (mm) of L. kaempferi seedlings subjected to extreme climate events, measured in October. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C extreme warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Results from two-way ANOVA are shown in each panel. Asterisks indicate significant differences between treatments; n.s.: not significant. Different letters in (c) indicate significant differences between treatments. Bars indicate standard errors of the mean.
Figure 4.
(a) Survival rate (%), (b) root collar diameter (mm), and (c) height (mm) of L. kaempferi seedlings subjected to extreme climate events, measured in October. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C extreme warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Results from two-way ANOVA are shown in each panel. Asterisks indicate significant differences between treatments; n.s.: not significant. Different letters in (c) indicate significant differences between treatments. Bars indicate standard errors of the mean.

Figure 5.
(a) Relationship between root collar diameter and height and (b) shoot to root ratios at the end of the treatment of L. kaempferi seedlings subjected to extreme climate events. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Results from the ANOVA and Pearson correlation analysis are shown in panel (a). Asterisks indicate significant differences between warming treatments. Different letters indicate significant differences between treatments. The boxes’ lower and upper limits indicate the 25% and 75% quartile, respectively. Bold lines within the boxes indicate the median, whiskers indicate the range of values from the minimum to maximum, and data points beyond the end of the whiskers are then plotted as filled dots.
Figure 5.
(a) Relationship between root collar diameter and height and (b) shoot to root ratios at the end of the treatment of L. kaempferi seedlings subjected to extreme climate events. TC: ambient temperature, T3: +3 °C warming, T6: +6 °C warming, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Results from the ANOVA and Pearson correlation analysis are shown in panel (a). Asterisks indicate significant differences between warming treatments. Different letters indicate significant differences between treatments. The boxes’ lower and upper limits indicate the 25% and 75% quartile, respectively. Bold lines within the boxes indicate the median, whiskers indicate the range of values from the minimum to maximum, and data points beyond the end of the whiskers are then plotted as filled dots.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Statistical significance (p-values) of the two-way ANOVA of the effects of experimental extreme climate events on the measured variables.
Table 1.
Statistical significance (p-values) of the two-way ANOVA of the effects of experimental extreme climate events on the measured variables.
| Treatment | d.f. 1 | Growth Responses | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Survival | RCD | Height | Volume | H/D | Total Biomass | S/R 2 | SQI 3 | ||
| Temperature (T) | 2 | 0.305 | 0.258 | 0.270 | 0.176 | 0.373 | 0.881 | 0.419 | 0.498 |
| Precipitation (P) | 2 | 0.576 | 0.598 | <0.001 | 0.155 | <0.001 | 0.955 | 0.390 | 0.145 |
| T × P | 4 | 0.967 | 0.035 | <0.001 | <0.001 | <0.001 | 0.785 | 0.788 | 0.562 |
1 d.f.: degree of freedom, 2 S/R: shoot to root biomass ratio, 3 SQI: seedling quality index.
Table 2.
Biomass and seedling quality index (SQI) of L. kaempferi (Lamb.) Carr. seedlings under extreme climate events regarding temperature and precipitation. TC: ambient temperature, T3: warming by 3 °C, T6: warming by 6 °C, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Values are means ± S.D. (n = 9). There was no significant difference in the variables between treatments at 5% levels using a Tukey HSD test.
Table 2.
Biomass and seedling quality index (SQI) of L. kaempferi (Lamb.) Carr. seedlings under extreme climate events regarding temperature and precipitation. TC: ambient temperature, T3: warming by 3 °C, T6: warming by 6 °C, DR: drought, PC: ambient precipitation, HR: heavy rainfall. Values are means ± S.D. (n = 9). There was no significant difference in the variables between treatments at 5% levels using a Tukey HSD test.
| Treatment | Leaf Biomass (g) | Stem Biomass (g) | Root Biomass (g) | Total Biomass (g) | SQI | |
|---|---|---|---|---|---|---|
| TC | DR | 4.06 ± 0.42 | 7.03 ± 1.27 | 3.76 ± 0.74 | 14.87 ± 2.00 | 1.23 ± 0.23 |
| PC | 4.80 ± 0.90 | 7.17 ± 1.43 | 4.21 ± 1.07 | 15.88 ± 3.25 | 1.37 ± 0.29 | |
| HR | 4.37 ± 0.71 | 6.30 ± 1.08 | 3.90 ± 0.92 | 14.57 ± 2.50 | 1.37 ± 0.29 | |
| T3 | DR | 4.39 ± 1.10 | 7.11 ± 1.82 | 3.74 ± 0.75 | 15.24 ± 3.47 | 1.24 ± 0.20 |
| PC | 4.38 ± 1.28 | 6.88 ± 1.73 | 4.12 ± 0.73 | 15.38 ± 3.35 | 1.34 ± 0.22 | |
| HR | 4.17 ± 1.10 | 7.12 ± 1.46 | 3.84 ± 1.42 | 15.13 ± 3.48 | 1.21 ± 0.32 | |
| T6 | DR | 4.21 ± 0.55 | 6.87 ± 1.38 | 3.84 ± 1.23 | 14.93 ± 2.95 | 1.20 ± 0.29 |
| PC | 5.03 ± 1.33 | 6.64 ± 1.22 | 4.08 ± 1.43 | 15.75 ± 3.78 | 1.42 ± 0.47 | |
| HR | 4.09 ± 0.58 | 6.03 ± 1.05 | 3.12 ± 0.55 | 13.24 ± 1.27 | 1.09 ± 0.09 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Noh, N.-J.; Kim, G.-J.; Son, Y.; Cho, M.-S. Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer. Forests 2021, 12, 1595. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111595
AMA Style
Noh N-J, Kim G-J, Son Y, Cho M-S. Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer. Forests. 2021; 12(11):1595. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111595
Chicago/Turabian StyleNoh, Nam-Jin, Gwang-Jung Kim, Yowhan Son, and Min-Seok Cho. 2021. "Early Growth Responses of Larix kaempferi (Lamb.) Carr. Seedling to Short-Term Extreme Climate Events in Summer" Forests 12, no. 11: 1595. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111595
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.