
The Radial Growth of Picea wilsonii Was More Restricted by Precipitation Due to Climate Warming on Mt. Guandi, China

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
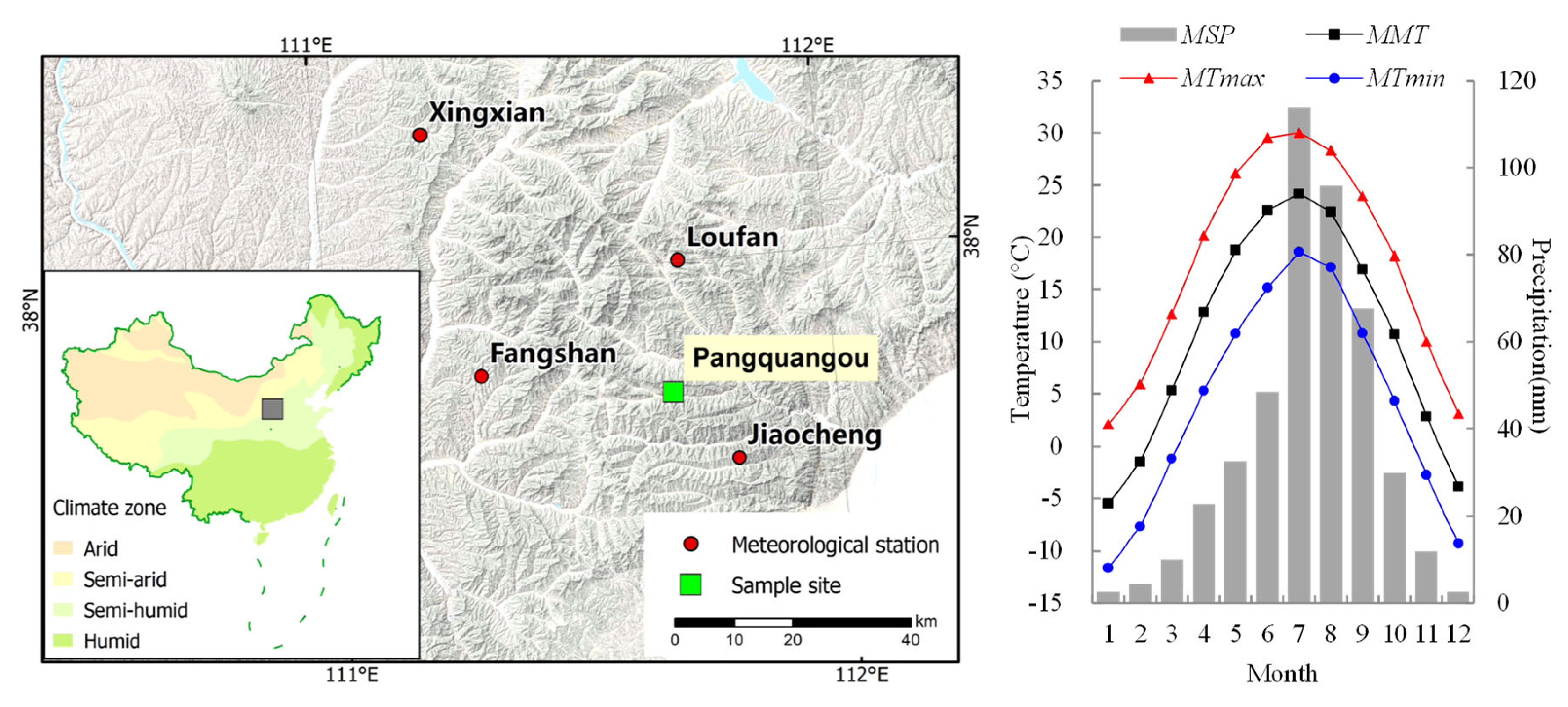
2.1. Study Area
2.2. Tree-Ring Data Sampling
2.3. Chronology Development
2.4. Climate Data
2.5. Data Analysis
3. Results
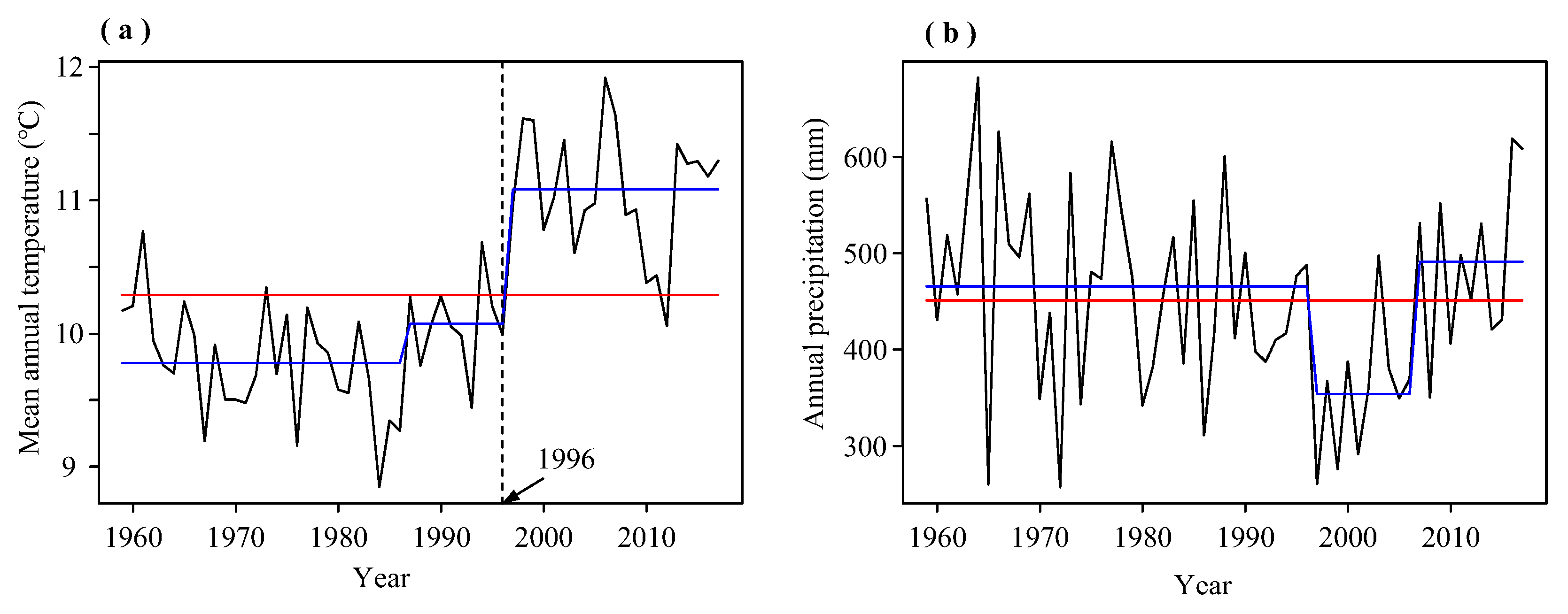
3.1. Climate Variations in the Study Area
3.2. Chronologies Statistics of Picea wilsonii
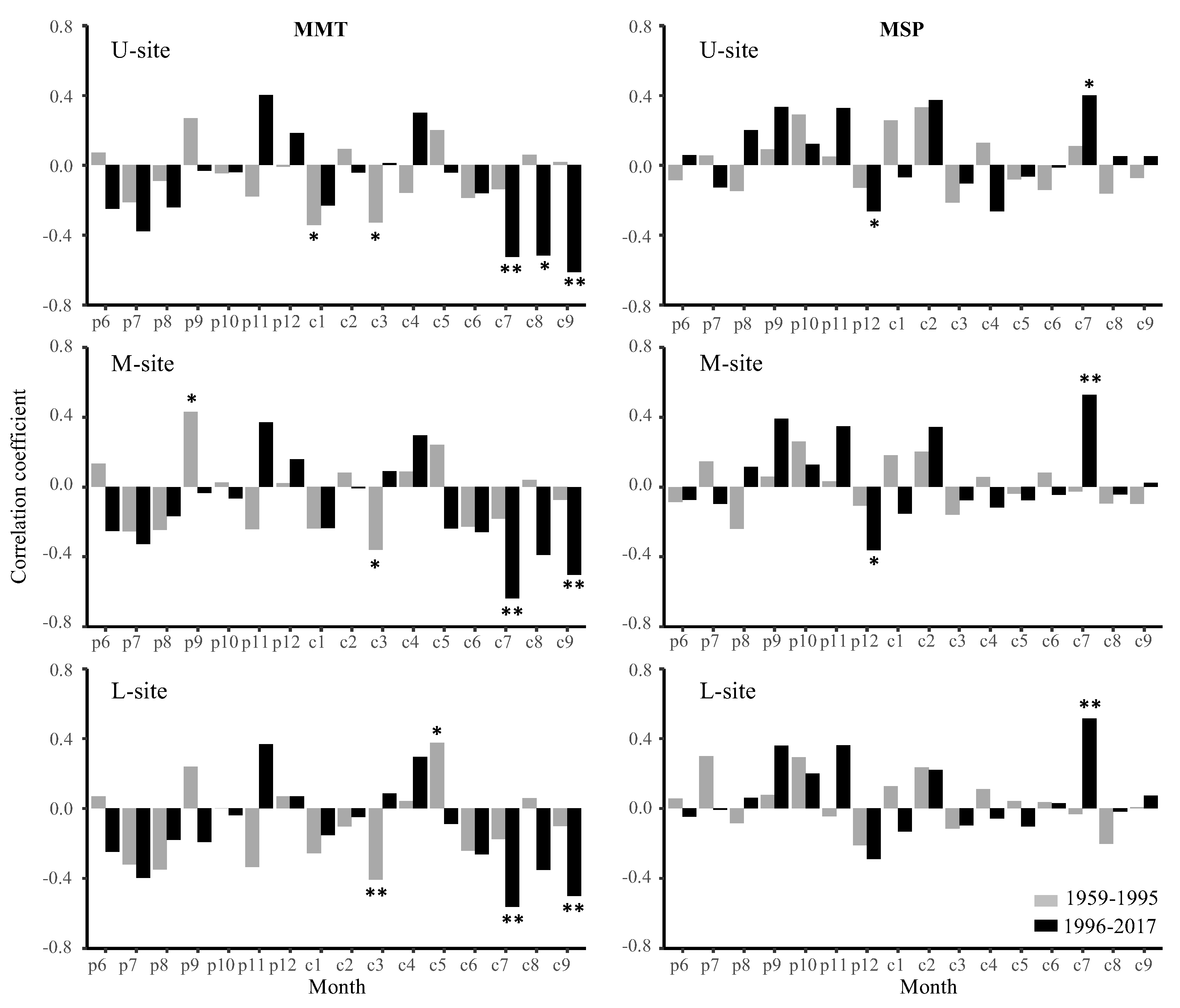
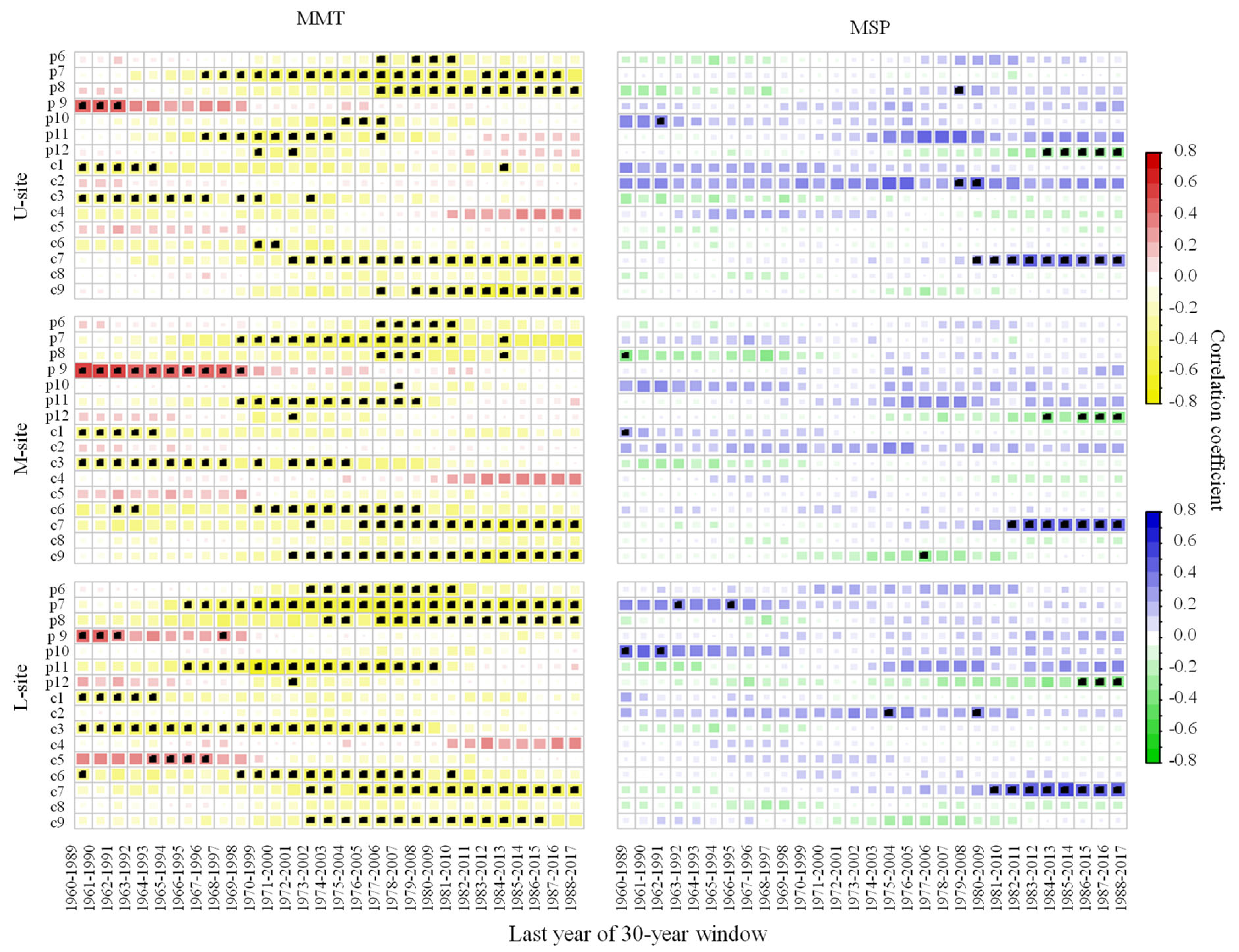
3.3. Tree Ring—Climate Relationship Changes with Increasing Temperature
3.3.1. Years 1959–1995 versus 1996–2017
3.3.2. Moving Correlations between Radial Growth and Climate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated Dryland Expansion under Climate Change. Nat. Clim. Chang. 2016, 6, 166–171. [Google Scholar] [CrossRef]
- Huang, W.; Yan, J.; Liu, C.; Xie, T. Changes in Climate Regimes over China Based on a High-Resolution Dataset. Sci. Bull. 2019, 64, 377–379. [Google Scholar] [CrossRef] [Green Version]
- Pachauri, R.K.; Meyer, L.A.; Core Writing Team (Eds.) Climate Change 2014 Synthesis Report. Contribution of Working Groups I, II, and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; IPCC: Geneva, Switzerland, 2014; ISBN 978-92-9169-143-2. [Google Scholar]
- Sun, C.; Jiang, Z.; Li, W.; Hou, Q.; Li, L. Changes in Extreme Temperature over China When Global Warming Stabilized at 1.5 °C and 2.0 °C. Sci. Rep. 2019, 9, 14982. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Ma, J.; Guan, X.; Li, Y.; He, Y. Progress in Semi-Arid Climate Change Studies in China. Adv. Atmos. Sci. 2019, 36, 922–937. [Google Scholar] [CrossRef]
- Wilmking, M.; Maaten-Theunissen, M.; Maaten, E.; Scharnweber, T.; Buras, A.; Biermann, C.; Gurskaya, M.; Hallinger, M.; Lange, J.; Shetti, R.; et al. Global Assessment of Relationships between Climate and Tree Growth. Glob. Chang. Biol. 2020, 26, 3212–3220. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bachelet, D.; McDowell, N.; Vennetier, M.; Kitzberger, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A Global Overview of Drought and Heat-Induced Tree Mortality Reveals Emerging Climate Change Risks for Forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- Choat, B.; Brodribb, T.J.; Brodersen, C.R.; Duursma, R.A.; López, R.; Medlyn, B.E. Triggers of Tree Mortality under Drought. Nature 2018, 558, 531–539. [Google Scholar] [CrossRef]
- Liu, H.; Park Williams, A.; Allen, C.D.; Guo, D.; Wu, X.; Anenkhonov, O.A.; Liang, E.; Sandanov, D.V.; Yin, Y.; Qi, Z.; et al. Rapid Warming Accelerates Tree Growth Decline in Semi-Arid Forests of Inner Asia. Glob. Chang. Biol. 2013, 19, 2500–2510. [Google Scholar] [CrossRef]
- Liang, E.; Leuschner, C.; Dulamsuren, C.; Wagner, B.; Hauck, M. Global Warming-Related Tree Growth Decline and Mortality on the North-Eastern Tibetan Plateau. Clim. Chang. 2016, 134, 163–176. [Google Scholar] [CrossRef]
- Zhao, S.; Jiang, Y.; Dong, M.; Xu, H.; Manzanedo, R.D.; Pederson, N. Early Monsoon Failure and Mid-Summer Dryness Induces Growth Cessation of Lower Range Margin Picea crassifolia. Trees 2018, 32, 1401–1413. [Google Scholar] [CrossRef]
- Salzer, M.W.; Hughes, M.K.; Bunn, A.G.; Kipfmueller, K.F. Recent Unprecedented Tree-Ring Growth in Bristlecone Pine at the Highest Elevations and Possible Causes. Proc. Natl. Acad. Sci. USA 2009, 106, 20348–20353. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Chen, Q.; Gou, X.; Du, M.; Wang, F.; Zhang, J. Climatic Control on the Growth and Regeneration of Juniperus Przewalskii at Alpine Treeline in the Eastern Qilian Mountains, Northwest China. Trees 2021, 35, 1085–1097. [Google Scholar] [CrossRef]
- Cao, J.; Liu, H.; Zhao, B.; Li, Z.; Drew, D.M.; Zhao, X. Species-Specific and Elevation-Differentiated Responses of Tree Growth to Rapid Warming in a Mixed Forest Lead to a Continuous Growth Enhancement in Semi-Humid Northeast Asia. For. Ecol. Manag. 2019, 448, 76–84. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Wang, M.; Zhang, L.; Dong, M. Topography- and Species-Dependent Climatic Responses in Radial Growth of Picea meyeri and Larix principis-rupprechtii in the Luyashan Mountains of North-Central China. Forests 2015, 6, 116–132. [Google Scholar] [CrossRef] [Green Version]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK; New York, NY, USA, 1976; ISBN 978-0-12-268450-0. [Google Scholar]
- Barber, V.A.; Juday, G.P.; Finney, B.P. Reduced Growth of Alaskan White Spruce in the Twentieth Century from Temperature-Induced Drought Stress. Nature 2000, 405, 668–673. [Google Scholar] [CrossRef] [PubMed]
- Jochner, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Tree Growth Responses to Changing Temperatures across Space and Time: A Fine-Scale Analysis at the Treeline in the Swiss Alps. Trees 2018, 32, 645–660. [Google Scholar] [CrossRef]
- Liang, E.; Camarero, J.J. Threshold-Dependent and Non-Linear Associations between Temperature and Tree Growth at and below the Alpine Treeline. Trees 2018, 32, 661–662. [Google Scholar] [CrossRef]
- Zhu, L.; Cooper, D.J.; Yang, J.; Zhang, X.; Wang, X. Rapid Warming Induces the Contrasting Growth of Yezo Spruce (Picea jezoensis var. microsperma) at Two Elevation Gradient Sites of Northeast China. Dendrochronologia 2018, 50, 52–63. [Google Scholar] [CrossRef]
- Gai, X.; Wang, S.; Zhou, L.; Wu, J.; Zhou, W.; Bi, J.; Cao, L.; Dai, L.; Yu, D. Spatiotemporal Evidence of Tree-Growth Resilience to Climate Variations for Yezo Spruce (Picea jezoensis var. Komarovii) on Changbai Mountain, Northeast China. J. For. Res. 2020, 31, 927–936. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, Y.; Dong, M.; Kang, M.; Yang, H. Relationship between the Radial Growth of Picea meyeri and Climate along Elevations of the Luyashan Mountain in North-Central China. For. Ecol. Manag. 2012, 265, 142–149. [Google Scholar] [CrossRef]
- Wang, B.; Yu, P.; Zhang, L.; Wang, Y.; Yu, Y.; Wang, S. Differential Trends of Qinghai Spruce Growth with Elevation in Northwestern China during the Recent Warming Hiatus. Forests 2019, 10, 712. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Zhang, W.; Wang, M.; Kang, M.; Dong, M. Radial Growth of Two Dominant Montane Conifer Tree Species in Response to Climate Change in North-Central China. PLoS ONE 2014, 9, e112537. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Chen, L.; Wang, J.; Wang, M.; Yang, S.; Zhao, C. Photosynthetic Acclimation to Long-Term High Temperature and Soil Drought Stress in Two Spruce Species (Picea crassifolia and P. Wilsonii) Used for Afforestation. J. For. Res. 2018, 29, 363–372. [Google Scholar] [CrossRef]
- Niu, H.G.; Zhang, F.; Yu, A.L.; Wang, F.; Zhang, J.Z.; Gou, X.H. Intra-annual stem radial growth dynamics of Picea wilsori in response to climate in the eastern Qilian Mountains. Acta Ecol. Sin. 2018, 38, 7412–7420. [Google Scholar] [CrossRef]
- Huang, J.; Li, Y.; Fu, C.; Chen, F.; Fu, Q.; Dai, A.; Shinoda, M.; Ma, Z.; Guo, W.; Li, Z.; et al. Dryland Climate Change: Recent Progress and Challenges: Dryland Climate Change. Rev. Geophys. 2017, 55, 719–778. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Raven, P.H. Flora of China; Science Press: Beijing, China, 1999; Volume 4, ISBN 978-0-915279-70-8. [Google Scholar]
- Bohner, J. General Climatic Controls and Topoclimatic Variations in Central and High Asia. Boreas 2008, 35, 279–295. [Google Scholar] [CrossRef]
- IUSS Working Group WRB. World Reference Base for Soil Resources 2006; World Soil Resources Reports No. 103; Food and Agriculture Organization: Rome, Italy, 2006; Available online: https://soilgrids.org/ (accessed on 15 October 2021).
- Grissino-Mayer, H. Evaluating Crossdating Accuracy: A Manual and Tutorial for the Computer Program COFECHA. Tree Ring Res. 2001, 57, 205–221. [Google Scholar]
- Cook, E.R.; Kairiukstis, L.A. (Eds.) Methods of Dendrochronology: Applications in the Environmental Sciences; Springer: Berlin/Heidelberg, Germany, 1990; ISBN 13978-0-7923-0586-6. [Google Scholar]
- Biondi, F.; Waikul, K. DENDROCLIM2002: A C++ Program for Statistical Calibration of Climate Signals in Tree-Ring Chronologies. Comput. Geosci. 2004, 30, 303–311. [Google Scholar] [CrossRef]
- Bunn, A.G. A Dendrochronology Program Library in R (DplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Navarro, R.M.; Camarero, J.J.; Fernández-Cancio, Á. Drought-Induced Growth Decline of Aleppo and Maritime Pine Forests in South-Eastern Spain. For. Syst. 2010, 19, 458–469. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct Effects of Climate Warming on Populations of Silver Fir (Abies Alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef] [Green Version]
- Jiao, L.; Jiang, Y.; Wang, M.; Zhang, W.; Zhang, Y. Age-Effect Radial Growth Responses of Picea schrenkiana to Climate Change in the Eastern Tianshan Mountains, Northwest China. Forests 2017, 8, 294. [Google Scholar] [CrossRef] [Green Version]
- Cook, E.R.; Pederson, N. Uncertainty, Emergence, and Statistics in Dendrochronology. In Dendroclimatology; Hughes, M.K., Swetnam, T.W., Diaz, H.F., Eds.; Developments in Paleoenvironmental Research; Springer: Dordrecht, The Netherlands, 2011; Volume 11, pp. 77–112. ISBN 978-1-4020-4010-8. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Bunn, A.G.; Jansma, E.; Korpela, M.; Westfall, R.D.; Baldwin, J. Using Simulations and Data to Evaluate Mean Sensitivity (ζ) as a Useful Statistic in Dendrochronology. Dendrochronologia 2013, 31, 250–254. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R.-project.org/ (accessed on 31 March 2021).
- Zhang, X.; Manzanedo, R.D.; Xu, C.; Hou, M.; Huang, X. How to Select Climate Data for Calculating Growth-Climate Correlation. Trees 2021, 35, 1199–1206. [Google Scholar] [CrossRef]
- Zeileis, A.; Leisch, F.; Hornik, K.; Kleiber, C. Strucchange: An R Package for Testing for Structural Change in Linear Regression Models. J. Stat. Soft 2002, 7, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Militino, A.F.; Moradi, M.; Ugarte, M.D. On the Performances of Trend and Change-Point Detection Methods for Remote Sensing Data. Remote Sens. 2020, 12, 1008. [Google Scholar] [CrossRef] [Green Version]
- Dang, H.; Zhang, Y.; Zhang, K.; Jiang, M.; Zhang, Q. Climate-Growth Relationships of Subalpine Fir (Abies Fargesii) across the Altitudinal Range in the Shennongjia Mountains, Central China. Clim. Chang. 2013, 117, 903–917. [Google Scholar] [CrossRef]
- Biondi, F. Are Climate-Tree Growth Relationships Changing in North-Central Idaho, U.S.A.? Arct. Antarct. Alp. Res. 2000, 32, 111–116. [Google Scholar] [CrossRef]
- Dixon, P.M. Bootstrap resampling. In The Encyclopedia of Environmetrics; El-Shaarawi, A.H., Piegorsch, W.W., Eds.; Wiley: New York, NY, USA, 2001. [Google Scholar]
- Zeileis, A.; Shah, A.; Patnaik, I. Testing, Monitoring, and Dating Structural Changes in Exchange Rate Regimes. Comput. Stat. Data Anal. 2010, 54, 1696–1706. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. Dendroclimatic Calibration in R: The BootRes Package for Response and Correlation Function Analysis. Dendrochronologia 2013, 31, 68–74. [Google Scholar] [CrossRef]
- Li, Z.S.; Liu, G.H.; Fu, B.J.; Zhang, Q.B.; Hu, C.J. Influence of Different Detrending Methods on Climate Signal in Tree-Ring Chronologies in Wolong National Natural Reserve, Western Sichuan, China. Chin. J. Plant Ecol. 2011, 35, 707. [Google Scholar] [CrossRef]
- Björklund, J.; Seftigen, K.; Schweingruber, F.; Fonti, P.; Arx, G.; Bryukhanova, M.V.; Cuny, H.E.; Carrer, M.; Castagneri, D.; Frank, D.C. Cell Size and Wall Dimensions Drive Distinct Variability of Earlywood and Latewood Density in Northern Hemisphere Conifers. New Phytol. 2017, 216, 728–740. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.T.; Jiang, Y.; Wang, M.C.; Zhang, L.N.; Dong, M.Y. Responses of radial growth in Larix principis-rupprechti to climate change along an elevation gradient on the southern slope of Luya Mountain. Acta Ecol. Sin. 2015, 35, 6481–6488. [Google Scholar]
- Fang, K.; Gou, X.; Chen, F.; Li, Y.; Zhang, F.; Kazmer, M. Tree Growth and Its Association with Climate between Individual Tree-Ring Series at Three Mountain Ranges in North Central China. Dendrochronologia 2012, 30, 113–119. [Google Scholar] [CrossRef]
- Schurman, J.S.; Babst, F.; Björklund, J.; Rydval, M.; Bače, R.; Čada, V.; Janda, P.; Mikolas, M.; Saulnier, M.; Trotsiuk, V.; et al. The Climatic Drivers of Primary Picea Forest Growth along the Carpathian Arc Are Changing under Rising Temperatures. Glob. Chang. Biol. 2019, 25, 3136–3150. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.H.; Bunn, A.G. Responses of the Circumpolar Boreal Forest to 20th Century Climate Variability. Environ. Res. Lett. 2007, 2, 045013. [Google Scholar] [CrossRef]
- Zhang, Y.; Wilmking, M.; Gou, X. Changing Relationships between Tree Growth and Climate in Northwest China. Plant Ecol. 2009, 201, 39–50. [Google Scholar] [CrossRef]
- Li, G.-Q.; Bai, F.; Sang, W.-G. Different Responses of Radial Growth to Climate Warming in Pinus Koraiensis and Picea jezoensis Var. komarovii at Their Upper Elevational Limits in Changbai Mountain, China. Chin. J. Plant Ecol. 2011, 35, 500–511. [Google Scholar] [CrossRef]
- Ribbons, R.R. Disturbance and Climatic Effects on Red Spruce Community Dynamics at its Southern Continuous Range Margin. PeerJ 2014, 2, e293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, K.; Gou, X.; Chen, F.; Yang, M.; Li, J.; He, M.; Zhang, Y.; Tian, Q.; Peng, J. Drought Variations in the Eastern Part of Northwest China over the Past Two Centuries: Evidence from Tree Rings. Clim. Res. 2009, 38, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Rathgeber, C.B.K. Conifer Tree-Ring Density Inter-Annual Variability—Anatomical, Physiological and Environmental Determinants. New Phytol. 2017, 216, 621–625. [Google Scholar] [CrossRef] [Green Version]
- Kagawa, A.; Sugimoto, A.; Maximov, T.C. 13CO2 Pulse-Labelling of Photoassimilates Reveals Carbon Allocation within and between Tree Rings. Plant Cell. Environ. 2006, 29, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Gessler, A.; Brandes, E.; Buchmann, N.; Helle, G.; Rennenberg, H.; Barnard, R.L. Tracing Carbon and Oxygen Isotope Signals from Newly Assimilated Sugars in the Leaves to the Tree-Ring Archive. Plant Cell. Environ. 2009, 32, 780–795. [Google Scholar] [CrossRef]
- Kuptz, D.; Fleischmann, F.; Matyssek, R.; Grams, T.E.E. Seasonal Patterns of Carbon Allocation to Respiratory Pools in 60-Yr-Old Deciduous (Fagus Sylvatica) and Evergreen (Picea abies) Trees Assessed via Whole-Tree Stable Carbon Isotope Labeling. New Phytol. 2011, 191, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Cuny, H.E.; Rathgeber, C.B.K.; Frank, D.; Fonti, P.; Fournier, M. Kinetics of Tracheid Development Explain Conifer Tree-Ring Structure. New Phytol. 2014, 203, 1231–1241. [Google Scholar] [CrossRef]
- Viherä-Aarnio, A.; Sutinen, S.; Partanen, J.; Häkkinen, R. Internal Development of Vegetative Buds of Norway Spruce Trees in Relation to Accumulated Chilling and Forcing Temperatures. Tree Physiol. 2014, 34, 547–556. [Google Scholar] [CrossRef] [Green Version]
- Delpierre, N.; Lireux, S.; Hartig, F.; Camarero, J.J.; Cheaib, A.; Čufar, K.; Cuny, H.; Deslauriers, A.; Fonti, P.; Gričar, J.; et al. Chilling and Forcing Temperatures Interact to Predict the Onset of Wood Formation in Northern Hemisphere Conifers. Glob. Chang. Biol. 2019, 25, 1089–1105. [Google Scholar] [CrossRef]
- Fu, Y.H.; Zhao, H.; Piao, S.; Peaucelle, M.; Peng, S.; Zhou, G.; Ciais, P.; Huang, M.; Menzel, A.; Peñuelas, J.; et al. Declining Global Warming Effects on the Phenology of Spring Leaf Unfolding. Nature 2015, 526, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Piao, S.; Liu, Q.; Chen, A.; Janssens, I.A.; Fu, Y.; Dai, J.; Liu, L.; Lian, X.; Shen, M.; Zhu, X. Plant Phenology and Global Climate Change: Current Progresses and Challenges. Glob. Chang. Biol. 2019, 25, 1922–1940. [Google Scholar] [CrossRef]
- Jeong, S.-H.; Eom, J.-Y.; Park, J.-Y.; Chun, J.-H.; Lee, J.-S. Effect of Precipitation on Soil Respiration in a Temperate Broad-Leaved Forest. J. Ecol. Environ. 2018, 42, 10. [Google Scholar] [CrossRef] [Green Version]
- Martin, J.; Looker, N.; Hoylman, Z.; Jencso, K.; Hu, J. Differential Use of Winter Precipitation by Upper and Lower Elevation Douglas Fir in the Northern Rockies. Glob. Chang. Biol. 2018, 24, 5607–5621. [Google Scholar] [CrossRef]
- Griesbauer, H.P.; Green, D.S. Geographic and Temporal Patterns in White Spruce Climate–Growth Relationships in Yukon, Canada. For. Ecol. Manag. 2012, 267, 215–227. [Google Scholar] [CrossRef]
- Buma, B.; Hennon, P.E.; Harrington, C.A.; Popkin, J.R.; Krapek, J.; Lamb, M.S.; Oakes, L.E.; Saunders, S.; Zeglen, S. Emerging Climate-Driven Disturbance Processes: Widespread Mortality Associated with Snow-to-Rain Transitions across 10 of Latitude and Half the Range of a Climate-Threatened Conifer. Glob. Chang. Biol. 2017, 23, 2903–2914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, I.T.; Cayan, D.R.; Dettinger, M.D. Changes in Snowmelt Runoff Timing in Western North America under a Business as Usual’ Climate Change Scenario. Clim. Chang. 2004, 62, 217–232. [Google Scholar] [CrossRef]
- Beier, C.M.; Sink, S.E.; Hennon, P.E.; D’Amore, D.V.; Juday, G.P. Twentieth-Century Warming and the Dendroclimatology of Declining Yellow-Cedar Forests in Southeastern Alaska. Can. J. For. Res. 2008, 38, 1319–1334. [Google Scholar] [CrossRef] [Green Version]
- Comeau, V.M.; Daniels, L.D.; Knochenmus, G.; Chavardès, R.D.; Zeglen, S. Tree-Rings Reveal Accelerated Yellow-Cedar Decline with Changes to Winter Climate after 1980. Forests 2019, 10, 1085. [Google Scholar] [CrossRef] [Green Version]
- Decker, K.L.M.; Wang, D.; Waite, C.; Scherbatskoy, T. Snow Removal and Ambient Air Temperature Effects on Forest Soil Temperatures in Northern Vermont. Soil Sci. Soc. Am. J. 2003, 67, 1234–1242. [Google Scholar] [CrossRef] [Green Version]
- Misson, L. Dendroecological Analysis of Climatic Effects on Quercus Petraea and Pinus Halepensis Radial Growth Using the Process-Based MAIDEN Model. Can. J. For. Res. 2004, 34, 888–898. [Google Scholar] [CrossRef] [Green Version]
- Skre, O.; Nes, K. Combined Effects of Elevated Winter Temperatures and CO2 on Norway Spruce Seedlings. Silva Fenn. 1996, 30, 135–143. [Google Scholar] [CrossRef]
- Zheng, L.; Gaire, N.P.; Shi, P. High-Altitude Tree Growth Responses to Climate Change across the Hindu Kush Himalaya. J. Plant Ecol. 2021, 14, 829–842. [Google Scholar] [CrossRef]
- Yang, B.; He, M.; Melvin, T.M.; Zhao, Y.; Briffa, K.R. Climate Control on Tree Growth at the Upper and Lower Treelines: A Case Study in the Qilian Mountains, Tibetan Plateau. PLoS ONE 2013, 8, e69065. [Google Scholar] [CrossRef]
- Yu, D.; Wang, Q.; Wang, Y.; Zhou, W.; Ding, H.; Fang, X.; Jiang, S.; Dai, L. Climatic Effects on Radial Growth of Major Tree Species on Changbai Mountain. Ann. For. Sci. 2011, 68, 921–933. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, W.; Huang, G.; Zeng, G. Changes of the Transitional Climate Zone in East Asia: Past and Future. Clim. Dyn. 2017, 49, 1463–1477. [Google Scholar] [CrossRef]
- Jiao, L.; Liu, X.; Wang, S.; Chen, K. Radial Growth Adaptability to Drought in Different Age Groups of Picea schrenkiana Fisch. & C.A. Mey in the Tianshan Mountains of Northwestern China. Forests 2020, 11, 455. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jiang, Y.; Zhao, S.; Kang, X.; Zhang, W.; Liu, T. Lingering Response of Radial Growth of Picea crassifolia to Climate at Different Altitudes in the Qilian Mountains, Northwest China. Trees 2017, 31, 455–465. [Google Scholar] [CrossRef]
- Zhang, L.; Li, H.; Ran, Y.; Wang, K.; Zeng, X.; Liu, X. Regional and Local Moisture Gradients Drive the Resistance to and Recovery from Drought of Picea crassifolia Kom. in the Qilian Mountains, Northwest China. Forests 2019, 10, 817. [Google Scholar] [CrossRef] [Green Version]
- Bose, A.K.; Gessler, A.; Bolte, A.; Bottero, A.; Buras, A.; Cailleret, M.; Camarero, J.J.; Haeni, M.; Hereş, A.; Hevia, A.; et al. Growth and Resilience Responses of Scots Pine to Extreme Droughts across Europe Depend on Predrought Growth Conditions. Glob. Chang. Biol. 2020, 26, 4521–4537. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Information | Time Span SSS > 0.85 | Eigenvalue of Residual Chronology | Common Interval Analysis (1949–2017) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | Altitude (m) | Slope | Trees | Cores | MS | SD | MC | AR1 | EPS | SNR | Rbar | Reff | |
| U-site | 2220 | 5° | 27 | 54 | 1923–2017 | 0.188 | 0.158 | 1.003 | −0.111 | 0.972 | 34.169 | 0.564 | 0.564 |
| M-site | 2090 | 15–30° | 25 | 50 | 1906–2017 | 0.176 | 0.159 | 0.984 | −0.077 | 0.967 | 29.156 | 0.440 | 0.543 |
| L-site | 2020 | 15–30° | 27 | 54 | 1917–2017 | 0.173 | 0.150 | 0.996 | −0.127 | 0.969 | 30.899 | 0.426 | 0.549 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, X.; Sun, X.; Jiang, Y.; Xue, F.; Cui, M.; Zhao, S.; Kang, M. The Radial Growth of Picea wilsonii Was More Restricted by Precipitation Due to Climate Warming on Mt. Guandi, China. Forests 2021, 12, 1602. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111602
Huang X, Sun X, Jiang Y, Xue F, Cui M, Zhao S, Kang M. The Radial Growth of Picea wilsonii Was More Restricted by Precipitation Due to Climate Warming on Mt. Guandi, China. Forests. 2021; 12(11):1602. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111602
Chicago/Turabian StyleHuang, Xiaoxia, Xiaoneng Sun, Yuan Jiang, Feng Xue, Minghao Cui, Shoudong Zhao, and Muyi Kang. 2021. "The Radial Growth of Picea wilsonii Was More Restricted by Precipitation Due to Climate Warming on Mt. Guandi, China" Forests 12, no. 11: 1602. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111602